

Abstract
DNA methylation is a key epigenetic mechanism controlling DNA accessibility and gene expression. Blockade of DNA methylation can significantly affect pain behaviors implicated in neuropathic and inflammatory pain. However, the role of DNA methylation with regard to postoperative pain has not yet been explored. In this study we sought to investigate the role of DNA methylation in modulating incisional pain and identify possible targets under DNA methylation and contributing to incisional pain. DNA methyltranferase (DNMT) inhibitor 5-Aza-2′-deoxycytidine significantly reduced incision-induced mechanical allodynia and thermal sensitivity. Aza-2′-deoxycytidine also reduced hindpaw swelling after incision, suggesting an anti-inflammatory effect. Global DNA methylation and DNMT3b expression were increased in skin after incision, but none of DNMT1, DNMT3a or DNMT3b was altered in spinal cord or DRG. The expression of proopiomelanocortin Pomc encoding β-endorphin and Oprm1 encoding the mu-opioid receptor were upregulated peripherally after incision; moreover, Oprm1 expression was further increased under DNMT inhibitor treatment. Finally, local peripheral injection of the opioid receptor antagonist naloxone significantly exacerbated incision-induced mechanical hypersensitivity. These results suggest that DNA methylation is functionally relevant to incisional nociceptive sensitization, and that mu-opioid receptor signaling might be one methylation regulated pathway controlling sensitization after incision.
Introduction
Postoperative pain of moderate to severe intensity is experienced by 30–40% of patients after surgeries [1, 2]. Unrelieved postoperative pain is a common cause for unplanned hospital admission and contributes to suboptimal functional outcomes. Current therapeutic approaches to postoperative pain are limited by our narrow understanding of the underlying biological mechanisms. Therefore, examining the mechanisms involved in supporting pain after surgical incision would have significant value in addressing these problems.
Epigenetics refers to environmentally-supported changes in DNA and chromatin structure that do not alter DNA sequence. Such changes include the chemical modification of histone proteins, DNA methylation and microRNA expression [3]. DNA methylation is a covalent modification and involves the transfer of a methyl group to cysteine residues at CpG sites, which are 5'-CG-3' dinucleotide sequences in the genome, and clusters of CpG sites known as CpG islands, which are often in the promoter regions of genes [4]. This modification has been linked to many physiological and pathological processes, such as embryo development [5], aging [6], cancer [7, 8], psychiatric disorders [9, 10] and drug addiction [11, 12].
In addition to modulating the conditions noted above, a growing body of evidence has shown that blockade of DNA methylation can affect pain behaviors [13–15]. For example, chronic constriction injury increased the spinal global DNA methylation in rats, while intrathecal injection of the DNA methyltransferase (DNMT) inhibitor 5-azacytidine reversed this up-regulation and simultaneously attenuated the mechanical allodynia and thermal hyperalgesia [14]. Similarly, Viet et al. reported that treatment with agents promoting DNA demethylation resulted in mechanical and thermal antinociception in a mouse oral cancer model. The behavioral changes were correlated with mu-opioid receptor expression in the tumor tissue and associated neurons [15]. Consistent with these data, administration of a DNMT inhibitor reversed the hypermethylation of the mu-opioid receptor gene (Oprm1) and improved the analgesic effect of morphine treatment in a neuropathic pain model [16]. Furthermore, DNA methylation regulates several additional pain and analgesia-related genes in various pain models [16–18].
Clinically, chronic opioid administration is associated with increased DNA methylation at the LINE-1 global methylation site, which is correlated with pain severity [19]. Chronic stress was discovered to be associated with up-regulation of DNMT1-associated methylation of the cannabinoid receptor 1 (Cnr1) promoter and reduced CNR1 expression in DRG mediated chronic stress-induced increases in visceral pain [13]. Qi et al. demonstrated that demethylation of CpG sites of the cystathionine-[beta]-synthetase (Cbs) gene promoter region caused increased hydrogen sulfide production in DRG samples and contributes to inflammatory pain in rats [17]. Despite these discoveries, there is little information available concerning how DNA methylation might regulate postoperative pain.
The goal of our study was to identify the role of DNA methylation in modulating pain after incision. In addition to identifying the behavioral effects of a DNMT inhibitor, we sought to identify and confirm methylation-regulated targets contributing to incisional pain including the Oprm1 gene and possibly others.
Materials and Methods
Animal use
All experimental protocols were reviewed and approved by Veterans Affairs Palo Alto Healthcare System Institutional Animal Care and Use Committee prior to beginning the work. All protocols conform to the guidelines for the study of pain in awake animals as established by the International Association for the Study of Pain. Male mice 8–9 weeks old of the C57BL/6J strain (weighting 20–25 gram) were obtained from Jackson Laboratory (Bar Harbor, ME). Mice were housed four per cage and maintained on a 12-h light/dark cycle and an ambient temperature of 22 ± 1°C, with food and tap water available ad libitum. For behavior tests and paw thickness measurement, mice were randomized into four groups (n = 6–8); for methylation measurement and genes expression, mice were randomized into two groups (n = 5–6).
Hindpaw incision
The hindpaw incision model in mice was performed in our laboratory as described in previous studies [20, 21]. Briefly, mice were anesthetized using isoflurane 2–3% delivered through a nose cone. After sterile preparation with alcohol, a 5 mm longitudinal incision was made with a number 11 scalpel on the plantar surface of the right hindpaw. The incision was sufficiently deep to divide deep tissue including the plantaris muscle longitudinally. After controlling bleeding, a single 6–0 nylon suture was placed through the midpoint of the wound and antibiotic ointment was applied. Mice used in these experiments did not show evidence of infection in the paws at the time of behavioral or biochemical assays.
Drug administration
5-AZA-CdR (5-Aza-2′-deoxycytidine)(Sigma-Aldrich, St. Louis, MO) was freshly dissolved in 0.9% saline (Sigma). The concentration was adjusted to 40 μg/100 μl so that a 4 mg/kg dose could be administrated intraperitoneally in a volume of 100 μl/10g body weight. Mice received either 5-AZA-CdR solution or matching vehicle 24h and 2h prior to incision and once daily for 3 days after incision. The opioid receptor antagonist naloxone hydrochloride dehydrate (Sigma-Aldrich) was freshly dissolved in 0.9% saline. Mice received either naloxone (10μg) or vehicle intraplantar on day 3 after incision in a volume of 15μl. The dosage selection of 5-AZA-CdR is based on the finding that at this dose it effectively reduced DNA methylation and produces little toxicity in mice in other experiments [22, 23].
Nociceptive testing
All nociceptive testing was done with the experimenter blind to drug treatment.
Mechanical hypersensitivity
Mechanical nociceptive thresholds were assayed using von Frey filaments according to a modification of the “up-down” algorithm described by Chaplan et al [24], as described previously [20, 21]. Mice were placed on wire mesh platforms in clear cylindrical plastic enclosures of 10 cm diameter and 30 cm height. After 20 minutes of acclimation, fibers of sequentially increasing stiffness with initial bending force of 0.2 gram were applied to the plantar surface of the hindpaw adjacent to the incision, just distal to the first set of foot pads and left in place 5 sec with enough force to slightly bend the fiber. Withdrawal of the hindpaw from the fiber was scored as a response. When no response was obtained, the next stiffer fiber in the series was applied in the same manner. If a response was observed, the next less stiff fiber was applied. Testing proceeded in this manner until 4 fibers had been applied after the first one causing a withdrawal response allowing the estimation of the mechanical withdrawal threshold using a curve fitting algorithm [25].
Thermal sensitization
Paw withdrawal response latencies to noxious thermal stimulation were measured using the method of Hargreaves et al. [26] as we have modified for use with mice 24. In this assay, mice were placed on a temperature-controlled glass platform (29°C) in a clear plastic enclosure. After 30 min of acclimation, a beam of focused light was directed towards the same area of the hindpaw as described for the von Frey assay. A 20 s cutoff was used to prevent tissue damage. The light beam intensity was adjusted to provide an approximate 10s baseline latency in control mice. Three measurements were made per animal per test session separated by at least one minute.
Paw edema
A laser (4381 Precicura, Limab, Goteborg, Sweden) sensor technique was used to measure the dorsal-ventral thickness of the hindpaw [27]. After induction of rapid and brief anesthesia with isoflurane, the mouse was held vertically with hindpaw resting on a table top under the laser. By applying a small metal rod to the top of ankle joint, the paw was gently held flat on the table. Using optical triangulation, a distance measuring sensor (200 mm range, 0.01 mm resolution) was used to determine the difference of the distance from the top of the hindpaw to the table top (dorsal-ventral paw thickness). Three measurements were made per paw per animal.
RNA isolation and real-time quantitative polymerase chain reaction (PCR) amplification
Mice were first euthanized by carbon dioxide asphyxiation and an ovular full-thickness patch of skin providing 1.5- to 2-mm margins surrounding the hindpaw incisions was collected. Spinal cord tissue was harvested by extrusion. Lumbar spinal cord segments were dissected on a chilled surface. Dorsal root ganglia (DRG) (L3-S1) were dissected using low power binocular magnification as described previously [28]. Dissected tissue was then quick-frozen in liquid nitrogen and stored at -80°C until required for analysis. For real-time quantitative PCR, total RNA was isolated from skin and spinal cord using the RNeasy Mini Kit (Qiagen, Valencia, CA) according to the manufacturer's instructions. The purity and concentration were determined spectrophotometrically. The total RNA samples were reverse transcribed into complementary DNA using a First Strand complementary DNA Synthesis Kit (Invitrogen, Carlsbad, CA). Real-time PCR was performed in an ABI prism 7900HT system (Applied Biosystems, Foster City, CA). All PCR experiments were performed using the SYBR Green I master kit (Applied Biosystems). The primer sequences for 18S message RNA (mRNA) are aagacgatcagataccgtcgtag (forward) and tccgtcaattcctttaagtttca (reverse). All of the other primer sets were purchased from SABiosciences (Valencia, CA). The amplification parameters were described previously [21]. Melting curves were performed to document single product formation and agarose electrophoresis confirmed product size. All the primers were purchased from SABiosciences (SABiosciences, Valencia, CA). As negative controls, RNA samples that were not reverse transcribed were run. Data were normalized to 18S mRNA expression.
DNA isolation and global DNA methylation
Genomic DNA was isolated with the GenElute™ Mammalian Genomic DNA Miniprep Kit (Sigma-Aldrich) according to manufacter's instructions. DNA was treated with RNAase to remove RNA contaminants. Isolated DNA was quantified using an ND-1000 NanoDrop spectrophotometer (Thermo Scientific, Wilmington, DE). Assessment of global DNA methylation status was accomplished by using the MethylFlash Methylated DNA Quantification Kit (Epigentek, Farmingdale, NY) according to manufacter's instructions. The methylated fraction of DNA was identified using 5-methylcytosine monoclonal antibodies and quantified by an enzyme-linked immunosorbent assay–like reaction. The levels of methylated DNA were measured at 450nm with a VERSmax Microplate Reader (Molecular Devices, Sunnyvale, CA). The percentage of 5-mC was calculated using the formula provided in the kit procedure and were normalized to percentage of control.
Statistical analysis
All data are expressed as mean ± standard error of the mean (SEM). The time course changes of gene expression and global methylation within each group was analyzed by one-way ANOVA with post-hoc Bonferroni's testing for multiple comparisons. Comparisons between two groups for behavior test, paw thickness and gene expression were analyzed by unpaired t-test at each timepoint. P values less than 0.05 were considered significant (Prism 5; GraphPad Software, La Jolla, CA).
Results
Effects of DNMT inhibition on incision-induced nociceptive sensitization and edema
To determine whether DNA methylation alters incision-induced sensitization, the DNMT inhibitor 5-AZA-CdR was administrated daily to incised mice. Fig 1A and 1B present data demonstrating that systemic administration of 5-AZA-CdR significantly attenuated incision-induced mechanical hypersensitivity and thermal sensitivity. 5-AZA-CdR had no effect on the nociceptive thresholds of control animals (Fig 1).
Fig 1. Assessment of DNMT inhibitor 5-Aza-2′-deoxycytidine on incision-induced mechanical and thermal sensitization.

Blocking DNMT attenuated incision-induced mechanical hypersensitivity (A) and thermal sensitization (B). Animals received intraperitoneal injection of 5-Aza-2′-deoxycytidine (4 mg/kg) or vehicle (saline) 24h and 2h prior to incision and once daily for 3 days after incision. Values are displayed as the mean ± SEM. N = 8. **p<0.01, 5-AZA-CdR + INC group vs. Veh + INC group. Veh = vehicle; INC = incision; 5-AZA-CdR = 5-Aza-2′-deoxycytidine.
To determine the ability of 5-AZA-CdR treatment to reduce indices of the inflammatory response in incised animals, we measured changes in paw thickness at time points up to 3 days post-incision. 5-AZA-CdR significant reduced incision-induced edema at the 1–3 day time points (Fig 2).
Fig 2. Effect of DNMT inhibitor 5-Aza-2′-deoxycytidine on paw edema induced by incision.
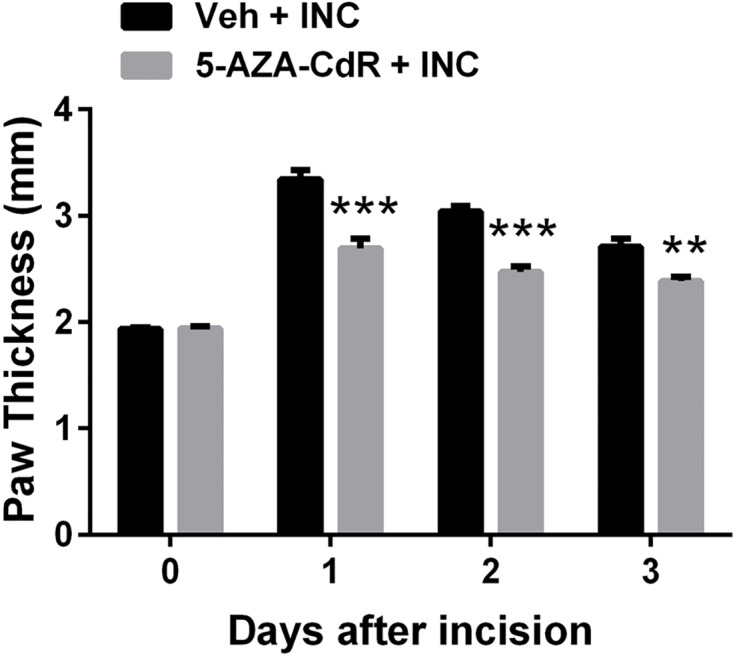
5-Aza-2′-deoxycytidine significantly reduced incision-induced hindpaw edema. Values are displayed as the mean ± SEM. N = 8. ** p<0.01, *** p<0.001, vs. vehicle treated group. Veh = vehicle; INC = incision; 5-AZA-CdR = 5-Aza-2′-deoxycytidine.
Alteration of DNMT expression and global DNA methylation after incision
Because a DNMT inhibition significantly attenuated incision-induced nociceptive sensitization, we hypothesized that pro-nociceptive changes may be associated with enhanced DNA methylation. To test this idea we first examined the expression of DNA methyltransferases (DNMTs) in incised skin and spinal cord tissue. The mammalian genome encodes three active DNMTs: DNMT1, DNMT3a and DNMT3b. DNMT1 is described as the maintenance methyltransferase, while DNMT3a and DNMT3b are methyltransferases often expressed de novo [29, 30]. We found that the mRNA level of DNMT3b was significantly increased (6h after incision in skin tissue), and the mRNA levels of DNMT1 and DNMT3a were not changed in skin tissue (Fig 3A–3C). In addition, none of the DNMTs’ expression was altered in the spinal cord (Fig 3D–3F) or DRG (Fig 3G–3I).
Fig 3. Expression of DNMTs in skin (A-C), spinal cord (D-F) and DRG (G-I) tissues after incision.

The mRNA level of DNMT3b (C) was significantly increased at 6h in skin after incision, while DNMT1 (A) and DNMT3a (B) did not change in skin. The mRNA levels of DNMT1 (D and G), DNMT3a (E and H), or DNMT3b (F and I) were not altered in spinal cord or DRG after incision, respectively. Values are displayed as the mean ± SEM, n = 6. ***p<0.001 vs. day 0 (before incision).
Since DNMT3b mRNA level was upregulated in skin after incision, we examined global DNA methylation in skin and found that the global DNA methylation was increased in skin tissue at day 1 and day 3 after incision (Fig 4). Additionally, we also examined the global methylation in spinal cord and no changes were found in spinal cord tissue across this time course (data not shown).
Fig 4. The changes of global DNA methylation after incision.
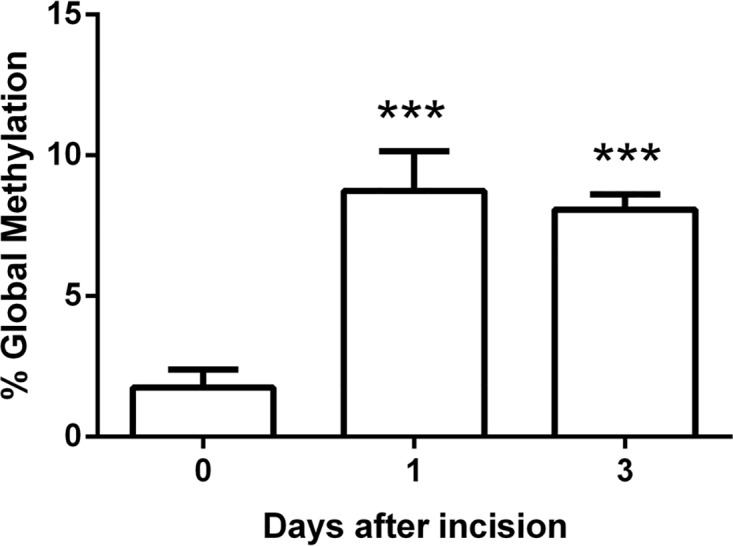
The global DNA methylation was increased in skin at day 1 and day 3 after incision. Values are displayed as the mean ± SEM, n = 5. ***p<0.001 vs. day 0 (before incision).
Effects of 5-AZA-CdR on anti-inflammatory cytokine gene expression
DNMT inhibition leads to the demethylation of gene promoters and subsequent gene activation. Here, we examined the expression of Il-1α, Il-4 and Il-10 mRNA levels after incision under 5-AZA-CdR treatment, because these anti-inflammatory cytokines have been demonstrated to have antinociceptive activities in various pain models [31–33], and are regulated via DNA methylation [34–36]. Both Il-10 and Il-4 genes were upregulated after incision. However, expression of none of these genes was altered by 5-AZA-CdR treatment (Fig 5).
Fig 5. Effect of DNMT inhibitor 5-Aza-2′-deoxycytidine on Il-1α (A), Il-4 (B) and Il-10 (C) mRNA levels in skin tissue after incision.

Incision induced the up-regulation of Il-4 and Il-10 expression, however, DNMT inhibitor failed to alter Il-1α (A), Il-4 (B) and Il-10 (C) expression compared with the vehicle treated group. Values are displayed as the mean ± SEM. N = 5. # p<0.05, ## p<0.01, ### p<0.001 vs. day 0 (before incision). Veh = vehicle; INC = incision; 5-AZA-CdR = 5-Aza-2′-deoxycytidine.
Effects of 5-AZA-CdR on mu-opioid receptor and endogenous opioid gene expression
Activation of peripheral or central mu-opioid receptors can produce analgesic effects in various animal pain models and humans [16, 37, 38]. Furthermore, mu-opioid receptor expression was previously demonstrated to be epigenetically regulated via DNA methylation [16, 19, 39]. Therefore, we examined the peripheral and central Oprm1 expression after incision and/or DNMT inhibitor treatment. The mRNA level of Oprm1 was increased in skin after incision and further increased in mice treated with 5-AZA-CdR (Fig 6A). Additionally, 5-AZA-CdR itself caused a transient upregulation of Oprm1 mRNA expression in skin at day 0 (Fig 6A), without significant changes at day 1 and day 3 (data not shown), which indicated that 5-AZA-CdR regulates Oprm1 gene expression in the control and incision settings. Neither incision nor 5-AZA-CdR altered the mRNA levels of Oprm1 in spinal cord (Fig 6B) or DRG (Fig 6C).
Fig 6. Changes in mu-opioid receptor mRNA (Oprm1) expression after incision.

(A) The mRNA level of Oprm1 significantly increased after incision in skin, and further increased in mice treated with 5-Aza-2′-deoxycytidine before and at day 1 after incision. The mRNA level of Oprm1 did not alter in spinal cord (B) or DRG (C) after incision. Values are displayed as the mean ± SEM. N = 6. **p<0.01, 5-AZA-CdR + INC group vs. Veh + INC group. ## p<0.01, ### p<0.001 vs. day 0 (before incision). Veh = vehicle; INC = incision; 5-AZA-CdR = 5-Aza-2′-deoxycytidine.
Next, since only peripheral Oprm1 expression was changed after incision and DNMT inhibitor treatment, we examined the peripheral expression of endogenous opioid peptide encoding genes, proopiomelanocortin (Pomc) and proenkephalin (Penk). As shown in Fig 7, incision induced the upregulation of Pomc mRNA expression in skin and the downregulation of Penk mRNA levels, however, 5-AZA-CdR failed to alter Pomc and Penk expression compared with vehicle treated group.
Fig 7. Changes in endogenous opioid peptides encoding genes, proopiomelanocortin (POMC) and proenkephalin (PENK) mRNA levels in skin after incision.

(A) The mRNA level of POMC significantly increased after incision in skin. (B) The mRNA level of PENK significantly decreased after incision in skin. Values are displayed as the mean ± SEM. N = 6. # p<0.05, ## p<0.01, ### p<0.001 vs. day 0 (before incision). Veh = vehicle; INC = incision; 5-AZA-CdR = 5-Aza-2′-deoxycytidine.
We examined the expression of other methylation-regulated pain-related genes in skin and spinal cord tissues, including microRNA203 [20, 40], encoding the endothelin B receptor (EDNRB) [41, 42], galanin [43, 44] and dipeptidyl peptidase 4 [45, 46]. However, none of them showed epigenetic regulation under DNMT inhibitor treatment (data not shown).
Effects of intraplantar injection of opioid antagonist, naloxone on 5-AZA-CdR attenuated incisional pain
Since Pomc and Oprm1 were up-regulated in skin tissue after incision and (for Oprm1) further increased in mice treated with 5-AZA-CdR, we determined whether the peripheral mu-opioid receptor signaling pathway was involved in nociceptive sensitization after incision. Peri-incisional administration of the opioid receptor antagonist naloxone (10 μg intraplantar) at day 3 after incision significantly exacerbated mechanical hypersensitivity consistent with an active process of local opioid analgesia (Fig 8A). Furthermore, local injection of naloxone (10 μg) significantly reduced threshold to the same level observed in the inhibitor treated mice (Fig 8B). This is consistent with opioid tone being involved in mediating the effects of methylation after incision. Naloxone did not have any effect on the nociceptive thresholds of non-incised control animals (Fig 8A).
Fig 8. Assessment of peripheral mu-opioid receptor signaling on nociceptive sensitization after incision.

Intraplantar injection of opioid receptor antagonist naloxone at day 3 after incision significantly reversed 5-Aza-2′-deoxycytidine attenuated, incision-induced mechanical hypersensitivity (A), and significantly exacerbated incisional mechanical hypersensitivity as well (B). Arrow indicates the time point for incision operation. Values are displayed as the mean ± SEM, n = 6, ** p<0.01, *** p<0.001, 5-AZA-CdR + INC + Naloxone group vs 5-AZA-CdR + INC + Veh group. Veh = vehicle; INC = incision; 5-AZA-CdR = 5-Aza-2′-deoxycytidine.
Discussion
Over the past decade, a significant amount of work has been done to elucidate the mechanisms involved in nociceptive sensitization after surgical incision. Recently, attention has to a degree shifted toward understanding how epigenetic mechanisms regulate postoperative pain [21, 47, 48]. DNA methylation is a key epigenetic mechanism controlling transcription factor binding and gene expression. However, the role of DNA methylation with regard to postoperative pain has not yet been explored. In this study, our principal observations were: (1) Blockade of DNMT attenuated incision-induced nociceptive sensitization as well as incision-induced edema. (2) Global DNA methylation and DNMT3b expression were increased in skin after incision, but none of DNMT1, DNMT3a or DNMT3b was altered in spinal cord or DRG after incision. (3) The expression of Pomc encoding β-endorphin and Oprm1 encoding mu-opioid receptor was upregulated peripherally after incision; moreover, Oprm1 was further increased under DNMT inhibitor treatment. (4) Peripheral injection of opioid receptor antagonist, naloxone significantly exacerbated incision-induced mechanical hypersensitivity. Together these observations suggest that DNA methylation is functionally relevant to incisional nociceptive sensitization, and that peripheral mu-opioid receptor signaling might be one DNA methylation regulated pathway controlling sensitization after incision.
Available reports from the literature are largely similar in their findings that DNMT inhibition provides antinociceptive effects [13–15]. 5-Azacytidine and its more selective deoxy analog, 5-aza-2-deoxycytidine (5-AZA-CdR), are well-known DNA methylation inhibitors, and both have been approved by the US food and Drug Administration for the treatment of myelodysplastic syndrome [49]. Both drugs inhibit the DNMTs’ activity by substituting for cytosine residues and forming covalent bonds with the DNMTs, resulting in demethylation [49]. A previous report showed that the systemic administration of 5-AZA-CdR produced significant antinociception to mechanical stimuli in the mice oral cancer model [15]. Spinal cord methylation was suggested to support nociception; intrathecal injection of 5-azacytidine attenuated mechanical and thermal sensitivity in chronic constriction injury-induced neuropathic pain model [14]. Additionally, knockdown of DNMT1expression prevented chronic stress induced increases in visceral pain [13]. Our data are functionally consistent with all of these observations in that 5-AZA-CdR reduced incision-induced mechanical and thermal sensitivity. However, since 5-AZA-CdR treatment started before incision and continued after, it is unclear at what specific time point DNMT inhibition might be most effective. Additionally, our data also showed that 5-AZA-CdR reduced incision-induced edema. However, since the levels of several pain-related anti-inflammatory cytokines were unchanged by DNMT inhibition, the mechanisms for the effects on edema are unclear.
DNMTs transfer a methyl group from S-adenosyl-methionine to the fifth carbon of cytosine residues in the genomic CpG dinucleotides. DNMT1, DNMT3a, DNMT3b are the principle isoforms that establish and maintain the patterns of DNA methylation in mammals [50]. Generally, DNMT1 is described as the maintenance methyltransferase, copying methylation marks throughout cell division [29]. DNMT3a and DNMT3b are de novo methyltransferases, methylating specific DNA marks in response to environmental factors [30]. It has been observed that persistent pain alters the expression of DNMTs. For example, CFA-induced inflammatory pain caused the global upregulation of DNMTs in spinal cord dorsal horn, while spared nerve injury lead to the downregulation of DNMT1 in spinal cord [51]. More recently, Pollema-Mays et al. using the spared nerve injury model found that the DNMT1 and DNMT3a were moderately upregulated in DRG, and DNMT3b showed a robust increase in DRG [52]. In our present study, we only found DNMT3b to be upregulated after incision, while none of DNMTs was changed in spinal cord. Baubec et al. observed that DNMT3b has a prominent genome-wide role in controlling transcription by selectively binding to the bodies of transcribed genes leading to their preferential methylation [53]. Interesting, among the DNMTs, the DNMT3B has the most alternatively spliced isoforms with about 40 different DNMT3B isoforms identified so far which influence the functional properties of this enzyme [54, 55]. It is not clear which of these variants is operative in our model.
Global methylation is a measure of the overall state of the DNA methylation and has long-range consequences on genome function [56, 57]. Consistent with the up-regulation of DNMT expression in peri-incisional tissue, we also observed the increased global DNA methylation. There were no such changes measured in spinal cord. Therefore, the changes in global DNA methylation and DNMT expression appear to be tissue- and disease-specific in response to various types of injuries. Similarly, it has been reported that chronic peripheral nerve injury caused decreases in global methylation in prefrontal cortex and amygdala, but not in thalamus and visual cortex [58]. Furthermore, overall tissue methylation levels do not always correlate with methylation changes at the local level. For example, Meller et al reported global methylation was reduced following preconditioning ischemia in primary neuronal culture; however, selective chromosomes showed enhanced hypermethylation [59]. In addition, opioid receptor and endogenous gene expression are regulated by the other epigenetic mechanisms, such as histone acetylation and microRNA regulation [60, 61]. Our observations were that Oprm1 expression actually increased after incision despite increased global skin methylation levels, but blockade of DNMT further increased Oprm1 expression. This suggests methylation limits or counterbalances other incision-induced processes.
Accumulating evidence shows that the expression of mu-opioid receptor and its relevant ligands are altered in peripheral and central nervous system tissue in various pain models and contribute to regulating nociceptive sensitization [37, 38, 62, 63]. Furthermore, this receptor is epigenetically regulated via DNA methylation [16, 19, 64]. In general, hypermethylation of gene promoters is associated with transcriptional repression by preventing the binding of transcription factors for their binding sites, whereas, hypomethylation results in gene activation [50]. For example, Zhou et al. observed that peripheral nerve injury caused a decreased mu-opioid receptor expression with an increased methylation status of Oprm1 gene promoter in DRG tissue. Treatment with the DNMT inhibitor 5-AZA-CdR reversed the decreased Oprm1 expression via demethylation of its promoter and improved the analgesic effects of morphine administration [16]. In the current study, we showed that both Oprm1 and its ligand Pomc were significantly increased in skin after incision, and Oprm1 gene expression was further increased when DNMT was blocked. Pharmacological experiments showed that intraplantar injection of an opioid receptor antagonist exaggerated 5-AZA-CdR-attenuated mechanical allodynia after incision, suggesting that the up-regulation of peripheral mu-opioid receptor leads to the decreased nociceptive sensitivity. Although mu-opioid receptor is generally considered to exert its anti-nociceptive actions within the CNS [65], peripheral mu-opioid receptor has become increasingly recognized as important for limiting sensitization in the settings of tissue injury and inflammation [66]. For example, it has been reported that Oprm1 is expressed in nerve terminals and keratinocytes in skin as well as on immune cells [67–69]. Furthermore, our previous studies demonstrated that endogenous opioid peptides (β-endorphin and others) were produced by neutrophils after incision [70]. Therefore, the upregulation of Oprm1 expression under 5-AZA-CdR treatment after incision may sensitize local or infiltrating cells to the effects of endogenous (or perhaps exogenous) opioid ligands. The reduction in edema similarly suggested local opioid receptor mediated anti-inflammatory effects. Additional studies have demonstrated that expression of pain-related genes microRNA203 [20, 40], encoding the endothelin B receptor (EDNRB) [41, 42], galanin [43, 44] and dipeptidyl peptidase 4 [45, 46] are also under the control of methylation-related epigenetic mechanisms. However, DNMT inhibition failed to alter their expression in setting of our incisional pain model.
In Summary, the present study is the first to show that incision induces changes in global DNA methylation and DNMT expression in skin, and suggests that regulation of DNA methylation can control nociceptive sensitization after incision. DNMT inhibition reduced incisional mechanical hypersensitivity which appears to be due at least in part to the up-regulation of Oprm1 expression. It is widely believed that large numbers of genes regulate nociception; DNA methylation can broadly control gene expression in setting of tissue injury potentially providing an overarching mechanism controlling some portion of this response [71–73]. Therefore, our theory that a DNMT inhibitor attenuated incisional sensitization via increasing Oprm1 expression likely explains only a portion of the inhibitor’s effects. Future studies might include the investigation of DNA methylation status of Oprm1 gene under DNMT regulation, as well as other groups of genes, e.g. by using array or sequencing technologies, and more broadly addressing the roles of DNMTs by using selective DNMT inhibitors or conditional knockdown strategies in the incisional pain model.
Data Availability
All relevant data are within the paper.
Funding Statement
The work was supported by NIH award GM079126 (Bethesda, MD, USA) and VA Merit Review award 5I01BX000881 (Washington D.C., USA). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Kehlet H, Holte K. Effect of postoperative analgesia on surgical outcome. British journal of anaesthesia. 2001;87(1):62–72. . [DOI] [PubMed] [Google Scholar]
- 2. Pyati S, Gan TJ. Perioperative pain management. CNS drugs. 2007;21(3):185–211. . [DOI] [PubMed] [Google Scholar]
- 3. Marsit CJ. Influence of environmental exposure on human epigenetic regulation. The Journal of experimental biology. 2015;218(Pt 1):71–9. 10.1242/jeb.106971 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Ushijima T. Detection and interpretation of altered methylation patterns in cancer cells. Nature reviews Cancer. 2005;5(3):223–31. 10.1038/nrc1571 . [DOI] [PubMed] [Google Scholar]
- 5. Razin A, Kafri T. DNA methylation from embryo to adult. Progress in nucleic acid research and molecular biology. 1994;48:53–81. . [DOI] [PubMed] [Google Scholar]
- 6. Jung M, Pfeifer GP. Aging and DNA methylation. BMC biology. 2015;13(1):7 10.1186/s12915-015-0118-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Ma X, Wang YW, Zhang MQ, Gazdar AF. DNA methylation data analysis and its application to cancer research. Epigenomics. 2013;5(3):301–16. 10.2217/epi.13.26 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Suzuki H, Maruyama R, Yamamoto E, Kai M. DNA methylation and microRNA dysregulation in cancer. Molecular oncology. 2012;6(6):567–78. 10.1016/j.molonc.2012.07.007 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Kato T, Iwamoto K. Comprehensive DNA methylation and hydroxymethylation analysis in the human brain and its implication in mental disorders. Neuropharmacology. 2014;80:133–9. 10.1016/j.neuropharm.2013.12.019 . [DOI] [PubMed] [Google Scholar]
- 10. Klengel T, Pape J, Binder EB, Mehta D. The role of DNA methylation in stress-related psychiatric disorders. Neuropharmacology. 2014;80:115–32. 10.1016/j.neuropharm.2014.01.013 . [DOI] [PubMed] [Google Scholar]
- 11. Tuesta LM, Zhang Y. Mechanisms of epigenetic memory and addiction. The EMBO journal. 2014;33(10):1091–103. 10.1002/embj.201488106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Bilinski P, Wojtyla A, Kapka-Skrzypczak L, Chwedorowicz R, Cyranka M, Studzinski T. Epigenetic regulation in drug addiction. Annals of agricultural and environmental medicine: AAEM. 2012;19(3):491–6. . [PubMed] [Google Scholar]
- 13. Hong S, Zheng G, Wiley JW. Epigenetic regulation of genes that modulate chronic stress-induced visceral pain in the peripheral nervous system. Gastroenterology. 2015;148(1):148–57 e7 10.1053/j.gastro.2014.09.032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Wang Y, Liu C, Guo QL, Yan JQ, Zhu XY, Huang CS, et al. Intrathecal 5-azacytidine inhibits global DNA methylation and methyl- CpG-binding protein 2 expression and alleviates neuropathic pain in rats following chronic constriction injury. Brain research. 2011;1418:64–9. 10.1016/j.brainres.2011.08.040 . [DOI] [PubMed] [Google Scholar]
- 15. Viet CT, Dang D, Ye Y, Ono K, Campbell RR, Schmidt BL. Demethylating drugs as novel analgesics for cancer pain. Clinical cancer research: an official journal of the American Association for Cancer Research. 2014;20(18):4882–93. 10.1158/1078-0432.CCR-14-0901 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Zhou XL, Yu LN, Wang Y, Tang LH, Peng YN, Cao JL, et al. Increased methylation of the MOR gene proximal promoter in primary sensory neurons plays a crucial role in the decreased analgesic effect of opioids in neuropathic pain. Molecular pain. 2014;10:51 10.1186/1744-8069-10-51 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Qi F, Zhou Y, Xiao Y, Tao J, Gu J, Jiang X, et al. Promoter demethylation of cystathionine-beta-synthetase gene contributes to inflammatory pain in rats. Pain. 2013;154(1):34–45. 10.1016/j.pain.2012.07.031 . [DOI] [PubMed] [Google Scholar]
- 18. Bell JT, Loomis AK, Butcher LM, Gao F, Zhang B, Hyde CL, et al. Differential methylation of the TRPA1 promoter in pain sensitivity. Nature communications. 2014;5:2978 10.1038/ncomms3978 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Doehring A, Oertel BG, Sittl R, Lotsch J. Chronic opioid use is associated with increased DNA methylation correlating with increased clinical pain. Pain. 2013;154(1):15–23. 10.1016/j.pain.2012.06.011 . [DOI] [PubMed] [Google Scholar]
- 20. Sun Y, Li XQ, Sahbaie P, Shi XY, Li WW, Liang DY, et al. miR-203 regulates nociceptive sensitization after incision by controlling phospholipase A2 activating protein expression. Anesthesiology. 2012;117(3):626–38. 10.1097/ALN.0b013e31826571aa [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Sun Y, Sahbaie P, Liang DY, Li WW, Li XQ, Shi XY, et al. Epigenetic regulation of spinal CXCR2 signaling in incisional hypersensitivity in mice. Anesthesiology. 2013;119(5):1198–208. 10.1097/ALN.0b013e31829ce340 . [DOI] [PubMed] [Google Scholar]
- 22. Momparler RL. Molecular, cellular and animal pharmacology of 5-aza-2'-deoxycytidine. Pharmacology & therapeutics. 1985;30(3):287–99. . [DOI] [PubMed] [Google Scholar]
- 23. Wilson VL, Jones PA, Momparler RL. Inhibition of DNA methylation in L1210 leukemic cells by 5-aza-2'-deoxycytidine as a possible mechanism of chemotherapeutic action. Cancer research. 1983;43(8):3493–6. . [PubMed] [Google Scholar]
- 24. Chaplan SR, Bach FW, Pogrel JW, Chung JM, Yaksh TL. Quantitative assessment of tactile allodynia in the rat paw. Journal of neuroscience methods. 1994;53(1):55–63. . [DOI] [PubMed] [Google Scholar]
- 25. Poree LR, Guo TZ, Kingery WS, Maze M. The analgesic potency of dexmedetomidine is enhanced after nerve injury: a possible role for peripheral alpha2-adrenoceptors. Anesthesia and analgesia. 1998;87(4):941–8. . [DOI] [PubMed] [Google Scholar]
- 26. Hargreaves K, Dubner R, Brown F, Flores C, Joris J. A new and sensitive method for measuring thermal nociception in cutaneous hyperalgesia. Pain. 1988;32(1):77–88. . [DOI] [PubMed] [Google Scholar]
- 27. Kingery WS, Davies MF, Clark JD. A substance P receptor (NK1) antagonist can reverse vascular and nociceptive abnormalities in a rat model of complex regional pain syndrome type II. Pain. 2003;104(1–2):75–84. . [DOI] [PubMed] [Google Scholar]
- 28. Jang JH, Liang D, Kido K, Sun Y, Clark DJ, Brennan TJ. Increased local concentration of complement C5a contributes to incisional pain in mice. Journal of neuroinflammation. 2011;8:80 10.1186/1742-2094-8-80 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Bestor TH. The DNA methyltransferases of mammals. Human molecular genetics. 2000;9(16):2395–402. . [DOI] [PubMed] [Google Scholar]
- 30. Okano M, Bell DW, Haber DA, Li E. DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell. 1999;99(3):247–57. . [DOI] [PubMed] [Google Scholar]
- 31. Mika J, Korostynski M, Kaminska D, Wawrzczak-Bargiela A, Osikowicz M, Makuch W, et al. Interleukin-1 alpha has antiallodynic and antihyperalgesic activities in a rat neuropathic pain model. Pain. 2008;138(3):587–97. 10.1016/j.pain.2008.02.015 . [DOI] [PubMed] [Google Scholar]
- 32. Vale ML, Marques JB, Moreira CA, Rocha FA, Ferreira SH, Poole S, et al. Antinociceptive effects of interleukin-4, -10, and -13 on the writhing response in mice and zymosan-induced knee joint incapacitation in rats. The Journal of pharmacology and experimental therapeutics. 2003;304(1):102–8. 10.1124/jpet.102.038703 . [DOI] [PubMed] [Google Scholar]
- 33. Milligan ED, Sloane EM, Langer SJ, Hughes TS, Jekich BM, Frank MG, et al. Repeated intrathecal injections of plasmid DNA encoding interleukin-10 produce prolonged reversal of neuropathic pain. Pain. 2006;126(1–3):294–308. 10.1016/j.pain.2006.07.009 . [DOI] [PubMed] [Google Scholar]
- 34. van Rietschoten JG, Verzijlbergen KF, Gringhuis SI, van der Pouw Kraan TC, Bayley JP, Wierenga EA, et al. Differentially methylated alleles in a distinct region of the human interleukin-1alpha promoter are associated with allele-specific expression of IL-1alpha in CD4+ T cells. Blood. 2006;108(7):2143–9. 10.1182/blood-2006-01-021147 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Mi XB, Zeng FQ. Hypomethylation of interleukin-4 and -6 promoters in T cells from systemic lupus erythematosus patients. Acta pharmacologica Sinica. 2008;29(1):105–12. 10.1111/j.1745-7254.2008.00739.x . [DOI] [PubMed] [Google Scholar]
- 36. Fu LH, Ma CL, Cong B, Li SJ, Chen HY, Zhang JG. Hypomethylation of proximal CpG motif of interleukin-10 promoter regulates its expression in human rheumatoid arthritis. Acta pharmacologica Sinica. 2011;32(11):1373–80. 10.1038/aps.2011.98 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Guan Y, Johanek LM, Hartke TV, Shim B, Tao YX, Ringkamp M, et al. Peripherally acting mu-opioid receptor agonist attenuates neuropathic pain in rats after L5 spinal nerve injury. Pain. 2008;138(2):318–29. 10.1016/j.pain.2008.01.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Kline RH, Wiley RG. Spinal mu-opioid receptor-expressing dorsal horn neurons: role in nociception and morphine antinociception. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2008;28(4):904–13. 10.1523/JNEUROSCI.4452-07.2008 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Wachman EM, Hayes MJ, Lester BM, Terrin N, Brown MS, Nielsen DA, et al. Epigenetic variation in the mu-opioid receptor gene in infants with neonatal abstinence syndrome. The Journal of pediatrics. 2014;165(3):472–8. 10.1016/j.jpeds.2014.05.040 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Han L, Witmer PD, Casey E, Valle D, Sukumar S. DNA methylation regulates MicroRNA expression. Cancer biology & therapy. 2007;6(8):1284–8. . [DOI] [PubMed] [Google Scholar]
- 41. Knight LJ, Burrage J, Bujac SR, Haggerty C, Graham A, Gibson NJ, et al. Epigenetic silencing of the endothelin-B receptor gene in non-small cell lung cancer. International journal of oncology. 2009;34(2):465–71. . [PubMed] [Google Scholar]
- 42. Viet CT, Ye Y, Dang D, Lam DK, Achdjian S, Zhang J, et al. Re-expression of the methylated EDNRB gene in oral squamous cell carcinoma attenuates cancer-induced pain. Pain. 2011;152(10):2323–32. 10.1016/j.pain.2011.06.025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Misawa K, Kanazawa T, Misawa Y, Uehara T, Imai A, Takahashi G, et al. Galanin has tumor suppressor activity and is frequently inactivated by aberrant promoter methylation in head and neck cancer. Translational oncology. 2013;6(3):338–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Xu X, Yang X, Zhang P, Chen X, Liu H, Li Z. Effects of exogenous galanin on neuropathic pain state and change of galanin and its receptors in DRG and SDH after sciatic nerve-pinch injury in rat. PloS one. 2012;7(5):e37621 10.1371/journal.pone.0037621 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Turcot V, Tchernof A, Deshaies Y, Perusse L, Belisle A, Marceau P, et al. Comparison of the dipeptidyl peptidase-4 gene methylation levels between severely obese subjects with and without the metabolic syndrome. Diabetology & metabolic syndrome. 2013;5(1):4 10.1186/1758-5996-5-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Grouzmann E, Monod M, Landis B, Wilk S, Brakch N, Nicoucar K, et al. Loss of dipeptidylpeptidase IV activity in chronic rhinosinusitis contributes to the neurogenic inflammation induced by substance P in the nasal mucosa. FASEB journal: official publication of the Federation of American Societies for Experimental Biology. 2002;16(9):1132–4. 10.1096/fj.01-0939fje . [DOI] [PubMed] [Google Scholar]
- 47. Sun Y, Sahbaie P, Liang D, Li W, Clark JD. opioids enhance CXCL1 expression and function after incision in mice. The journal of pain: official journal of the American Pain Society. 2014;15(8):856–66. 10.1016/j.jpain.2014.05.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Mauck M, Van de Ven T, Shaw AD. Epigenetics of chronic pain after thoracic surgery. Current opinion in anaesthesiology. 2014;27(1):1–5. 10.1097/ACO.0000000000000030 . [DOI] [PubMed] [Google Scholar]
- 49. Foulks JM, Parnell KM, Nix RN, Chau S, Swierczek K, Saunders M, et al. Epigenetic drug discovery: targeting DNA methyltransferases. Journal of biomolecular screening. 2012;17(1):2–17. 10.1177/1087057111421212 . [DOI] [PubMed] [Google Scholar]
- 50. Jin B, Robertson KD. DNA methyltransferases, DNA damage repair, and cancer. Advances in experimental medicine and biology. 2013;754:3–29. 10.1007/978-1-4419-9967-2_1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Tochiki KK, Cunningham J, Hunt SP, Geranton SM. The expression of spinal methyl-CpG-binding protein 2, DNA methyltransferases and histone deacetylases is modulated in persistent pain states. Molecular pain. 2012;8:14 10.1186/1744-8069-8-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Pollema-Mays SL, Centeno MV, Apkarian AV, Martina M. Expression of DNA methyltransferases in adult dorsal root ganglia is cell-type specific and up regulated in a rodent model of neuropathic pain. Frontiers in cellular neuroscience. 2014;8:217 10.3389/fncel.2014.00217 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Baubec T, Colombo DF, Wirbelauer C, Schmidt J, Burger L, Krebs AR, et al. Genomic profiling of DNA methyltransferases reveals a role for DNMT3B in genic methylation. Nature. 2015. 10.1038/nature14176 . [DOI] [PubMed] [Google Scholar]
- 54. Wang L, Wang J, Sun S, Rodriguez M, Yue P, Jang SJ, et al. A novel DNMT3B subfamily, DeltaDNMT3B, is the predominant form of DNMT3B in non-small cell lung cancer. International journal of oncology. 2006;29(1):201–7. . [PubMed] [Google Scholar]
- 55. Gopalakrishnan S, Van Emburgh BO, Shan J, Su Z, Fields CR, Vieweg J, et al. A novel DNMT3B splice variant expressed in tumor and pluripotent cells modulates genomic DNA methylation patterns and displays altered DNA binding. Molecular cancer research: MCR. 2009;7(10):1622–34. 10.1158/1541-7786.MCR-09-0018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Ji W, Hernandez R, Zhang XY, Qu GZ, Frady A, Varela M, et al. DNA demethylation and pericentromeric rearrangements of chromosome 1. Mutation research. 1997;379(1):33–41. . [DOI] [PubMed] [Google Scholar]
- 57. Chen RZ, Pettersson U, Beard C, Jackson-Grusby L, Jaenisch R. DNA hypomethylation leads to elevated mutation rates. Nature. 1998;395(6697):89–93. 10.1038/25779 . [DOI] [PubMed] [Google Scholar]
- 58. Tajerian M, Alvarado S, Millecamps M, Vachon P, Crosby C, Bushnell MC, et al. Peripheral nerve injury is associated with chronic, reversible changes in global DNA methylation in the mouse prefrontal cortex. PloS one. 2013;8(1):e55259 10.1371/journal.pone.0055259 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Meller R, Pearson A, Simon RP. Dynamic changes in DNA methylation in ischemic tolerance. Frontiers in neurology. 2015;6:102 10.3389/fneur.2015.00102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Kraus J, Lehmann L, Borner C, Hollt V. Epigenetic mechanisms involved in the induction of the mu opioid receptor gene in Jurkat T cells in response to interleukin-4. Molecular immunology. 2010;48(1–3):257–63. 10.1016/j.molimm.2010.08.002 . [DOI] [PubMed] [Google Scholar]
- 61. Wood EJ, Lipovich L. MicroRNAs in opioid addiction: elucidating evolution. Frontiers in genetics. 2012;3:241 10.3389/fgene.2012.00241 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Kohno T, Ji RR, Ito N, Allchorne AJ, Befort K, Karchewski LA, et al. Peripheral axonal injury results in reduced mu opioid receptor pre- and post-synaptic action in the spinal cord. Pain. 2005;117(1–2):77–87. 10.1016/j.pain.2005.05.035 . [DOI] [PubMed] [Google Scholar]
- 63. Truong W, Cheng C, Xu QG, Li XQ, Zochodne DW. Mu opioid receptors and analgesia at the site of a peripheral nerve injury. Annals of neurology. 2003;53(3):366–75. 10.1002/ana.10465 . [DOI] [PubMed] [Google Scholar]
- 64. Hwang CK, Kim CS, Kim do K, Law PY, Wei LN, Loh HH. Up-regulation of the mu-opioid receptor gene is mediated through chromatin remodeling and transcriptional factors in differentiated neuronal cells. Molecular pharmacology. 2010;78(1):58–68. 10.1124/mol.110.064311 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Corder G, Doolen S, Donahue RR, Winter MK, Jutras BL, He Y, et al. Constitutive mu-opioid receptor activity leads to long-term endogenous analgesia and dependence. Science. 2013;341(6152):1394–9. 10.1126/science.1239403 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Labuz D, Mousa SA, Schafer M, Stein C, Machelska H. Relative contribution of peripheral versus central opioid receptors to antinociception. Brain research. 2007;1160:30–8. 10.1016/j.brainres.2007.05.049 . [DOI] [PubMed] [Google Scholar]
- 67. Schmidt Y, Gaveriaux-Ruff C, Machelska H. mu-Opioid receptor antibody reveals tissue-dependent specific staining and increased neuronal mu-receptor immunoreactivity at the injured nerve trunk in mice. PloS one. 2013;8(11):e79099 10.1371/journal.pone.0079099 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Bigliardi-Qi M, Sumanovski LT, Buchner S, Rufli T, Bigliardi PL. Mu-opiate receptor and Beta-endorphin expression in nerve endings and keratinocytes in human skin. Dermatology. 2004;209(3):183–9. 10.1159/000079887 . [DOI] [PubMed] [Google Scholar]
- 69. Chuang TK, Killam KF Jr, Chuang LF, Kung HF, Sheng WS, Chao CC, et al. Mu opioid receptor gene expression in immune cells. Biochemical and biophysical research communications. 1995;216(3):922–30. 10.1006/bbrc.1995.2709 . [DOI] [PubMed] [Google Scholar]
- 70. Sahbaie P, Li X, Shi X, Clark JD. Roles of Gr-1+ leukocytes in postincisional nociceptive sensitization and inflammation. Anesthesiology. 2012;117(3):602–12. 10.1097/ALN.0b013e3182655f9f [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Spofford CM, Brennan TJ. Gene expression in skin, muscle, and dorsal root ganglion after plantar incision in the rat. Anesthesiology. 2012;117(1):161–72. 10.1097/ALN.0b013e31825a2a2b [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Nesic O, Lee J, Johnson KM, Ye Z, Xu GY, Unabia GC, et al. Transcriptional profiling of spinal cord injury-induced central neuropathic pain. Journal of neurochemistry. 2005;95(4):998–1014. 10.1111/j.1471-4159.2005.03462.x . [DOI] [PubMed] [Google Scholar]
- 73. Jones PL, Wolffe AP. Relationships between chromatin organization and DNA methylation in determining gene expression. Seminars in cancer biology. 1999;9(5):339–47. 10.1006/scbi.1999.0134 . [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the paper.