

Abstract
Spermatozoa are highly specialized cells. Adenosine triphosphate (ATP), which provides the energy for supporting the key functions of the spermatozoa, is formed by 2 metabolic pathways, namely glycolysis and oxidative phosphorylation (OXPHOS). It is produced in the mitochondria through OXPHOS as well as in the head and principal piece of the flagellum through glycolysis. However, there is a great discrepancy as to which method of ATP production is primarily utilized by the spermatozoa for successful fertilization. Mitochondrial respiration is considered to be a more efficient metabolic process for ATP synthesis in comparison to glycolysis. However, studies have shown that the diffusion potential of ATP from the mitochondria to the distal end of the flagellum is not sufficient to support sperm motility, suggesting that glycolysis in the tail region is the preferred pathway for energy production. It is suggested by many investigators that although glycolysis forms the major source of ATP along the flagellum, energy required for sperm motility is mainly produced during mitochondrial respiration. Nevertheless, some studies have shown that when glycolysis is inhibited, proper functioning and motility of spermatozoa remains intact although it is unclear whether such motility can be sustained for prolonged periods of time, or is sufficiently vigorous to achieve optimal fertilization. The purpose of this article is to provide an overview of mammalian sperm energy metabolism and identify the preferred metabolic pathway for ATP generation which forms the basis of energy production in human spermatozoa during fertilization.
Keywords: adenosine triphosphate, energy, glycolysis, oxidative phosphorylation, spermatozoa
INTRODUCTION
All living cells require energy for both development and function and male gametes, are no exception. Mammalian spermatozoa specifically use adenosine triphosphate (ATP) to maintain the intracellular milieu1 and for cellular processes such as motility, capacitation, hyperactivation and the acrosome reaction,2,3 all of which are needed for successful fertilization. ATP is formed from adenosine diphosphate (ADP), and therefore the addition and removal of phosphate groups to ADP molecules forms the basis of all life processes.
Throughout the years there has been ambiguity regarding the metabolic pathway that plays the most vital role in energy production in spermatozoa. According to Rees et al.4 mammalian spermatozoa share a common problem in that ATP must be delivered along the entire length of the flagellum, which means they must adopt a variety of metabolic strategies to generate it. The purpose of the present article is to provide an overview on mammalian sperm energy metabolism and identify the preferred metabolic pathway for ATP generation in human spermatozoa.
PATHWAYS OF ENERGY PRODUCTION IN SPERMATOZOA
Adenosine triphosphate is produced in the spermatozoon via two metabolic pathways – glycolysis and oxidative phosphorylation (OXPHOS) – with the former occurring in the head and principal piece of the flagellum and the latter occurring in the mitochondria. A glycolysis involves the breakdown of six carbon monosaccharides during a series of enzyme-catalyzed reactions that yield two molecules of three carbon compound pyruvates. In the above mentioned metabolic process the net yield of ATP is two molecules per molecule of glucose oxidized. In subsequent steps, pyruvate is further oxidized and the carboxyl group is lost as CO2 to yield the acetyl group of acetyl-coenzyme A. The acetyl group is then oxidized completely to CO2 by the citric acid cycle. Electron donors, namely NADH and FADH2, are formed as a result of metabolic processes such as glycolysis, fatty acid oxidation, and the citric acid cycle, and they are energy-rich molecules with high electron transfer potential. In sequential steps the flow of electrons thus proceeds from NADH or FADH2 to O2 through protein complexes located in the inner mitochondrial membrane. An uneven distribution of protons that leads to a proton-motive force is created as the electron transfer results in the pumping of protons out of the mitochondrial matrix. ATP is synthesized when protons flow back into the mitochondrial matrix through an enzyme complex, namely ATP synthase.5,6
Oxidative phosphorylation is a more complicated process that transpires in a coordinated fashion and involves two components of the inner mitochondrial membrane – the respiratory chain and ATP synthase.7 OXPHOS generates thirty molecules of ATP per molecule of oxidized glucose. The generation of ATP through OXPHOS is therefore 15 times more efficient than glycolysis. The most important factor that determines the rate of OXPHOS is the availability of ADP.6,8 To summarize, the oxidation of fuels and the phosphorylation of ADP, coupled with a proton gradient across the inner mitochondrial membrane are key factors in the generation of ATP.
Although a spermatozoon can survive purely on glycolytic energy,9 it requires OXPHOS for differentiation and maturation.10,11,12 The preferred metabolic pathway opted for by spermatozoa with regard to energy production is highly species specific.13 For example, various studies14,15 have shown that glycolysis is essential for fertilization in mice, rat, hamster and humans spermatozoa, but not so in bovine sperm.
ADENOSINE TRIPHOSPHATE PRODUCTION SITES IN SPERMATOZOA
Spermatozoa are highly differentiated cells consisting of three distinct regions: the head, mid-piece and tail. The sperm head contains the nucleus, which houses the highly compacted paternal genome.16 Extending posteriorly from the head is the mid-piece, which is followed by the flagellum. The flagellum comprises of the principal piece of the spermatozoa and inside it is the fibrous sheath - a characteristic feature of mammalian spermatozoa.17 Similarly, energy production in spermatozoa is also compartmentalized as it is in all eukaryotic cells:18 ATP generation through OXPHOS occur in the mitochondria whereas glycolysis occurs in the head and fibrous sheath of the flagellum (Figure 1).
Figure 1.
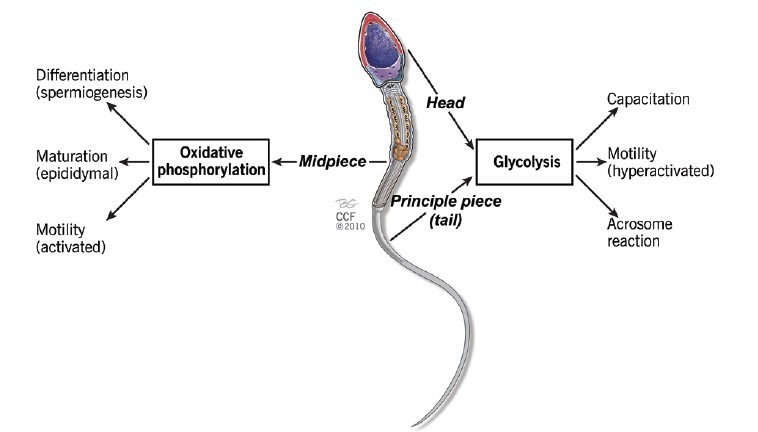
A proposed schematic representation of the compartmentalization of labor in the spermatozoon through different modes of adenosine triphosphate generation at varying sites.
Site of adenosine triphosphate production via oxidative phosphorylation
Mitochondria play a key role in the production of ATP through the process of OXPHOS. Therefore, they are appropriately referred to as the “power house” of the cell. They are known to occupy a major portion of the cytoplasmic volume of eukaryotic cells, and play an essential role in the evolution of complex animals.1
Located in the mitochondrial sheath and confined to the mid-piece region of the spermatozoa,19 mitochondria occupy a substantial portion of the total cellular volume (approximately 15%–22%).20 A mature spermatozoon contains approximately 72–80 mitochondria, which are involved in major processes such as the acrosome reaction and oocyte penetration.21 Although it consists of four distinct sub-compartments - the outer mitochondrial membrane, the inter-membrane space, an inner mitochondrial membrane and a matrix - a sperm mitochondrion varies from its somatic cell counterparts both morphologically and functionally.19,22 These differences can be attributed to the tight wrapping of the mitochondria around the axoneme of the spermatozoon. This tight wrapping results in a mitochondrial capsule consisting of selenoprotein and comprising of disulfide bridges,23 rendering stability to the mitochondrial sheath.13,24 The inner mitochondrial membrane is folded into structures called cristae, which are the principal sites for OXPHOS and ATP generation. Sperm mitochondria possess specific isoforms of proteins and isoenzymes such as cytochrome C,25,26,27 hexokinase subunit VIb of the cytochrome C oxidase,28 and lactate dehydrogenase (LDH)29 all of which are absent in somatic cell mitochondria.30
Site of adenosine triphosphate production via glycolysis
Glycolysis plays a key role in mammalian sperm energy production and thus, an understanding of the site of glycolitically produced ATP is therefore of utmost importance.31,32,33 Because the head and principal piece of the flagellum are devoid of respiratory enzymes, ATP production can therefore only occur through the process of glycolysis in these areas.1 Several glycolytic enzymes specific to the spermatogenic cell have been identified in the fibrous sheath of spermatozoa. These include hexokinase, phosphoglucokinase isomerase, phosphofructokinase, LDH and glyceraldehyde-3-phosphate dehydrogenase (GAPD).8,18,34,35,36,37
ENERGY NEEDS OF SPERMATOZOA
Mammalian fertilization is a multifaceted and complex process because the site of semen deposition is far from the site of fertilization in the female genital tract.38,39 Therefore, cellular processes such as motility, capacitation, hyperactivation and acrosome reaction must occur in a synchronized fashion.40 As all of these actions are energy dependent, the importance and role of energy in normal male gamete function cannot be overstated. The energy utilization of each of these processes will be discussed below.
Adenosine triphosphate utilization during sperm motility
Spermatozoa compete with one another during their transit in the female reproductive tract in order to reach the female gamete first. Adequate motility is therefore required for successful fertilization. Movement is generated by the flagellum that constitutes more than 90% of the length of the mammalian spermatozoon.21 Paoli et al.40 define sperm motility as the “result of propagation of transverse waves along the flagellum in a proximal – distal direction, thus producing a hydrodynamic impulse that pushes the spermatozoon through the female genital tract toward the oocyte.” Spermatozoa display two types of motility – activated and hyperactivated.41 Activated motility, as seen in the freshly ejaculated spermatozoa,20 refers to the low amplitude, symmetric waves that propagate along the length of the flagellum and result in linear propulsion of the sperm cell.42 Hyperactivated motility, another characteristic feature of the spermatozoa, occurs when flagellar movement becomes asymmetrical with higher amplitude, resulting in highly curved trajectories.43,44 Spermatozoa is dependent on ATP-fueled motility,45 yet how they are metabolically adapted to utilize ATP is less understood.46 ATP is required to support the coordinated movement of the central axoneme and the surrounding flagellar structures.20
The flagellar beat
The motile apparatus that propels a spermatozoon forward and enables it to penetrate an oocyte is contained in the flagellum.47 In general, speaking, flagellar movement is accompanied by the bending of the eukaryotic flagellum produced by the sliding movement of microtubules caused by the activity of dynein “arms.”48,49,50 Several experiments have been carried out to better understand the regulatory mechanism of flagellar beating of spermatozoa.51,52,53,54 Axonemal dynein is a protein that powers the rhythmic beat of the flagellum.55 Upon the activation of dynein ATPases, there is sliding of the outer axonemal doublet microtubules, causing the flagellum to bend.1 A force is thus generated when the dynein protein transduces the chemical energy of ATP hydrolysis into mechanical energy.55,56
Flagellum fuel
Because mitochondria are bulky structures, they are localized in the mid-piece region of the sperm cell and not distributed along the flagellum, as they may cause mechanical problems during flagellar beating.46 However, it is a known fact that efficient ATP synthesis occurs through mitochondrial respiration. The question then arises as to whether mitochondrial ATP can diffuse rapidly and sufficiently along the entire length of the flagellum to support the energy requirements needed for fast beating activity. Unfortunately, the answer is not clear.
Several biophysicists57,58 have calculated the required rate of diffusion of ATP to support motility in the spermatozoa of sea urchins and bulls and concluded that the diffusion rate of ATP is sufficient to keep pace with the beating frequency of the flagellum. Mitochondrial membrane potential is considered an indicator of sperm motility and a reduction in potential has resulted in reduced sperm motility and fertilization ability.59,60,61,62 In the studies of Paoli et al.40 which were conducted on asthenozoospermic patients, a positive correlation has been established between mitochondrial membrane potential and non-linear motility. Alterations in the mitochondrial respiratory chain enzyme activities can also have an effect on sperm motility.63 The electron transfer chain is composed of two mobile carriers (coenzyme Q and cytochrome C) and four multimeric complexes (I, II, III, and IV). Based on their experiments on sperm motility and mitochondrial enzyme specific activities, Ruiz-Pesini et al.11 suggested that mitochondrial dysfunction may lead to idiopathic asthenozoospermia.
This is in contradiction to the views of Tombes and Shapiro64 whose findings suggest that when ATP is produced by the distant cell body or mitochondrion, whether by glycolysis or OXPHOS, it does not meet the energy demands needed for proper flagellar development due to the insufficient diffusion capacity of the ATP to the area where the energy demand is high. Whether this is applicable to human spermatozoa is still unclear.20 Furthermore, the products of ATP hydrolysis such as ADP, inorganic phosphate Pi and H+ must be removed efficiently to avoid kinetic and thermodynamic stress.65 As such, several researchers have proposed an alternative pathway of energy production by the spermatozoa. They have suggested that there is a need for local ATP production closer to the site of ATP utilization, and that this is achieved by the glycolytic pathway of ATP production due to the presence of glycolytic enzymes in the fibrous sheath of the flagellum.18 Moreover, despite species specific differences,66 mammalian spermatozoa can use a variety of carbohydrates as substrate for ATP generation.67,68,69,70 This allows for ATP generation in the cytoplasm independent of mitochondrial activity.18
Glucose is considered to induce a high beat frequency in the flagellum and in this context Mukai and Okuno, for example, used a glucose analogue, 2-deoxyglucose (DOG), to inhibit glycolysis in spermatozoa. They found that DOG had no effect on mitochondrial respiration as assessed by a fluorescent probe - 5,5’,6,6’-tetrachloro-1,1’,3,3’-tetraethylbenzimidazolyl carbocyanine iodide (JC-1),32 whereas it did inhibit the activity of pyruvate and lactate when provided as substrates. This further resulted in low motility and decreased ATP content.32 The above evidence thus supports the fact that glycolysis acts as a key contributor to sperm motility and when suppressed, leads to decreased motility even though mitochondrial substrates are present.
The ATP content in the presence of DOG was also examined. Interestingly, the concentration of ATP in the presence and absence of pyruvate along with the DOG were found to be similar. Because DOG cannot inhibit ATP synthesis in mitochondria, this suggests that normal mitochondrial respiration is not sufficient to maintain the concentration of ATP required for sperm flagellar motility. Thus, the above facts strengthen the argument that glycolysis is the preferred energy source specifically for motility functions.
As mentioned earlier, spermatogenic specific isoforms of proteins that are essential for glycolysis are found throughout the fibrous sheath of the sperm flagellum. One of the most notable enzymes is the GAPD enzyme. GAPD-S is a gene that is specific to mouse spermatogenic cells;14 its human ortholog is GAPD-2. In fact, GAPD is the first catalytically active enzyme proven to be bound to the fibrous sheath of spermatozoa.18 It is a key regulator of glycolysis during spermatogenesis and is also the target of various environmental compounds that adversely affect male fertility.71 Miki et al.14 studied GAPD-S gene expression in knock-out mice and concluded that the sperm produced by GAPD-S-/- mice have low motility and exhibit no forward progression.14 Welch et al.72 also sequenced and cloned the cDNA for the human homologue GAPD-2 and suggested that it may have similar roles in human spermatozoa.
Carbonyl cyanide m-chlorophenylhydrazine (CCCP) acts as an uncoupler to inhibit mitochondrial ATP production. The addition of CCCP to highly active mice spermatozoa had no effect on ATP content and motility parameters, suggesting that mitochondrial respiration does not play a vital role in sperm motility.32 Similarly, LDH-C, a spermatogenic cell variant and a glycolytic enzyme,29,30 catalyzes the conversion of pyruvate to lactate, which is essential for sperm motility during anaerobic respiration. Disruption of the targeted LDH-C gene in mice showed neither tyrosine phosphorylation nor hyperactive motility necessary for capacitation.73 On the other hand, Hung et al.74 proposed that OXPHOS is not necessary for hyperactivated sperm motility in rhesus macaque monkeys. Moreover, Hereng et al.75 have also shown that during exogenous supplementation of pyruvate and glucose, the glycolytic pathway acted as the predominant source of energy to support progressive motility and capacitation through ATP generation.
Adenosine triphosphate utilization during capacitation and acrosome reaction
The process of fertilization includes several steps that occur in an orderly and synchronized fashion.76 After spermatozoa are deposited in the female reproductive tract, a series of events, collectively referred to as capacitation, take place before fertilization is possible.21 Capacitation can be defined as “the cellular and biochemical modifications that the spermatozoon undergoes in response to appropriate stimuli essential for acrosome reaction to occur.”77 When the human sperm are incubated in capacitation medium, an average of 10%–20% develops motility patterns that look like hyperactivation.78,79 Efficient generation of ATP is required by mammalian spermatozoa to fuel these crucial events.80 Modifications occur at both the surface of the head as well as throughout the flagellum of the spermatozoa.77 The acrosome reaction is an important aspect of the fertilization process and refers to the release of hydrolytic enzymes from the acrosome vesicles, ultimately leading to successful sperm binding and penetration of the oocyte.
The acrosome reaction is initiated when signals produced by the oocyte activate G-proteins inside the sperm thereby increasing intracellular Ca2+ levels, and finally activating several kinases and phosphorylation of various proteins. Ca2+ can be derived either through the plasma membrane channels formed by proteins in the CatSper family or from Ca2+ stored in organelles.81 It should be noted that although the CatSper channels are confined to the principal piece of the flagellum, the redundant nuclear envelope (RNE), which is a cluster of membrane vesicles82,83 that also serves as an important store for Ca2+ ions is localized to the base of the midpiece of the mitochondrial sheath. A rise in Ca2+ levels in acrosome-reacted spermatozoa has been observed to increase flagellar intensity leading to an increase in of hyperactivation.84
Adenosine triphosphate is required for ATPase activity, cyclic adenosine monophosphate formation and phosphorylation. Thus, it plays a crucial role in maintaining acrosomal integrity and inducting of the acrosome reaction.
Mitochondria play a key role in capacitation-dependent tyrosine phosphorylation in mammalian spermatozoa.85,86 In human spermatozoa, a secretory pathway – Ca2+-ATPase - has been immunolocalized to the midpiece and rear head and may play a role in clearance of Ca2+ released from the RNE store.87 In addition, selenium-dependent phospholipid hydroperoxide glutathione peroxidase is abundantly expressed in spermatids and displays high activity in the post-pubertal testis.88 In mature spermatozoa it is restricted to the mid-piece region23 which embeds the helix of mitochondria and plays a key role in sperm maturation and mitochondrial function.89 However, the exact mechanism of tyrosine phosphorylation during human sperm capacitation has not been described.
Mammalian spermatozoa rely on high concentrations of glucose, pyruvate and lactate,32,67,90 all of which are present in the oviductal fluid.91 The oviductal fluid of sheep and pigs is known to contain lactate, which serves as a substrate for spermatozoa activity whereas in mouse sperm, pyruvate and glucose could maintain vigorous motility for a longer duration.92,93 In fact, the use of metabolic substrates for ATP production varies amongst species and this ultimately determines the preferred metabolic pathway of ATP production in the spermatozoa to support capacitation.13 Depending on the presence of extracellular substrates, sperm metabolize those substrates by glycolysis to provide energy for flagellar movement. Alternatively, when there is a paucity of substrates for glycolysis, sperm metabolize respiratory substrates.32 In this context, respiratory substrates function as substrates for gluconeogenesis in the midpiece, resulting in glucose that can diffuse to other regions of the sperm flagellum. Although bull spermatozoa depend on OXPHOS to support capacitation,94 human spermatozoa seem to depend on glucose-derived ATP.67,95 Although glucose-derived ATP serves as an immediate source for energy, mitochondrial functionality increases during sperm capacitation.96,97
In order to determine the functionality of the mitochondria during capacitation, a comparative analysis of spermatozoa was done by taking samples before and after swim-up treatment and incubating them for different time intervals. The results showed mitochondrial respiratory activity in the cells incubated under capacitating conditions was significantly higher than that in the cells before swim-up. This might be due to efficient ATP generation through OXPHOS.98
CONCLUSIONS
A great deal of research has been conducted to understand the mechanisms of ATP utilization and identify the forms that act as the major source of ATP generation for human spermatozoa. Several authors have proposed that the cytosolic process of glycolysis forms the main source of ATP production in spermatozoa-even in the presence of oxygen. However, it is also believed that mitochondria significantly contribute to energy manufacturing based on their efficiency of ATP production during aerobic respiration in comparison to glycolysis. Understanding the molecular processes relating to the development of a functional and motile spermatozoon will help us to answer some of the questions related to idiopathic asthenozoospermia and possibly even idiopathic teratozoospermia. Further knowledge of molecular mechanisms related to sperm function during fertilization may help to target specific signaling pathways, and the disruption of the activity of certain proteins could even lead to the development of a safe and effective male contraceptive.
Our research suggests that (1) even though glycolysis is the predominant ATP generation pathway during hyperactivation, the mitochondria's role in sperm maturation and differentiation must not be underestimated; (2) both pathways seem to be essential for human sperm function and successful fertilization; and (3) metabolic energy production in human spermatozoa may be restricted to either glycolysis or mitochondrial respiration, or could take place in combination, depending on the surrounding environment and the availability of substrates.
AUTHOR CONTRIBUTIONS
SSDP conceived the idea and helped to write and edit the manuscript. GM helped to write and edit the manuscript. MVDL and AA helped to edit the manuscript. All authors discussed and commented on the manuscript at all stages.
COMPETING INTERESTS
The authors declare that they have no competing interests.
ACKNOWLEDGMENTS
This study was supported by the Harry Crossley foundation, the Cleveland Clinic Research Program and the INSPIRE program (DST) India.
REFERENCES
- 1.Mishro MM, Ramya T. Fuel/energy sources of spermatozoa. In: Parekattil JS, Agarwal A, editors. Male Infertility. New York: Springer-Verlag; 2012. pp. 209–23. [Google Scholar]
- 2.Mannowetz N, Wandernoth PM, Wennemuth G. Glucose is a pH-dependent motor for sperm beat frequency during early activation. PLoS One. 2012;7:e41030. doi: 10.1371/journal.pone.0041030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Mukai C, Travis AJ. What sperm can teach us about energy production. Reprod Domest Anim. 2012;47(Suppl 4):164–9. doi: 10.1111/j.1439-0531.2012.02071.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Rees JM, Ford WC, Hull MG. Effect of caffeine and of pentoxifylline on the motility and metabolism of human spermatozoa. J Reprod Fertil. 1990;90:147–56. doi: 10.1530/jrf.0.0900147. [DOI] [PubMed] [Google Scholar]
- 5.Gnaiger E. Bioenergetics at low oxygen: dependence of respiration and phosphorylation on oxygen and adenosine diphosphate supply. Respir Physiol. 2001;128:277–97. doi: 10.1016/s0034-5687(01)00307-3. [DOI] [PubMed] [Google Scholar]
- 6.Erecińska M, Wilson DF. On the mechanism of regulation of cellular respiration. The dependence of respiration on the cytosolic [ATP],[ADP] and [PI] Adv Exp Med Biol. 1977;94:271–8. doi: 10.1007/978-1-4684-8890-6_36. [DOI] [PubMed] [Google Scholar]
- 7.Piomboni P, Focarelli R, Stendardi A, Ferramosca A, Zara V. The role of mitochondria in energy production for human sperm motility. Int J Androl. 2012;35:109–24. doi: 10.1111/j.1365-2605.2011.01218.x. [DOI] [PubMed] [Google Scholar]
- 8.Kim YH, Haidl G, Schaefer M, Egner U, Mandal A, et al. Compartmentalization of a unique ADP/ATP carrier protein SFEC (Sperm Flagellar Energy Carrier, AAC4) with glycolytic enzymes in the fibrous sheath of the human sperm flagellar principal piece. Dev Biol. 2007;302:463–76. doi: 10.1016/j.ydbio.2006.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Spiropoulos J, Turnbull DM, Chinnery PF. Can mitochondrial DNA mutations cause sperm dysfunction? Mol Hum Reprod. 2002;8:719–21. doi: 10.1093/molehr/8.8.719. [DOI] [PubMed] [Google Scholar]
- 10.Nakada K, Sato A, Yoshida K, Morita T, Tanaka H, et al. Mitochondria-related male infertility. Proc Natl Acad Sci U S A. 2006;103:15148–53. doi: 10.1073/pnas.0604641103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ruiz-Pesini E, Diez C, Lapeña AC, Pérez-Martos A, Montoya J, et al. Correlation of sperm motility with mitochondrial enzymatic activities. Clin Chem. 1998;44:1616–20. [PubMed] [Google Scholar]
- 12.Ford WC. Glycolysis and sperm motility: does a spoonful of sugar help the flagellum go round? Hum Reprod Update. 2006;12:269–74. doi: 10.1093/humupd/dmi053. [DOI] [PubMed] [Google Scholar]
- 13.Storey BT. Mammalian sperm metabolism: oxygen and sugar, friend and foe. Int J Dev Biol. 2008;52:427–37. doi: 10.1387/ijdb.072522bs. [DOI] [PubMed] [Google Scholar]
- 14.Miki K, Qu W, Goulding EH, Willis WD, Bunch DO, et al. Glyceraldehyde 3-phosphate dehydrogenase-S, a sperm-specific glycolytic enzyme, is required for sperm motility and male fertility. Proc Natl Acad Sci U S A. 2004;101:16501–6. doi: 10.1073/pnas.0407708101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Galantino-Homer HL, Florman HM, Storey BT, Dobrinski I, Kopf GS. Bovine sperm capacitation: assessment of phosphodiesterase activity and intracellular alkalinization on capacitation-associated protein tyrosine phosphorylation. Mol Reprod Dev. 2004;67:487–500. doi: 10.1002/mrd.20034. [DOI] [PubMed] [Google Scholar]
- 16.Malo AF, Gomendio M, Garde J, Lang-Lenton B, Soler AJ, et al. Sperm design and sperm function. Biol Lett. 2006;2:246–9. doi: 10.1098/rsbl.2006.0449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Fouquet JP, Edde B, Kann ML, Wolff A, Desbruyeres E, et al. Differential distribution of glutamylated tubulin during spermatogenesis in mammalian testis. Cell Motil Cytoskeleton. 1994;27:49–58. doi: 10.1002/cm.970270106. [DOI] [PubMed] [Google Scholar]
- 18.Westhoff D, Kamp G. Glyceraldehyde 3-phosphate dehydrogenase is bound to the fibrous sheath of mammalian spermatozoa. J Cell Sci. 1997;110:1821–9. doi: 10.1242/jcs.110.15.1821. [DOI] [PubMed] [Google Scholar]
- 19.Ramalho-Santos J, Varum S, Amaral S, Mota PC, Sousa AP, et al. Mitochondrial functionality in reproduction: from gonads and gametes to embryos and embryonic stem cells. Hum Reprod Update. 2009;15:553–72. doi: 10.1093/humupd/dmp016. [DOI] [PubMed] [Google Scholar]
- 20.Turner RM. Tales from the tail: what do we really know about sperm motility? J Androl. 2003;24:790–803. doi: 10.1002/j.1939-4640.2003.tb03123.x. [DOI] [PubMed] [Google Scholar]
- 21.Rajender S, Rahul P, Mahdi AA. Mitochondria, spermatogenesis and male infertility. Mitochondrion. 2010;10:419–28. doi: 10.1016/j.mito.2010.05.015. [DOI] [PubMed] [Google Scholar]
- 22.Ferramosca A, Provenzano SP, Coppola L, Zara V. Mitochondrial respiratory efficiency is positively correlated with human sperm motility. Urology. 2012;79:809–14. doi: 10.1016/j.urology.2011.12.042. [DOI] [PubMed] [Google Scholar]
- 23.Calvin HI. Comparative labelling of rat epididymal spermatozoa by intratesticularly administered 65ZnCl2 and [35S] cysteine. J Reprod Fertil. 1981;61:65–73. doi: 10.1530/jrf.0.0610065. [DOI] [PubMed] [Google Scholar]
- 24.Ho HC, Suarez SS. Characterization of the intracellular Calcium store at the base of the sperm flagellum that regulates Hyperactiated motility. Biol Reprod. 2003;68:1590–6. doi: 10.1095/biolreprod.102.011320. [DOI] [PubMed] [Google Scholar]
- 25.Goldberg E, Sberna D, Wheat TE, Urbanski GJ, Margoliash E. Cytochrome c: immunofluorescent localization of the testis-specific form. Science. 1977;196:1010–2. doi: 10.1126/science.193188. [DOI] [PubMed] [Google Scholar]
- 26.Hess RA, Miller LA, Kirby JD, Margoliash E, Goldberg E. Immunoelectron microscopic localization of testicular and somatic cytochromes c in the seminiferous epithelium of the rat. Biol Reprod. 1993;48:1299–308. doi: 10.1095/biolreprod48.6.1299. Erratum in: Biol Reprod 1993; 49: 439. [DOI] [PubMed] [Google Scholar]
- 27.Narisawa S, Hecht NB, Goldberg E, Boatright KM, Reed JC, et al. Testis-specific cytochrome c-null mice produce functional sperm but undergo early testicular atrophy. Mol Cell Biol. 2002;22:5554–62. doi: 10.1128/MCB.22.15.5554-5562.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Hüttemann M, Jaradat S, Grossman LI. Cytochrome c oxidase of mammals contains a testes-specific isoform of subunit VIb - The counterpart to testes-specific cytochrome c? Mol Reprod Dev. 2003;66:8–16. doi: 10.1002/mrd.10327. [DOI] [PubMed] [Google Scholar]
- 29.Blanco A, Zinkham WH. Lactate dehydrogenases in human testes. Science. 1963;139:601–2. doi: 10.1126/science.139.3555.601. [DOI] [PubMed] [Google Scholar]
- 30.Goldberg E. Lactic and malic dehydrogenases in human spermatozoa. Science. 1963;139:602–3. doi: 10.1126/science.139.3555.602. [DOI] [PubMed] [Google Scholar]
- 31.Turner RM. Moving to the beat: a review of mammalian sperm motility regulation. Reprod Fertil Dev. 2006;18:25–38. doi: 10.1071/rd05120. [DOI] [PubMed] [Google Scholar]
- 32.Mukai C, Okuno M. Glycolysis plays a major role for adenosine triphosphate supplementation in mouse sperm flagellar movement. Biol Reprod. 2004;71:540–7. doi: 10.1095/biolreprod.103.026054. [DOI] [PubMed] [Google Scholar]
- 33.Albarracín JL, Fernández-Novell JM, Ballester J, Rauch MC, Quintero-Moreno A, et al. Gluconeogenesis-linked glycogen metabolism is important in the achievement of in vitro capacitation of dog spermatozoa in a medium without glucose. Biol Reprod. 2004;71:1437–45. doi: 10.1095/biolreprod.104.029041. [DOI] [PubMed] [Google Scholar]
- 34.Bradley MP, Geelan A, Leitch V, Goldberg E. Cloning, sequencing, and characterization of LDH-C4 from a fox testis cDNA library. Mol Reprod Dev. 1996;44:452–9. doi: 10.1002/(SICI)1098-2795(199608)44:4<452::AID-MRD4>3.0.CO;2-K. [DOI] [PubMed] [Google Scholar]
- 35.Bunch DO, Welch JE, Magyar PL, Eddy EM, O’Brien DA. Glyceraldehyde 3-phosphate dehydrogenase-S protein distribution during mouse spermatogenesis. Biol Reprod. 1998;58:834–41. doi: 10.1095/biolreprod58.3.834. [DOI] [PubMed] [Google Scholar]
- 36.Mori C, Nakamura N, Welch JE, Gotoh H, Goulding EH, et al. Mouse spermatogenic cell-specific type 1 hexokinase (mHk1-s) transcripts are expressed by alternative splicing from the mHk1 gene and the HK1-S protein is localized mainly in the sperm tail. Mol Reprod Dev. 1998;49:374–85. doi: 10.1002/(SICI)1098-2795(199804)49:4<374::AID-MRD4>3.0.CO;2-K. [DOI] [PubMed] [Google Scholar]
- 37.Travis AJ, Foster JA, Rosenbaum NA, Visconti PE, Gerton GL, et al. Targeting of a germ cell-specific type 1 hexokinase lacking a porin-binding domain to the mitochondria as well as to the head and fibrous sheath of murine spermatozoa. Mol Biol Cell. 1998;9:263–76. doi: 10.1091/mbc.9.2.263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hunter RH. Fallopian tube physiology: preliminaries to monospermic fertilization and cellular events post-fertilization. Ernst Schering Res Found Workshop. 2005;52:245–61. doi: 10.1007/3-540-27147-3_11. [DOI] [PubMed] [Google Scholar]
- 39.Wassarman PM. Mammalian fertilization: molecular aspects of gamete adhesion, exocytosis, and fusion. Cell. 1999;96:175–83. doi: 10.1016/s0092-8674(00)80558-9. [DOI] [PubMed] [Google Scholar]
- 40.Paoli D, Gallo M, Rizzo F, Baldi E, Francavilla S, et al. Mitochondrial membrane potential profile and its correlation with increasing sperm motility. Fertil Steril. 2011;95:2315–9. doi: 10.1016/j.fertnstert.2011.03.059. [DOI] [PubMed] [Google Scholar]
- 41.Suarez SS, Ho HC. Hyperactivation of mammalian sperm. Cell Mol Biol (Noisy-le-grand) 2003;49:351–6. [PubMed] [Google Scholar]
- 42.Curtis MP, Kirkman-Brown JC, Connolly TJ, Gaffney EA. Modelling a tethered mammalian sperm cell undergoing hyperactivation. J Theor Biol. 2012;309:1–10. doi: 10.1016/j.jtbi.2012.05.035. [DOI] [PubMed] [Google Scholar]
- 43.Yanagimachi R. The movement of golden hamster spermatozoa before and after capacitation. J Reprod Fertil. 1970;23:193–6. doi: 10.1530/jrf.0.0230193. [DOI] [PubMed] [Google Scholar]
- 44.Ishijima S, Baba SA, Mohri H, Suarez SS. Quantitative analysis of flagellar movement in hyperactivated and acrosome-reacted golden hamster spermatozoa. Mol Reprod Dev. 2002;61:376–84. doi: 10.1002/mrd.10017. [DOI] [PubMed] [Google Scholar]
- 45.Vigue C, Vigue L, Huszar G. Adenosine triphosphate (ATP) concentrations and ATP/adenosine diphosphate ratios in human sperm of normospermic, oligospermic, and asthenospermic specimens and in their swim-up fractions: lack of correlation between ATP parameters and sperm creatine kinase concentrations. J Androl. 1992;13:305–11. [PubMed] [Google Scholar]
- 46.Kamp G, Schmidt H, Stypa H, Feiden S, Mahling C, et al. Regulatory properties of 6-phosphofructokinase and control of glycolysis in boar spermatozoa. Reproduction. 2007;133:29–40. doi: 10.1530/REP-06-0082. [DOI] [PubMed] [Google Scholar]
- 47.Inaba K. Molecular architecture of the sperm flagella: molecules for motility and signaling. Zoolog Sci. 2003;20:1043–56. doi: 10.2108/zsj.20.1043. [DOI] [PubMed] [Google Scholar]
- 48.Summers KE, Gibbons IR. Adenosine triphosphate-induced sliding of tubules in trypsin-treated flagella of sea-urchin sperm. Proc Natl Acad Sci USA. 1971;68:3092–6. doi: 10.1073/pnas.68.12.3092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Shingyoji C, Murakami A, Takahashi K. Local reactivation of Triton-extracted flagella by iontophoretic application of ATP. Nature. 1977;265:269–70. doi: 10.1038/265269a0. [DOI] [PubMed] [Google Scholar]
- 50.Brokaw CJ. Calcium sensors in sea urchin sperm flagella. Cell Motil Cytoskeleton. 1991;18:123–30. doi: 10.1002/cm.970180207. [DOI] [PubMed] [Google Scholar]
- 51.Okuno M, Hiramoto Y. Mechanical stimulation of starfish sperm flagella. J Exp Biol. 1976;65:401–13. doi: 10.1242/jeb.65.2.401. [DOI] [PubMed] [Google Scholar]
- 52.Gibbons IR, Shingyoji C, Murakami A, Takahashi K. Spontaneous recovery after experimental manipulation of the plane of beat in sperm flagella. Nature. 1987;325:351–2. doi: 10.1038/325351a0. [DOI] [PubMed] [Google Scholar]
- 53.Shingyoji C, Yoshimura K, Eshel D, Takahashi K, Gibbons IR. Effect of beat frequency on the velocity of microtubule sliding in reactivated sea urchin sperm flagella under imposed head vibration. J Exp Biol. 1995;198:645–53. doi: 10.1242/jeb.198.3.645. [DOI] [PubMed] [Google Scholar]
- 54.Brokaw CJ. Calcium-induced asymmetrical beating of triton-demembranated sea urchin sperm flagella. J Cell Biol. 1979;82:401–11. doi: 10.1083/jcb.82.2.401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Burgess SA, Walker ML, Sakakibara H, Knight PJ, Oiwa K. Dynein structure and power stroke. Nature. 2003;421:715–8. doi: 10.1038/nature01377. [DOI] [PubMed] [Google Scholar]
- 56.Serohijos AW, Chen Y, Ding F, Elston TC, Dokholyan NV. A structural model reveals energy transduction in dynein. Proc Natl Acad Sci USA. 2006;103:18540–5. doi: 10.1073/pnas.0602867103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Nevo AC, Rikmenspoel R. Diffusion of ATP in sperm flagella. J Theor Biol. 1970;26:11–8. doi: 10.1016/s0022-5193(70)80027-3. [DOI] [PubMed] [Google Scholar]
- 58.Adam DE, Wei J. Mass transport of ATP within the motile sperm. J Theor Biol. 1975;49:125–45. doi: 10.1016/s0022-5193(75)80023-3. [DOI] [PubMed] [Google Scholar]
- 59.Troiano L, Granata AR, Cossarizza A, Kalashnikova G, Bianchi R, et al. Mitochondrial membrane potential and DNA stainability in human sperm cells: a flow cytometry analysis with implications for male infertility. Exp Cell Res. 1998;241:384–93. doi: 10.1006/excr.1998.4064. [DOI] [PubMed] [Google Scholar]
- 60.Donnelly ET, O’Connell M, McClure N, Lewis SE. Differences in nuclear DNA fragmentation and mitochondrial integrity of semen and prepared human spermatozoa. Hum Reprod. 2000;15:1552–61. doi: 10.1093/humrep/15.7.1552. [DOI] [PubMed] [Google Scholar]
- 61.Kasai T, Ogawa K, Mizuno K, Nagai S, Uchida Y, et al. Relationship between sperm mitochondrial membrane potential, sperm motility, and fertility potential. Asian J Androl. 2002;4:97–103. [PubMed] [Google Scholar]
- 62.Wang X, Sharma RK, Gupta A, George V, Thomas AJ, et al. Alterations in mitochondria membrane potential and oxidative stress in infertile men: a prospective observational study. Fertil Steril. 2003;80(Suppl 2):844–50. doi: 10.1016/s0015-0282(03)00983-x. [DOI] [PubMed] [Google Scholar]
- 63.Luft R. The development of mitochondrial medicine. Biochim Biophys Acta. 1995;1271:1–6. doi: 10.1016/0925-4439(95)00002-l. [DOI] [PubMed] [Google Scholar]
- 64.Tombes RM, Shapiro BM. Enzyme termini of a phosphocreatine shuttle. Purification and characterization of two creatine kinase isozymes from sea urchin sperm. J Biol Chem. 1987;262:16011–9. [PubMed] [Google Scholar]
- 65.Oberholzer M, Bregy P, Marti G, Minca M, Peier M, et al. Trypanosomes and mammalian sperm: one of a kind? Trends Parasitol. 2007;23:71–7. doi: 10.1016/j.pt.2006.12.002. [DOI] [PubMed] [Google Scholar]
- 66.Rodriguez-Gil JE. Mammalian sperm energy resources management and survival during conservation in refrigeration. Reprod Domest Anim. 2006;41(Suppl 2):11–20. doi: 10.1111/j.1439-0531.2006.00765.x. [DOI] [PubMed] [Google Scholar]
- 67.Williams AC, Ford WC. The role of glucose in supporting motility and capacitation in human spermatozoa. J Androl. 2001;22:680–95. [PubMed] [Google Scholar]
- 68.Frenette G, Thabet M, Sullivan R. Polyol pathway in human epididymis and semen. J Androl. 2006;27:233–9. doi: 10.2164/jandrol.05108. [DOI] [PubMed] [Google Scholar]
- 69.Urner F, Sakkas D. Characterization of glycolysis and pentose phosphate pathway activity during sperm entry into the mouse oocyte. Biol Reprod. 1999;60:973–8. doi: 10.1095/biolreprod60.4.973. [DOI] [PubMed] [Google Scholar]
- 70.Mann T, Lutwak-Mann C. Berlin: Springer Verlag; 1981. Male Reproductive Function and Semen; pp. 1–34. [Google Scholar]
- 71.Mohri H, Suter DA, Brown-Woodman PD, White IG, et al. Identification of the biochemical lesion produced by alpha-chlorohydrin in spermatozoa. Nature. 1975;255:75–7. doi: 10.1038/255075a0. [DOI] [PubMed] [Google Scholar]
- 72.Welch JE, Brown PL, O’Brien DA, Magyar PL, Bunch DO, et al. Human glyceraldehyde 3-phosphate dehydrogenase-2 gene is expressed specifically in spermatogenic cells. J Androl. 2000;21:328–38. [PubMed] [Google Scholar]
- 73.Odet F, Duan C, Willis WD, Goulding EH, Kung A, et al. Expression of the gene for mouse lactate dehydrogenase C (Ldhc) is required for male fertility. Biol Reprod. 2008;79:26–34. doi: 10.1095/biolreprod.108.068353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Hung PH, Miller MG, Meyers SA, VandeVoort CA. Sperm mitochondrial integrity is not required for hyperactivated motility, zona binding, or acrosome reaction in the rhesus macaque. Biol Reprod. 2008;79:367–75. doi: 10.1095/biolreprod.107.066357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Hereng TH, Elgstøen KB, Cederkvist FH, Eide L, Jahnsen T, et al. Exogenous pyruvate accelerates glycolysis and promotes capacitation in human spermatozoa. Hum Reprod. 2011;26:3249–63. doi: 10.1093/humrep/der317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Yanagimachi R. Fertility of mammalian spermatozoa: its development and relativity. Zygote. 1994;2:371–2. doi: 10.1017/s0967199400002240. [DOI] [PubMed] [Google Scholar]
- 77.de Lamirande E, Leclerc P, Gagnon C. Capacitation as a regulatory event that primes spermatozoa for the acrosome reaction and fertilization. Mol Hum Reprod. 1997;3:175–94. doi: 10.1093/molehr/3.3.175. [DOI] [PubMed] [Google Scholar]
- 78.Burkman LJ. Characterization of hyperactivated motility by human spermatozoa during capacitation: comparison of fertile and oligozoospermic sperm populations. Arch Androl. 1984;13:153–65. doi: 10.3109/01485018408987514. [DOI] [PubMed] [Google Scholar]
- 79.Buffone MG, Calamera JC, Verstraeten SV, Doncel GF. Capacitation-associated protein tyrosine phosphorylation and membrane fluidity changes are impaired in the spermatozoa of asthenozoospermic patients. Reproduction. 2005;129:697–705. doi: 10.1530/rep.1.00584. [DOI] [PubMed] [Google Scholar]
- 80.Ho HC, Granish KA, Suarez SS. Hyperactivated motility of bull sperm is triggered at the axoneme by Ca2+ and not cAMP. Dev Biol. 2002;250:208–17. doi: 10.1006/dbio.2002.0797. [DOI] [PubMed] [Google Scholar]
- 81.Kirichok Y, Navarro B, Clapham DE. Whole-cell patch-clamp measurements of spermatozoa reveal an alkaline-activated Ca2+channel. Nature. 2006;439:737–40. doi: 10.1038/nature04417. [DOI] [PubMed] [Google Scholar]
- 82.Franklin LE. Formation of the redundant nuclear envelope in monkey spermatids. Anat Rec. 1968;161:149–61. doi: 10.1002/ar.1091610202. [DOI] [PubMed] [Google Scholar]
- 83.Toshimori K, Higashi R, Oura C. Distribution of intramembranous particles and filipin-sterol complexes in mouse sperm membranes: polyene antibiotic filipin treatment. Am J Anat. 1985;174:455–70. doi: 10.1002/aja.1001740408. [DOI] [PubMed] [Google Scholar]
- 84.Suarez SS, Dai X. Intracellular calcium reaches different levels of elevation in hyperactivated and acrosome-reacted hamster sperm. Mol Reprod Dev. 1995;42:325–33. doi: 10.1002/mrd.1080420310. [DOI] [PubMed] [Google Scholar]
- 85.Ficarro S, Chertihin O, Westbrook VA, White F, Jayes F, et al. Phosphoproteome analysis of capacitated human sperm. Evidence of tyrosine phosphorylation of a kinase-anchoring protein 3 and valosin-containing protein/p97 during capacitation. J Biol Chem. 2003;278:11579–89. doi: 10.1074/jbc.M202325200. [DOI] [PubMed] [Google Scholar]
- 86.Kota V, Rai P, Weitzel JM, Middendorff R, Bhande SS, et al. Role of glycerol-3-phosphate dehydrogenase 2 in mouse sperm capacitation. Mol Reprod Dev. 2010;77:773–83. doi: 10.1002/mrd.21218. [DOI] [PubMed] [Google Scholar]
- 87.Harper CV, Publicover SJ. Reassessing the role of progesterone in fertilization – Compartmentalized calcium signalling in human spermatozoa? Hum Reprod. 2005;20:2675–80. doi: 10.1093/humrep/dei158. [DOI] [PubMed] [Google Scholar]
- 88.Roveri A, Casasco A, Maiorino M, Dalan P, Calligaro A, et al. Phospholipid hydroperoxide glutathione peroxidase of rat testis. Gonadotropin dependence and immunocytochemical identification. J Biol Chem. 1992;267:6142–6. [PubMed] [Google Scholar]
- 89.Mitra K, Shivaji S. Proteins implicated in sperm capacitation. Indian J Exp Biol. 2005;43:1001–15. [PubMed] [Google Scholar]
- 90.Hoshi K, Tsukikawa S, Sato A. Importance of Ca[2]+, K+ and glucose in the medium for sperm penetration through the human zona pellucida. Tohoku J Exp Med. 1991;165:99–104. doi: 10.1620/tjem.165.99. [DOI] [PubMed] [Google Scholar]
- 91.Ruiz-Pesini E, Díez-Sánchez C, López-Pérez MJ, Enríquez JA. The role of the mitochondrion in sperm function: Is there a place for oxidative phosphorylation or is this a purely glycolytic process? Curr Top Dev Biol. 2007;77:3–19. doi: 10.1016/S0070-2153(06)77001-6. [DOI] [PubMed] [Google Scholar]
- 92.Nichol R, Hunter RH, Gardner DK, Leese HJ, Cooke GM. Concentrations of energy substrates in oviductal fluid and blood plasma of pigs during the peri-ovulatory period. J Reprod Fertil. 1992;96:699–707. doi: 10.1530/jrf.0.0960699. [DOI] [PubMed] [Google Scholar]
- 93.Restall BJ, Wales RG. The fallopian tube of the sheep. 3. The chemical composition of the fluid from the fallopian tube. Aust J Biol Sci. 1966;19:687–98. doi: 10.1071/bi9660687. [DOI] [PubMed] [Google Scholar]
- 94.Hutson SM, Van Dop C, Lardy HA. Mitochondrial metabolism of pyruvate in bovine spermatozoa. J Biol Chem. 1977;252:1309–15. [PubMed] [Google Scholar]
- 95.Urner F, Sakkas D. Glucose participates in sperm-oocyte fusion in the mouse. Biol Reprod. 1996;55:917–22. doi: 10.1095/biolreprod55.4.917. [DOI] [PubMed] [Google Scholar]
- 96.Boell EJ. Oxygen consumption of mouse sperm and its relationship to capacitation. J Exp Zool. 1985;234:105–16. doi: 10.1002/jez.1402340113. [DOI] [PubMed] [Google Scholar]
- 97.Fraser LR, Lane MR. Capacitation- and fertilization-related alterations in mouse sperm oxygen consumption. J Reprod Fertil. 1987;81:385–93. doi: 10.1530/jrf.0.0810385. [DOI] [PubMed] [Google Scholar]
- 98.Stendardi A, Focarelli R, Piomboni P, Palumberi D, Serafini F, et al. Evaluation of mitochondrial respiratory efficiency during in vitro capacitation of human spermatozoa. Int J Androl. 2011;34:247–55. doi: 10.1111/j.1365-2605.2010.01078.x. [DOI] [PubMed] [Google Scholar]