

Abstract
The vast majority of species within the lanternshark genus Etmopterus harbour complex luminescent markings on their flanks, whose functional significance has long remained obscure. Recent studies, however, suggest these enigmatic photophore aggregations to play a role in intraspecific communication. Using visual modelling based on in vivo luminescence measurements from a common lanternshark species, we show that etmopterid flank markings can potentially work as a medium range signal for intraspecific detection/recognition. In addition, using molecular phylogenetic analyses, we demonstrate that the Etmopterus clade exhibits a greater than expected species richness given its age. This is not the case for other bioluminescent shark clades with no (or only few) species with flank markings. Our results therefore suggest that etmopterid flank markings may provide a way for reproductive isolation and hence may have facilitated speciation in the deep-sea.
Keywords: bioluminescence, diversification, evolution, shark, visual detection
1. Introduction
Sharks of the genus Etmopterus (Squaliformes: Etmopteridae) are among the most intriguing bioluminescent organisms. Their tiny photogenic organs contain an unknown light-producing chemistry [1], are controlled by a unique ‘hybrid’ physiological control involving both hormonal and neural components [2], and form a diversity of patterns whose function has puzzled ichthyologists for more than a century [3–5]. Recent advances, however, suggest their luminescence to be a versatile tool involved in varied behaviours including camouflage by counterillumination [6,7], aposematism [8] and intraspecific signalling [9–11]. While counterillumination and aposematism are well supported by behavioural, experimental and theoretical data, intraspecific signalling is currently only corroborated by circumstantial evidences i.e. sex- or clade/species-specific difference in bioluminescent patterns, especially at the level of the lateral photophore areas (‘flank markings’). Although they can be grouped into five main patterns, these bioluminescent markings are highly variable across species [12,13]. As a consequence, scientists have suggested these areas to favour sympatric speciation by promoting reproductive isolation [7,11]. Interestingly, species-specific bioluminescence has been recently demonstrated to increase speciation rate in deep-sea bony fishes [14]. A similar situation could explain the evolutionary success of the genus Etmopterus, which, with 38 currently described species, is among the most prolific shark genera [15].
In this work, we first use a recent theory for pelagic vision to test whether etmopterid bioluminescent flank markings could efficiently work as an intraspecific recognition device. Subsequently, we performed molecular phylogenetic analyses to test whether the presence of these lateral photophore areas correlates with a higher diversification rate within bioluminescent sharks as was demonstrated for myctophids, which also show lateral photophores [16].
2. Material and methods
2.1. In vivo bioluminescence recordings
Specimens from a common etmopterid species (Etmopterus spinax) were collected in the Raunefjord and transferred to seawater tanks placed in a dark cold (4°C) room at Espeland Marine Station (Norway).
A luminometer (Berthold FB12, Pforzheim, Germany) coupled to an optical fibre allowed in vivo recording of ventral and lateral luminescence intensities from several live specimens according to Claes et al. [6]. Values were corrected for fibre absorption and angular losses. For modelling purpose, all photophores from a single shark specimen were considered to have exactly the same intensity. This intensity was calculated by dividing lateral light output by lateral photophore density, which was estimated under a binocular microscope according to Claes et al. [10]. Photophore spacing was determined from photophore density assuming a square mosaic.
2.2. Visual modelling
The detection distances of lateral glows were calculated according to the theory developed by Nilsson et al. [17]. This distance depends on the intensity of downwelling daylight, thus on water depth and observer sighting direction. Therefore, as in Claes et al. [8], E. spinax was assumed to occur at ‘counterillumination depth’ where its silhouette, cloaked by ventral photophores, is invisible from below [6], and sighting direction was chosen to be horizontal given the body position of etmopterid lateral luminescent markings. Counterillumination depth was determined using the mean spacing (0.248 mm) and mean intensity (2.52×106 photons s−1) of ventral photophores from shark ‘α’ (a 43 cm total length (TL) male specimen that exhibited the brightest flank markings of our dataset) as inputs in the eqn 7 in Supplemental Information from Nilsson et al. [17]. Beam attenuation and back-scattering coefficients were set to 0.3 m−1 and 0.0385 m−1, respectively, to agree with the turbid waters of the fjords according to Nilsson et al. [16]. The horizontal detection distance of specimen α flank markings (photophore mean spacing=0.191 mm; photophore mean intensity=2.52×106 photons s−1), modelled as an extended bioluminescent source seen against a transparent background (the downwelling daylight), was subsequently calculated for a series of pupil diameters (0–150 mm) according to Nilsson et al. [17]. Measurements performed in a complete ontogenetic series further provided a precise delimitation of the shark's pupil diameter range. Photoreceptor cell diameter was set to 3 μm according to recent measurements of E. spinax rod photoreceptor [18].
2.3. Species richness curve
For estimating diversification and relative extinction rates, we used MEDUSA [19] implemented in the R module GEIGER [20]. Rate estimates are based on the chronogram in Straube et al. [11] pruned to squaliform sharks. The background diversification and extinction rates estimated in MEDUSA were subsequently used to calculate crown and stem limits during the last 70 Ma (5 Ma increments) using the bd.ms module in GEIGER [21]. The resulting data allowed generation of an expected species richness curve from different points in time. Using the crown.p and stem.p options of the bd.ms module in GEIGER [20,21], we further calculated the probabilities of obtaining the current bioluminescent shark clades Etmopterus, Centroscyllium and Aculeola, Trigonognathus, and Dalatiidae given species number, age, diversification rate r and extinction rate ε. Species numbers were derived from Pollerspöck & Straube [15] and clade ages refer to Straube et al. [11]; see the electronic supplementary material for details on the performed analysis.
3. Results
Luminescence from ventral and lateral (flank markings) photophores was recorded in eight adult E. spinax specimens (35.5–49.5 cm TL). Light intensities from these photogenic structures, which are under the same hormonal control, were significantly correlated (p=0.0116; figure 1a). Ventral photophore intensity, combined with the visual theory for pelagic vision developed by Nilsson et al. [16,17], allowed us to determine the counterillumination depth of specimen α, i.e. 203 and 291 m for heavily overcast and clear skies (sun at 45°), respectively. Using Nilsson et al.'s theory again but with lateral photophore intensity and pupil diameter measurements from a complete ontogenetic series of E. spinax (n=40; 12–55 cm TL), we then determined that, at counterillumination depth, flank markings of specimen α are detectable by conspecifics at 2.8–4.4 m, well resolved at 1.7–2.9 m and fully resolved at 1–1.7 m (figure 1b,c). In the absence of lateral photophores, specimen α would only be detected by conspecifics at 0.9–1.4 m (figure 1b,c).
Figure 1.

In vivo recordings and visual modelling of Etmopterus spinax luminescence. (a) Correlation between lateral and ventral (counterilluminating) photophore luminescence intensity. (b) Target (t) and background (b) pixels used in the theoretical visual modelling: t1=bioluminescent detection pixel, t2=gross discrimination pixel (allowing more than 50% of the lateral pattern to be seen), t3=fine discrimination pixel (allowing more than 95% of the lateral pattern to be seen), t4=non-bioluminescent (black) detection pixel. Scale bar, 5 cm. (c) Detection distance of target pixels at counterillumination depth according to the observer's pupil diameter. The shaded area represents the pupil diameter range observed from a complete E. spinax ontogenetic series.
MEDUSA estimated a background diversification rate r of 0.017 and a relative extinction rate ε of 0.82. A rate shift was detected at the split of the genus Etmopterus, increasing the net diversification rate r to 1. The Etmopterus clade exhibits exceptional species diversity given its age, whereas all other clades, which contain no or a minority of species with flank markings, fall within the 95% CI of expected species diversity given their ages (figure 2).
Figure 2.
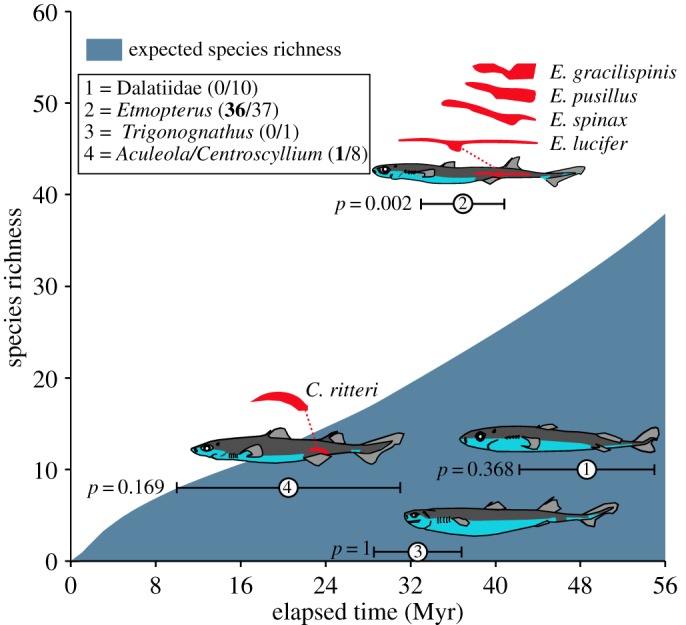
Expected species richness curve of bioluminescent sharks indicating the 95% CI for the expected number of species given clade age. Values in parentheses indicate, for each clade, the ratio between the number of species with flank markings (bold) versus the total species number. Only the Etmopterus clade, which contains the highest proportion of species with flank markings (97%), shows a significantly (p<0.01) higher species richness than expected. Flank markings are represented in red on shark drawings and on insets above them (enlarged view). To illustrate the morphological diversity of flank markings within Etmopterus genus, the flank marking shapes of four species were represented.
4. Discussion
Contrary to the photophores of other luminescent animals, which are often sparsely distributed or isolated, shark photophores form extended photogenic areas. Because they produce homogeneous bioluminescent emissions, these areas, when they occupy a ventral position, are particularly efficient to camouflage a silhouette [6,7,22]. Even though Etmopterus species display such ventral counterilluminating areas, they also display extended bioluminescent areas on their lateral sides, the so-called flank markings, which are hypothesized to be involved in intraspecific signalling [9–11]; their lateral body position and light kinetics (glow) indeed prevent them from being used in other bioluminescent behaviours such as camouflage, defence or prey capture. Collecting behavioural data demonstrating bioluminescent communication in elusive deep-sea animals such as bioluminescent sharks is logistically challenging. However, every luminous signal requires a target photoreceptor to be ecologically relevant [23]. This postulate allowed us to investigate bioluminescent shark communication via a physical modelling based on a recent theory for pelagic vision [18]. Using E. spinax as a model lanternshark species, this work demonstrates for the first time that these areas can be finely resolved at relatively long distance by conspecifics (figure 1c). Etmopterid flank markings exhibit a remarkable diversity of shapes that appears to be genetically supported [11]. When glowing, possibly in association with other clues (e.g. other bioluminescent areas), these lateral structures can therefore impact species recognition and potentially sexual selection in the darkness of the deep sea, as previously suggested [9,11]. Our theoretical approach also reveals that these markings are highly visible to predators with large pupils (e.g. large piscivorous fishes and marine mammals). Interestingly, captive E. spinax specimens often rotate their body right and left while swimming. This behaviour allows the continuous but directional photophore luminescence to generate intermittent signals similar to those of communicating fireflies or flashlight fishes, which are less conspicuous to predators [24,25].
From an evolutionary point of view, etmopterid flank photophores are considered to be an exaptation of ventral counterilluminating photophores [8]. In that context, the migration of ventral photophores towards the flanks would have occurred in the Palaeogene, potentially as an adaptation for bioluminescent signalling, probably during a deep-sea colonization event [7,11]. Here, we show that the extant Etmopterus clade exhibits a greater species richness than expected given its age, which is not the case for other bioluminescent shark clades with no (or only few) species with flank markings. Although the rapid diversification of Etmopterus sharks might be linked to a combination of different factors, our results agree with the idea that etmopterid flank markings are analogous to lateral photophores of lanternfishes (myctophids) [14], which may provide a way for reproductive isolation and facilitation of speciation in the darkness of the deep open oceans.
Supplementary Material
Acknowledgements
We thank T. Sørlie and A. Aadnesen for help in specimen collection and maintenance as well as the two anonymous reviewers whose constructive advices allowed substantial improvement of the quality of the manuscript. N.S. acknowledges the ichthyological staff at ZSM. J.M.C. and J.M. are, respectively, Postdoctoral Researcher and Research Associate at Fonds National de la Recherche Scientifique (FNRS, Belgium). This is a contribution to the Biodiversity Research Centre (BDIV) and to the Centre Interuniversitaire de Biologie Marine (CIBIM).
Ethics
Sharks were collected under Espeland Marine Station fishing permit (Permit 12/14048). Our protocol, including fish sacrifice, was approved by the National Animal Welfare Committee of Norway (fish handling approval no. 1664).
Data accessibility
All data supporting the paper are presented either in the main manuscript or in the electronic supplementary material.
Authors' contributions
J.M.C. and J.M. collected field data; D.-E.N. performed the modelling analysis; N.S. performed the phylogenetic analysis; J.M.C. wrote the manuscript and designed the figures. All authors gave final approval for publication.
Competing interests
The authors declare no competing interests.
Funding
Financial support was provided by the FNRS through a Fonds de la Recherche Fondamentale Collective grant (grant no. 2.4525.12) and travel grants to J.M.C. and J.M. D.-E.N. acknowledges The Swedish Research Council (grant no. 2011-4768) and the Knut and Alice Wallenberg Foundation (grant: Ultimate Vision).
References
- 1.Renwart M, Mallefet J. 2013. First study of the chemistry of the luminous system in a deep-sea shark, Etmopterus spinax Linnaeus, 1758 (Chondrichthyes: Etmopteridae). J. Exp. Mar. Biol. Ecol. 448, 214–219. (doi:10.1016/j.jembe.2013.07.010) [Google Scholar]
- 2.Claes JM, Mallefet J. 2015. Comparative control of luminescence in sharks: new insights from the slendertail lanternshark (Etmopterus molleri). J. Exp. Mar. Biol. Ecol. 467, 87–94. (doi:10.1016/j.jembe.2015.03.008) [Google Scholar]
- 3.Ohshima H. 1911. Some observations on the luminous organs of fishes. J. Coll. Sci. Imp. Univ. Tokyo. 27, 1–25. [Google Scholar]
- 4.Iwai T. 1960. Luminous organs of the deep-sea squaloid Centroscyllium ritteri Jordan and Fowler. Pac. Sci. 14, 51–54. [Google Scholar]
- 5.Reif WE. 1985. Function of scales and photophores in luminescent mesopelagic sharks. Acta Zool. 66, 111–118. (doi:10.1111/j.1463-6395.1985.tb00829.x) [Google Scholar]
- 6.Claes JM, Aksnes DL, Mallefet J. 2010. Phantom hunter of the fjords: camouflage by counterillumination in a shark (Etmopterus spinax). J. Exp. Mar. Biol. Ecol. 388, 28–32. (doi:10.1016/j.jembe.2010.03.009) [Google Scholar]
- 7.Claes JM, Nilsson DE, Straube N, Mallefet J. 2014. Iso-luminance counterillumination drove bioluminescent shark radiation. Sci. Rep. 4, 4328 (doi:10.1038/srep04328) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Claes JM, Dean MN, Nilsson DE, Hart NS, Mallefet J. 2013. A deepwater fish with lightsabers—dorsal spine-associated luminescence in a counterilluminating lanternshark. Sci. Rep. 3, 1308 (doi:10.1038/srep01308) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Claes JM, Mallefet J. 2009. Ontogeny of photophore pattern in the velvet belly lantern shark, Etmopterus spinax. Zoology 112, 433–441. (doi:10.1016/j.zool.2009.02.003) [DOI] [PubMed] [Google Scholar]
- 10.Claes JM, Mallefet J. 2010. Functional physiology of lantern shark (Etmopterus spinax) luminescent pattern: differential hormonal regulation of luminous zones. J. Exp. Biol. 213, 1852–1858. (doi:10.1242/jeb.041947) [DOI] [PubMed] [Google Scholar]
- 11.Straube N, Iglésias SP, Sellos DY, Kriwet J, Schliewen UK. 2010. Molecular phylogeny and node time estimation of bioluminescent lantern sharks (Elasmobranchii: Etmopteridae). Mol. Phylogen. Evol. 56, 905–917. (doi:10.1016/j.ympev.2010.04.042) [DOI] [PubMed] [Google Scholar]
- 12.Last PR, Burgess GH, Séret B. 2002. Description of six new species of lanternsharks of the genus Etmopterus (Squaloidea: Etmopteridae) from the Australasian region. Cybium 26, 203–223. [Google Scholar]
- 13.Ebert DA, Fowler S, Compagno L. 2013. Sharks of the world. Plymouth MA: Wild Nature Press. [Google Scholar]
- 14.Davis MP, Holcroft NI, Wiley EO, Sparks JS, Smith WL. 2014. Species-specific bioluminescence facilitates speciation in the deep sea. Mar. Biol. 161, 1139–1148. (doi:10.1007/s00227-014-2406-x) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Pollerspöck J, Straube N. 2015. www.shark-references.com, World Wide Web electronic publication, Version 2015 (accessed May 2015).
- 16.Nilsson DE, Warrant E, Johnsen S. 2014. Computational visual ecology in the pelagic realm. Phil. Trans. R. Soc. B 369, 20130038 (doi:10.1098/rstb.2013.0038) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Nilsson DE, Warrant EJ, Johnsen S, Hanlon R, Shashar N. 2012. A unique advantage for giant eyes in giant squid. Curr. Biol. 22, 683–688. (doi:10.1016/j.cub.2012.02.031) [DOI] [PubMed] [Google Scholar]
- 18.Claes JM, Partridge JC, Hart NS, Garza-Gisholt E, Ho HC, Mallefet J, Collin SP. 2014. Photon hunting in the twilight zone: visual features of mesopelagic bioluminescent sharks. PLoS ONE 9, e104213 (doi:10.1371/journal.pone.0104213) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Harmon LJ, Weir JT, Brock CD, Glor RE, Challenger W. 2008. GEIGER: investigating evolutionary radiations. Bioinformatics 24, 129–131. (doi:0.1093/bioinformatics/btm538) [DOI] [PubMed] [Google Scholar]
- 20.Alfaro ME, Santini F, Brock C, Alamillo H, Dornburg A, Rabosky DL, Carnevale G, Harmon LJ. 2009. Nine exceptional radiations plus high turnover explain species diversity in jawed vertebrates. Proc. Natl Acad. Sci. USA 106, 13 410–13 414. (doi:10.1073/pnas.0811087106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Magallon S, Sanderson MJ. 2001. Absolute diversification rates in angiosperm clades. Evolution 55, 1762–1780. (doi:10.1111/j.0014-3820.2001.tb00826.x) [DOI] [PubMed] [Google Scholar]
- 22.Claes JM, Ho H-C, Mallefet J. 2012. Control of luminescence from pygmy shark (Squaliolus aliae) photophores. J. Exp. Biol. 215, 1691–1699. (doi:10.1242/jeb.066704) [DOI] [PubMed] [Google Scholar]
- 23.Herring PJ. 1985. How to survive in the dark: bioluminescence in the deep-sea. Symp. Soc. Exp. Biol. 39, 323–350. [PubMed] [Google Scholar]
- 24.Morin JG, Harrington A, Nealson K, Krieger N, Baldwin TO, Hastings JW. 1975. Light for all reasons: versatility in the behavioral repertoire of the flashlight fish. Science 190, 74–76. (doi:10.1126/science.190.4209.74) [Google Scholar]
- 25.Lewis SM, Cratsley CK. 2008. Flash signal evolution, mate choice, and predation in fireflies. Annu. Rev. Entomol. 53, 293–321. (doi:10.1146/annurev.ento.53.103106.093346) [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
All data supporting the paper are presented either in the main manuscript or in the electronic supplementary material.