

Leaf senescence varies widely across species and sites

Leaf senescence marks the end of the temperate growing season, but climate change is altering leaf senescence phenology. Panchen et al. (pp. 865–873) record the leaf senescence phenology of 1360 deciduous species at six temperate botanical gardens in two consecutive years, and find that species tend to undergo leaf senescence in the same order at each garden in both years, but across gardens the senescence order is weakly correlated. Leaf senescence times are only minimally influenced by spring leaf out times, growth habit and phylogenetics, and show weaker patterns than leaf out times. Thus, in contrast to the broader temperature effects that determine leaf out times, leaf senescence times are probably determined by a larger or different suite of local environmental effects. Interspecific differences in leaf senescence responses to climate change could affect growing season length, micro-climate and community composition.
Meta-analysis of autumn senescence in the northern hemisphere

Past studies show that rising temperatures influence leaf senescence timing, but there is limited consensus on the magnitude, direction and spatial extent of the relationship. Gill et al. (pp. 875–888) conduct a meta-analysis of studies that report the timing of autumn leaf senescence in the northern hemisphere and show that increased temperatures are associated with larger senescence delays at low latitudes than at high latitudes. Senescence at high-latitude sites is more sensitive to photoperiod and at low-latitude sites it is more sensitive to temperature. Differences in the drivers of senescence between high- and low-latitude ecosystems have implications for both plant community interactions and ecosystem carbon storage.
Using dormant twigs to investigate climate change (Viewpoint)

Climate change has altered the spring phenology of many temperate plants, with important ecological consequences. However, studies of phenological shifts are generally restricted to field observations or remote-sensing methods. Twig experiments—which involve clipping dormant twigs of trees, shrubs and vines and bringing them into laboratory conditions for phenological observations—offer an underutilized opportunity to disentangle the drivers of plant phenology. Primack et al. (pp. 889–897) explore the power of twig experiments examining leaf-out, frost sensitivity, flowering and leaf senescence for use in ecology, climate change biology, horticulture and public health, and consider the challenges unique to this method.
Global warming may reduce plant nectar yield

Although flower nectar yield is well known to be temperature dependent, the effects of climate change on the availability of nectar to pollinators have rarely been studied. Mu et al. (pp. 899–906) apply long-term artificial asymmetric warming to a perennial herb, Saussurea nigrescens, in a Tibetan alpine meadow and find that winter warming weakens vernalization and decreases floret and capitulum number, and that night-time warming reduces floret nectar volume. The overall effect is a reduction in nectar production per unit area of about 90 %, and the results suggest that decreases in floral quantity and quality as a result of global warming may have increasingly negative effects on plant–pollinator interactions.
Climate warming and recruitment success in glacier forelands

Glacier foreland plants are highly threatened by global warming, and regeneration from seeds on deglaciated terrain will be crucial for successful migration and survival of these species. Mondoni et al. (pp. 907–916) sow seeds of eight foreland species at 2500 m altitude and at a site 400 m lower to simulate a +2·7 °C increase in mean annual temperature, and find that at the warmer site there is a shift from summer to autumn emergence in two species and a significant increase of summer emergence in all species except two. They conclude that warming will influence recruitment primarily via the extension of the snow-free period in spring, increasing seedling emergence and survival. The changes in recruitment success that are observed imply that range shifts or changes in abundance are possible in a future warmer climate.
Cold-induced forest dieback of rear-edge tree populations

The impact of extreme cold events on forest dynamics, especially at the low-latitude margin (‘rear edge’) of species distributional ranges, has received little attention. Camarero et al. (pp. 917–927) quantify how an extreme cold event in 2001 in Spain affected growth, needle loss and mortality of Scots pine (Pinus sylvestris) and compare it with the predictions of a species distribution model (SDM). For trees already stressed by a preceding dry summer and autumn, the record minimum temperatures caused dieback and large-scale mortality. The SDM, which predicts distribution mainly on the basis of responses to maximum and minimum monthly temperatures, failed to model this, leading to the conclusion that distribution modelling techniques such as SDM must incorporate climatic extremes in order to avoid biased predictions based solely on warmer climatic scenarios.
Temperature responses of photosynthesis under elevated CO2

Plants growing under elevated atmospheric CO2 concentrations often have reduced stomatal conductance and subsequently increased leaf temperature. Šigut et al. (pp. 929–939) find that this is indeed the case for broadleaved Fagus sylvatica and coniferous Picea abies saplings growing under elevated CO2, but that the higher leaf temperatures do not lead to an increased heat stress tolerance of primary photochemical reactions. The increases in temperature optima of photosynthetic CO2 uptake observed under elevated CO2 are instantaneous and are caused by reduced photorespiration and limitation of photosynthesis by RuBP regeneration. However, the increases disappear when plants are exposed to identical CO2 concentrations. The results therefore do not support the hypothesis that elevated CO2 leads to temperature acclimation of photosynthesis in the species studied.
Climate change and the decline of a foundation tree species
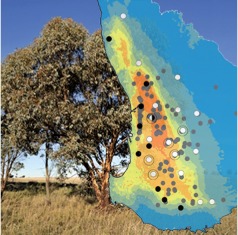
A worldwide increase in tree decline and mortality has been linked to climate change, and where these represent foundation species this can have important implications for ecosystem functions. Dalmaris et al. (pp. 941–952) undertake phylogeographic analyses across the species range of Eucalyptus wandoo, an endemic tree of south-western Australia, and find that a combination of phylogeography and paleo-distribution modelling can provide an evolutionary context for climate-driven tree decline, as both can be used to cross-validate evidence for refugia and contraction under harsh climatic conditions. The contraction of E. wandoo from lower rainfall areas is consistent with current observations of decline on the semi-arid margin of the range, and indicates low capacity to tolerate forecast climatic change.
Climate change and flowering phenology in mid- and high-elevation species

Climate change effects on flowering phenology in alpine environments with short growing seasons and cold temperatures are likely to make reproduction particularly challenging.
Gugger et al. (pp. 953–962) reciprocally transplant congeneric species pairs originating from mid and high elevations to common gardens at altitudes that mimic prospective climates and find considerable shifts in reproductive phenology, with highly advanced initiation and shortened phenophases at the lower, warmer site for both mid- and high-elevation species. Drought stress amplifies these responses. They conclude that while mid- and high-elevation species seem to adequately shift their reproductive phenology to track ongoing climate changes, those from high altitudes are less capable of doing so and are probably constrained by their specific adaptation to a cold environment with short growing seasons.
Native plant responses to enhanced rainfall

The environmental and biotic contexts within which plants grow have a great potential to modify responses to climatic changes. Eskelinen and Harrison (pp. 963–973) investigate how the effects of enhanced rainfall are modulated by above-ground competition and by edaphic variables for two grassland plants, Lasthenia californica and Calycadenia pauciflora. They find that increased competition outweighs the direct positive impacts of enhanced rainfall on most fitness measures for both species, resulting in no net effect of enhanced rainfall. Both species benefit from enhanced rainfall when the absence of competitors is accompanied by high soil water retention capacity. They conclude that the results emphasize the importance of considering abiotic as well as biotic context when making future climate change forecasts.
Phosphorus and elevated CO2 enhance drought tolerance
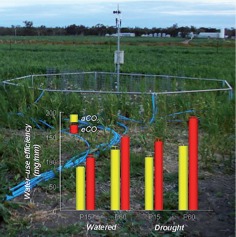
Benefits to crop productivity arising from elevated atmospheric CO2 concentration may be offset by nutrient deficiencies and detrimental effects of increasing frequency of drought under global climate change. Jin et al. (pp. 975–985) use a free-air CO2 enrichment system to study the effects of phosphorus (P) supply and elevated CO2 on responses to drought stress of field pea (Pisum sativum) grown in P-deficient vertisol. They find that P application and elevated CO2 interactively enhance water-use efficiency and improve tolerance to periodic drought as a result of decreased stomatal conductance, deep rooting and greater availability of inorganic P for carbon assimilation in leaves. An adequate supply of P may therefore help crops better withstand drought under future climate scenarios.
Plant phosphorus nutrition under elevated CO2 (Review)
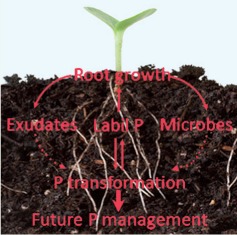
Significant increases in P demand by plants are likely to happen under elevated CO2 due to the stimulation of photosynthesis and subsequent growth responses. Jin et al.. (pp. 987–999) review possible mechanisms by which elevated CO2 might affect plant P acquisition and conclude that there will be increasing selection pressure for P-acquisition efficiency in a high-carbon environment by plants and plant–microbe associations. Plants will utilize and exploit the increased carbon flow to their roots to more efficiently mobilize and/or forage for labile P forms in the soil. The mechanisms for this selection might include the development of longer roots, more lateral roots and root hairs, changes in the quantity and composition of root exudates, and changes in the activities and/or functions of microbes and plant–microbe associations.
Drought strategies in perennial forage species

Severe droughts are expected to increase with climate change and to limit grassland perennity. Zwicke et al. (pp. 1001–1015) investigate the plant strategies involved in drought survival and recovery of six temperate herbaceous species in semi-controlled conditions and show that cell membrane stability, which ensures dehydration tolerance of roots and meristems, is positively and negatively correlated to fructan and sucrose contents, respectively. Species that survive and recover best combine high resource acquisition in spring and both high dehydration avoidance and tolerance strategies. The results suggest that most of the native forage species studied are able to survive and recover from extreme drought, but with various time lags, which emphasizes the need to assess recovery after drought over a relatively long time scale.
Communities on infertile soils are less sensitive to climate change

Much evidence suggests that plant communities on infertile soils are relatively insensitive to increased water deficit caused by increasing temperature and/or decreasing precipitation. Harrison et al. (pp. 1017–1022) undertake a re-analysis of a large, multi-decade study that examined forest understories on infertile serpentine and fertile soils in an aridifying climate in southern Oregon from 1949–51 to 2007–08. They conclude that the balance of evidence supports a hypothesis that links soil infertility to a stress-tolerant functional trait syndrome that tends to confer unusually high resistance of plant species and communities to climate change. This is qualified, however, by noting that the evidence only applies to communities that are becoming effectively drier because of declining precipitation and/or increasing temperatures leading to greater water deficits.
Snow cover and predicting alpine plant community diversity

Quantifying relationships between snow cover duration and plant community properties remains an important challenge in alpine ecology, especially in the context of climate change. Carlson et al. (pp. 1023–1034) utilize Landsat imagery to model snow cover dynamics and to estimate energy availability over the course of five growing seasons in a high-elevation watershed in the French Alps, and compare the explanatory power of environmental variables estimated with and without snow cover. They find that predictions of spatial variability in the functional and taxonomic diversity of plant communities improve when high-resolution data on snow cover duration are taken into account, and they conclude that studies seeking to predict the response of alpine plant communities to climate change need to consider shifts in both temperature and nival regimes.