

Abstract
Plasma membrane repair is a conserved cellular response mediating active resealing of membrane disruptions to maintain homeostasis and prevent cell death and progression of multiple diseases. Cell membrane repair repurposes mechanisms from various cellular functions, including vesicle trafficking, exocytosis, and endocytosis, to mend the broken membrane. Recent studies increased our understanding of membrane repair by establishing the molecular machinery contributing to membrane resealing. Here, we review some of the key proteins linked to cell membrane repair.
A eukaryotic cell is separated from the extracellular environment by a plasma membrane composed of a phospholipid bilayer containing proteins that regulate transit of molecules into and out of the cell. Loss of this barrier function can lead to compromised cellular homeostasis and death of the cell. Most cells are subjected to mechanical or chemical stresses that can disrupt the plasma membrane; thus there is strong selective pressure to ensure the integrity of this membrane. The inherent nature of phospholipids and early work with lysosomes indicated that the plasma membrane would thermodynamically reseal after disruption (112). While this is true of simple lipid bilayers or small membrane disruptions (FIGURE 1A), the plasma membrane contains integral proteins that interact with the cytoskeleton and extracellular matrix to support numerous cellular functions. These interactions create mechanical tension on the plasma membrane that holds the membrane open after disruption (25, 114). Such disruptions allow intracellular components to escape the cell and potentially permit toxic levels of Ca2+, oxidants, and other components of the extracellular milieu to enter the cell. Thus, if these disruptions are not closed rapidly, it may lead to the death of the cell. As a result, cells have evolved active methods to reseal plasma membrane disruptions in which normal cellular responses are repurposed to mend the broken membrane (70, 106, 130) through a process called membrane repair.
FIGURE 1.

Models of the plasma membrane repair process
A: thermodynamic resealing occurs spontaneously due to tension produced by the disordered arrangement of the membrane phospholipids at the open edge of the break. This process is the most likely route of resealing for membrane breaks of ≤1 μm in diameter. B: exocytosis can contribute by trafficking intracellular vesicles to the wounded area where they can fuse with each other and the injured membrane to form a repair patch. C: wound constriction is mediated by caveolae. During this process, caveolae cluster and fuse around larger wounds, leading to wound constriction and intracellular fusion of caveolar endosomes. D: budding/blebbing of the membrane portion containing the wound site with release of the newly formed vesicles into the extracellular space also involves exocytosis. E: exocytosis of an intracellular patch and fusion to the wound site could result in the extracellular release or “shedding” of the wound site. F: endocytosis of wounds occurs via invagination of caveolar vesicles and subsequent intracellular fusion of caveolae.
The idea of facilitated membrane repair was supported by earlier work (23, 59) before the concept was formally presented by McNeil and colleagues, who initially showed that plasma membrane disruptions and repair occur in vivo and that damaged cells reseal by recruitment of intracellular vesicles to form a repair patch in an extracellular Ca2+-dependent manner (FIGURE 1B) (12, 96–98, 106, 126). Membrane repair can also involve fusion of vesicles at the injury site or into the proximal plasma membrane. Constriction of the membrane around disruptions can also contribute to membrane repair (FIGURE 1C) (9). Endocytotic mechanisms may be involved in resealing of larger membrane disruptions, whereas smaller disruptions of <100 nm reseal through budding and exocytosis (FIGURE 1D). Repair through budding involves pinching the membrane at the injured site and shedding the injured membrane into the extracellular space (FIGURE 1E) (4, 74, 79). Endocytosis is also thought to contribute to membrane repair by internalization of the injured membrane (FIGURE 1F) (70, 74). How and under what specific conditions these mechanisms contribute to membrane repair is still an area of investigation. These non-exclusive mechanisms could be relevant in a cell-type and context-dependent fashion. It is clear that compromised membrane repair contributes to pathophysiology in a number of different tissues and that it is linked to muscular dystrophy, heart failure, neurodegeneration, and other diseases (5, 7, 24, 27, 50, 72, 133, 140). Despite the importance of membrane repair in cellular function, the field has only recently begun to expand with the discovery of more proteins linked to resealing damaged cell membranes. The focus of this review is to identify several proteins currently linked to membrane repair and to describe some of the key findings on their functions.
Ferlin Family (Dysferlin/Myoferlin/Otoferlin)
Dysferlin is a type II transmembrane protein from the ferlin family that localizes to the plasma membrane and T-tubules of muscle fibers (FIGURE 2) (1, 2). Dysferlin was initially identified as the target gene mutated in Myoshi Myopathy (MM) and limb-girdle muscular dystrophy (LGMD) type 2B (6, 71, 87, 132). Subsequent dysferlin knockout mouse studies showed that muscle fibers (63) and hearts (24, 56) from these animals demonstrated a decreased ability to reseal the plasma membrane (5), leading to muscular dystrophy and late-onset cardiomyopathy (5, 24). These results identified dysferlin as the first protein to contribute directly to membrane repair in striated muscles, resulting in dysferlin's prominent role in the membrane repair literature (26). Subsequent studies linked dysferlin to roles in membrane receptor recycling, endocytosis, vesicle trafficking, membrane turnover, focal adhesion, and ATP-dependent intercellular signaling and modulation of the immune system (31, 34, 36, 47, 58, 109, 139, 141). Recent in vivo studies indicated that dysferlin may function in the maintenance of T-tubule structure (78) and that the principal function of dysferlin may be at the T-tubules rather than in sarcolemmal membrane (77). Additional studies will be necessary to establish the extent that these findings alter the current understanding of dysferlin in membrane repair; however, it is clear that much is still unknown about dysferlin function.
FIGURE 2.

Major membrane repair proteins and their hypothesized roles in the repair process
Dysferlin's interaction with AHNAK is regulated by calpain, and cleavage of dysferlin can result in additional subunits that function in repair. Calpain may also regulate cytoskeletal structure and sarcomere remodeling. AHNAK may aid in cytoskeletal remodeling. MG53/TRIM72 and dysferlin form a vesicle lattice to close the wound. A: affixin, which also binds dysferlin, localizes to focal adhesions and may organize actin. B: ESCRT and acid sphingomyelinase (ASM) facilitate exocytosis and endocytosis. ESCRT has been found to be involved in both endocytosis and budding. ESCRT III can be recruited to the membrane, followed by blebbing of the membrane and shedding of the wound. ASM is secreted and cleaves sphingomyelin to generate ceramide, leading to membrane invagination of the injury site. C: the annexin and S100A10 complex binds dysferlin and may recruit AHNAK to the membrane due to annexin's ability to bind lipid rafts. Annexin/S100A10 may also bridge adjacent phospholipids to form endosomes. Annexin accumulates at the neck of membrane blebs to mediate microvesicle release. Annexin A6 may also “cap” the membrane repair patch. D: synaptotagmin and SNARE proteins interact at the plasma membrane via a conformational change in synaptotagmin present on synaptic vesicles to fuse the vesicles with the membrane.
Other members of the ferlin family also have been linked to membrane repair. For example, myoferlin shares a 56% homology to dysferlin and is expressed during muscle development to facilitate myoblast fusion (41, 139). Wild-type mice show low expression of myoferlin; however, increased expression was observed in mdx and gamma-sarcoglycan-null mouse models of muscular dystrophy (37, 141). Transgenic mouse experiments showed that myoferlin overexpression could compensate for dysferlin in membrane repair; however, myoferlin does not prevent all dystrophy symptoms (90). Another dysferlin family member, otoferlin, regulates synaptic vesicle exocytosis in cochlear hair cells and may be involved in membrane repair specifically within these cells (75, 121).
Calpain
After membrane disruption, rapid reorganization of cytoskeletal elements at the site of injury is necessary. Treatment with the actin-depolymerizing enzyme DNase I significantly enhanced resealing, demonstrating that disassembly of the actin skeleton is important in membrane resealing (107). Calpain 3 is a cysteine protease that cleaves cytoskeletal proteins, such as talin and vimentin, and may aid in early remodeling of various proteins during membrane repair as cleaved fragments of these cytoskeletal proteins could not be recovered in calpain-null cells (101). Calpain 3 cleaves AHNAK, inhibits AHNAK's interactions with dysferlin and myoferlin, and regulates AHNAK protein turnover (FIGURE 2). Through these actions, calpain regulates cytoskeletal structure and interaction of the cytoskeleton with the cell membrane (34, 67). In fact, loss of calpain 3 in particular has been shown to cause limb girdle muscular dystrophy (119). Calpain has been shown to be required for calcium-dependent membrane repair, since knockout of the small subunit of the calpain enzyme results in failure to reseal after laser-induced membrane disruption (101). Calpain could contribute to membrane repair through other mechanisms as well. Studies suggest that calpain can remodel sarcomeres since calpain 3 binds titin and is present in other regions of the sarcomere, such as the Z disk, costameres, and myotendinous junctions (45). Thus the role of calpain may be to increase local loosening of the sarcomere via proteolysis and facilitate removal of damaged and cleaved proteins by the proteosome (8). Other studies suggest a role for calpain 3 in post-membrane repair sarcolemmal remodeling because loss of calpain in limb girdle muscular dystrophy leads to disorganization of myofibers and a lack of organized sarcomeres and sarcomere proteins in myotubes (45, 67, 100). Finally, calpain activity leads to dysferlin cleavage to produce subunits that function in membrane repair (116).
MG53/TRIM72
Mitsugumin 53 (MG53/TRIM72) has been shown to be a vital component of the membrane repair machinery in several cell types (18, 21, 44, 80). It is a member of the tripartite motif family of E3 ubiquitin ligases (TRIM72) that was originally identified in an immunoproteomic library screen for muscle enriched proteins (136). Although native MG53/TRIM72 protein was initially thought to be found only in skeletal and cardiac muscle (18), recent studies have shown expression and membrane repair function in other tissues (44, 73, 80). Along with native protein repair capacity, overexpression of MG53/TRIM72 in cells that do not express MG53/TRIM72 also shows protective effects against membrane injury (137). MG53/TRIM72-null mice display defective membrane repair (18), progressive myopathy (18), and increased susceptibility to injury in the heart (21, 135), lungs (73, 80), and kidneys (44).
MG53/TRIM72 interacts with phosphatidylserine to associate with intracellular vesicles and the inner leaflet of the plasma membrane (18) (FIGURE 3). Once a cell is injured, MG53/TRIM72 is thought to react to the oxidized extracellular environment entering the cell by forming higher molecular weight units (69). MG53/TRIM72-tethered vesicles traffic to the membrane disruption, allowing vesicles to fuse and patch the injured membrane (18). This process appears to involve a dysferlin and Caveolin-3 (Cav-3) containing complex that regulates repair in the sarcolemma (20, 134). A study using ballistic injury and super-resolution microscopy of human myotubes showed that MG53/TRIM72 is recruited quickly to the membrane (2 s), followed by dysferlin ∼10 s post-injury (84). MG53/TRIM72 and dysferlin form a lattice that fills the wound area with vesicles and closes the wound (84).
FIGURE 3.
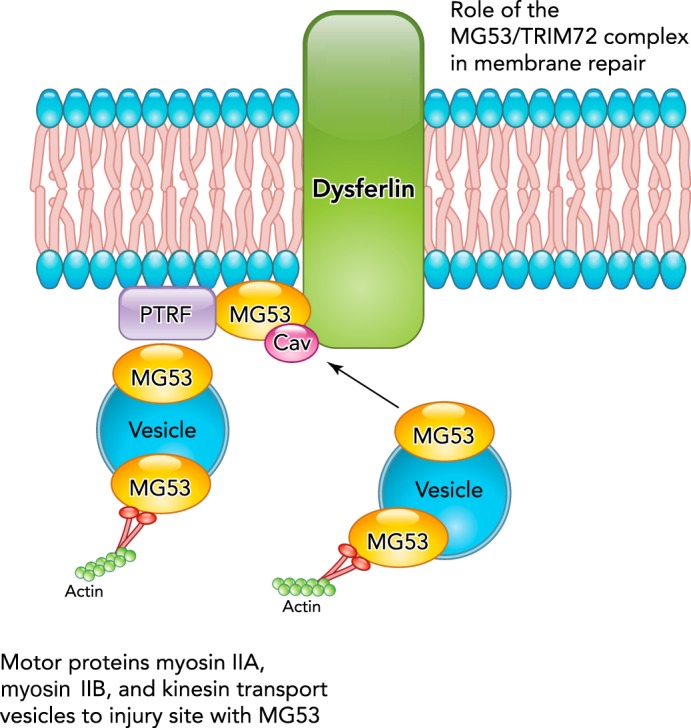
Proposed roles of MG53/TRIM72 in mediating membrane repair
MG53/TRIM72 interacts with phosphatidylserine in the plasma membrane in a complex containing dysferlin and Cav-3. MG53/TRIM72 and dysferlin close the membrane wound with vesicles. Vesicle transport is facilitated by myosin motor proteins. Cav-3 may regulate MG53/TRIM72-mediated membrane fusion and is enriched in caveolae or plasma membrane invaginations. PTRF may aid in the formation and stabilization of these caveolae by interaction with Cav-3 and MG53/TRIM72 through cholesterol. PTRF also binds and may help localize dysferlin.
Interestingly, when myotubes are injured in media containing exogenous recombinant human MG53/TRIM72 protein (rhMG53), the protein can be seen to localize to the injury site (137). This mechanism is similar to intracellular MG53/TRIM72 binding to exposed phosphatidylserine (18) and does not require endogenous MG53/TRIM72 or dysferlin (137). This association with injury sites can increase membrane resealing (137). Application of rhMG53 to animal models of various diseases, such as muscular dystrophy (137), myocardial infarct (88), lung injury (73, 80), compartment syndrome (28), and acute kidney injury (44) can reduce the pathology in these models. While this property of the protein is dependent on the phosphatidylserine binding capacity of rhMG53, it is possible that other aspects of the protein function, such as regulation of intracellular signaling, could also contribute to this ability to increase membrane repair capacity (55, 76, 83, 88). Further studies should determine the rhMG53 extracellular mechanism of action and assess its potential as a therapeutic agent.
Caveolin
Caveolin family (Cav-1, -2, and -3) proteins are 21- to 24-kDa integral membrane proteins enriched in invaginations of the plasma membrane (caveolae) and are involved in membrane transport (82) (FIGURES 1C AND 3). Cav-3 is muscle specific and has been most closely linked with membrane repair. Mutations in Cav-3 cause autosomal-dominant LGMD1C and autosomal-dominant rippling muscle disease (AD-RMD) (110). Cav-3 has been shown by co-immunoprecipitation to interact with MG53/TRIM72 and dysferlin (20), and Cav-3 overexpression regulates membrane fusion events by downregulating MG53/TRIM72-induced membrane budding and preventing development of filopodia-like structures (19). Disruption of the Cav-3/dysferlin/MG53 complex can affect the localization and membrane repair function of the other components. For example, dominant negative Cav-3 mutations associated with the development of muscular dystrophy have been shown to cause retention of MG53/TRIM72 (20) or dysferlin (61) in the Golgi apparatus and loss of membrane repair capacity. However, other studies using ultrastructural analysis of dysferlin trafficking showed that dysferlin can still reach the plasma membrane in the absence of Cav-3 but that it is rapidly endocytosed (60).
Polymerase-1 and Transcriptase Release Factor (PTRF/Cavin-1/Cav-p60)
PTRF may aid in the formation of caveolae at the plasma membrane since PTRF localizes to caveolae, expression of PTRF is sufficient for caveolae formation, and loss of PTRF results in a reduced number of caveolae and a dystrophic phenotype (17). Experiments using fluorescence lifetime imaging showed that cholesterol is required for the interaction between PTRF and caveolin. Since cholesterol depletion decreases the PTRF-caveolin interaction, this suggests that PTRF may be involved in stabilizing the membrane curvature of caveolae (62). Additionally, it was shown by immunoprecipitation that PTRF binds dysferlin and, in fact, may be required for the correct localization of dysferlin, since PTRF mutation results in decreased dysferlin at the cell membrane (17). Knockdown of PTRF by shRNA results in decreased membrane repair capacity, whereas overexpression of PTRF can rescue dystrophic muscle membrane repair (143). During the membrane repair process, PTRF binds to dysferlin and may anchor MG53/TRIM72 to cholesterol, since MG53/TRIM72 cannot bind cholesterol unless PTRF is present (92, 143). In this model, PTRF anchors MG53/TRIM72 by binding to exposed cholesterol at the membrane injury site (FIGURE 3).
Motor Proteins (Nonmuscle Myosin IIA and IIB/Kinesin)
Since vesicle trafficking is an important aspect of membrane repair, motor proteins must be involved in this process to allow trafficking to occur. Membrane repair is sensitive to inhibitors of both myosin and kinesin motor proteins (126, 131). Studies have specifically shown that non-muscle myosin IIA and B are important motor proteins that mediate vesicle trafficking during membrane repair (131). Antisense knockdown of myosin IIB suppressed exocytosis and membrane resealing, and knockdown of myosin IIA inhibited the rate of resealing at repeated wound sites. These studies are supported by the observation that non-muscle myosin IIA facilitates the transport of vesicles containing MG53/TRIM72 to the site of membrane injury (86) (FIGURE 3).
Affixin [β-Parvin/Integrin-Linked Kinase (ILK)-Binding Protein]
Affixin was first discovered to be an integrin-linked kinase binding protein that localizes to focal adhesions and likely contributes to their maturation. In muscle cells, affixin and ILK colocalize at sites where the Z band attaches to the sarcolemma due to an interaction with dysferlin (94). Affixin's interaction with dysferlin and altered immunoreactivity in MM, LGMD2B, and LGMD1C suggest a role for this protein in muscle membrane repair, possibly through organization of F-actin (FIGURE 2A) (94). For example, affixin has been shown by immunoprecipitation and pull-down assay to interact with guanine nucleotide exchange factor αPIX (ARHGEF6 or Cool-2), which functions to regulate actin skeleton adhesion (93, 94, 104) as well as α-actinin, which plays a role in organization of the cytoskeleton (142). Although it appears that affixin may participate in cytoskeletal remodeling during membrane repair, the precise role of affixin in this process remains speculative.
Acid Sphingomyelinase
Acid sphingomyelinase (ASM) is an enzyme that cleaves the phosphorylcholine head of sphingomyelin to generate ceramide, a molecule that leads to membrane invagination, and that contributes to the process of endocytosis during membrane repair (64, 124). In response to injury, lysosomes fuse with the injured cell membrane and release ASM from the cell. The action of ASM causes invagination of the membrane and endocytosis of the injury site (FIGURE 2B) (29, 35, 42, 129). Additional evidence of ASM contribution to membrane repair is provided by the finding that dysferlin-deficient C2C12 cells showed less secretion of ASM than control cells (35). When ASM was inhibited and cells were permeabilized using a pore-forming toxin, the cells could not sufficiently repair (35). Treatment with exogenous ASM was sufficient to restore the membrane integrity of ASM-depleted cells (35) and of a dystrophic patient-derived myoblast cell line (35).
ESCRT
Endosomal sorting complex required for transport (ESCRT) is involved in viral budding, cytokinesis, and spontaneous budding of the plasma membrane. ESCRT subunits are classified into five complexes (108). The ESCRT III complex recently has been shown to be involved in endocytosis and budding in response to membrane damage (30, 74, 122). After injury, apoptosis-linked gene (ALG)-2 binds Ca2+ and leads to the accumulation of ESCRT III and accessory proteins ALG-2-interacting protein X (ALIX), and vacuolar protein sorting-associated protein 4 (Vps4) (122). Assembly of this complex results in cleavage of the wounded membrane and shedding of the wound site into extracellular vesicles. In response to UV laser injury, ESCRT III is recruited to injury sites exercising calcium-dependent wound repair. The observed recruitment of ESCRT is followed by blebbing of the membrane and shedding of the wound (FIGURE 2B). ESCRT III is involved in endocytosis of the membrane after the insertion of the bacterial pore-forming toxin SLO (74). The complex appears to be critical for repair of injuries of <100 nm (74).
Annexin
Annexins are a protein family consisting of a COOH terminus containing phospholipid and Ca2+ binding sites, and a variable NH2 terminus (111). The annexins are unique in that they can bind lipids while in their Ca2+-bound conformation, and the concentration of the Ca2+ signal determines which particular annexin family member will bind to the phospholipid (53). Although the exact mechanism remains unclear, numerous studies suggest roles for annexins in vesicle movement, fusion, and patch formation during membrane repair (54, 57, 95). Annexins A1 and A2 bind dysferlin in a Ca2+-dependent manner and may contribute to membrane repair by their ability to assist in the aggregation and fusion of intracellular vesicles by their association with lipid rafts of the plasma membrane (3, 85). Density gradient centrifugation experiments confirmed the Ca2+- and annexin-dependent association of these rafts. Electron microscopic evidence shows that annexin may be involved in membrane fusion during exocytosis as well as serving as a scaffold for endosomes (FIGURE 2C) (11, 53). Annexins also have been shown to mediate microvesicle release and blebbing, an indirect repair mechanism that involves sealing off a damaged membrane segment by accumulation of annexin at the neck of the membrane bleb (43). Other studies indicate that annexin A5 (AnxA5) can bind to injured membranes and form a two-dimensional array that is thought to contribute to membrane repair, since AnxA5-null cells show defective repair capacity (15). Finally, annexin A6 has been shown by live imaging to locate to membrane disruptions and to assemble into a “cap” on the membrane repair patch to assist with membrane resealing (128).
AHNAK (Desmoyokin)
AHNAK, or desmoyokin, is a 629.1-kDa tripartite nucleoprotein with potential functions as diverse as fat metabolism, DNA repair, autoantigenicity, cell signaling, calcium channel regulation, and tumor metastasis (33). It has been proposed that interaction of AHNAK with annexin 2 and S100A10 regulates organization of the actin cytoskeleton and architecture of the cell membrane, since AHNAK-specific siRNA prevents actin cytoskeleton reorganization (10). In membrane repair, AHNAK has been associated with enlargeosomes, vesicles that rapidly exocytose in response to calcium influx; however, its exact role within these vesicles is unknown (14). AHNAK's ability to bind actin may signify a role for the protein in membrane resealing, although its presence within enlargeosomes during recruitment to the membrane injury suggests earlier involvement in the repair process (33). The co-localization of AHNAK with annexin 2 within vesicles is controversial, as studies also have reported that annexin is on the cytosolic face of the vesicle (89). In fact, calcium-sensitive annexin 2 may be involved in recruitment of AHNAK and S100A10 to the plasma membrane in response to calcium-annexin binding (33). Furthermore, co-immunoprecipitation and mass spectrometry studies confirm that AHNAK interacts with dysferlin, which may localize and stabilize AHNAK at the sarcolemma through its transmembrane domain, an interaction possibly regulated by calpain 3 (67, 68) (FIGURE 2).
S100A10
S100A10 is a small, ∼10-kDa EF-hand Ca2+ binding protein also known as annexin 2 light chain or p11. S100A10 forms a heterotetrameric complex with annexin A2 by forming an S100A10 dimer in the middle of two annexin A2 chains. This complex targets to the plasma membrane in a calcium-regulated manner that is dependent on its interaction with annexin 2 (FIGURE 2C). At the plasma membrane, S100A10 interacts with a number of proteins, where it may be essential for surface presentation of proteins such as ion channels (117). Furthermore, proteomic and structural analyses have identified that the heterotetrameric complex binds cytosolic proteins AHNAK and dysferlin, and that it is responsible for the recruitment of AHNAK to the cell membrane, where the entire complex acts as a scaffold for membrane repair (38, 81, 111, 118). Although S100A10 is known to be a central player in the membrane repair complex, its exact function in membrane repair has not been identified. One model suggests that membrane fusion may occur due to the ability of S100A10-annexin A2 to bridge adjacent phospholipid membranes (54).
Synaptotagmin/SNAREs
Synaptotagmins (Syt) are a family of proteins that contain two C2 domains (C2A and C2B), which some members use to bind Ca2+ or phospholipids (16, 65, 102, 123, 127). Much of the focus on synaptotagmins has been in synaptic neurotransmitter release in neurons through their interactions with soluble N-ethylmaleimide-sensitive factor attachment protein receptors (SNARE) (13, 32, 46, 49, 51, 52, 103). The general mechanism of this interaction begins with vesicular SNAREs interacting with membrane SNAREs to dock the vesicle to the membrane. Synaptotagmins bound to the vesicle bind Ca2+ and are then able to interact with the SNAREs to fuse the vesicle to the membrane and expel the contents of the vesicle (FIGURE 2D). Membrane repair is not solely dependent on synaptotagmins because studies using botulinum toxin A, which cleaves SNARE proteins, showed inhibition of resealing of sea urchin eggs (12). Disruption of the formation of the SNARE complex using the cytoplasmic domain of synaptobrevin 2, a SNARE protein, was also able to block membrane resealing (125).
The relationship between synaptotagmins and SNAREs is important because of their roles in Ca2+ sensing and vesicle membrane fusion. Syt I is found exclusively in the nervous system (48, 138), and antibodies against the C2A domain of Syt I caused inhibition of membrane repair in severed axons of squid and crayfish giant axons (40). This same study used antibodies against a domain of a SNARE protein, syntaxin, to inhibit interaction with synaptotagmin's C2A domain (40). Similar to inhibition of Syt I, blocking syntaxin inhibited resealing of the severed axon (40). Syt VII is ubiquitously expressed and found proximal to the membrane on lysosomal-associated membrane protein 1 (LAMP-1)-positive lysosomes. Cells injured in the presence of Syt VII blocking antibodies showed a decreased capacity to reseal membrane disruptions (115), and fibroblasts taken from Syt VII-deficient mice showed defective lysosomal exocytosis and decreased capacity to reseal their plasma membrane (22).
Conclusions
We have attempted to summarize the available literature concerning the roles of many of the proteins known to contribute to the membrane repair process. While clear evidence for the involvement of the discussed proteins in membrane repair exists, it should be noted that other molecules, such as Amphiphysin 2 (BIN1) (113), Lamp-1 (115), and others, have been speculated to play a role in membrane resealing due to the nature of their actions in the cell or due to their interactions with known members of the membrane repair machinery. For example, a recent study identified ATPase EH-domain containing 2 as a novel membrane repair protein that co-localizes at the site of membrane injury with F-actin and annexin A1 (91). Moesin (membrane-organizing extension spike protein) has been shown to interact with dysferlin and appears to cross-link plasma membranes and actin cytoskeletons (17). As more information about specific proteins and membrane repair in general is obtained, we expect that additional candidates will be discovered.
In considering the molecules known to be important in the membrane repair process, it becomes clear that many of the steps in membrane repair are processes necessary for normal cellular functions. For example, myosin, kinesin, annexin, and SNAREs are involved in vesicle trafficking and membrane fusion, integral events for numerous cell activities (39). Calpain and affixin are important for cytoskeletal remodeling during cell motility, responses to the cell environment and mitosis (66, 120). PTRF, ESCRT, ASM, and S100A10 are active at the plasma membrane to form caveolae, membrane buds, and other membrane invaginations as well as to mediate membrane fusion (17, 54, 74, 129). Dysferlin is integral to multiple activities, such as endocytosis, vesicle trafficking, membrane turnover, and others (31, 47, 58). Membrane repair can be considered an emergency response in which these cellular processes are used to reseal the membrane and allow cell survival (57, 99, 105). However, it is necessary to move beyond dissection of this supportive cellular machinery to gain a full understanding of membrane repair. Given the current understanding in the field, it is difficult to define the prevailing membrane repair hypothesis, and it is likely that there is no single mechanism that is at work under all injury scenarios. Our current understanding of membrane repair is limited to a subset of cellular functions and protein interactions, leaving compelling questions unanswered. For example, how does the cell differentiate the membrane repair process from normal cell functions and what are the specific signaling pathways that allow this specific response to occur? Furthermore, what are the signals that direct the repair machinery to the site of injury? While calcium-dependent mechanisms are known signals for the assembly of membrane repair proteins at the damage site and for fusion of membrane surfaces, additional mediators may exist. Discovery and characterization of these mediators and pathways are important next steps in understanding the membrane repair process.
Acknowledgments
Eric X Beck provided helpful comments in the preparation of this manuscript.
Footnotes
This work was supported by the Muscular Dystrophy Association, the OSU Physiology and Cell Biology Margaret T. Nishikawara Merit Scholarship Fund, and the National Institute of Arthritis and Musculoskeletal and Skin Diseases of the National Institutes of Health under Award No. AR-063084. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Noah Weisleder is Founder and Chief Scientific Officer of TRIM-edicine, a biotechnology company developing products targeting membrane repair, including rhMG53.
Author contributions: A.D.B., B.J.P., and N.W. prepared figures; A.D.B., B.J.P., and N.W. drafted manuscript; A.D.B., B.J.P., and N.W. edited and revised manuscript; N.W. approved final version of manuscript.
References
- 1.Ampong BN, Imamura M, Matsumiya T, Yoshida M, Takeda S. Intracellular localization of dysferlin and its association with the dihydropyridine receptor. Acta Myol 24: 134–144, 2005. [PubMed] [Google Scholar]
- 2.Anderson LV, Davison K, Moss JA, Young C, Cullen MJ, Walsh J, Johnson MA, Bashir R, Britton S, Keers S, Argov Z, Mahjneh I, Fougerousse F, Beckmann JS, Bushby KM. Dysferlin is a plasma membrane protein and is expressed early in human development. Hum Mol Genet 8: 855–861, 1999. [DOI] [PubMed] [Google Scholar]
- 3.Babiychuk EB, Draeger A. Annexins in cell membrane dynamics. Ca2+-regulated association of lipid microdomains. J Cell Biol 150: 1113–1124, 2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Babiychuk EB, Monastyrskaya K, Potez S, Draeger A. Intracellular Ca2+ operates a switch between repair and lysis of streptolysin O-perforated cells. Cell Death Differ 16: 1126–1134, 2009. [DOI] [PubMed] [Google Scholar]
- 5.Bansal D, Miyake K, Vogel SS, Groh S, Chen CC, Williamson R, McNeil PL, Campbell KP. Defective membrane repair in dysferlin-deficient muscular dystrophy. Nature 423: 168–172, 2003. [DOI] [PubMed] [Google Scholar]
- 6.Bashir R, Britton S, Strachan T, Keers S, Vafiadaki E, Lako M, Richard I, Marchand S, Bourg N, Argov Z, Sadeh M, Mahjneh I, Marconi G, Passos-Bueno MR, Moreira EeS, Zatz M, Beckmann JS, Bushby K. A gene related to Caenorhabditis elegans spermatogenesis factor fer-1 is mutated in limb-girdle muscular dystrophy type 2B. Nat Genet 20: 37–42, 1998. [DOI] [PubMed] [Google Scholar]
- 7.Bazan NG, Marcheselli VL, Cole-Edwards K. Brain response to injury and neurodegeneration: endogenous neuroprotective signaling. Ann NY Acad Sci 1053: 137–147, 2005. [DOI] [PubMed] [Google Scholar]
- 8.Beckmann JS, Spencer M. Calpain 3, the “gatekeeper” of proper sarcomere assembly, turnover and maintenance. Neuromuscul Disord 18: 913–921, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bement WM, Forscher P, Mooseker MS. A novel cytoskeletal structure involved in purse string wound closure and cell polarity maintenance. J Cell Biol 121: 565–578, 1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Benaud C, Gentil BJ, Assard N, Court M, Garin J, Delphin C, Baudier J. AHNAK interaction with the annexin 2/S100A10 complex regulates cell membrane cytoarchitecture. J Cell Biol 164: 133–144, 2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Bharadwaj A, Bydoun M, Holloway R, Waisman D. Annexin A2 heterotetramer: structure and function. Int J Mol Sci 14: 6259–6305, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bi GQ, Alderton JM, Steinhardt RA. Calcium-regulated exocytosis is required for cell membrane resealing. J Cell Biol 131: 1747–1758, 1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bommert K, Charlton MP, DeBello WM, Chin GJ, Betz H, Augustine GJ. Inhibition of neurotransmitter release by C2-domain peptides implicates synaptotagmin in exocytosis. Nature 363: 163–165, 1993. [DOI] [PubMed] [Google Scholar]
- 14.Borgonovo B, Cocucci E, Racchetti G, Podini P, Bachi A, Meldolesi J. Regulated exocytosis: a novel, widely expressed system. Nat Cell Biol 4: 955–962, 2002. [DOI] [PubMed] [Google Scholar]
- 15.Bouter A, Gounou C, Bérat R, Tan S, Gallois B, Granier T, d'Estaintot BL, Pöschl E, Brachvogel B, Brisson AR. Annexin-A5 assembled into two-dimensional arrays promotes cell membrane repair. Nat Commun 2: 270, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Brose N, Petrenko AG, Südhof TC, Jahn R. Synaptotagmin: a calcium sensor on the synaptic vesicle surface. Science 256: 1021–1025, 1992. [DOI] [PubMed] [Google Scholar]
- 17.Cacciottolo M, Belcastro V, Laval S, Bushby K, di Bernardo D, Nigro V. Reverse engineering gene network identifies new dysferlin-interacting proteins. J Biol Chem 286: 5404–5413, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Cai C, Masumiya H, Weisleder N, Matsuda N, Nishi M, Hwang M, Ko JK, Lin P, Thornton A, Zhao X, Pan Z, Komazaki S, Brotto M, Takeshima H, Ma J. MG53 nucleates assembly of cell membrane repair machinery. Nat Cell Biol 11: 56–64, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Cai C, Masumiya H, Weisleder N, Pan Z, Nishi M, Komazaki S, Takeshima H, Ma J. MG53 regulates membrane budding and exocytosis in muscle cells. J Biol Chem 284: 3314–3322, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Cai C, Weisleder N, Ko JK, Komazaki S, Sunada Y, Nishi M, Takeshima H, Ma J. Membrane repair defects in muscular dystrophy are linked to altered interaction between MG53, caveolin-3, and dysferlin. J Biol Chem 284: 15894–15902, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Cao CM, Zhang Y, Weisleder N, Ferrante C, Wang X, Lv F, Song R, Hwang M, Jin L, Guo J, Peng W, Li G, Nishi M, Takeshima H, Ma J, Xiao RP. MG53 constitutes a primary determinant of cardiac ischemic preconditioning. Circulation 121: 2565–2574, 2010. [DOI] [PubMed] [Google Scholar]
- 22.Chakrabarti S, Kobayashi KS, Flavell RA, Marks CB, Miyake K, Liston DR, Fowler KT, Gorelick FS, Andrews NW. Impaired membrane resealing and autoimmune myositis in synaptotagmin VII-deficient mice. J Cell Biol 162: 543–549, 2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Chambers R, Chambers EL. Explorations into the nature of the living cell. Acad Med 36: 966, 1961. [Google Scholar]
- 24.Chase TH, Cox GA, Burzenski L, Foreman O, Shultz LD. Dysferlin deficiency and the development of cardiomyopathy in a mouse model of limb-girdle muscular dystrophy 2B. Am J Pathol 175: 2299–2308, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Chernomordik LV, Melikyan GB, Chizmadzhev YA. Biomembrane fusion: a new concept derived from model studies using two interacting planar lipid bilayers. Biochim Biophys Acta 906: 309–352, 1987. [DOI] [PubMed] [Google Scholar]
- 26.Cooper ST, Head SI. Membrane injury and repair in the muscular dystrophies. Neuroscientist. In press. [DOI] [PubMed]
- 27.Cooper ST, Kizana E, Yates JD, Lo HP, Yang N, Wu ZH, Alexander IE, North KN. Dystrophinopathy carrier determination and detection of protein deficiencies in muscular dystrophy using lentiviral MyoD-forced myogenesis. Neuromuscul Disord 17: 276–284, 2007. [DOI] [PubMed] [Google Scholar]
- 28.Corona BT, Garg K, Roe JL, Zhu H, Park KH, Ma J, Walters TJ. Effect of recombinant human MG53 protein on tourniquet-induced ischemia-reperfusion injury in rat muscle. Muscle Nerve 49: 919–921, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Corrotte M, Almeida PE, Tam C, Castro-Gomes T, Fernandes MC, Millis BA, Cortez M, Miller H, Song W, Maugel TK, Andrews NW. Caveolae internalization repairs wounded cells and muscle fibers. Elife 2: e00926, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Corrotte M, Fernandes MC, Tam C, Andrews NW. Toxin pores endocytosed during plasma membrane repair traffic into the lumen of MVBs for degradation. Traffic 13: 483–494, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Covian-Nares JF, Koushik SV, Puhl HL, Vogel SS. Membrane wounding triggers ATP release and dysferlin-mediated intercellular calcium signaling. J Cell Sci 123: 1884–1893, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Davis AF, Bai J, Fasshauer D, Wolowick MJ, Lewis JL, Chapman ER. Kinetics of synaptotagmin responses to Ca2+ and assembly with the core SNARE complex onto membranes. Neuron 24: 363–376, 1999. [DOI] [PubMed] [Google Scholar]
- 33.Davis TA, Loos B, Engelbrecht AM. AHNAK: the giant jack of all trades. Cell Signal 26: 2683–2693, 2014. [DOI] [PubMed] [Google Scholar]
- 34.de Morrée A, Hensbergen PJ, van Haagen HH, Dragan I, Deelder AM, 't Hoen PA, Frants RR, van der Maarel SM. Proteomic analysis of the dysferlin protein complex unveils its importance for sarcolemmal maintenance and integrity. PLos One 5: e13854, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Defour A, Van der Meulen JH, Bhat R, Bigot A, Bashir R, Nagaraju K, Jaiswal JK. Dysferlin regulates cell membrane repair by facilitating injury-triggered acid sphingomyelinase secretion. Cell Death Dis 5: e1306, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Demonbreun AR, Fahrenbach JP, Deveaux K, Earley JU, Pytel P, McNally EM. Impaired muscle growth and response to insulin-like growth factor 1 in dysferlin-mediated muscular dystrophy. Hum Mol Genet 20: 779–789, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Demonbreun AR, Lapidos KA, Heretis K, Levin S, Dale R, Pytel P, Svensson EC, McNally EM. Myoferlin regulation by NFAT in muscle injury, regeneration and repair. J Cell Sci 123: 2413–2422, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Dempsey BR, Rezvanpour A, Lee TW, Barber KR, Junop MS, Shaw GS. Structure of an asymmetric ternary protein complex provides insight for membrane interaction. Structure 20: 1737–1745, 2012. [DOI] [PubMed] [Google Scholar]
- 39.Derby MC, Gleeson PA. New insights into membrane trafficking and protein sorting. Intl Rev Cytol 261: 47–116, 2007. [DOI] [PubMed] [Google Scholar]
- 40.Detrait E, Eddleman CS, Yoo S, Fukuda M, Nguyen MP, Bittner GD, Fishman HM. Axolemmal repair requires proteins that mediate synaptic vesicle fusion. J Neurobiol 44: 382–391, 2000. [DOI] [PubMed] [Google Scholar]
- 41.Doherty KR, Cave A, Davis DB, Delmonte AJ, Posey A, Earley JU, Hadhazy M, McNally EM. Normal myoblast fusion requires myoferlin. Development 132: 5565–5575, 2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Draeger A, Babiychuk EB. Ceramide in plasma membrane repair. Handb Exp Pharmacol 341–353, 2013. [DOI] [PubMed] [Google Scholar]
- 43.Draeger A, Monastyrskaya K, Babiychuk EB. Plasma membrane repair and cellular damage control: the annexin survival kit. Biochem Pharmacol 81: 703–712, 2011. [DOI] [PubMed] [Google Scholar]
- 44.Duann P, Li H, Lin P, Tan T, Wang Z, Chen K, Zhou X, Gumpper K, Zhu H, Ludwig T, Mohler PJ, Rovin B, Abraham WT, Zeng C, Ma J. MG53-mediated cell membrane repair protects against acute kidney injury. Sci Transl Med 7: 279ra236, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Duguez S, Bartoli M, Richard I. Calpain 3: a key regulator of the sarcomere? FEBS J 273: 3427–3436, 2006. [DOI] [PubMed] [Google Scholar]
- 46.Earles CA, Bai J, Wang P, Chapman ER. The tandem C2 domains of synaptotagmin contain redundant Ca2+ binding sites that cooperate to engage t-SNAREs and trigger exocytosis. J Cell Biol 154: 1117–1123, 2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Evesson FJ, Peat RA, Lek A, Brilot F, Lo HP, Dale RC, Parton RG, North KN, Cooper ST. Reduced plasma membrane expression of dysferlin mutants is attributed to accelerated endocytosis via a syntaxin-4-associated pathway. J Biol Chem 285: 28529–28539, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Fox MA, Sanes JR. Synaptotagmin I and II are present in distinct subsets of central synapses. J Comp Neurol 503: 280–296, 2007. [DOI] [PubMed] [Google Scholar]
- 49.Fukuda M, Moreira JE, Lewis FM, Sugimori M, Niinobe M, Mikoshiba K, Llinás R. Role of the C2B domain of synaptotagmin in vesicular release and recycling as determined by specific antibody injection into the squid giant synapse preterminal. Proc Natl Acad Sci USA 92: 10708–10712, 1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Gajic O, Lee J, Doerr CH, Berrios JC, Myers JL, Hubmayr RD. Ventilator-induced cell wounding and repair in the intact lung. Am J Respir Crit Care Med 167: 1057–1063, 2003. [DOI] [PubMed] [Google Scholar]
- 51.Geppert M, Goda Y, Hammer RE, Li C, Rosahl TW, Stevens CF, Südhof TC. Synaptotagmin I: a major Ca2+ sensor for transmitter release at a central synapse. Cell 79: 717–727, 1994. [DOI] [PubMed] [Google Scholar]
- 52.Geppert M, Goda Y, Stevens CF, Südhof TC. The small GTP-binding protein Rab3A regulates a late step in synaptic vesicle fusion. Nature 387: 810–814, 1997. [DOI] [PubMed] [Google Scholar]
- 53.Gerke V, Creutz CE, Moss SE. Annexins: linking Ca2+ signalling to membrane dynamics. Nat Rev Mol Cell Biol 6: 449–461, 2005. [DOI] [PubMed] [Google Scholar]
- 54.Gerke V, Moss SE. Annexins: from structure to function. Physiol Rev 82: 331–371, 2002. [DOI] [PubMed] [Google Scholar]
- 55.Ham YM, Mahoney SJ. Compensation of the AKT signaling by ERK signaling in transgenic mice hearts overexpressing TRIM72. Exp Cell Res 319: 1451–1462, 2013. [DOI] [PubMed] [Google Scholar]
- 56.Han R, Bansal D, Miyake K, Muniz VP, Weiss RM, McNeil PL, Campbell KP. Dysferlin-mediated membrane repair protects the heart from stress-induced left ventricular injury. J Clin Invest 117: 1805–1813, 2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Han R, Campbell KP. Dysferlin and muscle membrane repair. Curr Opin Cell Biol 19: 409–416, 2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Han R, Frett EM, Levy JR, Rader EP, Lueck JD, Bansal D, Moore SA, Ng R, Beltrán-Valero de Bernabé D, Faulkner JA, Campbell KP. Genetic ablation of complement C3 attenuates muscle pathology in dysferlin-deficient mice. J Clin Invest 120: 4366–4374, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Heilbrunn LV. The Dynamics of Living Protoplasm. New York: Academic, 1956, p. 480–481. [Google Scholar]
- 60.Hernandez-Deviez DJ, Howes MT, Laval SH, Bushby K, Hancock JF, Parton RG. Caveolin regulates endocytosis of the muscle repair protein, dysferlin. J Biol Chem 283: 6476–6488, 2008. [DOI] [PubMed] [Google Scholar]
- 61.Hernández-Deviez DJ, Martin S, Laval SH, Lo HP, Cooper ST, North KN, Bushby K, Parton RG. Aberrant dysferlin trafficking in cells lacking caveolin or expressing dystrophy mutants of caveolin-3. Hum Mol Genet 15: 129–142, 2006. [DOI] [PubMed] [Google Scholar]
- 62.Hill MM, Bastiani M, Luetterforst R, Kirkham M, Kirkham A, Nixon SJ, Walser P, Abankwa D, Oorschot VM, Martin S, Hancock JF, Parton RG. PTRF-Cavin, a conserved cytoplasmic protein required for caveola formation and function. Cell 132: 113–124, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ho M, Post CM, Donahue LR, Lidov HG, Bronson RT, Goolsby H, Watkins SC, Cox GA, Brown RH Jr. Disruption of muscle membrane and phenotype divergence in two novel mouse models of dysferlin deficiency. Hum Mol Genet 13: 1999–2010, 2004. [DOI] [PubMed] [Google Scholar]
- 64.Holopainen JM, Angelova MI, Kinnunen PK. Vectorial budding of vesicles by asymmetrical enzymatic formation of ceramide in giant liposomes. Biophys J 78: 830–838, 2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Holz RW, Hlubek MD, Sorensen SD, Fisher SK, Balla T, Ozaki S, Prestwich GD, Stuenkel EL, Bittner MA. A pleckstrin homology domain specific for phosphatidylinositol 4,5-bisphosphate (PtdIns-4,5-P2) and fused to green fluorescent protein identifies plasma membrane PtdIns-4,5-P2 as being important in exocytosis. J Biol Chem 275: 17878–17885, 2000. [DOI] [PubMed] [Google Scholar]
- 66.Honda S, Marumoto T, Hirota T, Nitta M, Arima Y, Ogawa M, Saya H. Activation of m-calpain is required for chromosome alignment on the metaphase plate during mitosis. J Biol Chem 279: 10615–10623, 2004. [DOI] [PubMed] [Google Scholar]
- 67.Huang Y, de Morrée A, van Remoortere A, Bushby K, Frants RR, den Dunnen JT, van der Maarel SM. Calpain 3 is a modulator of the dysferlin protein complex in skeletal muscle. Hum Mol Genet 17: 1855–1866, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Huang Y, Laval SH, van Remoortere A, Baudier J, Benaud C, Anderson LV, Straub V, Deelder A, Frants RR, den Dunnen JT, Bushby K, van der Maarel SM. AHNAK, a novel component of the dysferlin protein complex, redistributes to the cytoplasm with dysferlin during skeletal muscle regeneration. FASEB J 21: 732–742, 2007. [DOI] [PubMed] [Google Scholar]
- 69.Hwang M, Ko JK, Weisleder N, Takeshima H, Ma J. Redox-dependent oligomerization through a leucine zipper motif is essential for MG53-mediated cell membrane repair. Am J Physiol Cell Physiol 301: C106–C114, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Idone V, Tam C, Goss JW, Toomre D, Pypaert M, Andrews NW. Repair of injured plasma membrane by rapid Ca2+-dependent endocytosis. J Cell Biol 180: 905–914, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Illa I, Serrano-Munuera C, Gallardo E, Lasa A, Rojas-García R, Palmer J, Gallano P, Baiget M, Matsuda C, Brown RH. Distal anterior compartment myopathy: a dysferlin mutation causing a new muscular dystrophy phenotype. Ann Neurol 49: 130–134, 2001. [PubMed] [Google Scholar]
- 72.Jaiswal JK, Marlow G, Summerill G, Mahjneh I, Mueller S, Hill M, Miyake K, Haase H, Anderson LV, Richard I, Kiuru-Enari S, McNeil PL, Simon SM, Bashir R. Patients with a non-dysferlin Miyoshi myopathy have a novel membrane repair defect. Traffic 8: 77–88, 2007. [DOI] [PubMed] [Google Scholar]
- 73.Jia Y, Chen K, Lin P, Lieber G, Nishi M, Yan R, Wang Z, Yao Y, Li Y, Whitson BA, Duann P, Li H, Zhou X, Zhu H, Takeshima H, Hunter JC, McLeod RL, Weisleder N, Zeng C, Ma J. Treatment of acute lung injury by targeting MG53-mediated cell membrane repair. Nat Commun 5: 4387, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Jimenez AJ, Maiuri P, Lafaurie-Janvore J, Divoux S, Piel M, Perez F. ESCRT machinery is required for plasma membrane repair. Science 343: 1247136, 2014. [DOI] [PubMed] [Google Scholar]
- 75.Johnson CP, Chapman ER. Otoferlin is a calcium sensor that directly regulates SNARE-mediated membrane fusion. J Cell Biol 191: 187–197, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Jung SY, Ko YG. TRIM72, a novel negative feedback regulator of myogenesis, is transcriptionally activated by the synergism of MyoD (or myogenin) and MEF2. Biochem Biophys Res Commun 396: 238–245, 2010. [DOI] [PubMed] [Google Scholar]
- 77.Kerr JP, Ward CW, Bloch RJ. Dysferlin at transverse tubules regulates Ca2+ homeostasis in skeletal muscle. Front Physiol 5: 89, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Kerr JP, Ziman AP, Mueller AL, Muriel JM, Kleinhans-Welte E, Gumerson JD, Vogel SS, Ward CW, Roche JA, Bloch RJ. Dysferlin stabilizes stress-induced Ca2+ signaling in the transverse tubule membrane. Proc Natl Acad Sci USA 110: 20831–20836, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Keyel PA, Loultcheva L, Roth R, Salter RD, Watkins SC, Yokoyama WM, Heuser JE. Streptolysin O clearance through sequestration into blebs that bud passively from the plasma membrane. J Cell Sci 124: 2414–2423, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Kim SC, Kellett T, Wang S, Nishi M, Nagre N, Zhou B, Flodby P, Shilo K, Ghadiali SN, Takeshima H, Hubmayr RD, Zhao X. TRIM72 is required for effective repair of alveolar epithelial cell wounding. Am J Physiol Lung Cell Mol Physiol 307: L449–L459, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Kobayashi K, Izawa T, Kuwamura M, Yamate J. Dysferlin and animal models for dysferlinopathy. J Toxicol Pathol 25: 135–147, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Kovtun O, Tillu VA, Ariotti N, Parton RG, Collins BM. Cavin family proteins and the assembly of caveolae. J Cell Sci 128: 1269–1278, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Lee CS, Yi JS, Jung SY, Kim BW, Lee NR, Choo HJ, Jang SY, Han J, Chi SG, Park M, Lee JH, Ko YG. TRIM72 negatively regulates myogenesis via targeting insulin receptor substrate-1. Cell Death Differ 17: 1254–1265, 2010. [DOI] [PubMed] [Google Scholar]
- 84.Lek A, Evesson FJ, Lemckert FA, Redpath GM, Lueders AK, Turnbull L, Whitchurch CB, North KN, Cooper ST. Calpains, cleaved mini-dysferlinC72, and L-type channels underpin calcium-dependent muscle membrane repair. J Neurosci 33: 5085–5094, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Lennon NJ, Kho A, Bacskai BJ, Perlmutter SL, Hyman BT, Brown RH Jr. Dysferlin interacts with annexins A1 and A2 and mediates sarcolemmal wound-healing. J Biol Chem 278: 50466–50473, 2003. [DOI] [PubMed] [Google Scholar]
- 86.Lin P, Zhu H, Cai C, Wang X, Cao C, Xiao R, Pan Z, Weisleder N, Takeshima H, Ma J. Nonmuscle myosin IIA facilitates vesicle trafficking for MG53-mediated cell membrane repair. FASEB J 26: 1875–1883, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Liu J, Aoki M, Illa I, Wu C, Fardeau M, Angelini C, Serrano C, Urtizberea JA, Hentati F, Hamida MB, Bohlega S, Culper EJ, Amato AA, Bossie K, Oeltjen J, Bejaoui K, McKenna-Yasek D, Hosler BA, Schurr E, Arahata K, de Jong PJ, Brown RH. Dysferlin, a novel skeletal muscle gene, is mutated in Miyoshi myopathy and limb girdle muscular dystrophy. Nat Genet 20: 31–36, 1998. [DOI] [PubMed] [Google Scholar]
- 88.Liu J, Zhu H, Zheng Y, Xu Z, Li L, Tan T, Park KH, Hou J, Zhang C, Li D, Li R, Liu Z, Weisleder N, Zhu D, Lin P, Ma J. Cardioprotection of recombinant human MG53 protein in a porcine model of ischemia and reperfusion injury. J Mol Cell Cardiol 80: 10–19, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Lorusso A, Covino C, Priori G, Bachi A, Meldolesi J, Chieregatti E. Annexin2 coating the surface of enlargeosomes is needed for their regulated exocytosis. EMBO J 25: 5443–5456, 2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Lostal W, Bartoli M, Roudaut C, Bourg N, Krahn M, Pryadkina M, Borel P, Suel L, Roche JA, Stockholm D, Bloch RJ, Levy N, Bashir R, Richard I. Lack of correlation between outcomes of membrane repair assay and correction of dystrophic changes in experimental therapeutic strategy in dysferlinopathy. PLos One 7: e38036, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Marg A, Schoewel V, Timmel T, Schulze A, Shah C, Daumke O, Spuler S. Sarcolemmal repair is a slow process and includes EHD2. Traffic 13: 1286–1294, 2012. [DOI] [PubMed] [Google Scholar]
- 92.Mariano A, Henning A, Han R. Dysferlin-deficient muscular dystrophy and innate immune activation. FEBS J 280: 4165–4176, 2013. [DOI] [PubMed] [Google Scholar]
- 93.Matsuda C, Kameyama K, Suzuki A, Mishima W, Yamaji S, Okamoto H, Nishino I, Hayashi YK. Affixin activates Rac1 via betaPIX in C2C12 myoblast. FEBS Lett 582: 1189–1196, 2008. [DOI] [PubMed] [Google Scholar]
- 94.Matsuda C, Kameyama K, Tagawa K, Ogawa M, Suzuki A, Yamaji S, Okamoto H, Nishino I, Hayashi YK. Dysferlin interacts with affixin (beta-parvin) at the sarcolemma. J Neuropathol Exp Neurol 64: 334–340, 2005. [DOI] [PubMed] [Google Scholar]
- 95.McNeil AK, Rescher U, Gerke V, McNeil PL. Requirement for annexin A1 in plasma membrane repair. J Biol Chem 281: 35202–35207, 2006. [DOI] [PubMed] [Google Scholar]
- 96.McNeil PL, Ito S. Gastrointestinal cell plasma membrane wounding and resealing in vivo. Gastroenterology 96: 1238–1248, 1989. [DOI] [PubMed] [Google Scholar]
- 97.McNeil PL, Ito S. Molecular traffic through plasma membrane disruptions of cells in vivo. J Cell Sci 96: 549–556, 1990. [DOI] [PubMed] [Google Scholar]
- 98.McNeil PL, Khakee R. Disruptions of muscle fiber plasma membranes. Role in exercise-induced damage. Am J Pathol 140: 1097–1109, 1992. [PMC free article] [PubMed] [Google Scholar]
- 99.McNeil PL, Kirchhausen T. An emergency response team for membrane repair. Nat Rev Mol Cell Biol 6: 499–505, 2005. [DOI] [PubMed] [Google Scholar]
- 100.Mellgren RL, Miyake K, Kramerova I, Spencer MJ, Bourg N, Bartoli M, Richard I, Greer PA, McNeil PL. Calcium-dependent plasma membrane repair requires m- or mu-calpain, but not calpain-3, the proteasome, or caspases. Biochim Biophys Acta 1793: 1886–1893, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Mellgren RL, Zhang W, Miyake K, McNeil PL. Calpain is required for the rapid, calcium-dependent repair of wounded plasma membrane. J Biol Chem 282: 2567–2575, 2007. [DOI] [PubMed] [Google Scholar]
- 102.Micheva KD, Holz RW, Smith SJ. Regulation of presynaptic phosphatidylinositol 4,5-biphosphate by neuronal activity. J Cell Biol 154: 355–368, 2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Mikoshiba K, Fukuda M, Moreira JE, Lewis FM, Sugimori M, Niinobe M, Llinás R. Role of the C2A domain of synaptotagmin in transmitter release as determined by specific antibody injection into the squid giant synapse preterminal. Proc Natl Acad Sci USA 92: 10703–10707, 1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Mishima W, Suzuki A, Yamaji S, Yoshimi R, Ueda A, Kaneko T, Tanaka J, Miwa Y, Ohno S, Ishigatsubo Y. The first CH domain of affixin activates Cdc42 and Rac1 through alphaPIX, a Cdc42/Rac1-specific guanine nucleotide exchanging factor. Genes Cells 9: 193–204, 2004. [DOI] [PubMed] [Google Scholar]
- 105.Miyake K, McNeil PL. Mechanical injury and repair of cells. Crit Care Med 31: S496–S501, 2003. [DOI] [PubMed] [Google Scholar]
- 106.Miyake K, McNeil PL. Vesicle accumulation and exocytosis at sites of plasma membrane disruption. J Cell Biol 131: 1737–1745, 1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Miyake K, McNeil PL, Suzuki K, Tsunoda R, Sugai N. An actin barrier to resealing. J Cell Sci 114: 3487–3494, 2001. [DOI] [PubMed] [Google Scholar]
- 108.Nabhan JF, Hu R, Oh RS, Cohen SN, Lu Q. Formation and release of arrestin domain-containing protein 1-mediated microvesicles (ARMMs) at plasma membrane by recruitment of TSG101 protein. Proc Natl Acad Sci USA 109: 4146–4151, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Nagaraju K, Rawat R, Veszelovszky E, Thapliyal R, Kesari A, Sparks S, Raben N, Plotz P, Hoffman EP. Dysferlin deficiency enhances monocyte phagocytosis: a model for the inflammatory onset of limb-girdle muscular dystrophy 2B. Am J Pathol 172: 774–785, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Ohsawa Y, Okada T, Kuga A, Hayashi S, Murakami T, Tsuchida K, Noji S, Sunada Y. Caveolin-3 regulates myostatin signaling. Mini-review. Acta Myol 27: 19–24, 2008. [PMC free article] [PubMed] [Google Scholar]
- 111.Ozorowski G, Milton S, Luecke H. Structure of a C-terminal AHNAK peptide in a 1:2:2 complex with S100A10 and an acetylated N-terminal peptide of annexin A2. Acta Crystallogr D Biol Crystallogr 69: 92–104, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Parsegian VA, Rand RP, Gingell D. Lessons for the study of membrane fusion from membrane interactions in phospholipid systems. Ciba Found Symp 103: 9–27, 1984. [DOI] [PubMed] [Google Scholar]
- 113.Prokic I, Cowling BS, Laporte J. Amphiphysin 2 (BIN1) in physiology and diseases. J Mol Med 92: 453–463, 2014. [DOI] [PubMed] [Google Scholar]
- 114.Raucher D, Sheetz MP. Characteristics of a membrane reservoir buffering membrane tension. Biophys J 77: 1992–2002, 1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Reddy A, Caler EV, Andrews NW. Plasma membrane repair is mediated by Ca2+-regulated exocytosis of lysosomes. Cell 106: 157–169, 2001. [DOI] [PubMed] [Google Scholar]
- 116.Redpath GM, Woolger N, Piper AK, Lemckert FA, Lek A, Greer PA, North KN, Cooper ST. Calpain cleavage within dysferlin exon 40a releases a synaptotagmin-like module for membrane repair. Mol Biol Cell 25: 3037–3048, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Rescher U, Gerke V. S100A10/p11: family, friends and functions. Pflügers Arch 455: 575–582, 2008. [DOI] [PubMed] [Google Scholar]
- 118.Rezvanpour A, Santamaria-Kisiel L, Shaw GS. The S100A10-annexin A2 complex provides a novel asymmetric platform for membrane repair. J Biol Chem 286: 40174–40183, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Richard I, Roudaut C, Marchand S, Baghdiguian S, Herasse M, Stockholm D, Ono Y, Suel L, Bourg N, Sorimachi H, Lefranc G, Fardeau M, Sébille A, Beckmann JS. Loss of calpain 3 proteolytic activity leads to muscular dystrophy and to apoptosis-associated IkappaBalpha/nuclear factor kappaB pathway perturbation in mice. J Cell Biol 151: 1583–1590, 2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Rosenberger G, Gal A, Kutsche K. AlphaPIX associates with calpain 4, the small subunit of calpain, and has a dual role in integrin-mediated cell spreading. J Biol Chem 280: 6879–6889, 2005. [DOI] [PubMed] [Google Scholar]
- 121.Roux I, Safieddine S, Nouvian R, Grati M, Simmler MC, Bahloul A, Perfettini I, Le Gall M, Rostaing P, Hamard G, Triller A, Avan P, Moser T, Petit C. Otoferlin, defective in a human deafness form, is essential for exocytosis at the auditory ribbon synapse. Cell 127: 277–289, 2006. [DOI] [PubMed] [Google Scholar]
- 122.Scheffer LL, Sreetama SC, Sharma N, Medikayala S, Brown KJ, Defour A, Jaiswal JK. Mechanism of Ca2+-triggered ESCRT assembly and regulation of cell membrane repair. Nat Commun 5: 5646, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Schiavo G, Gu QM, Prestwich GD, Söllner TH, Rothman JE. Calcium-dependent switching of the specificity of phosphoinositide binding to synaptotagmin. Proc Natl Acad Sci USA 93: 13327–13332, 1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Schissel SL, Schuchman EH, Williams KJ, Tabas I. Zn2+-stimulated sphingomyelinase is secreted by many cell types and is a product of the acid sphingomyelinase gene. J Biol Chem 271: 18431–18436, 1996. [DOI] [PubMed] [Google Scholar]
- 125.Shen SS, Tucker WC, Chapman ER, Steinhardt RA. Molecular regulation of membrane resealing in 3T3 fibroblasts. J Biol Chem 280: 1652–1660, 2005. [DOI] [PubMed] [Google Scholar]
- 126.Steinhardt RA, Bi G, Alderton JM. Cell membrane resealing by a vesicular mechanism similar to neurotransmitter release. Science 263: 390–393, 1994. [DOI] [PubMed] [Google Scholar]
- 127.Sutton RB, Davletov BA, Berghuis AM, Südhof TC, Sprang SR. Structure of the first C2 domain of synaptotagmin I: a novel Ca2+/phospholipid-binding fold. Cell 80: 929–938, 1995. [DOI] [PubMed] [Google Scholar]
- 128.Swaggart KA, Demonbreun AR, Vo AH, Swanson KE, Kim EY, Fahrenbach JP, Holley-Cuthrell J, Eskin A, Chen Z, Squire K, Heydemann A, Palmer AA, Nelson SF, McNally EM. Annexin A6 modifies muscular dystrophy by mediating sarcolemmal repair. Proc Natl Acad Sci USA 111: 6004–6009, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Tam C, Idone V, Devlin C, Fernandes MC, Flannery A, He X, Schuchman E, Tabas I, Andrews NW. Exocytosis of acid sphingomyelinase by wounded cells promotes endocytosis and plasma membrane repair. J Cell Biol 189: 1027–1038, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Terasaki M, Miyake K, McNeil PL. Large plasma membrane disruptions are rapidly resealed by Ca2+-dependent vesicle-vesicle fusion events. J Cell Biol 139: 63–74, 1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Togo T, Steinhardt RA. Nonmuscle myosin IIA and IIB have distinct functions in the exocytosis-dependent process of cell membrane repair. Mol Biol Cell 15: 688–695, 2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Ueyama H, Kumamoto T, Nagao S, Masuda T, Horinouchi H, Fujimoto S, Tsuda T. A new dysferlin gene mutation in two Japanese families with limb-girdle muscular dystrophy 2B and Miyoshi myopathy. Neuromuscul Disord 11: 139–145, 2001. [DOI] [PubMed] [Google Scholar]
- 133.van der Kooi AJ, Frankhuizen WS, Barth PG, Howeler CJ, Padberg GW, Spaans F, Wintzen AR, Wokke JH, van Ommen GJ, de Visser M, Bakker E, Ginjaar HB. Limb-girdle muscular dystrophy in the Netherlands: gene defect identified in half the families. Neurology 68: 2125–2128, 2007. [DOI] [PubMed] [Google Scholar]
- 134.Waddell LB, Lemckert FA, Zheng XF, Tran J, Evesson FJ, Hawkes JM, Lek A, Street NE, Lin P, Clarke NF, Landstrom AP, Ackerman MJ, Weisleder N, Ma J, North KN, Cooper ST. Dysferlin, annexin A1, and mitsugumin 53 are upregulated in muscular dystrophy and localize to longitudinal tubules of the T-system with stretch. J Neuropathol Exp Neurol 70: 302–313, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Wang X, Xie W, Zhang Y, Lin P, Han L, Han P, Wang Y, Chen Z, Ji G, Zheng M, Weisleder N, Xiao RP, Takeshima H, Ma J, Cheng H. Cardioprotection of ischemia/reperfusion injury by cholesterol-dependent MG53-mediated membrane repair. Circ Res 107: 76–83, 2010. [DOI] [PubMed] [Google Scholar]
- 136.Weisleder N, Takeshima H, Ma J. Immuno-proteomic approach to excitation: contraction coupling in skeletal and cardiac muscle: molecular insights revealed by the mitsugumins. Cell Calcium 43: 1–8, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Weisleder N, Takizawa N, Lin P, Wang X, Cao C, Zhang Y, Tan T, Ferrante C, Zhu H, Chen PJ, Yan R, Sterling M, Zhao X, Hwang M, Takeshima M, Cai C, Cheng H, Takeshima H, Xiao RP, Ma J. Recombinant MG53 protein modulates therapeutic cell membrane repair in treatment of muscular dystrophy. Sci Transl Med 4: 139ra185, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Wendland B, Miller KG, Schilling J, Scheller RH. Differential expression of the p65 gene family. Neuron 6: 993–1007, 1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Wenzel K, Carl M, Perrot A, Zabojszcza J, Assadi M, Ebeling M, Geier C, Robinson PN, Kress W, Osterziel KJ, Spuler S. Novel sequence variants in dysferlin-deficient muscular dystrophy leading to mRNA decay and possible C2-domain misfolding. Hum Mutat 27: 599–600, 2006. [DOI] [PubMed] [Google Scholar]
- 140.Wenzel K, Geier C, Qadri F, Hubner N, Schulz H, Erdmann B, Gross V, Bauer D, Dechend R, Dietz R, Osterziel KJ, Spuler S, Ozcelik C. Dysfunction of dysferlin-deficient hearts. J Mol Med 85: 1203–1214, 2007. [DOI] [PubMed] [Google Scholar]
- 141.Wenzel K, Zabojszcza J, Carl M, Taubert S, Lass A, Harris CL, Ho M, Schulz H, Hummel O, Hubner N, Osterziel KJ, Spuler S. Increased susceptibility to complement attack due to down-regulation of decay-accelerating factor/CD55 in dysferlin-deficient muscular dystrophy. J Immunol 175: 6219–6225, 2005. [DOI] [PubMed] [Google Scholar]
- 142.Yamaji S, Suzuki A, Kanamori H, Mishima W, Yoshimi R, Takasaki H, Takabayashi M, Fujimaki K, Fujisawa S, Ohno S, Ishigatsubo Y. Affixin interacts with alpha-actinin and mediates integrin signaling for reorganization of F-actin induced by initial cell-substrate interaction. J Cell Biol 165: 539–551, 2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Zhu H, Lin P, De G, Choi KH, Takeshima H, Weisleder N, Ma J. Polymerase transcriptase release factor (PTRF) anchors MG53 protein to cell injury site for initiation of membrane repair. J Biol Chem 286: 12820–12824, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]