

Graphical abstract
Keywords: Vitamin D, Deficiency, Dermatology, Immunological
Abstract
The “sunshine” vitamin is a hot topic that attracted ample attention over the past decades, specially that a considerable proportion of the worldwide population are deficient in this essential nutrient. Vitamin D was primarily acknowledged for its importance in bone formation, however; increasing evidence point to its interference with the proper function of nearly every tissue in our bodies including brain, heart, muscles, immune system and skin. Thereby its deficiency has been incriminated in a long panel of diseases including cancers, autoimmune diseases, cardiovascular and neurological disorders. Its involvement in the pathogenesis of different dermatological diseases is no exception and has been the subject of much research over the recent years. In the current review, we will throw light on this highly disputed vitamin that is creating a significant concern from a dermatological perspective. Furthermore, the consequences of its deficiency on the skin will be in focus.
Introduction
It is somewhat ironic that vitamin D, through a historical accident, became classified as a ‘vitamin’, owing to the fact that vitamin is conventionally defined as ‘essential item needed in the diet’. The paradox with ‘vitamin D’ is that diet per se is usually poor in vitamin D except for cod or other fish oils or food fortified with this vitamin [1].
Vitamin D is actually a fat-soluble prohormone steroid that has endocrine, paracrine and autocrine functions [2]. The endocrine effects of vitamin D are mainly involved in serum calcium homeostasis. Vitamin D and calcium are often used in the same sentence because they work closely together, vitamin D’s primary role is to control the levels of calcium found in the bloodstream by constantly allowing calcium and phosphate absorption from the intestine or taking calcium from bones. Furthermore, vitamin D is an enabling agent that, when present in optimal concentrations, has no perceptible effect on calcium absorption in its own right; however, it permits or facilitates flexible physiologic response to varying calcium need [3].
The paracrine and autocrine effects of vitamin D depend on genetic transcription, unique to the type of cell expressing nuclear vitamin D receptors. These potential effects include inhibition of cell proliferation, promotion of cell differentiation, and apoptosis which may in turn have roles in cancer, immunity, and many organ systems [4], [5], [6], [7], [8]. The potential myriad effects of this vitamin in human health and disease have led to an escalating interest in vitamin D inadequacy and the best methods to normalize suboptimal levels.
Sources of vitamin D
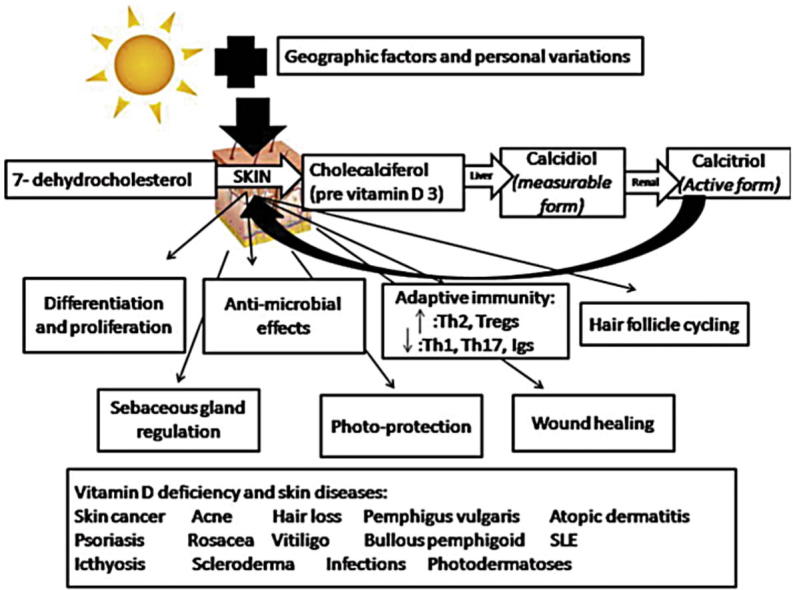
There are only 3 known sources of vitamin D; sunlight, diet, and vitamin D supplements (Fig. 1) [2], [9], [10].
Fig. 1.

A diagram illustrating the different sources and forms of vitamin D.
Sunlight
The most well-known source of vitamin D is via synthesis in the skin induced by sun exposure. The first reference to the physiological effect of sunlight on vitamin D was illustrated by the Greek historian Herodotus. He visited the battlefield where Cambyses (525 BC) overcame the Egyptians, and inspected the skulls of slain Persians and Egyptians. He noted that the Persian skulls were so fragile that they broke even when struck with a pebble, whereas those of the Egyptians were strong and could scarcely be broken even when struck with a stone. The Egyptians’ explanation to Herodotus was that they went bareheaded from childhood exposing their heads to sunlight, whereas Persians covered their heads with turbans shading them from the sun resulting in skull bone weakness. Later on, in the mid 17th century Francis Glisson, Professor of Physics at Cambridge University, in his treatise on rickets observed that the disease was common among infants and young children of country farmers who ate well, and whose diets were known to include eggs and butter, but who lived in rainy, misty parts of the country and who were kept indoors during long severe winters [11].
Vitamin D synthesis in the skin
According to the Commission Internationale de l’Eclairage (CIE) [12], the vitamin D effective radiation is described in terms of its action spectrum (i.e., the efficiency of each wavelength to synthesize vitamin D in skin) which covers the spectral range (255–330 nm) with a maximum at about 295 nm (UVB). A whole body exposure to UVB radiation inducing the light pink color of the minimal erythema dose for 15–20 min is able to induce the production of up to 250 μg vitamin D (10,000 IU) [13], [14].
Its precursor 7-dehydrocholesterol in the plasma membranes of both epidermal basal and suprabasal keratinocytes and dermal fibroblasts is converted to previtamin D3. Cutaneously synthesized vitamin D3 is released from the plasma membrane and enters the systemic circulation bound to vitamin D-binding protein (DBP) [15]. Serum concentrations of vitamin D3 peak 24–48 h following exposure to UV radiation [13]. Thereafter, vitamin D3 levels decline exponentially with a serum half-life ranging from 36 to 78 h [13], [14]. As a lipid-soluble molecule, vitamin D3 can be taken up by adipocytes and stored in subcutaneous or omental fat for later use [16]. The distribution of vitamin D3 into adipose tissue prolongs its total-body half-life to approximately two months as first detected on experiments on submarine personnel [17], [18], [19].
Once in the circulation, vitamin D is converted by a hepatic hydroxylase into 25-hydroxyvitamin D (25(OH)D; calcidiol). The circulating 25(OH)D level is an indicator of the vitamin D status. This level reflects both ultraviolet exposure and dietary vitamin D intake. The serum half-life of 25(OH)D is approximately 15 days [2]. 25(OH)D is not biologically active except at very high, non-physiological levels [20]. As needed, 25(OH)D is converted in the kidney to its active hormonal form 1,25-dihydroxyvitamin D (1,25(OH)2D; calcitriol) in a process which is usually tightly controlled by the parathyroid hormone which levels start rising at 25(OH)D cutoff levels of 75 nmol/L or lower. In spite of this, inadequate vitamin D supply lowers the circulating level of calcitriol [16]. Circulating calcitriol is also adversely affected by a reduced number of viable nephrons, high serum concentrations of fibroblast growth factor-23, and high levels of inflammatory cytokines such as interleukin (IL)-1, IL-6, and tumor necrosis factor-alpha (TNF-α) [19], [21].
It is important to know that the conversion of previtamin D3 to the inactive photoproducts lumisterol and tachysterol balances the cutaneous biosynthesis of vitamin D3 as a feedback loop. This mechanism ensures that one cannot “overdose” on vitamin D3 by photoexposure alone. After less than 1 minimal erythema dose (MED; i.e., the amount of photoexposure required to produce faint pinkness in the skin at 24 h after exposure), the concentration of previtamin D3 reaches maximal levels and further UV radiation merely results in the production of inactive metabolites [2].
Dietary sources and supplements
Vitamin D is available in 2 distinct forms, ergocalciferol (vitamin D2) and cholecalciferol (vitamin D3). Sunshine exposure provides vitamin D in the form of D3 only, while dietary sources are able to provide both forms, which are officially regarded by many as equivalent and interchangeable [22], [23], [24]. However, several reasons have been suggested to argue against this presumption including that both are different in their efficacy at raising serum 25-hydroxyvitamin D, with diminished binding of vitamin D2 metabolites to vitamin D binding protein in plasma, as well as the detection of a nonphysiologic metabolism and shorter shelf life for vitamin D2. Nevertheless, still to this day, the major preparations of vitamin D for prescription are in the form of vitamin D2, not vitamin D3. Multivitamins may contain either vitamin D2 or vitamin D3, but most companies are now reformulating their products to contain vitamin D in the D3 form [25].
There are only few natural sources of vitamin D including cod liver oil, cheese, egg yolks, mackerel, salmon, tuna fish, and beef liver. Because it is not easy for many individuals to obtain adequate vitamin D intake from natural dietary sources alone, many countries fortify foods such as orange juice, milk, yogurt, and cereal with vitamin D. Many inexpensive supplemental vitamin D forms are readily available over the counter in both vitamin D3 and vitamin D2 forms and with or without calcium [26], [27].
Vitamin D levels
Different cut-off values for the normal threshold of vitamin D have been used until recently [28]. A level of 50 nmol/L has been widely used to define 25(OH)D insufficiency, while some studies have used 37.5 nmol/L as the lowest level of sufficiency [29], [30], [31]. Further studies, however, suggest that a 25-(OH)D level as high as 75 nmol/L or higher is needed to cover all physiological functions of vitamin D and should therefore be considered optimal [32], [33], [34], [35], [36].
Factors influencing vitamin D levels
Nutrient deficiencies are usually the result of dietary inadequacy, impaired absorption and use, increased requirement, or increased excretion. Vitamin D deficiency can occur when usual intake is lower than recommended levels over time, exposure to sunlight is limited, the kidneys cannot convert 25(OH)D to its active form, or absorption of vitamin D from the digestive tract is inadequate. Vitamin D-deficient diets are associated with milk allergy, lactose intolerance, ovo-vegetarianism, and veganism [37].
Regarding the amount of vitamin D production in human skin, it depends on several variables including environmental factors such as geographic latitude, season, time of day, weather conditions (cloudiness), amount of air pollution and surface reflection which can all interfere with the amount of UVB radiation reaching the skin [38], [39], [40], [41].
Personal variations represent another group of influential factors affecting the vitamin D production in the skin, including age as elderly people have thinner skin, and consequently are less capable of synthesizing vitamin D [7], [38], [39] and obesity as overweight individuals have reduced vitamin D levels [42]. It is also noteworthy that skin type determines a person’s effectiveness in producing vitamin D. Light skins (type I) produce up to six fold the amount of vitamin D produced by dark skins (type VI). In addition, clothing habits, lifestyle, workplace (e.g., indoor versus outdoor), and sun avoidance practices have a strong impact on vitamin D synthesis [38], [39], [40], [41].
The influence of some common practices as using sunblocks or receiving sunbeds on vitamin D production is another point of interest. Sunblocks are known to block UVB radiation effectively. However, it is questionable whether sunscreen in practice causes any vitamin D deficiency. Absolute full-body coverage of sunscreen is uncommon. Some areas of the skin are always left out. At times and locations where the sun is intense and the temperature is high enough to make the population use sunscreen, its vitamin D status is generally very satisfactory [39], [40], [41]. On the other hand the use of sun beds is controversial, but regardless, subjects who regularly use tanning beds that emit UVB radiation are likely to have higher 25(OH)D concentrations. Nevertheless, there is a trend toward discouraging the use of such tanning beds for fear of melanoma and non-melanoma skin cancer [43].
Vitamin D and the skin: What’s beyond its synthesis and metabolism?
The skin is unique in being not only the source of vitamin D for the body but also in being capable of responding to the active metabolite of vitamin D, 1,25(OH)2D. Both 1,25(OH)2D and its receptor (VDR) play essential roles in the skin.
Skin differentiation and proliferation
Both calcium and 1,25(OH)2D perform important and interacting functions in regulating the skin differentiation process. 1,25(OH)2D increases the expression of involucrin, transglutaminase, loricrin, and filaggrin and increases keratinocyte cornified envelope formation while inhibiting proliferation [44], [45]. These actions are due to, at least in part, the ability of 1,25(OH)2D to increase intracellular calcium levels achieved by induction of the calcium receptor [46], and the phospholipase C [47] that are critical for the ability of calcium to stimulate keratinocyte differentiation [48], [49]. Mice lacking the VDR show defective epidermal differentiation manifesting as reduced levels of involucrin and loricrin and loss of keratohyaline granules [50], [51].
Cutaneous antimicrobial effects
1,25(OH)2D and its receptor regulate the processing of the long chain glycosylceramides that are critical for the skin barrier formation [52] which is crucial in defending the skin. Furthermore, they induce toll like receptor 2 (TLR2) and its coreceptor CD14, that initiate the innate immune response in skin [53]. Activation of these receptors leads to the induction of CYP27B1, which in turn induces cathelicidin resulting in the killing of invasive organisms [53], [54]. Mice lacking the VDR or the enzyme (CYP27B1) show decreased lipid content of the lamellar bodies leading to a defective permeability barrier [52], and a defective response of the innate immune system to invading infections [53].
Vitamin D and cutaneous innate immunity
The historical link between vitamin D and innate immune function stemmed initially from the use of cod liver oil as treatment for tuberculosis (TB) [54]. More recent work has focused on the cellular and molecular machinery that underpins the actions of vitamin D on the pathogen that causes TB, Mycobacterium tuberculosis (M. TB). In the first of these studies, carried out 25 years ago, active 1,25(OH)2D was shown to reduce the proliferation of M. TB in macrophages with this effect being enhanced by the cytokine interferon γ (IFNγ), a known stimulator of macrophages [55]. However, the major advance in our understanding of how vitamin D directs antibacterial responses in TB arose from much more recent studies aiming at defining the way by which monocytes and macrophages, key cells in directing bacterial killing, respond to an encounter with M. TB [56]. These data suggested that monocytes promote localized activation of vitamin D in response to M. TB, with the resulting 1,25(OH)2D binding to endogenous VDR. In this way, vitamin D can act to modulate gene expression in response to M. TB immune challenge – a classical intracrine mechanism [57], [58]. Functional analyses showed that 25OHD-mediated induction of cathelicidin is coincident with enhanced killing of M. TB in monocytes. Naturally occurring variations in serum 25OHD have been shown to correlate with induction of monocyte cathelicidin expression [59]. The conclusion from these studies was that individuals with low serum 25OHD will be less able to support monocyte induction of antibacterial activity and may therefore be at greater risk of infection. Conversely, supplementation of vitamin D-insufficient individuals in vivo has been shown to improve TLR-mediated induction of monocyte cathelicidin [60] and may therefore help to protect against infection (Fig. 2).
Fig. 2.

A diagram illustrating the influences of vitamin D on the cutaneous innate and adaptive immunity.
Studies have shown that T-cell cytokines play a pivotal role in both amplifying and attenuating vitamin D-mediated cathelicidin production [61]. Indeed, cytokine production by monocytes themselves may be central to the intracrine metabolism of vitamin D in this cell type [62], [63]. Thus, it seems likely that the ability to mount an appropriate response to infection will be highly dependent on the availability of vitamin D, with additional tuning of this response by other components of the normal human immune response.
Vitamin D can also influence innate immune responses to pathogens via effects on antigen presentation by macrophages or dendritic cells (DCs) (Fig. 2). These cells are known to express VDR [64], and treatment with 1,25(OH)2D has been shown to inhibit DC maturation, suppress antigen presentation and promote a tolerogenic T-cell response [65], [66].
Vitamin D and cutaneous adaptive immunity
Early studies of vitamin D and the immune system demonstrated VDR expression in both T and B cells (Fig. 2) [67]. Notably, VDR expression by these cells was only immunologically functional in active, proliferating cells, suggesting an antiproliferative role for 1,25(OH)2D on these cells [68]. T helper (Th) cells appear to be the principal target for 1,25(OH)2D which can suppress Th cell proliferation as well as modulating cytokines production by these cells [69]. Activation of naive Th cells by antigen in turn leads to the generation of Th cell subgroups with distinct cytokine profiles: Th1 (IL-2, IFN γ, tumor necrosis factor alpha) and Th2 (IL-3, IL-4, IL-5, IL-10) that respectively support cell-mediated and humoral immunity [70], [71].
In vitro 1,25(OH)2D inhibits Th1 cytokines [72], while promoting Th2 cytokines [73]. A third group of Th cells known to be influenced by vitamin D are interleukin-17 (IL-17)-secreting T cells (Th17 cells). Autoimmune disease-susceptible non obese diabetic (NOD) mice treated with 1,25D exhibit lower levels of IL-17 [74], and 1,25(OH)2D-mediated suppression of murine retinal autoimmunity appears to involve inhibition of Th17 activity [75]. Furthermore, subsequent studies have shown that 1,25(OH)2D suppresses IL-17 production via direct transcriptional suppression of IL-17 gene expression [76].
Another group of T cells known to be potently induced by 1,25(OH)2D are regulatory T cells (Tregs) [77]. Although part of the Th cell family, Tregs act to suppress immune responses by other T cells as part of the machinery to prevent over-exuberant or autoimmune responses [78]. Recent studies have underlined the importance of Tregs in mediating the immunoregulatory actions of vitamin D. Administration of 1,25(OH)2D systemically to patients who underwent renal transplantation has been shown to expand circulating Treg populations [79].
Studies of vitamin D and T-cell function have to date focused primarily on the response of these cells to active 1,25(OH)2D. What is less clear is the mechanism by which variations in vitamin D status can also influence T cells, despite reports linking serum levels of 25OHD with specific T-cell populations [56]. For example, circulating levels of 25OHD have been shown to correlate with Tregs activity in patients with multiple sclerosis [80], [81]. There are four potential mechanisms by which serum 25OHD is believed to influence T-cell function; (i) direct effects on T cells mediated via systemic 1,25(OH)2D; (ii) indirect effects on antigen presentation to T cells mediated via localized DC expression of CYP27B1 and intracrine synthesis of 1,25(OH)2D; (iii) direct effects of 1,25(OH)2D on T cells following synthesis of the active form of vitamin D by CYP27B1-expressing monocytes or DCs – a paracrine mechanism; (iv) Intracrine conversion of 25OHD to 1,25(OH)2D by T cells. As yet, it is unclear whether one or more of these mechanisms will apply to the regulation of specific T-cell types. For example, the effects of 1,25(OH)2D on Tregs can occur indirectly via effects on DCs [82], but may also involve direct effects on the Tregs [83]. However, as DCs also express CYP27B1 [84] and may therefore act as the conduit for 25OHD effects on Tregs. Interestingly, reports have also described expression of CYP27B1 by T cells [85], suggesting that 25OHD may also influence the function of these cells via an intracrine mechanism, although the precise relevance of this to specific T-cell types remains unclear [56].
Despite the fact that expression of VDR by B cells has been recognized for many years [67], the ability of 1,25(OH)2D to suppress B-cell proliferation and immunoglobulin (Ig) production was initially considered to be an indirect effect mediated via Th cells [68]. However, more recent studies have confirmed direct effects of1,25(OH)2D on B-cell homoeostasis [86], with notable effects including inhibition of plasma cells and class switched memory cells differentiation. These effects lend further support for vitamin D’s proposed role in B-cell-related autoimmune disorders such as systemic lupus erythematosis. Other B-cell targets known to be modulated by for 1,25(OH)2D include IL-10 [87] and CCR10 [88], suggesting that the repertoire of B-cell responses to vitamin D extends beyond its effects on B-cell proliferation and Ig synthesis [56].
Hair follicle cycling
In vitro studies have supported the concept that VDR may play a vital role in the postnatal maintenance of the hair follicle. Mesodermal papilla cells and the outer root sheath (ORS) epidermal keratinocytes express VDR in varied degrees in correlation with the stages of the hair cycle. In both the late anagen and catagen stages there is an increase in VDR, which is associated with decreased proliferation and increased differentiation of the keratinocytes. These changes are thought to promote the progression of the hair cycle [89].
Limited studies have been done in humans to elaborate the role of vitamin D in the hair cycle. A potential application for vitamin D is in chemotherapy-induced alopecia. Topical calcitriol has been shown to protect against chemotherapy-induced alopecia caused by paclitaxel and cyclophosphamide. However, topical calcitriol failed to protect against chemotherapy-induced alopecia caused by a combination of 5-fluorouracil, doxorubicin, and cyclophosphamide and a combination of cyclophosphamide, methotrexate, and 5-fluorouracil. The ability of topical calcitriol to prevent chemotherapy-induced alopecia may therefore depend on the chemotherapy agents used. Of note, the studies in which no effects were observed, were small and may have used doses of vitamin D that were inadequate to protect against chemotherapy-induced alopecia [90].
The sebaceous gland
It has been reported that incubation of the human sebaceous gland cell line with 1,25OH2D results in a dose-dependent suppression of cell proliferation. Using real-time PCR, it was demonstrated that key components of the vitamin D system (VDR, 25OHase, 1αOHase, and 24OHase) are strongly expressed in such cells. It has been concluded that local synthesis or metabolism of vitamin D metabolites may be of importance for growth regulation and various other cellular functions in sebaceous glands and that sebaceous glands represent promising targets for therapy with vitamin D analogs or for pharmacological modulation of calcitriol synthesis/metabolism [91], [92].
Photoprotection
Photodamage refers to skin damage induced by ultraviolet (UV) light. Depending on the dose, UV light can lead to DNA damage, inflammatory responses, skin cell apoptosis (programmed cell death), skin aging, and skin cancer. Some studies, mainly in vitro (cell culture) studies [93], [94], [95], [96] and mouse studies where 1,25-dihydroxyvitamin D3 was topically applied to skin before or immediately following irradiation [93], [97], [98], have found that vitamin D exhibits photoprotective effects. Documented effects on skin cells include decreased DNA damage, reduced apoptosis, increased cell survival, and decreased erythema. The mechanisms for such effects are not known, but one mouse study found that 1,25-dihydroxyvitamin D3 induced expression of metallothionein (a protein that protects against free radicals and oxidative damage) in the stratum basale [93]. It has also been postulated that non-genomic actions of vitamin D contribute to the photoprotection [99]; such effects of vitamin D involve cell-signaling cascades that open calcium channels [100].
Wound healing
1,25-Dihydroxyvitamin D3 regulates the expression of cathelicidin (LL-37/hCAP18) [53], [57], an antimicrobial protein that appears to mediate innate immunity in skin by promoting wound healing and tissue repair. One human study found that cathelicidin expression is upregulated during early stages of normal wound healing [58]. Other studies have shown that cathelicidin modulates inflammation in skin [101], induces angiogenesis [102], and improves reepithelialization (the process of restoring the epidermal barrier to re-establish a functional barrier that protects underlying cells from environmental exposures) [103]. The active form of vitamin D and its analogs have been shown to upregulate cathelicidin expression in cultured keratinocytes [58], [104]. However, more research is needed to determine the role of vitamin D in wound healing and epidermal barrier function, and whether oral vitamin D supplementation or topical treatment with vitamin D analogs is helpful in healing surgical wounds.
Vitamin D and skin diseases
Based on the afore mentioned facts concerning the intertwined bonding that exists between vitamin D and skin, it seems only “natural” to incriminate vitamin D deficiency in a long list of cutaneous disorders including skin cancer, psoriasis, ichthyosis, autoimmune skin disorders such as vitiligo, blistering disorders, scleroderma and systemic lupus erythematosus, as well as atopic dermatitis, acne, hair loss, infections and photodermatoses. Nevertheless, it remains speculative whether vitamin D deficiency primarily contributes to disease pathogenesis or merely represents a consequential event to the inflammatory processes involved. According to a recent systematic review including 290 prospective cohort studies and 172 randomized trials of major health outcomes and of physiological parameters related to disease risk or inflammatory status, one solid fact is emphasized; vitamin D deficiency appears to be a marker of ill health [105] regardless of being an actual cause or an association. In the current review we will highlight the most commonly studied dermatological diseases.
Skin cancer
A number of epidemiologic studies have suggested that vitamin D may have a protective effect decreasing cancer risk and cancer-associated mortality [106], [107], [108], [109], [110]. Adequate vitamin D status has been linked to decreased risks of developing specific cancers, including cancers of the esophagus, stomach, colon, rectum, gallbladder, pancreas, lung, breast, uterus, ovary, prostate, urinary bladder, kidney, skin, thyroid, and hematopoietic system (e.g., Hodgkin’s lymphoma, non-Hodgkin’s lymphoma, multiple myeloma) [110]. With regards to skin cancer, epidemiologic and laboratory studies have reported mixed findings, with some reporting an association between higher vitamin D levels and increased skin cancer risk [111], others showing a decreased skin cancer risk [106], [107], [108], [109], and still others showing no association [106]. The key findings that point to the role of vitamin D in the prevention of the initiation and progression of lethal skin cancers are the involvement of vitamin D in regulation of multiple signaling pathways that have implications in carcinogenesis [109], among which are the inhibition of the hedgehog signaling pathway, the pathway underlying development of basal cell carcinomas, and upregulation of nucleotide excision repair enzymes [106]. Furthermore, vitamin D induces cellular arrest, triggers apoptotic pathways, inhibits angiogenesis, and alters cellular adhesion [108]. Another point is that skin cancer metastasis depends on the tumor microenvironment, where vitamin D metabolites play a key role in prevention of certain molecular events involved in tumor progression [109]. The key factor complicating the association between vitamin D and skin cancer is ultraviolet B radiation. The same spectrum of ultraviolet B radiation that catalyzes the production of vitamin D in the skin also causes DNA damage that can lead to epidermal malignancies. Overall, there is some evidence that vitamin D may play a role in nonmelanoma skin cancer (NMSC) including basal cell and squamous cell carcinoma as well as melanoma prevention, although as of yet there is no direct evidence to show a protective effect [106].
Psoriasis
Psoriasis is a chronic inflammatory skin disease that affects 2–3% of the population worldwide and causes significant morbidity [112]. Although the pathogenesis of psoriasis is not fully understood, there is ample evidence suggesting that the dysregulation of the immune cells in the skin, particularly T cells, plays a critical role in psoriasis development [113].
Several studies have focused on the possible role of vitamin D deficiency in psoriasis [114], [115], [116]. The exact mechanism by which vitamin D deficiency contributes in such a complex pathogenesis is not fully understood. Several pathways have been established including, loss of the anti-proliferative function of vitamin D, as it has been found that human cultured keratinocytes exposed to calcitriol showed marked inhibition of growth and accelerated maturation [117]. Moreover, as inflammation and angiogenesis represent cornerstones in the pathogenesis of psoriasis [118], [119], the loss of the anti-inflammatory and anti-angiogenic activity of vitamin D [108] could represent another explanation to the contribution of the vitamin D deficiency in psoriasis. As 1α,25-dihydroxyvitamin D3 is known to suppress the Th1 and Th17 cell proliferation [69], as well as induce the Tregs [120], another proposed pathway through which vitamin D deficiency could share in the psoriatic predicament would be the unchecked proliferation of Th1 and 17 cells on one hand and unchecked inhibition of Tregs on the other hand. Topical treatment with calcipotriol has been shown to significantly decrease cutaneous levels of human beta defensins (HBD) 2 and HBD3 as well as IL-17A, IL-17F and IL-8, which play significant roles in psoriasis [121], further linking vitamin D deficiency to the pathogenesis of psoriasis.
Owing to this postulated role played by vitamin D in the pathogenesis of psoriasis, it is no wonder that it is one of the most popularly prescribed topical medications for this disease, singly or in combination with betamethasone, and numerous studies documented the efficacy and safety of using topical calcipotriol in the treatment of cases of localized plaque psoriasis [122], [123], [124], [125], [126].
Acne and rosacea
Acne vulgaris is the most common skin disorder affecting millions of people worldwide. Inflammation resulting from the immune response targeting Propionibacterium acnes (P. acnes) has a significant role in acne pathogenesis. In a recent study, it has been demonstrated that P. acnes is a potent inducer of Th17, and that 1,25OH2D inhibits P. acnes-induced Th17 differentiation, and thereby could be considered as an effective tool in modulating acne [127]. Furthermore, sebocytes were identified as 1,25OH2D responsive target cells, indicating that vitamin D analogs may be effective in the treatment of acne. In another recent study, the expression of inflammatory biomarkers have been shown to be influenced by treatment with vitamin D in cultured sebocytes, but not through VDR [128].
In the same spectrum of acne, another study demonstrated relatively high serum levels of vitamin D in patients with rosacea which is a common chronic skin condition affecting the face, in comparison with controls, suggesting that increased vitamin D levels may lead to the development of rosacea [129].
Hair loss
The role of vitamin D in hair might be explained by the fact that an optimal concentration of vitamin D has been suggested to be necessary to delay the aging phenomena, including hair loss [130]. Recently it has been shown that 1,25OH2D/VDR promotes the ability of β-catenin to stimulate hair follicle differentiation [131]. Moreover extensive data from animal models clearly show that the VDR activation plays an important role in the hair follicle cycle, specifically anagen initiation [132]. Interestingly, in VDR ablated mice it did not seem that normalization of mineral ion homeostasis by a diet high in calcium and phosphorous prevented alopecia suggesting that the mechanism for alopecia is unrelated to mineral levels but rather to the vitamin D levels [133]. Furthermore, recent data suggested that VDR regulates directly or indirectly the expression of genes required for hair follicle cycling, including the hedgehog signaling pathway [134].
A recent study conducted on eighty female patients demonstrated that low serum vitamin D2 is associated with both common types of hair loss in females namely; telogen effluvium and androgenetic female pattern hair loss. It was suggested that screening for vitamin D level and supplementation with vitamin D in cases with deficiency would be beneficial in the management of these conditions [135].
In contradistinction to the proposal of the important role played by vitamin D in hair loss, a placebo-controlled trial on 26 patients showed that calcipotriol did not affect the telogen to anagen ratio after 6 weeks of treatment in patients with scalp psoriasis. It is to be noted that the optimal effect of calcipotriol on psoriasis was not seen until 8 weeks, thus, follow up might have been too brief to detect an effect of calcipotriol on hair loss [136]. Furthermore, a cross sectional study of 296 healthy men was done to explore a possible association between male pattern baldness and serum 25-hydroxyvitamin D levels. The severity and extent of the baldness did not appear to be associated with serum 25-hydroxyvitamin D levels [130]. This raises the speculation about the real value of vitamin D levels in hair loss, and whether the story could be intrinsic, closely related to the receptor itself rather than to the level of vitamin D.
Vitiligo
Vitiligo is a common pigmentary disorder characterized by well-demarcated depigmented patches or macules of different shapes and sizes and is caused by the destruction of functional melanocytes in the epidermis [137].
Vitamin D protects the epidermal melanin unit and restores melanocyte integrity via several mechanisms including controlling the activation, proliferation, migration of melanocytes and pigmentation pathways by modulating T cell activation, which is apparently correlated with melanocyte disappearance in vitiligo. The mechanism through which vitamin D exerts its effects on melanocytes is not yet fully understood. Vitamin D is believed to be involved in melanocyte physiology by coordinating melanogenic cytokines [most likely endothelin-3 (ET-3)] and the activity of the SCF/c-Kit system, which is one of the most important regulators of melanocyte viability and maturation [138]. Furthermore, a proposed mechanism involving vitamin D in the protection of vitiliginous skin is based on its antioxidant properties and regulatory function toward the reactive oxygen species that are produced in excess in vitiligo epidermis [139]. Another point is that the active form of vitamin D reduces the apoptotic activity induced by UVB in keratinocytes and melanocytes [140], that has been reported to remove melanocytes from the skin [141]. Moreover, vitamin D might exert immunomodulatory effects by inhibiting the expression of IL-6, IL-8, TNF-α, and TNF-γ, modulate dendritic cell maturation, differentiation, and activation as well as induce the inhibition of antigen presentation [65], thereby dampen the autoimmune pathway incriminated in the pathogenesis of vitiligo.
It is still unknown if vitamin D deficiency plays a role in causing vitiligo, as it does in other autoimmune diseases. In 2010 Silverberg and Silverberg [142] assessed serum 25-hydroxyvitamin D (25(OH)D) levels in 45 patients with vitiligo and it appeared that 55.6% were insufficient (22.5–75 nmol/L) and 13.3% were very low (<.22.5 nmo/L) a finding that was re-demonstrated by others [143]. However, another study showed no correlation between 25(OH)D and vitiligo [144].
Regardless the existing controversy, topical vitamin D3 analogs are members of the armamentarium of therapeutic modalities for vitiligo. The use of vitamin D analogs in combination with PUVAsol and topical calcipotriol for the treatment of vitiligo was first reported by Parsad et al. [145]. Subsequently, a number of studies have reported on the treatment of vitiligo with vitamin D analogs alone or in combination with ultraviolet light or corticosteroids to enhance repigmentation [142], [146], [147] with some contradictory results [148], [149], [150].
Pemphigus vulgaris and bullous pemphigoid
Pemphigus vulgaris and bullous pemphigoid are potentially fatal autoimmune bullous disorders caused by keratinocyte acantholysis as a result of pathogenic antibody production by B cells. Vitamin D, through its participation in several immune modulatory functions including B cells apoptosis, Th2 cell differentiation, apoptotic enzyme regulation and Tregs functions, may be actively involved in the immune regulation of such diseases. Several recent studies demonstrated that patients with pemphigus vulgaris and bullous pemphigoid have significantly lower serum vitamin D levels in comparison with controls regardless age, body mass index or pattern of sun exposure [151], [152]. In addition, it was suggested that this lower level of vitamin D might account for the increased prevalence of fractures in such patients and therefore should be taken into consideration in patients who must be given corticosteroids [152].
Atopic dermatitis
Atopic dermatitis (AD) is a common chronic inflammatory type of eczema. Several studies have shown initial epidermal barrier dysfunction with subsequent immune activation as the underlying mechanism. Animal studies, case reports, and randomized clinical trials have suggested that vitamin D, through various mechanisms including immunomodulation, may alleviate the symptoms of AD. The majority of these studies indicate an inverse relationship between the severity of atopic dermatitis and vitamin D levels. Furthermore, studies have shown that, in individuals with AD who are deficient in vitamin D, repletion of vitamin D results in improvement and decreased severity of the disease [153], [154].
Should vitamin D be scripted on every prescription?
The answer to this question is still far from clear, but at least we could clearly recommend routine evaluation of its level, with particular focus on those who are at risk of its deficiency e.g. elderly, obese, lacking proper sun exposure or with malabsorption disorders. Vitamin D supplementation could represent an important adjuvant treatment if deficient or insufficient.
Conclusions
In conclusion one could clearly sense the unique relationship that entangles vitamin D to dermatology. On one hand, our skin is one source for this important vitamin and on the other hand all available data point to its important impact on the health of our skin and the involvement of its deficiency in the pathway of many dermatological diseases. Several factors are responsible for maintaining it in optimum levels; therefore sunny climates are by far not a guarantee for providing a “comfort zone” regarding the possibility of this vitamin deficiency, a concern documented by several epidemiological studies carried out in areas close to the equator [155], [156], [157], [158]. On the basis of currently available data, it is clear that supplemental vitamin D should be the preferred recommendation toward achieving its normal serum levels, thereby avoiding the deleterious effects accompanied by its deficiency. Still more research is needed to unravel its complicated ties to dermatological diseases and create clear guidelines and recommendations for its supplementation.
Conflict of Interest
The authors have declared no conflict of interest.
Compliance with Ethics Requirements
This article does not contain any studies with human or animal subjects.
Biographies

Dr. Wedad Z. Mostafa, Emeritus Professor at Cairo University Department of Dermatology, obtained her Masters and MD degrees in Dermatology in 1978 and 1984. She taught at Dammam University, Saudi Arabia during the eighties and won the Janssen Research Council Award in 1993. Interests in research led to over 50 publications in renowned international and regional journals including the Journal of the American Academy of Dermatology, Pediatric Dermatology and the International Journal of Dermatology. She was a visiting lecturer at the Department of Dermatology RWTH Aachen, Germany during 2011 and 2012. An active member of Vitiligo Groups, Dr Mostafa resides in Cairo, Egypt.

Dr. Rehab A. Hegazy, Associate Professor at Cairo University, Department of Dermatology graduated from CU Medical School in 2001. She obtained Masters and MD degrees in Dermatology in 2006, 2009. She has over 40 publications and is a reviewer in a number of international and national journals, as well as a member in a group of medical societies. She won “The John Stratigos Memorial Scholarship”; 2012, “Omar Ibn Abd ElAziz Al Sheikh prize for Scientific Research”; 2013, “Best abstract award in 3rd 5 continent congress for Lasers and Aesthetic Medicine, Cannes”; 2013 and “Samy ElSogeir prize for scientific research”; 2015.
Footnotes
Peer review under responsibility of Cairo University.

References
- 1.Norman A. Vitamin D. In: Bowman B.A., Russel R.E., editors. Present knowledge of nutrition. 8th ed. ILSI Press; Washington, DC: 2001. [chapter 13] [Google Scholar]
- 2.Vanchinathan V., Lim H.W. A dermatologist’s perspective on vitamin D. Mayo Clin Proc. 2012;87:372–380. doi: 10.1016/j.mayocp.2011.12.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bikle D.D. Vitamin D metabolism and function in the skin. Mol Cell Endocrinol. 2011;347:80–89. doi: 10.1016/j.mce.2011.05.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Heaney R.P. Vitamin D in health and disease. Clin J Am Soc Nephrol. 2008;3:1535–1541. doi: 10.2215/CJN.01160308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Lehmann B., Meurer M. Vitamin D metabolism. Dermatol Ther. 2010;23:2–12. doi: 10.1111/j.1529-8019.2009.01286.x. [DOI] [PubMed] [Google Scholar]
- 6.Reddy K.K., Gilchrest B.A. What is all this commotion about vitamin D? J Invest Dermatol. 2010;130:321–326. doi: 10.1038/jid.2009.353. [DOI] [PubMed] [Google Scholar]
- 7.Tsiaras W.G., Weinstock M.A. Factors influencing vitamin D status. Acta Derma Venereol. 2011;91:115–124. doi: 10.2340/00015555-0980. [DOI] [PubMed] [Google Scholar]
- 8.Armas L.G., Heaney R.P. Vitamin D: the iceberg nutrient. J Ren Nutr. 2011;21:134–139. doi: 10.1053/j.jrn.2010.09.002. [DOI] [PubMed] [Google Scholar]
- 9.Kochupillai N. The physiology of vitamin D: current concepts. Indian J Med Res. 2008;127:256–262. [PubMed] [Google Scholar]
- 10.Gilchrest B.A. Sun exposure and vitamin D sufficiency. Am J Clin Nutr. 2008;88:570S–577S. doi: 10.1093/ajcn/88.2.570S. [DOI] [PubMed] [Google Scholar]
- 11.Holick M.F., Vitamin D. In: Modern nutrition in health and disease. Shils M.E., Olson J.A., Shike M., Ross A.C., editors. Lippincott; Williams & Wilkins: 1999. pp. 228–239. [Google Scholar]
- 12.CIE. Action spectrum for the production of previtamin D3 in human skin. Technical report 174; 2006.
- 13.Krause R., Bohring M., Hopfenmüller W., Holick M.F., Sharma A.M. Ultraviolet B and blood pressure. Lancet. 1998;352:709–710. doi: 10.1016/S0140-6736(05)60827-6. [DOI] [PubMed] [Google Scholar]
- 14.Stamp T.C., Haddad J.G., Twigg C.A. Comparison of oral 25-hydroxycholecalciferol, vitamin D, and ultraviolet light as determinants of circulating 25-hydroxyvitamin D. Lancet. 1977;1:1341–1343. doi: 10.1016/s0140-6736(77)92553-3. [DOI] [PubMed] [Google Scholar]
- 15.Clemens T.L., Adams J.S., Henderson S.L., Holick M.F. Increased skin pigment reduces the capacity of skin to synthesize vitamin D3. Lancet. 1982;1:74–76. doi: 10.1016/s0140-6736(82)90214-8. [DOI] [PubMed] [Google Scholar]
- 16.Zittermann A., Frisch S., Berthold H.K., Götting C., Kuhn J., Kleesiek K. Vitamin D supplementation enhances the beneficial effects of weight loss on cardiovascular risk markers. Am J Clin Nutr. 2009;89:1321–1327. doi: 10.3945/ajcn.2008.27004. [DOI] [PubMed] [Google Scholar]
- 17.Preece M.A., Tomlinson S., Ribot C.A., Pietrek J., Korn H.T., Davies D.M. Studies of vitamin D deficiency in man. Q J Med. 1975;44:575–589. [PubMed] [Google Scholar]
- 18.Dlugos D.J., Perrotta P.L., Horn W.G. Effects of the submarine environment on renal-stone risk factors and vitamin D metabolism. Undersea Hyperb Med. 1995;22:145–152. [PubMed] [Google Scholar]
- 19.Zittermann A., Koerfer R. Protective and toxic effects of vitamin D on vascular calcification: clinical implications. Mol Aspects Med. 2008;29:423–432. doi: 10.1016/j.mam.2008.04.002. [DOI] [PubMed] [Google Scholar]
- 20.Shahriari M., Kerr P.E., Slade K., Grant-Kels J.E. Vitamin D and the skin. Clin Dermatol. 2010;28:663–668. doi: 10.1016/j.clindermatol.2010.03.030. [DOI] [PubMed] [Google Scholar]
- 21.Antoniucci D.M., Yamashita T., Portaloe A.A. Dietary phosphorus regulates serum fibroblast growth factor-23 concentrations in healthy men. J Clin Endocrinol Metab. 2006;91:3144–3149. doi: 10.1210/jc.2006-0021. [DOI] [PubMed] [Google Scholar]
- 22.Committee of Revision. Drug information for the health care professional. Rockville, MD: United States Pharmacopeial Convention Inc.; 1997.
- 23.Institute of Medicine. Standing Committee on the Scientific Evaluation of Dietary Reference Intakes. Vitamin D. Dietary reference intakes: calcium, phosphorus, magnesium, vitamin D, and fluoride. Washington, DC: National Academy Press; 1997. p. 250–87.
- 24.Medicines Commission. British Pharmacopoeia 1980. London, United Kingdom: Her Majesty’s Stationery Office; 1980.
- 25.Houghton L.A., Vieth R. The case against ergocalciferol (vitamin D2) as a vitamin supplement. Am J Clin Nutr. 2006;84:694–697. doi: 10.1093/ajcn/84.4.694. [DOI] [PubMed] [Google Scholar]
- 26.LoPiccolo M.C., Lim H.W. Vitamin D in health and disease. Photodermatol Photoimmunol Photomed. 2010;26:224–229. doi: 10.1111/j.1600-0781.2010.00524.x. [DOI] [PubMed] [Google Scholar]
- 27.Holick M.F., Binkley N.C., Bischoff-Ferrari H.A., Gordon C.M., Hanley D.A., Heaney R.P. Evaluation, treatment, and prevention of vitamin D deficiency: an Endocrine Society clinical practice guideline. J Clin Endocrinol Metab. 2011;96:1911–1930. doi: 10.1210/jc.2011-0385. [DOI] [PubMed] [Google Scholar]
- 28.Kull M., Jr, Kallikorm R., Tamm A., Lember M. Seasonal variance of 25-(OH) vitamin D in the general population of Estonia, a Northern European country. BMC Public Health. 2009;19(9):22. doi: 10.1186/1471-2458-9-22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Tangpricha V., Pearce E.N., Chen T.C., Holick M.F. Vitamin D insufficiency among free-living healthy young adults. Am J Med. 2002;112:659–662. doi: 10.1016/s0002-9343(02)01091-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.MacFarlane G.D., Sackrison J.L., Jr, Body J.J., Ersfeld D.L., Fenske J.S., Miller A.B. Hypovitaminosis D in a normal, apparently healthy urban European population. J Steroid Biochem Mol Biol. 2004;89–90:621–622. doi: 10.1016/j.jsbmb.2004.03.042. [DOI] [PubMed] [Google Scholar]
- 31.Malabanan A., Veronikis I.E., Holick M.F. Redefining vitamin D insufficiency. Lancet. 1998;351:805–806. doi: 10.1016/s0140-6736(05)78933-9. [DOI] [PubMed] [Google Scholar]
- 32.Chapuy M.C., Preziosi P., Maamer M., Arnaud S., Galan P., Hercberg S. Prevalence of vitamin D insufficiency in an adult normal population. Osteoporos Int. 1997;7:439–443. doi: 10.1007/s001980050030. [DOI] [PubMed] [Google Scholar]
- 33.Bischoff-Ferrari H.A., Dietrich T., Orav E.J., Dawson-Hughes B. Positive association between 25-hydroxy vitamin D levels and bone mineral density: a population-based study of younger and older adults. Am J Med. 2004;116:634–639. doi: 10.1016/j.amjmed.2003.12.029. [DOI] [PubMed] [Google Scholar]
- 34.Bischoff-Ferrari H.A. The 25-hydroxyvitamin D threshold for better health. J Steroid Biochem Mol Biol. 2007;103:614–619. doi: 10.1016/j.jsbmb.2006.12.016. [DOI] [PubMed] [Google Scholar]
- 35.Dawson-Hughes B., Heaney R.P., Holick M.F., Lips P., Meunier P.J., Vieth R. Estimates of optimal vitamin D status. Osteoporos Int. 2005;16:713–716. doi: 10.1007/s00198-005-1867-7. [DOI] [PubMed] [Google Scholar]
- 36.Bischoff-Ferrari H.A., Giovannucci E., Willett W.C., Dietrich T., wson-Hughes B. Estimation of optimal serum concentrations of 25-hydroxyvitamin D for multiple health outcomes. Am J Clin Nutr. 2006;84:18–28. doi: 10.1093/ajcn/84.1.18. [DOI] [PubMed] [Google Scholar]
- 37.Institute of Medicine, Food and Nutrition Board. Dietary reference intakes for calcium and vitamin D. Washington, DC: National Academy Press; 2010.
- 38.Holick M.F. Vitamin D deficiency. N Engl J Med. 2007;357:266–281. doi: 10.1056/NEJMra070553. [DOI] [PubMed] [Google Scholar]
- 39.Engelsen O. The relationship between ultraviolet radiation exposure and vitamin D status. Nutrients. 2010;2:482–495. doi: 10.3390/nu2050482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Greene-Finestone L.S., Berger C., de Groh M., Hanley D.A., Hidiroglou N., Sarafin K. 25-Hydroxyvitamin D in Canadian adults: biological, environmental, and behavioral correlates. Osteoporos Int. 2011;22:1389–1399. doi: 10.1007/s00198-010-1362-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Gozdzik A., Barta J.L., Wu H., Wagner D., Cole D.E., Vieth R. Low wintertime vitamin D levels in a sample of healthy young adults of diverse ancestry living in the Toronto area: associations with vitamin D intake and skin pigmentation. BMC Public Health. 2008;8:336. doi: 10.1186/1471-2458-8-336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Ardawi M.S., Qari M.H., Rouzi A.A., Maimani A.A., Raddadi R.M. Vitamin D status in relation to obesity, bone mineral density, bone turnover markers and vitamin D receptor genotypes in healthy Saudi pre- and postmenopausal women. Osteoporos Int. 2011;22:463–475. doi: 10.1007/s00198-010-1249-7. [DOI] [PubMed] [Google Scholar]
- 43.Parkin D.M., Mesher D., Sasieni P. Cancers attributable to solar (ultraviolet) radiation exposure in the UK in 2010. Br J Cancer. 2011;105:S66–S69. doi: 10.1038/bjc.2011.486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Bikle D.D., Pillai S. Vitamin D, calcium, and epidermal differentiation. Endocr Rev. 1993;14:3–19. doi: 10.1210/edrv-14-1-3. [DOI] [PubMed] [Google Scholar]
- 45.Hawker N.P., Pennypacker S.D., Chang S.M., Bikle D.D. Regulation of human epidermal keratinocyte differentiation by the vitamin D receptor and its coactivators DRIP205, SRC2, and SRC3. J Invest Dermatol. 2007;127:874. doi: 10.1038/sj.jid.5700624. [DOI] [PubMed] [Google Scholar]
- 46.Ratnam A.V., Bikle D.D., Cho J.K. 1,25 Dihydroxyvitamin D3 enhances the calcium response of keratinocytes. J Cell Physiol. 1999;178:188–196. doi: 10.1002/(SICI)1097-4652(199902)178:2<188::AID-JCP8>3.0.CO;2-4. [DOI] [PubMed] [Google Scholar]
- 47.Pillai S., Bikle D.D., Su M.J., Ratnam A., Abe J. 1,25 Dihydroxyvitamin D upregulates the phosphatidyl inositol signalling pathway in human keratinocytes by increasing phospholipase C levels. J Clin Invest. 1995;96:602–609. doi: 10.1172/JCI118075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Tu C.L., Chang W., Bikle D.D. The extracellular calcium-sensing receptor is required for calcium-induced differentiation in human keratinocytes. J Biol Chem. 2001;276:41079–41085. doi: 10.1074/jbc.M107122200. [DOI] [PubMed] [Google Scholar]
- 49.Xie Z., Bikle D.D. Inhibition of 1,25-dihydroxyvitamin-D-induced keratinocyte differentiation by blocking the expression of phospholipase C-gamma1. J Invest Dermatol. 2001;117:1250–1254. doi: 10.1046/j.0022-202x.2001.01526.x. [DOI] [PubMed] [Google Scholar]
- 50.Bikle D.D., Elalieh H., Chang S., Xie Z., Sundberg J.P. Development and progression of alopecia in the vitamin D receptor null mouse. J Cell Physiol. 2006;207:340–353. doi: 10.1002/jcp.20578. [DOI] [PubMed] [Google Scholar]
- 51.Xie Z., Komuves L., Yu Q.C., Elalieh H., Ng D.C., Leary C. Lack of the vitamin D receptor is associated with reduced epidermal differentiation and hair follicle growth. J Invest Dermatol. 2002;118:11–16. doi: 10.1046/j.1523-1747.2002.01644.x. [DOI] [PubMed] [Google Scholar]
- 52.Oda Y., Uchida Y., Moradian S., Crumrine D., Elias P.M., Bikle D.D. Vitamin D receptor and coactivators SRC 2 and 3 regulate epidermis-specific sphingolipid production and permeability barrier formation. J Invest Dermatol. 2009;129:1367–1378. doi: 10.1038/jid.2008.380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Schauber J., Dorschner R.A., Coda A.B., Büchau A.S., Liu P.T., Kiken D. Injury enhances TLR2 function and antimicrobial peptide expression through a vitamin D-dependent mechanism. J Clin Invest. 2007;117:803–811. doi: 10.1172/JCI30142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Grad R. Cod and the consumptive: a brief history of cod-liver oil in the treatment of pulmonary tuberculosis. Pharm Hist. 2004;46:106–120. [PubMed] [Google Scholar]
- 55.Rook G.A., Steele J., Fraher L., Barker S., Karmali R., O’Riordan J. Vitamin D3, gamma interferon, and control of proliferation of Mycobacterium tuberculosis by human monocytes. Immunology. 1986;57:159–163. [PMC free article] [PubMed] [Google Scholar]
- 56.Hewison M. An update on vitamin D and human immunity. Clin Endocrinol (Oxf) 2012;76:315–325. doi: 10.1111/j.1365-2265.2011.04261.x. [DOI] [PubMed] [Google Scholar]
- 57.Wang T.T., Nestel F.P., Bourdeau V., Nagai Y., Wang Q., Liao J. Cutting edge: 1,25-dihydroxyvitamin D3 is a direct inducer of antimicrobial peptide gene expression. J Immunol. 2004;173:2909–2912. doi: 10.4049/jimmunol.173.5.2909. [DOI] [PubMed] [Google Scholar]
- 58.Gombart A.F., Borregaard N., Koeffler H.P. Human cathelicidin antimicrobial peptide (CAMP) gene is a direct target of the vitamin D receptor and is stronglyup-regulated in myeloid cells by 1,25-dihydroxyvitamin D3. FASEB J. 2005;19:1067–1077. doi: 10.1096/fj.04-3284com. [DOI] [PubMed] [Google Scholar]
- 59.Liu P.T., Stenger S., Li H., Wenzel L., Tan B.H., Krutzik S.R. Toll-like receptor triggering of a vitamin D-mediated human antimicrobial response. Science. 2006;311:1770–1773. doi: 10.1126/science.1123933. [DOI] [PubMed] [Google Scholar]
- 60.Adams J.S., Ren S., Liu P.T., Chun R.F., Lagishetty V., Gombart A.F. Vitamin d-directed rheostatic regulation of monocyte antibacterial responses. J Immunol. 2009;182:4289–4295. doi: 10.4049/jimmunol.0803736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Edfeldt K., Liu P.T., Chun R., Fabri M., Schenk M., Wheelwright M. T-cell cytokines differentially control human monocyte antimicrobial responses by regulating vitamin Dmetabolism. Proc Natl Acad Sci USA. 2010;107:22593–22598. doi: 10.1073/pnas.1011624108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Krutzik S.R., Hewison M., Liu P.T., Robles J.A., Stenger S., Adams J.S. IL-15 links TLR2/1-induced macrophage differentiation to the vitamin D-dependent antimicrobial pathway. J Immunol. 2008;181:7115–7120. doi: 10.4049/jimmunol.181.10.7115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Liu P.T., Schenk M., Walker V.P., Dempsey P.W., Kanchanapoomi M., Wheelwright M. Convergence of IL-1beta and VDR activation pathways in human TLR2/1-induced antimicrobial responses. PLoS One. 2009;4:e5810. doi: 10.1371/journal.pone.0005810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Brennan A., Katz D.R., Nunn J.D., Barker S., Hewison M., Fraher L.J. Dendritic cells from human tissues express receptors for the immunoregulatory vitamin D3 metabolite, dihydroxycholecalciferol. Immunology. 1987;61:457–461. [PMC free article] [PubMed] [Google Scholar]
- 65.Penna G., Adorini L. 1 Alpha,25-dihydroxyvitamin D3 inhibits differentiation, maturation, activation, and survival of dendritic cells leading to impaired alloreactive T cell activation. J Immunol. 2000;164:2405–2411. doi: 10.4049/jimmunol.164.5.2405. [DOI] [PubMed] [Google Scholar]
- 66.Adorini L., Penna G., Giarratana N., Uskokovic M. Tolerogenic dendritic cells induced by vitamin D receptor ligands enhance regulatory T cells inhibiting allograftrejection and autoimmune diseases. J Cell Biochem. 2003;88:227–233. doi: 10.1002/jcb.10340. [DOI] [PubMed] [Google Scholar]
- 67.Provvedini D.M., Tsoukas C.D., Deftos L.J., Manolagas S.C. 1,25-Dihydroxyvitamin D3 receptors in human leukocytes. Science. 1983;221:1181–1183. doi: 10.1126/science.6310748. [DOI] [PubMed] [Google Scholar]
- 68.Lemire J.M., Adams J.S., Sakai R., Jordan S.C. 1 Alpha,25-dihydroxyvitamin D3 suppresses proliferation and immunoglobulin production by normal human peripheral blood mononuclear cells. J Clin Invest. 1984;74:657–661. doi: 10.1172/JCI111465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Lemire J.M., Adams J.S., Kermani-Arab V., Bakke A.C., Sakai R., Jordan S.C. 1,25-Dihydroxyvitamin D3 suppresses human T helper/inducer lymphocyte activity in vitro. J Immunol. 1985;134:3032–3035. [PubMed] [Google Scholar]
- 70.Abbas A.K., Murphy K.M., Sher A. Functional diversity of helper T lymphocytes. Nature. 1996;383:787–793. doi: 10.1038/383787a0. [DOI] [PubMed] [Google Scholar]
- 71.Romagnani S. Regulation of the T cell response. Clin Exp Allergy. 2006;36:1357–1366. doi: 10.1111/j.1365-2222.2006.02606.x. [DOI] [PubMed] [Google Scholar]
- 72.Lemire J.M., Archer D.C., Beck L., Spiegelberg H.L. Immunosuppressive actions of 1,25-dihydroxyvitamin D3: preferential inhibition of Th1 functions. J Nutr. 1995;125:1704S–1708S. doi: 10.1093/jn/125.suppl_6.1704S. [DOI] [PubMed] [Google Scholar]
- 73.Boonstra A., Barrat F.J., Crain C., Heath V.L., Savelkoul H.F., O’Garra A. 1 Alpha,25-dihydroxyvitamin d3 has a direct effect on naive CD4(+) T cells to enhance the development of Th2 cells. J Immunol. 2001;167:4974–4980. doi: 10.4049/jimmunol.167.9.4974. [DOI] [PubMed] [Google Scholar]
- 74.Penna G., Amuchastegui S., Cossetti C., Aquilano F., Mariani R., Sanvito F. Treatment of experimental autoimmune prostatitis in nonobese diabetic mice by the vitamin D receptor agonist elocalcitol. J Immunol. 2006;177:8504–8511. doi: 10.4049/jimmunol.177.12.8504. [DOI] [PubMed] [Google Scholar]
- 75.Tang J., Zhou R., Luger D., Zhu W., Silver P.B., Grajewski R.S. Calcitriol suppresses antiretinal autoimmunity through inhibitory effects on the Th17 effector response. J Immunol. 2009;182:4624–4632. doi: 10.4049/jimmunol.0801543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Joshi S., Pantalena L.C., Liu X.K., Gaffen S.L., Liu H., Rohowsky-Kochan C. 1,25-Dihydroxyvitamin D(3) ameliorates Th17 autoimmunity via transcriptional modulation of interleukin-17A. Mol Cell Biol. 2011;31:3653–3669. doi: 10.1128/MCB.05020-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Barrat F.J., Cua D.J., Boonstra A., Richards D.F., Crain C., Savelkoul H.F. In vitro generation of interleukin 10-producing regulatory CD4(+) T cells is induced by immunosuppressive drugsand inhibited by T helper type 1 (Th1)- and Th2-inducing cytokines. J Exp Med. 2002;195:603–616. doi: 10.1084/jem.20011629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Rudensky A.Y. Regulatory T cells and Foxp3. Immunol Rev. 2011;241:260–268. doi: 10.1111/j.1600-065X.2011.01018.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Ardalan M.R., Maljaei H., Shoja M.M., Piri A.R., Khosroshahi H.T., Noshad H. Calcitriol started in the donor, expands the population of CD4+ CD25+ T cells in renal transplant recipients. Transplant Proc. 2007;39:951–953. doi: 10.1016/j.transproceed.2007.04.012. [DOI] [PubMed] [Google Scholar]
- 80.Royal W., 3rd, Mia Y., Li H., Naunton K. Peripheral blood regulatory T cell measurements correlate with serum vitamin D levels in patients with multiple sclerosis. J Neuroimmunol. 2009;213:135–141. doi: 10.1016/j.jneuroim.2009.05.012. [DOI] [PubMed] [Google Scholar]
- 81.Smolders J., Thewissen M., Peelen E., Menheere P., Tervaert J.W., Damoiseaux J. Vitamin D status is positively correlated with regulatory T cell function in patients with multiple sclerosis. PLoS One. 2009;4:e6635. doi: 10.1371/journal.pone.0006635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Penna G., Amuchastegui S., Giarratana N., Daniel K.C., Vulcano M., Sozzani S. 1,25-Dihydroxyvitamin D3 selectively modulates tolerogenic properties in myeloid but not plasmacytoid dendritic cells. J Immunol. 2007;178:145–153. doi: 10.4049/jimmunol.178.1.145. [DOI] [PubMed] [Google Scholar]
- 83.Jeffery L.E., Burke F., Mura M., Zheng Y., Qureshi O.S., Hewison M. 1,25-Dihydroxyvitamin D3 and IL-2 combine to inhibit T cell production of inflammatory cytokines and promote development of regulatory T cells expressing CTLA-4 and FoxP3. J Immunol. 2009;183:5458–5467. doi: 10.4049/jimmunol.0803217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Hewison M., Freeman L., Hughes S.V., Evans K.N., Bland R., Eliopoulos A.G. Differential regulation of vitamin D receptor and its ligand in human monocyte-derived dendritic cells. J Immunol. 2003;170:5382–5390. doi: 10.4049/jimmunol.170.11.5382. [DOI] [PubMed] [Google Scholar]
- 85.Sigmundsdottir H., Pan J., Debes G.F., Alt C., Habtezion A., Soler D. DCs metabolize sunlight-induced vitamin D3 to ‘program’ T cell attraction to the epidermal chemokine CCL27. Nat Immunol. 2007;8:285–293. doi: 10.1038/ni1433. [DOI] [PubMed] [Google Scholar]
- 86.Chen S., Sims G.P., Chen X.X., Gu Y.Y., Chen S., Lipsky P.E. Modulatory effects of 1,25-dihydroxyvitamin D3 on human B cell differentiation. J Immunol. 2007;179:1634–1647. doi: 10.4049/jimmunol.179.3.1634. [DOI] [PubMed] [Google Scholar]
- 87.Heine G., Niesner U., Chang H.D., Steinmeyer A., Zügel U., Zuberbier T. 1,25-Dihydroxyvitamin D(3) promotes IL-10 production in human B cells. Eur J Immunol. 2008;38:2210–2218. doi: 10.1002/eji.200838216. [DOI] [PubMed] [Google Scholar]
- 88.Shirakawa A.K., Nagakubo D., Hieshima K., Nakayama T., Jin Z., Yoshie O. 1,25-Dihydroxyvitamin D3 induces CCR10 expression in terminally differentiating human B cells. J Immunol. 2008;180:2786–2795. doi: 10.4049/jimmunol.180.5.2786. [DOI] [PubMed] [Google Scholar]
- 89.Amor K.T., Rashid R.M., Mirmirani P. Does D matter? The role of vitamin D in hair disorders and hair follicle cycling. Dermatol Online J. 2010;16:3. [PubMed] [Google Scholar]
- 90.Wang J., Lu Z., Au J.L. Protection against chemotherapy-induced alopecia. Pharm Res. 2006;23:2505–2514. doi: 10.1007/s11095-006-9105-3. [DOI] [PubMed] [Google Scholar]
- 91.Reichrath J., Schuler Ch., Seifert M., Zouboulis Ch., Tilgen W. The vitamin D endocrine system of human sebocytes. Exp Dermatol. 2006;15:643. [Google Scholar]
- 92.Reichrath J. Vitamin D and the skin: an ancient friend, revisited. Exp Dermatol. 2007;16:618–625. doi: 10.1111/j.1600-0625.2007.00570.x. [DOI] [PubMed] [Google Scholar]
- 93.Lee J., Youn J.I. The photoprotective effect of 1,25-dihydroxyvitamin D3 on ultraviolet light B-induced damage in keratinocyte and its mechanism of action. J Dermatol Sci. 1998;18:11–18. doi: 10.1016/s0923-1811(98)00015-2. [DOI] [PubMed] [Google Scholar]
- 94.Wong G., Gupta R., Dixon K.M., Deo S.S., Choong S.M., Halliday G.M. 1,25-Dihydroxyvitamin D and three low-calcemic analogs decrease UV-induced DNA damage via the rapid response pathway. J Steroid Biochem Mol Biol. 2004;89–90:567–570. doi: 10.1016/j.jsbmb.2004.03.072. [DOI] [PubMed] [Google Scholar]
- 95.De Haes P., Garmyn M., Degreef H., Vantieghem K., Bouillon R., Segaert S. 1,25-Dihydroxyvitamin D3 inhibits ultraviolet B-induced apoptosis, Jun kinase activation, and interleukin-6 production in primary human keratinocytes. J Cell Biochem. 2003;89:663–673. doi: 10.1002/jcb.10540. [DOI] [PubMed] [Google Scholar]
- 96.Dixon K.M., Deo S.S., Wong G., Slater M., Norman A.W., Bishop J.E. Skin cancer prevention: a possible role of 1,25dihydroxyvitamin D3 and its analogs. J Steroid Biochem Mol Biol. 2005;97:137–143. doi: 10.1016/j.jsbmb.2005.06.006. [DOI] [PubMed] [Google Scholar]
- 97.Dixon K.M., Deo S.S., Norman A.W., Bishop J.E., Halliday G.M., Reeve V.E. In vivo relevance for photoprotection by the vitamin D rapid response pathway. J Steroid Biochem Mol Biol. 2007;103:451–456. doi: 10.1016/j.jsbmb.2006.11.016. [DOI] [PubMed] [Google Scholar]
- 98.Gupta R., Dixon K.M., Deo S.S., Holliday C.J., Slater M., Halliday G.M. Photoprotection by 1,25 dihydroxyvitamin D3 is associated with an increase in p53 and a decrease in nitric oxideproducts. J Invest Dermatol. 2007;127:707–715. doi: 10.1038/sj.jid.5700597. [DOI] [PubMed] [Google Scholar]
- 99.Mason R.S., Sequeira V.B., Dixon K.M., Gordon-Thomson C., Pobre K., Dilley A. Photoprotection by 1alpha,25-dihydroxyvitamin D and analogs: further studies on mechanisms and implications for UV-damage. J Steroid Biochem Mol Biol. 2010;121:164–168. doi: 10.1016/j.jsbmb.2010.03.082. [DOI] [PubMed] [Google Scholar]
- 100.Revelli A., Massobrio M., Tesarik J. Nongenomic effects of 1alpha,25-dihydroxyvitamin D(3) Trends Endocrinol Metab. 1998;9:419–427. doi: 10.1016/s1043-2760(98)00100-3. [DOI] [PubMed] [Google Scholar]
- 101.Heilborn J.D., Nilsson M.F., Kratz G., Weber G., Sørensen O., Borregaard N. The cathelicidin anti-microbial peptide LL-37 is involved in re-epithelialization of human skin wounds and is lacking in chronic ulcer epithelium. J Invest Dermatol. 2003;120:379–389. doi: 10.1046/j.1523-1747.2003.12069.x. [DOI] [PubMed] [Google Scholar]
- 102.Frohm M., Agerberth B., Ahangari G., Stâhle-Bäckdahl M., Lidén S., Wigzell H. The expression of the gene coding for the antibacterial peptide LL-37 is induced in human keratinocytes duringinflammatory disorders. J Biol Chem. 1997;272:15258–15263. doi: 10.1074/jbc.272.24.15258. [DOI] [PubMed] [Google Scholar]
- 103.Koczulla R., von Degenfeld G., Kupatt C., Krötz F., Zahler S., Gloe T. An angiogenic role for the human peptide antibiotic LL-37/hCAP-18. J Clin Invest. 2003;111:1665–1672. doi: 10.1172/JCI17545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Weber G., Heilborn J.D., Chamorro Jimenez C.I., Hammarsjo A., Törmä H., Stahle M. Vitamin D induces the antimicrobial protein hCAP18 in human skin. J Invest Dermatol. 2005;124:1080–1082. doi: 10.1111/j.0022-202X.2005.23687.x. [DOI] [PubMed] [Google Scholar]
- 105.Autier P., Boniol M., Pizot C., Mullie P. Vitamin D status and ill health: a systematic review. Lancet Diabetes Endocrinol. 2014;2:76–89. doi: 10.1016/S2213-8587(13)70165-7. [DOI] [PubMed] [Google Scholar]
- 106.Tang JY, Fu T, Lau C, Oh DH, Bikle DD, Asgari MM. Vitamin D in cutaneous carcinogenesis: Part II. J Am Acad Dermatol 2012; 67: 817.e1–11; quiz 827–8. [DOI] [PMC free article] [PubMed]
- 107.Chiang K.C., Chen T.C. The anti-cancer actions of vitamin D. Anticancer Agents Med Chem. 2013;13:126–139. [PubMed] [Google Scholar]
- 108.Picotto G., Liaudat A.C., Bohl L., Talamoni N.T. Molecular aspects of vitamin d anticancer activity. Cancer Invest. 2012;30:604–614. doi: 10.3109/07357907.2012.721039. [DOI] [PubMed] [Google Scholar]
- 109.Nemazannikova N., Antonas K., Dass C.R. Role of vitamin D metabolism in cutaneous tumour formation and progression. J Pharm Pharmacol. 2013;65:2–10. doi: 10.1111/j.2042-7158.2012.01527.x. [DOI] [PubMed] [Google Scholar]
- 110.Glade MJ. Vitamin D: health panacea or false prophet? Nutrition 2012. pii: S0899-9007(12)00252-3. [DOI] [PubMed]
- 111.Afzal S., Nordestgaard B.G., Bojesen S.E. Plasma 25-hydroxyvitamin D and risk of non-melanoma and melanoma skin cancer: a prospective cohort study. J Invest Dermatol. 2013;133:629–636. doi: 10.1038/jid.2012.395. [DOI] [PubMed] [Google Scholar]
- 112.Fitch E., Harper E., Skorcheva I., Kurtz S.E., Blauvelt A. Pathophysiology of psoriasis: recent advances on IL-23 and Th17 cytokines. Curr Rheumatol Rep. 2007;9:461–467. doi: 10.1007/s11926-007-0075-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Cai Y., Fleming C., Yan J. New insights of T cells in the pathogenesis of psoriasis. Cell Mol Immunol. 2012;9:302–309. doi: 10.1038/cmi.2012.15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Gisondi P., Rossini M., Di Cesare A., Idolazzi L., Farina S., Beltrami G. Vitamin D status in patients with chronic plaque psoriasis. Br J Dermatol. 2012;166:505–510. doi: 10.1111/j.1365-2133.2011.10699.x. [DOI] [PubMed] [Google Scholar]
- 115.Orgaz-Molina J., Buendía-Eisman A., Arrabal-Polo M.A., Ruiz J.C., Arias-Santiago S. Deficiency of serum concentration of 25-hydroxyvitamin D in psoriatic patients: a case-control study. J Am Acad Dermatol. 2012;67:931–938. doi: 10.1016/j.jaad.2012.01.040. [DOI] [PubMed] [Google Scholar]
- 116.El-Moaty Zaher H.A., El-Komy M.H., Hegazy R.A., Mohamed El Khashab H.A., Ahmed H.H. Assessment of interleukin-17 and vitamin D serum levels in psoriatic patients. J Am Acad Dermatol. 2013;69:840–842. doi: 10.1016/j.jaad.2013.07.026. [DOI] [PubMed] [Google Scholar]
- 117.Holick M.F. Vitamin D: a millenium perspective. J Cell Biochem. 2003;88:296–307. doi: 10.1002/jcb.10338. [DOI] [PubMed] [Google Scholar]
- 118.Botti E., Spallone G., Caruso R., Monteleone G., Chimenti S., Costanzo A. Psoriasis, from pathogenesis to therapeutic strategies: IL-21 as a novel potential therapeutic target. Curr Pharm Biotechnol. 2012;13:1861–1867. doi: 10.2174/138920112802273281. [DOI] [PubMed] [Google Scholar]
- 119.Creamer D., Sullivan D., Bicknell R., Barker J. Angiogenesis in psoriasis. Angiogenesis. 2002;5:231–236. doi: 10.1023/a:1024515517623. [DOI] [PubMed] [Google Scholar]
- 120.Sugiyama H., Gyulai R., Toichi E., Garaczi E., Shimada S., Stevens S.R. Dysfunctional blood and target tissue CD4+ CD25 high regulatory T cells in psoriasis: mechanism underlying unrestrained pathogenic effector T cell proliferation. J Immunol. 2005;174:164–173. doi: 10.4049/jimmunol.174.1.164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Peric M., Koglin S., Dombrowski Y., Gross K., Bradac E., Büchau A. Vitamin D analogs differentially control antimicrobial peptide/“Alarmin” expression in psoriasis. PLoS One. 2009;4:e6340. doi: 10.1371/journal.pone.0006340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Mason A., Mason J., Cork M., Hancock H., Dooley G. Topical treatments for chronic plaque psoriasis: an abridged Cochrane Systematic Review. J Am Acad Dermatol. 2013;69:799–807. doi: 10.1016/j.jaad.2013.06.027. [DOI] [PubMed] [Google Scholar]
- 123.Prieto-Pérez R., Cabaleiro T., Daudén E., Ochoa D., Román M., Abad-Santos F. Pharmacogenetics of topical and systemic treatment of psoriasis. Pharmacogenomics. 2013;14:1623–1634. doi: 10.2217/pgs.13.163. [DOI] [PubMed] [Google Scholar]
- 124.Ahn C.S., Awadalla F., Huang K.E., Yentzer B., Dabade T.S., Feldman S.R. Patterns of vitamin d analog use for the treatment of psoriasis. J Drugs Dermatol. 2013;12:906–910. [PubMed] [Google Scholar]
- 125.Mason A.R., Mason J.M., Cork M.J., Hancock H., Dooley G. Topical treatments for chronic plaque psoriasis of the scalp: a systematic review. Br J Dermatol. 2013;169:519–527. doi: 10.1111/bjd.12393. [DOI] [PubMed] [Google Scholar]
- 126.Oquendo M., Abramovits W., Morrell P. Topical vitamin D analogs available to treat psoriasis. Skinmed. 2012;10:356–360. Review. [PubMed] [Google Scholar]
- 127.Agak G.W., Qin M., Nobe J., Kim M.H., Krutzik S.R., Tristan G.R. Propionibacterium acnes induces an IL-17 response in acne vulgaris that is regulated by vitamin A and vitamin D. J Invest Dermatol. 2014;134(2):366–373. doi: 10.1038/jid.2013.334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Lee W.J., Choi Y.H., Sohn M.Y., Lee S.J., Kim do W. Expression of inflammatory biomarkers from cultured sebocytes was influenced by treatment with vitamin D. Indian J Dermatol. 2013;58:327. doi: 10.4103/0019-5154.113959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Ekiz O., Balta I., Sen B.B., Dikilitaş M.C., Ozuğuz P., Rifaioğlu E.N. Vitamin D status in patients with rosacea. Cutan Ocul Toxicol. 2013:28. doi: 10.3109/15569527.2013.797907. [DOI] [PubMed] [Google Scholar]
- 130.Bolland M.J., Ames R.W., Grey A.B., Horne A.M., Mason B.H., Gamble G.D. Does degree of baldness influence vitamin D status? Med J Aust. 2008;189:674–675. doi: 10.5694/j.1326-5377.2008.tb02241.x. [DOI] [PubMed] [Google Scholar]
- 131.Hu L, Bikle DD, Oda Y. Reciprocal role of vitamin D receptor on β-catenin regulated keratinocyte proliferation and differentiation. J Steroid Biochem Mol Biol 2014;144 Pt A:237–41. [DOI] [PMC free article] [PubMed]
- 132.Demay M.B. The hair cycle and vitamin D receptor. Arch Biochem Biophys. 2012;523:19–21. doi: 10.1016/j.abb.2011.10.002. [DOI] [PubMed] [Google Scholar]
- 133.Li Y.C., Amling M., Pirro A.E., Priemel M., Meuse J., Baron R. Normalization of mineral ion homeostasis by dietary means prevents hyperparathyroidism, rickets, and osteomalacia, but not alopecia in vitamin D receptor-ablated mice. Endocrinology. 1998;139:4391–4396. doi: 10.1210/endo.139.10.6262. [DOI] [PubMed] [Google Scholar]
- 134.Teichert A., Elalieh H., Bikle D. Disruption of the hedgehog signaling pathway contributes to the hair follicle cycling deficiency in Vdr knockout mice. J Cell Physiol. 2010;225:482–489. doi: 10.1002/jcp.22227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Rasheed H., Mahgoub D., Hegazy R., El-Komy M., Abdel Hay R., Hamid M.A. Serum ferritin and vitamin D in female hair loss: do they play a role? Skin Pharmacol Physiol. 2013;26:101–107. doi: 10.1159/000346698. [DOI] [PubMed] [Google Scholar]
- 136.van der Vleuten C.J., van de Kerkhof P.C. Management of scalp psoriasis: guidelines for corticosteroid use in combination treatment. Drugs. 2001;61:1593–1598. doi: 10.2165/00003495-200161110-00006. Review. [DOI] [PubMed] [Google Scholar]
- 137.AlGhamdi K., Kumar A. Depigmentation therapies for normal skin in vitiligo universalis. J Eur Acad Dermatol Venereol. 2011;25:749–757. doi: 10.1111/j.1468-3083.2010.03876.x. [DOI] [PubMed] [Google Scholar]
- 138.Birlea S.A., Costin G.E., Norris D.A. Cellular and molecular mechanisms involved in the action of vitamin D analogs targeting vitiligo depigmentation. Curr Drug Targets. 2008;9:345–359. doi: 10.2174/138945008783954970. [DOI] [PubMed] [Google Scholar]
- 139.Alghamdi K., Kumar A., Moussa N. The role of vitamin D in melanogenesis with an emphasis on vitiligo. Indian J Dermatol Venereol Leprol. 2013;79:750–758. doi: 10.4103/0378-6323.120720. [DOI] [PubMed] [Google Scholar]
- 140.Mason R.S., Holliday C.J. 1,25-Dihydroxyvitamin D contributes to photoprotection in skin cells. In: Norman A., Bouillon R., Thomasset M., editors. Vitamin D endocrine system: structural, biological, genetic and clinical aspects. University of California; Riverside: 2000. pp. 605–608. [Google Scholar]
- 141.Huang C.L., Nordlund J.J., Boissy R. Vitiligo: a manifestation of apoptosis? Am J Clin Dermatol. 2002;3:301–308. doi: 10.2165/00128071-200203050-00001. [DOI] [PubMed] [Google Scholar]
- 142.Silverberg J., Silverberg A. A pilot study assessing the role of 25 hydroxyvitamin D levels in patients with vitiligo vulgaris. J Am Acad Dermatol. 2010;626:938–941. doi: 10.1016/j.jaad.2009.11.024. [DOI] [PubMed] [Google Scholar]
- 143.Saleh H.M., Abdel Fattah N.S., Hamza H.T. Evaluation of serum 25-hydroxyvitamin D levels in vitiligo patients with and without autoimmune diseases. Photodermatol Photoimmunol Photomed. 2013;29:34–40. doi: 10.1111/phpp.12016. [DOI] [PubMed] [Google Scholar]
- 144.Xu X., Fu W.W., Wu W.Y. Serum 25-hydroxyvitamin D deficiency in Chinese patients with vitiligo: a case-control study. PLoS One. 2012;7:e52778. doi: 10.1371/journal.pone.0052778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Parsad D., Saini R., Verma N. Combination of PUVAsol and topical calcipotriol in vitiligo. Dermatology. 1998;197:167–170. doi: 10.1159/000017991. [DOI] [PubMed] [Google Scholar]
- 146.Parsad D., Kanwar A.J. Topical vitamin D3 analogues in the treatment of vitiligo. Pigm Cell Melanoma Res. 2009;22:487–488. doi: 10.1111/j.1755-148X.2009.00579.x. [DOI] [PubMed] [Google Scholar]
- 147.Oh S.H., Kim T., Jee H., Do J.E., Lee J.H. Combination treatment of nonsegmental vitiligo with a 308-nm xenon chloride excimer laser and topical high-concentration tacalcitol: a prospective, single-blinded, paired, comparative study. J Am Acad Dermatol. 2011;65:428–430. doi: 10.1016/j.jaad.2010.12.007. [DOI] [PubMed] [Google Scholar]
- 148.Arca E., Taştan H.B., Erbil A.H., Sezer E., Koç E., Kurumlu Z. Narrow-band ultraviolet B as monotherapy and in combination with topical calcipotriol in the treatment of vitiligo. J Dermatol. 2006;33:338–343. doi: 10.1111/j.1346-8138.2006.00079.x. [DOI] [PubMed] [Google Scholar]
- 149.Hartmann A., Lurz C., Hamm H., Bröcker E.B., Hofmann U.B. Narrow-band UVB311 nm vs. broad-band UVB therapy in combination with topical calcipotriol vs. placebo in vitiligo. Int J Dermatol. 2005;44:736–742. doi: 10.1111/j.1365-4632.2004.02154.x. [DOI] [PubMed] [Google Scholar]
- 150.Ada S., Sahin S., Boztepe G., Karaduman A., Kölemen F. No additional effect of topical calcipotriol on narrow-band UVB phototherapy in patients with generalized vitiligo. Photodermatol Photoimmunol Photomed. 2005;21:79–83. doi: 10.1111/j.1600-0781.2005.00139.x. [DOI] [PubMed] [Google Scholar]
- 151.El-Komy MH, Samir N, Shaker OG. Estimation of vitamin D levels in patients with pemphigus vulgaris. J Eur Acad Dermatol Venereol 2014:28(7):859–63. [DOI] [PubMed]
- 152.Marzano A.V., Trevisan V., Eller-Vainicher C., Cairoli E., Marchese L., Morelli V. Evidence for vitamin D deficiency and increased prevalence of fractures in autoimmune bullous skin diseases. Br J Dermatol. 2012;167:688–691. doi: 10.1111/j.1365-2133.2012.10982.x. [DOI] [PubMed] [Google Scholar]
- 153.Mutgi K., Koo J. Update on the role of systemic vitamin d in atopic dermatitis. Pediatr Dermatol. 2013;30:303–307. doi: 10.1111/j.1525-1470.2012.01850.x. [DOI] [PubMed] [Google Scholar]
- 154.Heimbeck I., Wjst M., Apfelbacher C.J. Low vitamin D serum level is inversely associated with eczema in children and adolescents in Germany. Allergy. 2013;68:906–910. doi: 10.1111/all.12167. [DOI] [PubMed] [Google Scholar]
- 155.Aly WW, Hussein MA, Moahamed Ebeid S, Mortagy AK. Prevalence of vitamin D insufficiency among community dwelling elderly in Dakahlia as a representative of rural areas in Egypt. Aging Clin Exp Res 2013 [Epub ahead of print]. [DOI] [PubMed]
- 156.Ragab W.S., Saleh W.F., Tawfik L.T. Vitamin D deficiency in postmenopausal Egyptian women. Int J Gynaecol Obstet. 2013;123:75–76. doi: 10.1016/j.ijgo.2013.04.016. [DOI] [PubMed] [Google Scholar]
- 157.Amr N., Hamid A., Sheta M., Elsedfy H. Vitamin D status in healthy Egyptian adolescent girls. Georgian Med News. 2012;210:65–71. [PubMed] [Google Scholar]
- 158.Brink E.W., Aly H.E., Dakroury A.M., Said A.K., Ghoneme F.M., Hussein M.A. The Egyptian National Nutrition Survey, 1978. Bull World Health Organ. 1983;61:853–860. [PMC free article] [PubMed] [Google Scholar]