

Abstract
As biofilms grow, resident cells inevitably face the challenge of resource limitation. In the opportunistic pathogen Pseudomonas aeruginosa PA14, electron acceptor availability affects matrix production and, as a result, biofilm morphogenesis. The secreted matrix polysaccharide Pel is required for pellicle formation and for colony wrinkling, two activities that promote access to O2. We examined the exploitability and evolvability of Pel production at the air-liquid interface (during pellicle formation) and on solid surfaces (during colony formation). Although Pel contributes to the developmental response to electron acceptor limitation in both biofilm formation regimes, we found variation in the exploitability of its production and necessity for competitive fitness between the two systems. The wild type showed a competitive advantage against a non-Pel-producing mutant in pellicles but no advantage in colonies. Adaptation to the pellicle environment selected for mutants with a competitive advantage against the wild type in pellicles but also caused a severe disadvantage in colonies, even in wrinkled colony centers. Evolution in the colony center produced divergent phenotypes, while adaptation to the colony edge produced mutants with clear competitive advantages against the wild type in this O2-replete niche. In general, the structurally heterogeneous colony environment promoted more diversification than the more homogeneous pellicle. These results suggest that the role of Pel in community structure formation in response to electron acceptor limitation is unique to specific biofilm models and that the facultative control of Pel production is required for PA14 to maintain optimum benefit in different types of communities.
INTRODUCTION
Most bacteria form multicellular communities called biofilms by producing and encasing themselves in matrices of secreted polymers (1). The National Institutes of Health has estimated that more than half of all bacterial infections involve such biofilm formation, a feature that complicates treatment due to a variety of associated mechanisms that confer increased antibiotic resistance and tolerance in these communities (2). Steep chemical gradients that form within cellular aggregates give rise to microenvironmental heterogeneity; consequently, cells in biofilms exist in diverse physiological states, at least some of which are unique to this lifestyle. For example, in the opportunistic pathogen Pseudomonas aeruginosa PA14, some genes expressed specifically in biofilms are critical for the establishment of lung infections in a mouse model (3). A better understanding of the physiology of biofilm development is required for rational approaches to new therapies for many types of bacterial infections.
Using a colony biofilm model, we have found that a primary factor influencing P. aeruginosa PA14 community morphology is the availability of electron acceptors (4–6). As colonies increase in thickness, the formation of O2 gradients renders the community anoxic at depth and leads to an increase in intracellular reducing power. O2-limited P. aeruginosa can use two other electron acceptors, nitrate or endogenously produced redox-active compounds called phenazines, to balance the intracellular redox state (7–9). If these oxidants are not available, the community forms a thin biofilm with pronounced wrinkle structures, which increase the surface-to-volume ratio of the biofilm and improve access to O2. Wrinkle formation in strain PA14 depends on the increased production of a cationic matrix polysaccharide called Pel (10, 11). When grown on media without nitrate, a phenazine- and Pel-null double mutant forms a smooth colony with a reduced intracellular redox state, indicating the importance of Pel-mediated wrinkling for redox balancing in colonies (5). Pel also is required for the formation of pellicles, biofilms that float on standing liquid at the interface with air (10). The overexpression of the pel operon, which encodes the machinery required for Pel polysaccharide production, transport, and processing in several P. aeruginosa isolates (12–16), potentiates both the formation of pellicle biofilms and the rugose morphotype of colony biofilms in P. aeruginosa strain PAK (14). The structures formed by Pel production allow for increased access to air in both of these model systems (5, 6, 17). Together, these findings suggest that Pel-mediated structure formation is regulated in response to electron acceptor availability.
Secreted products such as Pel are intriguing from the perspective of social evolution because they are shared within the community; thus, they may be exploited by nonproducing cells. Because the production of polysaccharides is energetically costly, it is plausible that nonproducing cells in biofilms could gain a fitness advantage by benefitting from the formation of the biofilm matrix without contributing to it (i.e., cheating). Nonetheless, both theoretical and empirical studies suggest that cheaters actually are at a disadvantage in biofilms. Agent-based computer simulations show that by producing extracellular polymeric substances (EPS) like Pel, bacteria build protruding structures that allow them to reach O2-rich regions on top of the biofilm, where they outcompete nonproducers by suffocation (18). This model, which conceptually builds on Hamilton's insights on population viscosity and limited dispersal (19, 20), has received some empirical support from experiments with Vibrio cholerae flow cell biofilms (21). Additional experiments conducted in V. cholerae have revealed a mechanism in which the modification of a matrix-associated protein allows biofilm residents to mediate invasion by nonproducing cells (22, 23). Moreover, whether cheaters in biofilms can gain an advantage by avoiding the cost of matrix production is likely to depend on variables including the density of the cell suspension that initially forms the biofilm and the time scale over which competitive fitness is assessed (24).
Competitive smothering resulting from the production of EPS provides a compelling model to account for the stabilization of this trait. However, an extension to the original agent-based model predicts that, over long time scales or with sufficiently high mutation rates, new mutants lacking the ability to produce EPS could emerge within a biofilm and take over the community (25). Experiments with Pseudomonas fluorescens, a nonpathogenic relative of P. aeruginosa, show that spontaneous nonproducer mutants can grow to predominate within the population, causing pellicle collapse (26). Mutation-driven adaptation is relevant for P. aeruginosa communities, as biofilms grown in drip flow reactors have been shown to give rise to mutants at high rates (27). It is also potentially an issue in the chronic infections that occur in cystic fibrosis patients, where the clonal diversity of P. aeruginosa has been reported (28–30). Such clonal diversity could contribute to the adaptation and persistence of P. aeruginosa infections in these patients by enhancing antibiotic tolerance/resistance or immune system evasion (31).
In this study, we interrogated the relationship between electron acceptor availability and the evolutionary stability of Pel polysaccharide production in biofilms. We found that cells that produce Pel have a competitive advantage over nonproducers in static liquid cultures, where pellicle biofilms form in a gradient of O2 (32) but not in shaken liquid cultures where O2 is homogenously provided. In contrast, we observed that Pel producers and nonproducers are equally competitive in mixed-colony biofilms, suggesting that the roles of Pel in pellicle and colony-wrinkle formation contribute differentially to competitive fitness. Using evolutionary experiments, we demonstrate that matrix production can be selected both for and against in specific biofilm conditions depending on electron acceptor availability. Competition experiments revealed that adaptation to growth in pellicle biofilms leads to a disadvantage in colony biofilms, while adaptation to growth at the colony edge leads to a competitive advantage in this specific colony subregion. Our analysis demonstrates that electron acceptor availability and the production of EPS are evolutionarily concurrent in P. aeruginosa.
MATERIALS AND METHODS
Strains and plasmids.
The clinical isolate Pseudomonas aeruginosa UCBPP-PA14 and derivatives thereof were the focus of this work; all strains and plasmids used in this study are listed in Table 1. Unless otherwise specified, liquid cultures were grown in lysogeny broth (LB) (Miller) at 37°C, with shaking at 250 rpm, while LB solidified with 1.5% agar was used for routine plating and for dilution plating/CFU counting. Static cultures were grown at 37°C without shaking. Specific incubation conditions for individual assays are described in relevant sections below and in figure legends where appropriate. In routine cloning procedures, 15 μg/ml gentamicin was used to select for E. coli transformants, while 100 μg/ml gentamicin was used to select for P. aeruginosa transformants.
TABLE 1.
Strains and plasmids used in this study
| Name | Description | Reference or source |
|---|---|---|
| P. aeruginosa PA14 | ||
| WT | Clinical isolate UCBPP-PA14, wild type | 59 |
| Δphz (LD24) | ΔphzA1-G1 ΔphzA2-G2 deletion mutant | 60 |
| Δpel (LD82) | ΔpelB-G deletion mutant | 5 |
| Δphz Δpel (LD83) | ΔphzA1-G1 ΔphzA2-G2 ΔpelB-G deletion mutant | 5 |
| phz+ (DKN370) | Contains two copies of phzM (pyocyanin overproducer) | 4 |
| pel+ (LD1471) | ΔPpel + PA1/04/03 promoter swap mutant (constitutive Pel producer) | This study |
| eShak (LD1482) | Strain isolated after three passages of shaken culture growth | This study |
| ePel (LD1480) | Strain isolated after three passages of pellicle growth | This study |
| eColcenter (LD2243, LD2246 LD2249, LD2252) | Evolved populations obtained after three passages of colony center growth | This study |
| eColedge (LD2231, LD2234 LD2237, LD2240) | Evolved populations obtained after three passages of colony edge growth | This study |
| eColwhole (LD2255, LD2258, LD2261, LD2264) | Evolved populations obtained after three passages of whole colony growth | This study |
| E. coli | ||
| UQ950 | E. coli DH5α λ(pir) host for cloning; F− Δ(argF-lac)169 φ80dlacZ58ΔM15 glnV44(AS) rfbD1 gyrA96(Nalr) recA1 endA1 spoT1 thi-1 hsdR17 deoR λpir | D. Lies, Caltech |
| BW29427 | thrB1004 pro thi rpsL hsdS lacZΔM15 RP4-1360 Δ(araBAD)567 ΔdapA1341::[erm pir] | W. Metcalf, University of Illinois |
| ß2155 | thrB1004 pro thi strA hsdsS lacZΔM15 (F′ lacZΔM15 lacIq traD36 proA+ proB+) ΔdapA::erm (Ermr) pir::RP4 [::kan (Kmr) from SM10] | 61 |
| S. cerevisiae | ||
| InvSc1 | MATa/MATα leu2/leu2 trp1-289/trp1-289 ura3-52/ura3-52 his3-Δ1/his3-Δ1 | Invitrogen |
| Plasmids | ||
| pMQ30 | sacB CEN/ARSH URA3+ Gmr, yeast-based vector , used for allelic exchange | 33 |
| pAKN69-eYFP | Contains mini-Tn7(Gm)PA1/04/03::eYFP fusion, used to genomically integrate constitutive YFP in PA14 strains | 34 |
Construction of strains constitutively producing Pel or yellow fluorescent protein (YFP).
The pel+ mutant was constructed by first amplifying the regions flanking the native Ppel promoter and the constitutive PA1/04/03 promoter using PCR and the primers listed in Table 2. The flanking regions were connected to the PA1/04/03 promoter in the pMQ30 plasmid by gap repair cloning in the Saccharomyces cerevisiae strain InvSc1 (Invitrogen) (33, 34). The resulting promoter swap plasmid was transformed into the Escherichia coli donor strain BW29427 and mobilized into PA14 via biparental conjugation. The pel+ mutant then was created by two successive recombination events, one in each of the two flanking regions, which replaced the chromosomal Ppel with PA1/04/03. The correct pel+ mutant was selected for by successive screening, first with gentamicin and then with 10% sucrose, and was confirmed by PCR. This method is similar to the markerless deletion method described in Dietrich et al. (5), which was used to make the Δpel and Δphz Δpel strains.
TABLE 2.
Primers used for generation of the pel+ mutant
| Primer | Sequence (5′ to 3′) |
|---|---|
| Upstream region flanking Ppel | |
| 5′ | CCAGGCAAATTCTGTTTTATCAGACCGCTTCTGCGTTCTGATAGCAAGATCGACCTCAAGGA |
| 3′ | AAAAAGGGGACCTCTAGGGTCCGCCAGAATGAAAGAGCC |
| PA1/04/03 | |
| 5′ | GGCTCTTTCATTCTGGCGGACCCTAGAGGTCCCCTTTTT |
| 3′ | CCTTTCTTGCTGAACCGCATGCTTAATTTCTCCTCTTTAATTCTAGATGTG |
| Downstream region flanking Ppel | |
| 5′ | CACATCTAGAATTAAAGAGGAGAAATTAAGCATGCGGTTCAGCAAGAAAGG |
| 3′ | GGAATTGTGAGCGGATAACAATTTCACACAGGAAACAGCTGCAGGTAGTCGACCCGATAA |
For many of the PA14 strains used in this study, corresponding tagged strains were generated that produced YFP constitutively so that different strains could be distinguished in coculture experiments. This was done using the pAKN69-eYFP plasmid (34), which was mobilized into PA14 via triparental conjugation (donor strain, BW29427; helper strain, ß2155).
Colony morphology assay.
Ten microliters of overnight precultures were spotted on 9-cm by 9-cm by 1.5-cm square plates containing 60 ml colony morphology assay medium (1% tryptone; Teknova) with 40 μg/ml Congo red (CR) and 20 μg/ml Coomassie brilliant blue dyes (EMB), solidified with 1% agar (Teknova). Colonies were grown at room temperature (23 to 25°C). Images of the colonies were taken daily using a Keyence VHX-1000 digital microscope or a CanoScan 5600F scanner.
For the colony morphology assay at various O2 concentrations, plates were grown at room temperature in C-Chamber hypoxia chambers (BioSpherix). Pure O2 and N2 gasses were mixed to achieve the desired concentrations, regulated by a ProOx P110 O2 controller (BioSpherix).
Quantitative Congo red binding assay.
The relative amount of matrix present in resuspended pellicles or colonies was assessed using a Congo red binding assay. Congo red was added at a final concentration of 20 μg/ml in 0.5 ml of 1% tryptone (Fig. 1D) or 40 μg/ml in 1.5 ml of PBS (see Fig. 7D) and incubated with shaking for 2 h at 37°C. After this incubation, the cells were pelleted via centrifugation, and the absorbance (490 nm) of the supernatant was measured. One percent tryptone plus 20 μg/ml Congo red (Fig. 1D) or PBS plus 40 μg/ml CR (see Fig. 7D) was used as the reference to determine the amount of CR that was removed from the solution (i.e., bound to sample) (35, 36).
FIG 1.

Electron acceptor availability, Pel production, and biofilm architecture are linked in P. aeruginosa PA14. (A) Reduced availability of O2 and/or phenazines induced a red and wrinkled colony morphology, while colonies grown with elevated availability of these electron acceptors were smoother and exhibited less Congo red (CR) staining. Δphz, phenazine-deficient mutant; phz+, phenazine overproducer. (B) A Δpel mutant formed smooth and pale colonies (with less CR bound), while pel+, a constitutive Pel producer, formed highly wrinkled, dark red colonies. Colonies shown are 5 days old and were grown on colony morphology assay medium. Colonies were grown at 21% O2 unless otherwise stated. (C) Pellicles formed in static monocultures. Overnight precultures were diluted to an optical density (OD) at 500 nm of 0.5 and then diluted 1:100 in LB medium. These new cultures were grown statically (in 18-mm by 150-mm glass tubes) for 2 days at 37°C. The pellicle and planktonic fractions have been highlighted. (D) CR binding by pellicles grown as described for panel C. CR binding was calculated as the difference between absorbance for reference solution containing the starting concentration of CR and the absorbance of the sample. (E) CFU counts of both the pellicle and planktonic fractions of WT, Δphz, Δpel, and Δphz Δpel strains. Fewer CFU were found in the planktonic fractions of Pel producers (WT and Δpel strains) than non-Pel producers (Δpel and Δphz Δpel strains) (P = 1.6 × 10−4). More total CFU were observed in cultures of Pel producers (error bars denote ± standard errors of the means [SEM]; n = 3; 2-tailed heteroscedastic student t test).
FIG 7.

Experimental evolution in the O2-limited (center) and O2-replete (edge) regions of colony biofilms gives rise to mutants with diverse Pel production phenotypes. (A) O2 profiles taken in the Pel-producing and non-Pel-producing regions (as indicated in the photo, inset) of a WT colony on day 6 (n = 5; error bars denote SD). O2 was measured with a Clark-type microelectrode inserted at the top of the colony, taking readings every 5 μm as the electrode tip moved toward the agar medium. (B) Cartoon showing the experimental evolution regimes used to generate the eColwhole, eColcenter, and eColedge lineages. (C) Representative scans of colony biofilms grown from the eColwhole, eColcenter, and eColedge lineages. For each passaging regime, scans are shown for 4 replicates after the 1st, 2nd, and 3rd passages. Scans were taken after 3 days of growth. (D) Pel production, quantified using the CR binding assay, for colonies formed by WT, Δpel, and eColedge strains. n = 4, and error bars represent SD. P values were calculated using unpaired, 2-tailed t tests.
Competition assays: liquid cultures and colonies.
Each competition assay was conducted by coculturing one strain constitutively expressing YFP (the tagged strain) with one untagged strain. Controls with the opposite tagged/untagged strain combination, as well as controls competing with the tagged and untagged versions of the same strain, were carried out simultaneously.
Overnight precultures were diluted 1:100 in fresh LB and grown for 2.5 to 3 h to reach an optical density at 500 nm (OD500) of 0.5 to 0.7. Each culture was diluted to an OD500 of 0.5, and the two strains were mixed at the desired ratio (1:1, unless otherwise specified). Fifty microliters of this mix then was diluted again into two new 5-ml cocultures. One of these cultures was grown with shaking for 20 h at 37°C, while the other was grown statically for 48 h at 37°C in 18- by 150-mm glass tubes.
Colony counts were used to determine the ratio of the YFP-tagged strain to the non-YFP-tagged strain following coculture. YFP-expressing colonies were identified using a Typhoon Trio fluorescence scanner (excitation, 473-nm laser; emission, 520-nm filter; GE). For static cultures, the pellicle that formed on the surface of the culture was aspirated and resuspended in 1 ml of 1% tryptone. Colonies were sampled using a flame-sterilized razor blade and a 1,000-μl pipet tip before resuspension in 1 ml of 1% tryptone. Resuspensions were homogenized using a BeadBug (Benchmark Scientific) for a total of 5 min (10 30-s pulses; samples were kept on ice between pulses) at 400 rpm before dilution and plating. One to two hundred microliters each of 10−6 and 10−7 dilutions of biofilm resuspensions or shaken-culture samples was spread on LB agar plates and grown overnight at 37°C.
Pellicles were imaged using a Zeiss Axio Zoom.V16 microscope (excitation, 488 nm; emission, 509 nm) fitted with an ApoTome.2 slider module (exposure time, 50 ms per slice; slice thickness, 10 μm).
For colony biofilm competition assays, the cocultures were prepared as described above, but the coculture mixes were spotted directly onto plates and grown as in the colony morphology assay. YFP images of the colony biofilms were taken using a Typhoon Trio scanner (GE Life Sciences) unless otherwise noted.
Evolution experiments: liquid cultures.
Five milliliters of LB in a glass tube (18 by 150 mm) was inoculated with 50 μl of an overnight preculture. After 2 days of static incubation at 37°C, the pellicle from this culture was aspirated, washed with phosphate-buffered saline, and resuspended in 1 ml LB. Fifty microliters of this suspension was used to inoculate a new 5-ml static culture, and the pellicle collection and transferring process was repeated for a total of three passages. After the third passage, the resuspended pellicle was streaked on an LB plate and an isolated colony was picked at random to represent the “evolved in pellicle” (ePel) strain.
For the corresponding control experiment, the initial inoculation was the same as that described above, but the culture was incubated at 37°C with shaking at 250 rpm for 1 day before 50 μl was sampled and transferred into 5 ml of fresh LB. This process was repeated for a total of three passages. After the third passage, the culture was sampled and streaked on an LB plate. An isolated colony was picked at random to represent the “evolved in shaken culture” (eShak) control.
Microelectrode measurements.
Profiles of O2 concentrations were generated using a miniaturized Clark-type O2 sensor (10-μm tip diameter; Unisense) connected to a picoampere amplifier multimeter (Unisense) and polarized with −0.8 V. Calibration was performed as previously described (5). For each of the two regions (Pel producing and non-Pel producing), 5 O2 profiles were generated from 3 different WT colony biofilms.
Evolution experiments: colonies.
Colonies were grown as described for the colony morphology assay described above. After 14 days of incubation, biofilm growth from either the edge, the center, or the entire colony was collected and resuspended in 50 μl of LB. Ten microliters of this was spotted and grown into a new colony biofilm. This was repeated for a total of three passages. After the third passage, a final sample was taken in a manner consistent with the passaging protocol, resuspended, and preserved as a population. Individual replicates of this process are referred to as eColedge (for “evolved at the colony edge”), eColcenter, or eColwhole lineage.
RESULTS
P. aeruginosa PA14 communities enhance colony wrinkling and pellicle formation by producing more Pel polysaccharide under electron acceptor limitation.
Previous studies examining P. aeruginosa colony biofilm development have demonstrated a correlation between electron acceptor limitation and colony wrinkling. They employed a colony morphology assay in which colony biofilms are grown on a medium containing the EPS-binding dye Congo red (CR). A Δphz mutant unable to produce phenazines wrinkles earlier and to a greater extent over the course of development. Conversely, a phz+ mutant that overproduces one of the phenazines forms a smoother colony that exhibits less CR binding than the wild type (WT) (Fig. 1A) (4). Growth on media containing nitrate, another electron acceptor for P. aeruginosa, also leads to less EPS production and a smoother colony morphology (5). Therefore, we have proposed that colony wrinkling is a strategic response to electron acceptor limitation that enhances access to O2 (5, 37). To confirm the link between colony wrinkling and EPS production, we constructed a P. aeruginosa PA14 mutant that constitutively expresses the pel operon (the pel+ mutant). Colonies of this mutant show hyperwrinkled structures, as expected (Fig. 1B). Taken together, these results reveal a trend in which the limited availability of exogenously supplied or endogenously produced electron acceptors leads to colony wrinkling and suggest that Pel upregulation is required for this process (Fig. 1A and B) (4, 5).
We further investigated the relationship between electron acceptor availability, Pel production, and biofilm development using a static liquid culture assay in which a pellicle biofilm forms at the air-liquid interface (10). This assay would be expected to establish a more uniform O2 limitation pattern than the colony morphology assay, as cells at the periphery are constrained by the wall of the culture vessel. We assessed Pel production in pellicle biofilms of P. aeruginosa PA14 WT and the Δphz mutant using a CR binding assay (35, 36). The Δphz mutant formed a thicker pellicle than the WT and exhibited higher CR binding levels, indicating increased production of EPS (Fig. 1C and D; also see Fig. S1 in the supplemental material). These observations are consistent with our observations in colony biofilms (5) and suggest that EPS production is upregulated in the context of electron acceptor limitation in both pellicles and colonies. As expected, mutants lacking the pel genes (the Δpel and Δphz Δpel mutants) did not form pellicles in static cultures (Fig. 1C and D) (10).
Pel production is beneficial to the static liquid culture community under electron acceptor limitation.
We next assessed whether pellicle formation is advantageous for a P. aeruginosa population. We collected the pellicle and planktonic fractions of static cultures, plated for CFU, and found that pellicle-forming cultures contain more total CFU than cultures of mutants unable to produce pellicles (Fig. 1E). This difference can be attributed to the high numbers of CFU observed specifically in the pellicle fractions of these cultures. Comparing the planktonic fractions, significantly fewer CFU were found in those of Pel producers than in those of non-Pel producers, suggesting that the pellicle biofilm consumes O2 that otherwise would have been available to the planktonic cells and limits growth in this region. This effect is consistent with experiments in P. fluorescens, where cells in static culture fully deplete O2 at 1 cm from the air-liquid interface within hours of inoculation (32) and the development of a pellicle inhibits growth in the underlying liquid medium (26). In addition, more cells were found in the pellicle fraction of the Δphz mutant than in that of the WT, indicating that higher levels of EPS production are linked to greater population growth in the pellicle itself (Fig. 1E; also see Fig. S1 in the supplemental material). Indeed, this suggests that the Δphz mutant compensates for its defect in phenazine production and, as a result, lower electron acceptor levels by producing more Pel and enhancing Pel-dependent structure (pellicle) formation in static liquid culture much in the same way as that shown for growth on solid surfaces (Pel-dependent colony wrinkle formation). To further verify the effects of electron acceptor availability on pellicle development, we grew PA14 static cultures using media amended with nitrate, an alternative terminal electron acceptor for P. aeruginosa, and found that it inhibited pellicle formation (see Fig. S1). Taken together, these results show that there is a fitness advantage to forming a pellicle biofilm in the context of electron acceptor limitation and support the model that electron acceptor limitation leads to higher Pel production (26, 32).
Pel confers a direct benefit to producers in pellicle biofilms.
As a secreted product, Pel has the potential to act as a shared resource that benefits both individuals responsible for its production as well as cheaters, individuals that do not contribute to EPS production and avoid the associated energy cost but nevertheless reside in the pellicle and benefit from increased access to O2. However, theoretical modeling predicts that the synthesis of EPS can directly benefit producing lineages by repositioning these cells and allowing them to smother nonproducing neighbors (18), and experiments in other bacterial species suggest that extracellular polymers are not public goods at all (22, 23). To test this possibility in P. aeruginosa biofilms, we examined whether mutants unable to produce Pel could exploit and take up residence in a pellicle produced by WT cells. We competed the WT and the Δphz mutant against their corresponding Δpel mutants in both pellicle biofilms and shaken cultures. In contrast to pellicle biofilms, which contain O2 gradients, shaken cultures have high, homogenous O2 availability throughout the population. The competitions were inoculated with a mixed population containing equal numbers of the two strains being tested. CFU counts at the end of the competition were used to calculate a final ratio between the strains. In order to distinguish the two strains during colony counts, one of the strains was tagged with constitutively expressed YFP. To control for the effects of YFP labeling, experiments were repeated with the other strain tagged (reverse tagging), and each strain was competed against its corresponding tagged version.
In shaken cultures, the ratio between the Pel-producing strains and the non-Pel-producing strains remained approximately 1:1 for all combinations tested (Fig. 2A). This indicates that in the presence of abundant electron acceptor (O2), the ability to produce Pel does not confer a competitive advantage. However, in static cultures (where O2 is limited), Pel-producing strains strongly outcompeted non-Pel-producing strains in both phenazine-producing and nonproducing backgrounds (Fig. 2B). Reverse tagging experiments yielded consistent results, and controls in which a YFP-labeled strain was competed against its unlabeled counterpart showed no significant effect of YFP expression under the conditions used for this experiment. Final total CFU counts obtained from shaken cultures were higher than those obtained from pellicles, indicating that the competitive difference observed in pellicles cannot be attributed to a higher number of generations achieved in this mode of growth (see Fig. S2A in the supplemental material). The fluorescence intensities of these pellicles (Fig. 2C) correlated well with the CFU-based quantification of relative strain success, and the differences in strain success did not arise from fundamental growth defects (see Fig. S2B in the supplemental material). Thus, we conclude that Pel confers a producer-specific fitness advantage in static culture, where it enhances access to O2.
FIG 2.

Pel production is advantageous in competition with non-Pel producers but only under electron acceptor limitation. (Top) Final percentages of each strain in shaken (A) and static (pellicle) (B) cocultures. All cocultures were inoculated with an initial ratio of 1:1 (n = 3; error bars denote standard deviations [SD]; P values are from two-tailed unpaired equal-variance t test). (C) Imaging of coculture pellicles reveals variations in fluorescence intensity consistent with relative strain success, as indicated by the CFU counts for static cultures reported in panel B. Pellicles were imaged using a Zeiss AXIO Zoom.V16 microscope fitted with an ApoTome.2 module (excitation, 488 nm; emission, 509 nm).
In the Pel producer versus non-Pel-producer pellicle competition experiments inoculated with 1:1 mixing ratios, there was no successful cheating by non-Pel-producing cells in either WT or Δphz mutant backgrounds (Fig. 2B). However, all of the cultures for these experiments were inoculated with 1:1 mixing ratios. It is possible that the outcome of such a competition would change at different initial mixing ratios, since social traits such as cheating often have frequency-dependent selection (38). To investigate this, we carried out WT-Δpel strain competitions using a range of initial mixing ratios and confirmed that the WT (Pel-producing) strain retains its advantage over the Δpel strain regardless of its starting frequency (Fig. 3A). We found that the final Δpel strain percentage was substantially lower than the initial percentage for all mixing ratios tested, indicating a strong selection for Pel production. Total pellicle CFU counts (for WT and Δpel strains combined) were roughly constant across the range of starting Δpel strain percentages, suggesting that the Pel-producing strain divides until it reaches a plateau in population size (a fixed carrying capacity) regardless of the number of cells initially present (Fig. 3B). These results demonstrate that in P. aeruginosa PA14 pellicle biofilms, selection for Pel production is stable irrespective of the initial frequency of producers.
FIG 3.

Δpel strain shows a competitive disadvantage against the WT in pellicles formed on static cultures regardless of the initial coculture mixing ratio. (A) WT and Δpel strains were mixed at different initial ratios and used to start static cultures incubated under the same conditions as those used for results shown in Fig. 2. The initial percentage of the Δpel strain is plotted against its final percentage in the pellicle. Each data point represents an individual coculture, with the YFP tag on either the WT or the Δpel strain. (B) Pellicles reach a generally consistent final population size regardless of the initial mixing ratio of WT and Δpel strains. The initial percentage of the Δpel strain is plotted against the final total CFU (WT plus Δpel strains) in the pellicle.
Pel does not confer a strong competitive advantage to producers in colony biofilms.
Colonies develop O2 gradients due primarily to the consumption of O2 by cells closest to the periphery (5, 39). Pel production in colonies enables the formation of wrinkle structures that increase the colony surface area-to-volume ratio, improving access to O2 for resident cells. We again took advantage of constitutive YFP expression to distinguish between strains and grew colonies from mixtures of WT and Δpel precultures. We expected the Pel-producing cells to possess an increased ability to form wrinkle structures and, as a result, to exhibit increased success in colonies relative to nonproducing mutant cells, as in the pellicle competition experiments described above. The earlier and more intricate wrinkling of the colony center suggests a role for Pel in this region specifically, so we took separate samples from the center and the edge (Fig. 4A). We found that sampling from the colony edge produced data with greater variability than sampling from the colony center (Fig. 4B). This also was reflected in fluorescence scans of colony cocultures and may be due to range expansion (40) (see Fig. S3 in the supplemental material). The large variability in the final frequencies of WT and Δpel strains do not allow us to conclude that either strain has the advantage. Nonetheless, the competition result differs significantly from that obtained in the pellicle model, where WT clearly wins the competition (Fig. 4B; also see Fig. S3 in the supplemental material). We conclude that the exploitability of mechanisms whereby Pel contributes to structure formation differs between the pellicle and colony environments.
FIG 4.

Ability to produce Pel does not provide a competitive advantage against Δpel strains in mixed-colony biofilms. Samples were obtained from the colony center or edge (A) after 5 days of growth on colony morphology assay medium and plated for CFU (B). n = 3. Error bars represent SD. P values are from two-tailed unpaired equal-variance t test.
A recent study using V. cholerae showed that the adhesive property of matrix can cause producing cells to become fixed to the bottom of a biofilm and displace nonproducers (41). We prepared thin sections of mixed-colony biofilms containing WT and Δpel strains in which one strain was tagged with YFP. Fluorescence microscopy did not reveal an apparent accumulation of matrix producers at the base of the biofilm (see Fig. S3).
Repeated rounds of growth in pellicles select for Pel overproducers that show a competitive advantage against the wild type.
Pel production is involved in the responses of both static cultures and colony biofilms to electron acceptor limitation (Fig. 1 and 2). Therefore, we wondered whether repeated culturing in these biofilm systems would promote the evolution of strains with altered Pel production phenotypes that affected fitness in each system. First, we hypothesized that repeated culturing under pellicle formation conditions would give rise to mutants that overproduced Pel, which would confer an advantage in pellicle cocultures.
To test this, we carried out repeated rounds of growth in pellicles. We started by growing a monoclonal population of the WT in static culture for 2 days; we then harvested, washed, and resuspended the pellicle. This suspension was used to inoculate fresh medium for the next round of pellicle growth. This process was repeated for a total of three passages (Fig. 5A). As a control, we conducted a second evolutionary experiment with three passages in shaken culture. Clones were isolated at the end of the third passage in each case and evaluated for changes in Pel production using the colony morphology assay.
FIG 5.

Experimental evolution in pellicles selects for strains that show an advantage in competitions for pellicle residence and in shaken liquid cultures. (A) Cartoon showing the experimental evolution regimes used to generate the ePel strain and the shaken-culture control. Representative scans of eShak, WT, and ePel colonies after 48 h of growth are shown. The ePel strain then was evaluated for its competitive fitness against the WT in pellicles of static cultures (B) and shaken cultures (C). n = 6 for static cultures, n = 3 for shaken cultures. Error bars represent SD. P values were calculated using unpaired, 2-tailed t tests.
Repeated growth in shaken cultures did not alter the biofilm morphotypes of isolates compared to those of the parent strain. In contrast, growth in pellicles promoted the emergence of mutants that formed hyperwrinkled colony biofilms (Fig. 5A). We refer to the representative isolate from this experimental evolution regime as ePel (for evolved in pellicle) and the isolate from shaken culture conditions as eShak (for evolved in shaken culture). As ePel is phenotypically similar to small-colony variant (SCV) and wrinkly spreader isolates previously described in studies examining the evolution of other Pseudomonas species and strains in pellicles and biofilms grown in tube reactors (42, 43), we repeated the selection protocol to examine the dynamics of ePel-type colony evolution. The ePel phenotype arose after just one round of pellicle growth and in consistent proportions after each passage (see Fig. S4A in the supplemental material). The CR binding assay confirmed that the ePel strain produced more Pel than the WT, and that this increase was particularly strong in pellicles (see Fig. S4B in the supplemental material). This finding indicates that the pellicle biofilm environment exerts a strong selective pressure for Pel production.
To test whether the evolved Pel overproduction phenotype conferred an advantage in pellicles, we generated a YFP-tagged version of the ePel strain and conducted competition assays. We found that pellicles gave rise to data with more variability than those of shaken cultures in competition assays and attribute this to the expansion of SCVs, which propagate more rapidly than WT cells and can spontaneously arise in labeled or unlabeled strains. Still, our results clearly show that ePel outcompeted its WT parent for residence in pellicles (Fig. 5B), consistent with our findings (described above) demonstrating an advantage conferred by Pel production in pellicles (Fig. 2B). Notably, we also found that ePel showed a selective advantage in shaken cultures, although it was not as pronounced as that in static cultures (Fig. 5C). This advantage was not due to an inherent ability to grow faster than the WT, as the two strains exhibited similar growth dynamics in monocultures (see Fig. S4C in the supplemental material). Whether Pel itself or a trait that is coregulated with Pel production is the factor that gives Pel producers an advantage over nonproducers remains to be investigated.
Pellicle adaptation is detrimental to competitive fitness in colony biofilms.
We next asked if adaptation to the pellicle biofilm environment also conferred an advantage in other biofilm environments. We have observed that Pel production is critical to the formation of wrinkle structures in the colony morphology assay and is associated with electron acceptor limitation. Therefore, we suspected that the evolutionary history of the ePel strain would enhance its competitive fitness in colony biofilms. We prepared mixed suspensions of YFP-tagged and untagged versions of the ePel strain and the WT and used these to inoculate colony morphology assay plates. As with colony competition experiments between WT and Δpel strains, we observed more variability in results for samples taken from the colony edge than for those taken from the colony center. We also observed deviations from the 1:1 ratio for the ePel labeling control, possibly due to the expansion of SCVs. Nevertheless, the results showed a strong competitive disadvantage for the ePel strain relative to that of the WT, both at the colony center and at the colony edge (Fig. 6; also see Fig. S5 in the supplemental material). Although the ePel strain showed Pel overproduction in colony biofilms and pellicles, the observed increase in Pel production was much greater in pellicles, indicating that this strain functions differently in the two environments (see Fig. S4B in the supplemental material). In addition, the ePel strain showed severe defects in swarming and twitching (see Fig. S4D in the supplemental material), which may have contributed to its compact colony phenotype. However, these defects are insufficient to explain the disadvantage in colony competitions against the WT, as this was apparent both at the colony edge and the colony center (where spreading does not occur). These findings further support the notion that cells in pellicle and colony biofilms, despite exposure to apparently similar O2 limitation conditions, require different adaptations for success in the presence of such stress.
FIG 6.

ePel strain exhibits a competitive disadvantage in colony biofilms, both in the colony center (A) and at the colony edge (B). Colonies were grown for 7 days before sampling. n = 3. Error bars represent SD.
Repeated rounds of growth in different subregions of colony biofilms yield diverse Pel production phenotypes.
Repeated growth in the pellicle biofilm system gave rise to a mutant that produced higher levels of Pel and that exhibited an advantage over the wild type specifically in this type of biofilm. We hypothesized that conditions in colony biofilms also would promote diversification in Pel production phenotypes. We reasoned that success in specific colony biofilm subregions (microenvironments) might depend on the balance between the cost (resources used in Pel synthesis) and benefit (closer proximity to the colony-air interface) of Pel production. This in turn would depend on electron acceptor availability, since the need for O2 access is greater under electron acceptor limitation. Using Clark-type microelectrodes, we measured O2 concentrations at the edge (in the area of lower Pel production, indicated by CR staining) and away from the edge toward the center (in the area of higher Pel production/CR staining) of the colony (see Fig. S6 in the supplemental material). In the Pel-producing (center) region of the colony, O2 became undetectable at 80 μm below the colony surface. In the region with little or no Pel production (at the colony edge), however, O2 levels initially decreased but reached a plateau at approximately 40 μm below the colony surface, and full O2 depletion was not detected at any point (Fig. 7A). We note that these profiles do not indicate where the electrode passes from the colony into the underlying agar. Nevertheless, these measurements indicate that, in contrast to the Pel-producing regions closer to the center of the colony, the edge of the colony constitutes a more electron acceptor-rich region where, according to our model, increased Pel production would not be specifically favored.
To ask whether repeated growth in specific colony subregions would promote the evolution of strains with altered Pel production, we conducted evolution experiments starting with a WT colony grown using the standard colony morphology assay protocol. Three different sampling regimes were followed, involving either collection and resuspension of the entire colony, excision and resuspension of the colony center, or excision and resuspension of the colony edge (Fig. 7B). Ten microliters of each sample was used as the inoculum for a new colony, and each passage was repeated three times. Representative images of colony morphologies for lineages transferred under each sampling regime are shown in Fig. 7C. Colonies that grew after the last transfer were collected and saved as whole populations, and these strains are referred to as eColwhole, eColcenter, and eColedge, respectively. eColcenter lineages yielded colony biofilms with diverse phenotypes: some were similar to the ancestral strain, while others appeared somewhat hyperwrinkled and bound more CR than the WT. eColwhole lineages also yielded colony biofilms with diverse phenotypes, showing even greater variability than eColcenter lineages. eColedge lineages consistently formed colonies that were smoother and bound less CR than the WT (see Fig. S7 in the supplemental material). Using the quantitative CR binding assay, we found that EPS production by eColedge strain colonies was comparable to that of the Δpel mutant (Fig. 7D). Overall, lineages obtained from colony evolution experiments exhibited a greater diversity of phenotypes, both between and within specific colony regions, than lineages obtained from pellicle evolution experiments. This difference in the extent of lineage diversification could stem from the more complex structure and more heterogeneous O2 availability of colonies compared to pellicles.
Adaptation to the colony edge environment confers a competitive advantage in this region.
Although other colony regions gave rise to mutants with various phenotypes, the eColedge lineages consistently showed decreased Pel production, as indicated by colony morphology and reduced CR staining (Fig. 7C and D; see Fig. S7 in the supplemental material). This suggested strong selection for decreased Pel production at the colony periphery. We performed competition experiments to test whether the evolved traits, including Pel underproduction, conferred an advantage in this zone. These experiments could be conducted only with the YFP-tagged version of the WT and untagged eColedge populations, because the eColedge lineages are not clonal. We observed that eColedge strains excluded the WT from the colony edge, demonstrating a clear competitive advantage for eColedge lineages in this region (Fig. 8; also see Fig. S8A). Interestingly, when two different lineages were cocultured with the WT in shaken and pellicle-forming cultures, they differed in their competitive fitness relative to that of the ancestral strain (see Fig. S8B and C in the supplemental material) despite showing similar abilities to exclude the WT from the colony edge in colony biofilm competitions.
FIG 8.
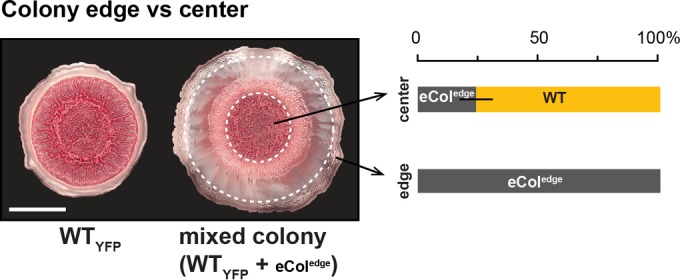
eColedge lineages exclude the ancestral strain from the colony edge. Colonies shown at the left were grown for 7 days on colony morphology assay medium. Sampling was performed as diagrammed in Fig. 4A, and the percentage of each strain represented by CFU is shown at the right. Experiments performed with the eColedge-1 and eColedge-2 lineages yielded similar results; results for eColedge-1 are shown. n = 3. Error bars represent SD. The scale bar is 1 cm.
The advantages of the eColedge lineages over the WT strain at the colony edge are somewhat surprising given that the Δpel strain did not show a competitive advantage in this region. Considering that Pel levels in the eColedge lineages were comparable to those of the Δpel strain, we might expect these strains to have similar competitive fitness outcomes (Fig. 7D). Thus, this discrepancy suggests that either the specific regulation of Pel production in the eColedge lineages or other phenotypic changes in these lineages contribute to their success at the colony edge.
Strains evolved in biofilm environments show additional phenotypic changes.
To more fully understand the evolved strains used in this study, we asked whether they showed alterations in other phenotypic traits associated with biofilm formation. First, we tested whether differences in motility could account for differences in the competitive fitness of the evolved strains. In the case of the eColedge strains, we suspected that increased motility allows these strains to reach the colony edge faster than the WT and contribute to their ability to exclude the WT from this region. The eColedge strains did not show increased swarming or twitching motility relative to that of the WT, consistent with the phenotype of the Δpel mutant (see Fig. S9A and B in the supplemental material).
However, in contrast to the Δpel mutant (see Fig. S9C and D in the supplemental material), all of the eColedge lineages did show increased colony spreading relative to that of the WT when grown on colony morphology assay medium (see Fig. S7 and S9D in the supplemental material). It has been suggested that Pel production confers adhesiveness that leads to cellular immobilization and the formation of more compact communities. In this context, the increased spreading exhibited by the eColedge strains could be interpreted as a consequence of decreased Pel production. However, the fact that the Δpel mutant does not exhibit increased spreading indicates that decreased matrix-dependent adhesiveness is not the sole determinant of spreading ability. Although the cause of enhanced colony spreading in the eColedge lineages is unclear, it may confer a competitive advantage for eColedge lineages in the colony edge region.
We additionally examined phenazine production in the evolved strains. We have observed that phenazine production, which can relieve electron acceptor limitation, leads to decreased Pel production in both pellicle and colony biofilms. Therefore, one might expect an inverse correlation between changes in phenazine production and changes in Pel production in the evolved strains. However, we did not observe such a trend. When grown in shaken liquid cultures, the ePel isolate produced more of the phenazine pyocyanin per OD unit than the WT, while the eColedge strains varied in their pyocyanin production phenotypes (see Fig. S10A in the supplemental material). Consistent results were observed in colony biofilms: the intensity of the pigmentation in the agar after these strains were grown as colonies on a 1% tryptone, 1% agar plate correlated with the pyocyanin quantification from liquid cultures, suggesting that phenazine production phenotypes for shaken liquid cultures corresponded to those for the colony biofilm mode of growth (see Fig. S10B in the supplemental material). Together, these results indicate that the altered Pel production phenotypes of the evolved strains cannot be attributed solely to changes in phenazine production and its effects on redox homeostasis.
DISCUSSION
Historically, many different types of culture regimes have been used as laboratory models for biofilm formation (44), but these models often represent very distinct environmental conditions. A universal challenge faced by cells in communities is the development of chemical gradients that lead to resource limitation for individuals at depth. The production of polysaccharides has been shown to be important for development and morphogenesis in pellicles and colonies formed by diverse microbes (13, 45–48). Following a decrease in electron acceptor availability, P. aeruginosa PA14 increases the production of the polysaccharide Pel, which is required for pellicle formation and colony wrinkling (5, 10). In both of these systems, Pel has the effect of increasing O2 availability, although the physical basis of this effect differs in the two regimes. In colonies, increased access to O2 results from the increased colony surface area of a wrinkled morphology compared to a smooth one, which exposes a higher proportion of resident cells to the atmosphere (49). In pellicles, increased access to O2 is achieved because the formation of the floating biofilm retains cells at the air-liquid interface in physical proximity to the atmosphere. We acknowledge that pellicles can wrinkle at later time points, mimicking the morphologies seen in colonies. However, in this study we focused on the early stages of pellicle formation.
As electron acceptor limitation stimulates Pel production and the effects of Pel enhance access to O2 in both pellicle and colony biofilms, we predicted that the exploitability of Pel production and any advantage Pel production confers would be consistent between these models. However, we found that nonproducing cells were at a severe disadvantage in competition with the WT in pellicles but at no disadvantage against the WT in colonies. This suggests that Pel contributes differentially to fitness in pellicle and colony biofilms. Differences in the physical characteristics of the two environments may affect the mechanisms by which extracellular matrix production enhances access to electron acceptors. A steep O2 gradient arises relatively early in the static incubation of a liquid culture, before pellicle formation is observed, while the development of an O2 gradient in a colony occurs only after significant biofilm growth (32). Therefore, the different abilities of the Δpel mutant to thrive in coculture with the WT in the respective systems may arise from differences in the timing and localization of O2 gradient development, the relative proximity of cells to each other, structural differences between pellicles and colony biofilms, or other subtle differences between the growth regimes. We found that adaptation to the pellicle environment conferred an advantage against the WT parent in pellicle cocultures but a pronounced disadvantage against the WT in mixed colonies, further underscoring the differences between the two types of biofilms (Fig. 5B and C and 6).
We note that the inability of the Δpel strain to cheat in static cocultures with the WT appears to contradict results obtained by Rainey and Rainey demonstrating the ability of a smooth-colony-forming strain to cheat at some time points in prolonged static coculture experiments with a wrinkly colony-forming strain. However, the smooth strain studied by Rainey and Rainey was obtained through evolution in static culture, in contrast to our engineered Δpel strain; therefore, it may have been adapted to cheat in this setting (26).
Distinct levels of CR staining at the center and edge of P. aeruginosa PA14 biofilms suggested that Pel production and its associated benefits would differ in these subregions. According to our microelectrode measurements, these regions are characterized by low and high O2 availability, respectively (Fig. 7A). We predicted that repeated growth in the colony center would select for increased Pel production. However, although some strains that evolved in the colony center showed this phenotype, it was not consistently obtained (Fig. 7; see Fig. S7 in the supplemental material). Structural and sampling differences between pellicle and colony biofilm evolution regimes may provide some insight into this discrepancy. In static liquid cultures, only the cells with significant Pel production take up residence in the pellicle, and only the pellicle was sampled for transferring in the evolution experiments. In colonies, cells with significant Pel production may elevate themselves to higher portions of the wrinkle, leaving cells with lower levels of Pel production in the base; however, in colony evolution experiments, both the wrinkle and the base were sampled for transferring. Therefore, the relatively heterogeneous selective pressures of colonies compared to those of pellicles could explain their decreased potential to select for Pel-overproducing strains.
Recent findings by Kim et al. indicate that polysaccharide production provides a selective advantage in colony biofilms of a nonpathogenic relative of P. aeruginosa, the soil isolate P. fluorescens Pf0-1 (50). Within-biofilm evolution of this strain leads to mutations in the gene for the global repressor protein RsmE and gave rise to a mucoid phenotype. In diverse pseudomonad species, mucoidy is caused by the increased production of extracellular polysaccharides such as alginate (51, 52). Competition experiments by Kim et al. revealed a strong selective advantage for the mucoid phenotype during growth in colony biofilms. These results differ from our observations in P. aeruginosa PA14, where increased Pel polysaccharide production did not confer an advantage in colonies and within-biofilm evolution did not select for this trait. We suggest that differences in the structural properties of EPS compounds formed by P. fluorescens and P. aeruginosa, and/or differences in the exploitability of each polysaccharide, contribute to discrepancies between observations made with the two species and may be relevant for the pathogenesis of P. aeruginosa.
Because Pel production is facultative, we did not expect adaptation to the colony edge to affect this trait. Surprisingly, mutants evolved in this colony subregion showed a consistent phenotype that indicated decreased Pel production. When mixed with the ancestral strain and inoculated for growth in the colony morphology assay, strains evolved at the colony edge excluded the wild type from this region. We note that full removal of Pel production did not affect the competitiveness of the Δpel mutant (Fig. 4), so either specific regulatory effects on Pel or other changes, such as increased colony spreading, gave rise to the increased fitness that we observed in the eColedge lineages.
Our evolution experiments revealed that Pel production is a trait that is tunable by mutation. Although wild-type P. aeruginosa has a facultative strategy that renders it able to respond to local variations in electron acceptor availability by adjusting EPS production (a single genotype enabling different phenotypes), microenvironments in biofilms may select for constitutive strategies (different genotypes with fixed phenotypes). Evolutionary theory predicts that a flexible response to environmental conditions is favored in changing environments (53). Obligate strategists, with increased or decreased matrix output, were observed in the population after only one round of growth in O2-limited (pellicle) and O2-replete (edge of colony) biofilm conditions, respectively. In subsequent competition experiments, these two mutant types were highly successful in the niches to which they were adapted. Therefore, clonal diversity arises in biofilm environments in part due to the distinct static conditions emerging in different regions of the same biofilm, but facultative regulation of traits such as Pel production may be retained over longer time scales due to the differential contributions of these traits in different types of biofilms or biofilm subregions. We found that although the pellicle and colony systems appear to impose similar constraints on resources (in this case, O2 availability), Pel production provided a competitive advantage—and therefore was subjected to positive selection—in only one of these biofilm formation regimes. Therefore, facultative control of Pel production may allow P. aeruginosa PA14 to optimize the contributions of this trait to survivability in different environments.
We evaluate fitness by counting CFU, which represent the viable cells in a sample, at the end of the competition. We do not presently have data addressing whether the strains investigated here differ in their level of survival, which could vary, for example, due to differential levels of cell death. However, as CFU measurements represent the net result of reproduction and survival, we feel they provide an appropriate estimation of Darwinian fitness. Another crucial point is whether we can ensure that mutations occur during our experiments as opposed to being present in the founder populations. This issue, which is central to all evolutionary experiments, is one that cannot be fully addressed deterministically. We start our cultures from a streaked inoculum, which reduces the amount of generations from clonal isolation to inoculation, but there is always the possibility that mutations occur in the meantime. Nevertheless, it is only during the biofilm growth that selection will act on the genetic diversity created from mutations, and it is this selection that is the focus of our study.
It has been suggested that the emergence and maintenance of clonal diversity in biofilms provides an overall benefit to the biofilm community by increasing survivability in the face of physiological stress (54). However, according to social evolution theory, group-level benefits alone are not sufficient to explain the evolution and stability of a social trait (38, 55, 56). Our results imply that spontaneous mutants with different Pel production phenotypes arise not as a result of group-level selection but rather because niches within the biofilm simultaneously select for distinct traits that confer benefits locally and at the individual level.
Electron acceptor limitation occurs in most biofilms and in sites of infection (57); thus, it is likely to be one common selective pressure that can induce clonal diversification in microbes. Our results suggest that the competitive fitness of strains with differences in Pel production depend on unique features of the biofilm models employed, such as the timing of electron acceptor gradient formation. Nevertheless, the imposed electron acceptor heterogeneity promotes phenotypic diversification, including diverse Pel production phenotypes, in both biofilm regimes. Diversity, genetic or otherwise, underpins the persistence of P. aeruginosa infections, such as those colonizing the lungs of cystic fibrosis patients (31). In this context, bacterial populations become spatially isolated (58) and are subjected to variations in environmental redox potential (57). Since drug efficacy is largely dependent on the genotype of the target, clonal diversity can lead to further complications when treating bacterial infections with antibiotics.
Supplementary Material
ACKNOWLEDGMENTS
We thank Maria Svistunova for expert laboratory assistance, Josephine Chandler for insightful discussions, and Chinweike Okegbe for help with electrode measurements.
This study was partly funded by the Danish Innovation Consortium, SiB, grant 11804520, and the Danish Council for Independent Research, grant 11106571 (to J.S.M.); the Amgen Scholars Program at Columbia University/Barnard College (supporting G.R.S.); grant R01 AI103369 from the National Institute of Allergy and Infectious Diseases (to L.E.P.D.); and by the Office of the Director, National Institutes of Health, under award DP2OD008440 (to J.B.X.).
We have no competing interests to declare.
Footnotes
Supplemental material for this article may be found at http://dx.doi.org/10.1128/AEM.02628-15.
REFERENCES
- 1.Lopez D, Vlamakis H, Kolter R. 2010. Biofilms. Cold Spring Harb Perspect Biol 2:a000398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Lewis K. 2001. Riddle of biofilm resistance. Antimicrob Agents Chemother 45:999–1007. doi: 10.1128/AAC.45.4.999-1007.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Recinos DA, Sekedat MD, Hernandez A, Cohen TS, Sakhtah H, Prince AS, Price-Whelan A, Dietrich LE. 2012. Redundant phenazine operons in Pseudomonas aeruginosa exhibit environment-dependent expression and differential roles in pathogenicity. Proc Natl Acad Sci U S A 109:19420–19425. doi: 10.1073/pnas.1213901109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Dietrich LE, Teal TK, Price-Whelan A, Newman DK. 2008. Redox-active antibiotics control gene expression and community behavior in divergent bacteria. Science 321:1203–1206. doi: 10.1126/science.1160619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Dietrich LE, Okegbe C, Price-Whelan A, Sakhtah H, Hunter RC, Newman DK. 2013. Bacterial community morphogenesis is intimately linked to the intracellular redox state. J Bacteriol 195:1371–1380. doi: 10.1128/JB.02273-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kempes CP, Okegbe C, Mears-Clarke Z, Follows MJ, Dietrich LE. 2014. Morphological optimization for access to dual oxidants in biofilms. Proc Natl Acad Sci U S A 111:208–213. doi: 10.1073/pnas.1315521110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Price-Whelan A, Dietrich LE, Newman DK. 2007. Pyocyanin alters redox homeostasis and carbon flux through central metabolic pathways in Pseudomonas aeruginosa PA14. J Bacteriol 189:6372–6381. doi: 10.1128/JB.00505-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wang Y, Kern SE, Newman DK. 2010. Endogenous phenazine antibiotics promote anaerobic survival of Pseudomonas aeruginosa via extracellular electron transfer. J Bacteriol 192:365–369. doi: 10.1128/JB.01188-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Arai H. 2011. Regulation and function of versatile aerobic and anaerobic respiratory metabolism in Pseudomonas aeruginosa. Front Microbiol 2:103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Friedman L, Kolter R. 2004. Genes involved in matrix formation in Pseudomonas aeruginosa PA14 biofilms. Mol Microbiol 51:675–690. [DOI] [PubMed] [Google Scholar]
- 11.Jennings LK, Storek KM, Ledvina HE, Coulon C, Marmont LS, Sadovskaya I, Secor PR, Tseng BS, Scian M, Filloux A, Wozniak DJ, Howell PL, Parsek MR. 2015. Pel is a cationic exopolysaccharide that cross-links extracellular DNA in the Pseudomonas aeruginosa biofilm matrix. Proc Natl Acad Sci U S A 112:11353–11358. doi: 10.1073/pnas.1503058112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Franklin MJ, Nivens DE, Weadge JT, Howell PL. 2011. Biosynthesis of the Pseudomonas aeruginosa extracellular polysaccharides, alginate, Pel, and Psl. Front Microbiol 2:167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Friedman L, Kolter R. 2004. Two genetic loci produce distinct carbohydrate-rich structural components of the Pseudomonas aeruginosa biofilm matrix. J Bacteriol 186:4457–4465. doi: 10.1128/JB.186.14.4457-4465.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Vasseur P, Vallet-Gely I, Soscia C, Genin S, Filloux A. 2005. The pel genes of the Pseudomonas aeruginosa PAK strain are involved at early and late stages of biofilm formation. Microbiology 151:985–997. doi: 10.1099/mic.0.27410-0. [DOI] [PubMed] [Google Scholar]
- 15.Colvin KM, Gordon VD, Murakami K, Borlee BR, Wozniak DJ, Wong GC, Parsek MR. 2011. The pel polysaccharide can serve a structural and protective role in the biofilm matrix of Pseudomonas aeruginosa. PLoS Pathog 7:e1001264. doi: 10.1371/journal.ppat.1001264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Colvin KM, Alnabelseya N, Baker P, Whitney JC, Howell PL, Parsek MR. 2013. PelA deacetylase activity is required for Pel polysaccharide synthesis in Pseudomonas aeruginosa. J Bacteriol 195:2329–2339. doi: 10.1128/JB.02150-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Yamamoto K, Arai H, Ishii M, Igarashi Y. 2011. Trade-off between oxygen and iron acquisition in bacterial cells at the air-liquid interface. FEMS Microbiol Ecol 77:83–94. doi: 10.1111/j.1574-6941.2011.01087.x. [DOI] [PubMed] [Google Scholar]
- 18.Xavier JB, Foster KR. 2007. Cooperation and conflict in microbial biofilms. Proc Natl Acad Sci U S A 104:876–881. doi: 10.1073/pnas.0607651104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Nadell CD, Xavier JB, Foster KR. 2009. The sociobiology of biofilms. FEMS Microbiol Rev 33:206–224. doi: 10.1111/j.1574-6976.2008.00150.x. [DOI] [PubMed] [Google Scholar]
- 20.Hamilton WD. 1964. Genetical evolution of social behaviour I. J Theor Biol 7:1–16. doi: 10.1016/0022-5193(64)90038-4. [DOI] [PubMed] [Google Scholar]
- 21.Nadell CD, Bassler BL. 2011. A fitness trade-off between local competition and dispersal in Vibrio cholerae biofilms. Proc Natl Acad Sci U S A 108:14181–14185. doi: 10.1073/pnas.1111147108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Nadell CD, Drescher K, Wingreen NS, Bassler BL. 2015. Extracellular matrix structure governs invasion resistance in bacterial biofilms. ISME J 9:1700–1709. doi: 10.1038/ismej.2014.246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Smith DR, Maestre-Reyna M, Lee G, Gerard H, Wang AH, Watnick PI. 2015. In situ proteolysis of the Vibrio cholerae matrix protein RbmA promotes biofilm recruitment. Proc Natl Acad Sci U S A 112:10491–10496. doi: 10.1073/pnas.1512424112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.van Gestel J, Weissing FJ, Kuipers OP, Kovacs AT. 2014. Density of founder cells affects spatial pattern formation and cooperation in Bacillus subtilis biofilms. ISME J 8:2069–2079. doi: 10.1038/ismej.2014.52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Foster KR, Xavier JB. 2007. Cooperation: bridging ecology and sociobiology. Curr Biol 17:R319–R321. doi: 10.1016/j.cub.2007.03.005. [DOI] [PubMed] [Google Scholar]
- 26.Rainey PB, Rainey K. 2003. Evolution of cooperation and conflict in experimental bacterial populations. Nature 425:72–74. doi: 10.1038/nature01906. [DOI] [PubMed] [Google Scholar]
- 27.Boles BR, Thoendel M, Singh PK. 2004. Self-generated diversity produces “insurance effects” in biofilm communities. Proc Natl Acad Sci U S A 101:16630–16635. doi: 10.1073/pnas.0407460101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Workentine ML, Sibley CD, Glezerson B, Purighalla S, Norgaard-Gron JC, Parkins MD, Rabin HR, Surette MG. 2013. Phenotypic heterogeneity of Pseudomonas aeruginosa populations in a cystic fibrosis patient. PLoS One 8:e60225. doi: 10.1371/journal.pone.0060225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Ashish A, Paterson S, Mowat E, Fothergill JL, Walshaw MJ, Winstanley C. 2013. Extensive diversification is a common feature of Pseudomonas aeruginosa populations during respiratory infections in cystic fibrosis. J Cyst Fibros 12:790–793. doi: 10.1016/j.jcf.2013.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Markussen T, Marvig RL, Gomez-Lozano M, Aanaes K, Burleigh AE, Hoiby N, Johansen HK, Molin S, Jelsbak L. 2014. Environmental heterogeneity drives within-host diversification and evolution of Pseudomonas aeruginosa. mBio 5:e01592-14. doi: 10.1128/mBio.01592-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Rodriguez-Rojas A, Oliver A, Blazquez J. 2012. Intrinsic and environmental mutagenesis drive diversification and persistence of Pseudomonas aeruginosa in chronic lung infections. J Infect Dis 205:121–127. doi: 10.1093/infdis/jir690. [DOI] [PubMed] [Google Scholar]
- 32.Koza A, Moshynets O, Otten W, Spiers AJ. 2011. Environmental modification and niche construction: developing O2 gradients drive the evolution of the Wrinkly Spreader. ISME J 5:665–673. doi: 10.1038/ismej.2010.156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Shanks RM, Caiazza NC, Hinsa SM, Toutain CM, O'Toole GA. 2006. Saccharomyces cerevisiae-based molecular tool kit for manipulation of genes from gram-negative bacteria. Appl Environ Microbiol 72:5027–5036. doi: 10.1128/AEM.00682-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lambertsen L, Sternberg C, Molin S. 2004. Mini-Tn7 transposons for site-specific tagging of bacteria with fluorescent proteins. Environ Microbiol 6:726–732. doi: 10.1111/j.1462-2920.2004.00605.x. [DOI] [PubMed] [Google Scholar]
- 35.Ueda A, Wood TK. 2009. Connecting quorum sensing, c-di-GMP, pel polysaccharide, and biofilm formation in Pseudomonas aeruginosa through tyrosine phosphatase TpbA (PA3885). PLoS Pathog 5:e1000483. doi: 10.1371/journal.ppat.1000483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Spiers AJ, Bohannon J, Gehrig SM, Rainey PB. 2003. Biofilm formation at the air-liquid interface by the Pseudomonas fluorescens SBW25 wrinkly spreader requires an acetylated form of cellulose. Mol Microbiol 50:15–27. doi: 10.1046/j.1365-2958.2003.03670.x. [DOI] [PubMed] [Google Scholar]
- 37.Okegbe C, Sakhtah H, Sekedat MD, Price-Whelan A, Dietrich LE. 2012. Redox eustress: roles for redox-active metabolites in bacterial signaling and behavior. Antioxid Redox Signal 16:658–667. doi: 10.1089/ars.2011.4249. [DOI] [PubMed] [Google Scholar]
- 38.de Vargas Roditi L, Boyle KE, Xavier JB. 2013. Multilevel selection analysis of a microbial social trait. Mol Syst Biol 9:684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Werner E, Roe F, Bugnicourt A, Franklin MJ, Heydorn A, Molin S, Pitts B, Stewart PS. 2004. Stratified growth in Pseudomonas aeruginosa biofilms. Appl Environ Microbiol 70:6188–6196. doi: 10.1128/AEM.70.10.6188-6196.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hallatschek O, Hersen P, Ramanathan S, Nelson DR. 2007. Genetic drift at expanding frontiers promotes gene segregation. Proc Natl Acad Sci U S A 104:19926–19930. doi: 10.1073/pnas.0710150104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Schluter J, Nadell CD, Bassler BL, Foster KR. 2015. Adhesion as a weapon in microbial competition. ISME J 9:139–149. doi: 10.1038/ismej.2014.174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Spiers AJ, Kahn SG, Bohannon J, Travisano M, Rainey PB. 2002. Adaptive divergence in experimental populations of Pseudomonas fluorescens. I. Genetic and phenotypic bases of wrinkly spreader fitness. Genetics 161:33–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kirisits MJ, Prost L, Starkey M, Parsek MR. 2005. Characterization of colony morphology variants isolated from Pseudomonas aeruginosa biofilms. Appl Environ Microbiol 71:4809–4821. doi: 10.1128/AEM.71.8.4809-4821.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.McBain AJ. 2009. Chapter 4. In vitro biofilm models: an overview. Adv Appl Microbiol 69:99–132. [DOI] [PubMed] [Google Scholar]
- 45.Serra DO, Richter AM, Hengge R. 2013. Cellulose as an architectural element in spatially structured Escherichia coli biofilms. J Bacteriol 195:5540–5554. doi: 10.1128/JB.00946-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Ali A, Rashid MH, Karaolis DK. 2002. High-frequency rugose exopolysaccharide production by Vibrio cholerae. Appl Environ Microbiol 68:5773–5778. doi: 10.1128/AEM.68.11.5773-5778.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Vlamakis H, Chai Y, Beauregard P, Losick R, Kolter R. 2013. Sticking together: building a biofilm the Bacillus subtilis way. Nat Rev Microbiol 11:157–168. doi: 10.1038/nrmicro2960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Fong JC, Yildiz FH. 2007. The rbmBCDEF gene cluster modulates development of rugose colony morphology and biofilm formation in Vibrio cholerae. J Bacteriol 189:2319–2330. doi: 10.1128/JB.01569-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Okegbe C, Price-Whelan A, Dietrich LE. 2014. Redox-driven regulation of microbial community morphogenesis. Curr Opin Microbiol 18:39–45. doi: 10.1016/j.mib.2014.01.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kim W, Racimo F, Schluter J, Levy SB, Foster KR. 2014. Importance of positioning for microbial evolution. Proc Natl Acad Sci U S A 111:E1639–E1647. doi: 10.1073/pnas.1323632111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Hay ID, Wang Y, Moradali MF, Rehman ZU, Rehm BH. 2014. Genetics and regulation of bacterial alginate production. Environ Microbiol 16:2997–3011. doi: 10.1111/1462-2920.12389. [DOI] [PubMed] [Google Scholar]
- 52.McElroy KE, Hui JG, Woo JK, Luk AW, Webb JS, Kjelleberg S, Rice SA, Thomas T. 2014. Strain-specific parallel evolution drives short-term diversification during Pseudomonas aeruginosa biofilm formation. Proc Natl Acad Sci U S A 111:E1419–E1427. doi: 10.1073/pnas.1314340111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Levins R. 1968. Evolution in changing environments: some theoretical explorations. Princeton University Press, Princeton, NJ. [Google Scholar]
- 54.Boles BR, Singh PK. 2008. Endogenous oxidative stress produces diversity and adaptability in biofilm communities. Proc Natl Acad Sci U S A 105:12503–12508. doi: 10.1073/pnas.0801499105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Foster KR. 2009. A defense of sociobiology. Cold Spring Harbor Symp Quant Biol 74:403–418. doi: 10.1101/sqb.2009.74.041. [DOI] [PubMed] [Google Scholar]
- 56.West SA, Griffin AS, Gardner A, Diggle SP. 2006. Social evolution theory for microorganisms. Nat Rev Microbiol 4:597–607. doi: 10.1038/nrmicro1461. [DOI] [PubMed] [Google Scholar]
- 57.Cowley ES, Kopf SH, LaRiviere A, Ziebis W, Newman DK. 2015. Pediatric cystic fibrosis sputum can be chemically dynamic, anoxic, and extremely reduced due to hydrogen sulfide formation. mBio 6:e00767-15. doi: 10.1128/mBio.00767-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Jorth P, Staudinger BJ, Wu X, Hisert KB, Hayden H, Garudathri J, Harding CL, Radey MC, Rezayat A, Bautista G, Berrington WR, Goddard AF, Zheng C, Angermeyer A, Brittnacher MJ, Kitzman J, Shendure J, Fligner CL, Mittler J, Aitken ML, Manoil C, Bruce JE, Yahr TL, Singh PK. 2015. Regional isolation drives bacterial diversification within cystic fibrosis lungs. Cell Host Microbe 18:307–319. doi: 10.1016/j.chom.2015.07.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Rahme LG, Stevens EJ, Wolfort SF, Shao J, Tompkins RG, Ausubel FM. 1995. Common virulence factors for bacterial pathogenicity in plants and animals. Science 268:1899–1902. doi: 10.1126/science.7604262. [DOI] [PubMed] [Google Scholar]
- 60.Dietrich LE, Price-Whelan A, Petersen A, Whiteley M, Newman DK. 2006. The phenazine pyocyanin is a terminal signalling factor in the quorum sensing network of Pseudomonas aeruginosa. Mol Microbiol 61:1308–1321. doi: 10.1111/j.1365-2958.2006.05306.x. [DOI] [PubMed] [Google Scholar]
- 61.Dehio C, Meyer M. 1997. Maintenance of broad-host-range incompatibility group P and group Q plasmids and transposition of Tn5 in Bartonella henselae following conjugal plasmid transfer from Escherichia coli. J Bacteriol 179:538–540. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.