

Abstract
Background:
Oxidative stress-induced cell damage is common in the etiology of several neurobiological disorders, including Alzheimer's disease and Parkinson's disease. In a case study, nobiletin-rich Citrus reticulata peels could prevent the progression of cognitive impairment in donepezil-preadministered Alzheimer's disease patients.
Objective:
In this study, we investigated the effects and underlying mechanism of nobiletin and Citrus unshiu immature peel (CUIP) water extract, which contains nobiletin as a major compound, on hydrogen peroxide-induced oxidative stress in HT22 cells, a murine hippocampal neuronal model.
Materials and Methods:
HT22 cells were treated with hydrogen peroxide in the presence or absence of various concentrations of CUIP and nobiletin. Cytotoxicity and apoptotic protein levels were measured by 3-(4,5-dimethythiazol-2-yl)-2,5-diphenyl tetrazolium bromide assay and Western blotting.
Results:
Pretreatment with CUIP and nobiletin inhibited cell death due to hydrogen peroxide. Hydrogen peroxide-induced the expression of phospho-Jun N-terminal kinases (p-JNK) and p-p38 proteins in HT22 cells; however CUIP and nobiletin suppressed p-JNK and p-p38 without changing JNK or p38. Regarding apoptosis, caspase 3, B-cell lymphoma 2 (Bcl-2), and Bax protein expression was determined. CUIP and nobiletin suppressed caspase 3 and Bax expression, but they induced Bcl-2 expression in HT22 cells.
Conclusion:
These results show that CUIP and nobiletin can protect against hydrogen peroxide-induced cell death in HT22 neurons via mitogen-activated protein kinases and apoptotic pathways.
Keywords: Citrus unshiu immature peel, neuroprotective effect, nobiletin, oxidative stress
INTRODUCTION
Oxidative stress-induced cell damage is common in the etiology of several neurobiological disorders, such as ischemia, Alzheimer's disease, and Parkinson's disease.[1,2,3] In these pathological processes, oxidative stress leads to many biological consequences, including cell death, which involves the mitogen-activated protein kinases (MAPKs) and apoptotic signals.[4] H2O2 has also been shown to activate several intracellular signaling pathways, including poly (adenosine diphosphate-ribose) polymerase in response to DNA damage,[5] nuclear factor-κB,[6] activator protein 1.[7] HT22 cells, an immortalized mouse hippocampal cell line, have been widely used as in vitro model for studying the mechanism of oxidative stress-induced neuronal cell death.[8,9]
Nobiletin is a polymethoxyflavone. It is well-known that nobiletin has biological activities such as anti-inflammatory effects, anti-carcinoma effects, and ameliorating scratching behavior in mice.[10,11,12] In a recently report, nobiletin rescued memory deterioration in Alzheimer's disease model rats by restoring β-amyloid-impaired cAMP response element binding protein phosphorylation.[13] Citrus unshiu peel is the dried peel of C. unshiu Markovich or C. reticulata Blanco. It has been used as traditional medicine in Korea, China, and Japan and also divided as immature peel or mature peel. C. unshiu immature peel (CUIP) (Cheong pi or Chung-pi in Korea, Qing pi in China, Jyōhi in Japan) has been used for relief of pleuralgia caused by liver disease and remedy of indigestion. On the other hand, C. unshiu mature peel (Jin pi in Korea, Chen pi in china, Chinpi in Japan) has been used for improving bronchial, asthmatic conditions, cardiac and blood circulation in these countries.[14,15] CUIP and mature peel contains flavonoids including hesperidin, rutin, nobiletin, and naringenin.[16] In a case study, nobiletin-rich C. reticulata peels could prevent the progression of cognitive impairment in donepezil-preadministered Alzheimer's disease patients.[17] Moreover, there are evidences that nobiletin content in CUIP is higher than C. unshiu mature peel.[18,19] However, the mechanisms of action of CUIP and nobiletin's neuroprotective efficacy against oxidative stress in HT22 cells have not been investigated. These encouraged us to focus on the relationship between CUIP and neuroprotective effect of nobiletin. In this study, we investigated how CUIP and nobiletin inhibited hydrogen peroxide-induced cell death in HT22 cells through MAPK activity, and the Bax, and Bcl-2 pathway.
MATERIALS AND METHODS
Materials
Nobiletin, hesperidin, and H2O2 (30%) were purchased from Sigma (St. Louis, MO, USA). Fetal bovine serum (FBS), Dulbecco's modified Eale's medium (DMEM) ⇒ erase, penicillin/streptomycin were obtained from Gibco (NY, USA). Cell proliferation assay solution was obtained from Promega (Madison, USA). phospho-Jun N-terminal kinases (p-JNK), JNK, p-p38, p38, caspase-3, and horseradish peroxidase-linked anti-mouse and rabbit antibodies were purchased from cell signaling technology (Danvers, MA, USA). Bax, Bcl-2, and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) were purchased from Assay Designs (MI, USA).
Preparation of Citrus unshiu immature peel
Dried CUIP was provided by JBT Co. Ltd., in Jangheung, Republic of Korea. It was identified by Dr. Yeo (Medicinal Crop Seed Supply Center, Jangheung, Republic of Korea). The quality CUIP (100g) was boiled in 1 L distilled water for 3 h. The water extract was filtered and then removed under reduced pressure and lyophilized. In total, 12.4 g of extract powder of CUIP was recovered and kept at −20°C (JKTM-13-02). The powder was redissolved in distilled water (200 mg/mL) and stored in a freezer.
Cell line and culture
The HT22 cells, a murine hippocampal cell line, were provided by Dr. Soo Hyun Park (Jeonnam National University, Republic of Korea). HT22 cells were cultured in DMEM, supplemented with 10% heat-inactivated FBS, 100 U/mL penicillin, and 100 µg/mL streptomycin and were maintained in a humidified incubator with 5% CO2. In all experiments, cells were incubated in the presence of the indicated concentrations of CUIP or nobiletin for 1 h before the addition of H2O2 (250 µM).
Cell viability assay
Cell viability was assessed using the 3-(4,5-dimethythiazol-2-yl)-2,5-diphenyl tetrazolium bromide assay (CellTiter96 Aqueous One Solution, Promega) according to the manufacturer's protocol. Briefly, HT22 cells were plated in 96-well culture plates (1 × 104 cells/well). After 24 h, the medium was changed, and the cells were treated with CUIP or nobiletin at various concentrations for 1 h. The plates were then cultured for 24 h with 250 µM H2O2 or not. Then, 20 µL MTS (5.0 µg/µL) was added to each well for an additional 4 h of incubation at 37°C. Absorbance was measured at 490 nm with a microplate reader (infinite M200 Pro, Tecan Group Ltd. Salzburg, Austria).
Western blot analysis
Cells were lysed with radioimmunoprecipitation assay buffer (150 mM sodium chloride, 1 mM ethylenediaminetetraacetic acid, 1 mM ethylene glycol tetraacetic acid, 1.2% Triton X-100, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulfate (SDS), and 50 mM Tris-HCl, pH 7.4) with protease inhibitor cocktail (Sigma). Total protein (40 µg) was separated by 10% or 12% SDS-polyacrylamide gel electrophoresis and transferred to a polyvinylidene fluoride membrane. Western blot procedures were performed routinely. pJNK, JNK, p-p38, p38, caspase3, Bax, Bcl-2, and GAPDH were detected with the respective antibodies and a chemiluminescence detector (LiCor 2600, Lincoln, NE, USA) after enhanced chemiluminescence treatment.
High-performance liquid chromatography analysis of nobiletin in Citrus unshiu immature peel
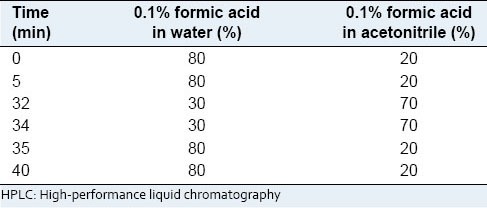
To assay nobiletin and hesperidin in CUIP, we performed high-performance liquid chromatography (HPLC) (Shimadzu, Japan) by using a diode array detector. Analytical HPLC was run on a Luna C-18 reverse phase column, 100A, 25 cm × 4.6 mm, 5 mm. The mobile phase consisted of two solvents, water (0.1% formic acid) and acetonitrile. Table 1. shows the ratio of mobile phases for 40 min; the flow rate was 1 mL/min and the injection volume was 10 mL. 10 mg/mL CUIP, 100 mg/mL nobiletin, and hesperidin were used for HPLC.
Table 1.
The ratio of mobile phases for 40 min for HPLC
Statistics
The data are expressed as means ± standard deviation. Statistical analyses used student's t-test, assuming unequal variances. Values of P < 0.05 were considered to indicate statistical significance.
RESULTS
Cell viability with Citrus unshiu immature peel and nobiletin
HT22 cells were treated for 24 h with different concentrations of CUIP or nobiletin. Neither CUIP nor nobiletin showed cytotoxicity at the concentrations tested. When HT22 cells were treated with 250 µM H2O2, cell viability was decreased by 84% (P < 0.05). However, cell viability with 1 mg/mL CUIP or 50 µM nobiletin pretreatment were 95.8% and 97.02%, respectively. Thus, pretreatment with CUIP and nobiletin could protect against cell death induced by oxidative stress in HT22 cells [Figure 1].
Figure 1.

Effects of Citrus unshiu immature peel and nobiletin on H2O2-induced cytotoxicity in HT22 cells. (a) Cell viability was reduced by H2O2, dose-dependently. (b) C. unshiu immature peel and nobiletin protected the cell death by 250 µM H2O2. Data are presented as means ± S.D. from three experiments. *P < 0.05, **P<0.005; Blank vs. H2O2 treated group, #P <0.05; H2O2 treated group vs. CUIP or nobiletin treated group with H2O2
Inhibitory effects of Citrus unshiu immature peel and nobiletin on phosphorylation of Jun N-terminal kinases and p38
In the previous studies, H2O2 induced phosphorylation of MAPKs.[4] In this study, pJNK and p-p38 were overexpressed with 250 µM H2O2 treatment for 6 h. However, CUIP or nobiletin pretreatment inhibited the expression of phosphorylation of JNK and p38 without changing total JNK and p38 expression. These result showed that CUIP and nobiletin could affect the MAPKs pathway in protecting HT22 cells against death [Figure 2].
Figure 2.

Effects of Citrus unshiu immature peel and nobiletin on H2O2-induced phosphorylation of Jun N-terminal kinases and p38 in HT22 cells. (a) Protein bands and (b) relative intensities of protein bands. Cells were treated with C. unshiu immature peel and nobiletin 1 h after 250 µM H2O2 was added to the cells and incubated for 6 h. Then, 30 µg cell lysates were subjected to Western blot analysis for phospho-Jun N-terminal kinases, Jun N-terminal kinases, pp38, and p38 with monoclonal antibodies
Inhibitory effects of Citrus unshiu immature peel and nobiletin on apoptosis
To investigate mitochondrial dysfunction associated apoptosis in HT22 cells due to CUIP and nobiletin, cells were pretreated with CUIP (1 mg/mL) or nobiletin (50 µM) for 1 h and then 250 µM H2O2 was added for 24 h to activate the caspase-3, Bax, and Bcl-2 pathway. H2O2 activated caspase-3 and Bax but suppressed Bcl-2 expression. However, CUIP and nobiletin inhibited caspase-3 and Bax expression but activated Bcl-2 expression [Figure 3]. These results indicated that CUIP and nobiletin can protect the cells against death through mitochondria-associated apoptosis after oxidative stress.
Figure 3.

Effects of Citrus unshiu immature peel and nobiletin on H2O2-induced Bax, Bcl-2, and caspase-3 in HT22 cells. (a) Protein bands and (b) relative intensities of protein bands. Cells were treated with C. unshiu immature peel and nobiletin 1 h after 250 µM H2O2 was added to the cells and incubated for 24 h. Then, 30 µg cell lysates were subjected to Western blot analysis for Bax, Bcl-2, and capase-3 with monoclonal antibodies. glyceraldehyde 3-phosphate dehydrogenase was used as a control
Identification of nobiletin and hesperidin in Citrus unshiu immature peel by high-performance liquid chromatography
To identify nobiletin and hesperidin in CUIP, we performed HPLC analysis using a C18 cartridge column and eluted with increasing concentrations of acetonitrile. HPLC revealed that the yield of nobiletin and hesperidin from CUIP was 0.64% and 1.41%, respectively [Figure 4].
Figure 4.

Analysis of nobiletin and hesperidin in Citrus unshiu immature peel by high-performance liquid chromatography at 280 nm. (a) Standard of hesperidin; (b) standard of nobiletin; (c) C. unshiu immature peel in methanol
DISCUSSION
In the present study, we found that CUIP and nobiletin prevent the H2O2-induced cell death through inhibiting JNK and p38 phosphorylation, caspase-3 and Bax activation, and activating Bcl-2 suppression in HT22 cells.
Jun N-terminal kinases and p38 are major members of the MAPKs, a family of serine/threonine protein kinases, and involved in many cellular processes, including cell growth, differentiation, inflammation, and cell death.[20] Inhibition of pJNK and p-p38 pathway by diverse stimulators such as glutamate, amyloid beta, and H2O2 is involved in neuroprotective effects.[4,21,22] In this study, we found that H2O2 induced phosphorylation of JNK and p38 in HT22 cells; however, pretreatment with CUIP and nobiletin inhibited the phosphorylation of JNK and p38. These results accorded with the previous study, which showed nobiletin modulated p38 and JNK in PC12 cells challenged by H2O2.[23] In this study, we reconfirmed that the neuroprotective effect of nobiletin involved in MAPKs pathway also CUIP protected against cell death by oxidative stress through inhibiting the phosphorylation of p38 and JNK.
Because, apoptosis is known to be one the most sensitive biological markers for evaluating oxidative stress, representing an imbalance between free radical generation and the efficacy of the antioxidant system,[24,25] we determined whether effects on apoptosis-related proteins could be seen in H2O2-induced apoptosis in HT22 cells. H2O2 activates the “intrinsic apoptosis” or mitochondrial apoptotic pathway. When Bcl-2 family proteins translocate to mitochondria, intrinsic apoptosis are initiated. Bax and Bcl-2 belong to the Bcl-2 family. Bax plays as a pro-apoptotic protein. On the other hand, Bcl2 plays as anti-apoptotic protein.[26] Bax translocate to the mitochondrial outer membrane, where it oligomerizes or forms complexes with Bid or Bad. Bax oligomers facilitate the release of proapoptotic molecules such as cytochrome c or Smac from mithochondria membrane. Cytochrome c leads caspase-9 activation result in caspase-3 activation.[27] In this study, we observed CUIP and nobiletin activated Bcl-2 expression but inhibited caspase-3 and Bax expression against H2O2 stimulation in HT22 cells. In the previous studies, nobiletin also regulated caspase-3 levels in PC12 cells stimulated by H2O2.[23,28] In this study, we showed that CUIP and nobiletin can protect against H2O2-induced oxidative stress in HT22 cells by upregulating Bcl-2, and an anti-apoptotic protein downregulating cleaved caspase-3 and Bax.
Hesperidin and nobiletin are well known main flavonoids of Citrus species. Zhang et al., reported 95% ethanol extract of citrus peel, purchased from a local factory in Jinhua, Zhejiang, China in November 2004, contained 2.81 ± 0.09% hesperidin and 0.27 ± 0.03% nobiletin.[29] In another report, Lu et al., analyzed flavonoids in fresh fruit and traditional chinese medicine ingredients of 27 citrus and three not-citrus cultivars from Zhejiang, Fujian and Jiangxi Province in China. According to the report, fresh peel of C. unshiu contained 5.86% and 6.25% hesperidin and 0.01% and 0.02% nobiletin from C. unshiu peel.[18] Choi et al., analyzed flavonoids in immature peel and mature of C. unshiu grown on Jeju Island in Korea. They reported that CUIP ethanol extract contained 17.23 ± 0.34% hesperidin and 1.21 ± 0.48% nobiletin. However, its mature peel ethanol extract had 4.47 ± 1.29% hesperidin and 0.75 ± 0.00% nobiletin.[19] These reports showed that the flavonoid in C. unshiu were different by collecting time or growing place. Also immature peel of C. unshiu contained hesperidin and nobiletin more than mature peel. In our study, we detected 1.41% hesperidin and 0.64% nobiletin in CUIP water extract. Because hesperidin and nobiletin are hard to be extracted by hot water, contents of these compounds were detected lower in other reports.
CONCLUSION
In conclusion, CUIP and nobiletin protected against cell death induced by H2O2 in HT22 cells through inhibiting the activation of JNK, p38 MAPKs, caspase-3, and Bax but upregulation of Bcl-2. These results suggest that CUIP and nobiletin may be useful to protect against neurodegenerative disorders involving oxidative stress. Moreover nobiltein may play an active compound of CUIP for preventing the cell death in HT22 by H2O2. However, further studies are required to clarify the anti-apoptotic mechanisms of CUIP and nobiletin.
ACKNOWLEDGEMENTS
This research was financially supported by the Ministry Of Trade, Industry and Energy (MOTIE) and Korea Institute for Advancement of Technology(KIAT) through the Inter-ER Cooperation Projects (R0002020).
Footnotes
Source of Support: Nil
Conflict of Interest: None declared.
REFERENCES
- 1.Allen CL, Bayraktutan U. Oxidative stress and its role in the pathogenesis of ischaemic stroke. Int J Stroke. 2009;4:461–70. doi: 10.1111/j.1747-4949.2009.00387.x. [DOI] [PubMed] [Google Scholar]
- 2.Gackowski D, Rozalski R, Siomek A, Dziaman T, Nicpon K, Klimarczyk M, et al. Oxidative stress and oxidative DNA damage is characteristic for mixed Alzheimer disease/vascular dementia. J Neurol Sci. 2008;266:57–62. doi: 10.1016/j.jns.2007.08.041. [DOI] [PubMed] [Google Scholar]
- 3.Zuo L, Motherwell MS. The impact of reactive oxygen species and genetic mitochondrial mutations in Parkinson's disease. Gene. 2013;532:18–23. doi: 10.1016/j.gene.2013.07.085. [DOI] [PubMed] [Google Scholar]
- 4.Zhao ZY, Luan P, Huang SX, Xiao SH, Zhao J, Zhang B, et al. Edaravone protects HT22 neurons from H2O2-induced apoptosis by inhibiting the MAPK signaling pathway. CNS Neurosci Ther. 2013;19:163–9. doi: 10.1111/cns.12044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Xu H, Luo P, Zhao Y, Zhao M, Yang Y, Chen T, et al. Iduna protects HT22 cells from hydrogen peroxide-induced oxidative stress through interfering poly (ADP-ribose) polymerase-1-induced cell death (parthanatos) Cell Signal. 2013;25:1018–26. doi: 10.1016/j.cellsig.2013.01.006. [DOI] [PubMed] [Google Scholar]
- 6.Ishimura A, Ishige K, Taira T, Shimba S, Ono S, Ariga H, et al. Comparative study of hydrogen peroxide- and 4-hydroxy-2-nonenal-induced cell death in HT22 cells. Neurochem Int. 2008;52:776–85. doi: 10.1016/j.neuint.2007.09.008. [DOI] [PubMed] [Google Scholar]
- 7.Meyer M, Schreck R, Baeuerle PA. H2O2 and antioxidants have opposite effects on activation of NF-kappa B and AP-1 in intact cells: AP-1 as secondary antioxidant-responsive factor. EMBO J. 1993;12:2005–15. doi: 10.1002/j.1460-2075.1993.tb05850.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Heo SJ, Cha SH, Kim KN, Lee SH, Ahn G, Kang DH, et al. Neuroprotective effect of phlorotannin isolated from Ishige okamurae against H2O2-induced oxidative stress in murine hippocampal neuronal cells, HT22. Appl Biochem Biotechnol. 2012;166:1520–32. doi: 10.1007/s12010-012-9545-7. [DOI] [PubMed] [Google Scholar]
- 9.Kang SM, Cha SH, Ko JY, Kang MC, Kim D, Heo SJ, et al. Neuroprotective effects of phlorotannins isolated from a brown alga, Ecklonia cava, against H2O2-induced oxidative stress in murine hippocampal HT22 cells. Environ Toxicol Pharmacol. 2012;34:96–105. doi: 10.1016/j.etap.2012.03.006. [DOI] [PubMed] [Google Scholar]
- 10.Lin N, Sato T, Takayama Y, Mimaki Y, Sashida Y, Yano M, et al. Novel anti-inflammatory actions of nobiletin, a citrus polymethoxy flavonoid, on human synovial fibroblasts and mouse macrophages. Biochem Pharmacol. 2003;65:2065–71. doi: 10.1016/s0006-2952(03)00203-x. [DOI] [PubMed] [Google Scholar]
- 11.Yoshimizu N, Otani Y, Saikawa Y, Kubota T, Yoshida M, Furukawa T, et al. Anti-tumour effects of nobiletin, a citrus flavonoid, on gastric cancer include: Antiproliferative effects, induction of apoptosis and cell cycle deregulation. Aliment Pharmacol Ther. 2004;20(Suppl 1):95–101. doi: 10.1111/j.1365-2036.2004.02082.x. [DOI] [PubMed] [Google Scholar]
- 12.Jang SE, Ryu KR, Park SH, Chung S, Teruya Y, Han MJ, et al. Nobiletin and tangeretin ameliorate scratching behavior in mice by inhibiting the action of histamine and the activation of NF-?B, AP-1 and p38. Int Immunopharmacol. 2013;17:502–7. doi: 10.1016/j.intimp.2013.07.012. [DOI] [PubMed] [Google Scholar]
- 13.Matsuzaki K, Yamakuni T, Hashimoto M, Haque AM, Shido O, Mimaki Y, et al. Nobiletin restoring beta-amyloid-impaired CREB phosphorylation rescues memory deterioration in Alzheimer's disease model rats. Neurosci Lett. 2006;400:230–4. doi: 10.1016/j.neulet.2006.02.077. [DOI] [PubMed] [Google Scholar]
- 14.Pharmacognosy. Paju, Korea: Dongmyung; 2008. The Pharmacognosy Compilation Committee; pp. 409–12. [Google Scholar]
- 15.Densky D, Clavey S, Stoger E, translators. Chinese Herbal Medicine Materia Medica. 3rd ed. WA, USA: Eastland Press, Inc; 2004. pp. 510–2. 514-6. [Google Scholar]
- 16.Kawaii S, Tomono Y, Katase E, Ogawa K, Yano M. Quantitation of flavonoid constituents in citrus fruits. J Agric Food Chem. 1999;47:3565–71. doi: 10.1021/jf990153+. [DOI] [PubMed] [Google Scholar]
- 17.Seki T, Kamiya T, Furukawa K, Azumi M, Ishizuka S, Takayama S, et al. Nobiletin-rich Citrus reticulata peels, a kampo medicine for Alzheimer's disease: A case series. Geriatr Gerontol Int. 2013;13:236–8. doi: 10.1111/j.1447-0594.2012.00892.x. [DOI] [PubMed] [Google Scholar]
- 18.Lu Y, Zhang C, Bucheli P, Wei D. Citrus flavonoids in fruit and traditional Chinese medicinal food ingredients in China. Plant Foods Hum Nutr. 2006;61:57–65. doi: 10.1007/s11130-006-0014-8. [DOI] [PubMed] [Google Scholar]
- 19.Choi SY, Ko HC, Ko SY, Hwang JH, Park JG, Kang SH, et al. Correlation between flavonoid content and the NO production inhibitory activity of peel extracts from various citrus fruits. Biol Pharm Bull. 2007;30:772–8. doi: 10.1248/bpb.30.772. [DOI] [PubMed] [Google Scholar]
- 20.Pearson G, Robinson F, Beers Gibson T, Xu BE, Karandikar M, Berman K, et al. Mitogen-activated protein (MAP) kinase pathways: Regulation and physiological functions. Endocr Rev. 2001;22:153–83. doi: 10.1210/edrv.22.2.0428. [DOI] [PubMed] [Google Scholar]
- 21.Tian X, Sui S, Huang J, Bai JP, Ren TS, Zhao QC. Neuroprotective effects of Arctium lappa L. roots against glutamate-induced oxidative stress by inhibiting phosphorylation of p38, JNK and ERK 1/2 MAPKs in PC12 cells. Environ Toxicol Pharmacol. 2014;38:189–98. doi: 10.1016/j.etap.2014.05.017. [DOI] [PubMed] [Google Scholar]
- 22.Ma B, Meng X, Wang J, Sun J, Ren X, Qin M, et al. Notoginsenoside R1 attenuates amyloid-ß-induced damage in neurons by inhibiting reactive oxygen species and modulating MAPK activation. Int Immunopharmacol. 2014;22:151–9. doi: 10.1016/j.intimp.2014.06.018. [DOI] [PubMed] [Google Scholar]
- 23.Hwang SL, Yen GC. Modulation of Akt, JNK, and p38 activation is involved in citrus flavonoid-mediated cytoprotection of PC12 cells challenged by hydrogen peroxide. J Agric Food Chem. 2009;57:2576–82. doi: 10.1021/jf8033607. [DOI] [PubMed] [Google Scholar]
- 24.Gutteridge JM. Lipid peroxidation and antioxidants as biomarkers of tissue damage. Clin Chem. 1995;41:1819–28. [PubMed] [Google Scholar]
- 25.Kassie F, Parzefall W, Knasmüller S. Single cell gel electrophoresis assay: A new technique for human biomonitoring studies. Mutat Res. 2000;463:13–31. doi: 10.1016/s1383-5742(00)00041-7. [DOI] [PubMed] [Google Scholar]
- 26.Liu NS, Du X, Lu J, He BP. Diva reduces cell death in response to oxidative stress and cytotoxicity. PLoS One. 2012;7:e43180. doi: 10.1371/journal.pone.0043180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wu CC, Bratton SB. Regulation of the intrinsic apoptosis pathway by reactive oxygen species. Antioxid Redox Signal. 2013;19:546–58. doi: 10.1089/ars.2012.4905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lu YH, Su MY, Huang HY, Lin-Li, Yuan CG. Protective effects of the citrus flavanones to PC12 cells against cytotoxicity induced by hydrogen peroxide. Neurosci Lett. 2010;484:6–11. doi: 10.1016/j.neulet.2010.07.078. [DOI] [PubMed] [Google Scholar]
- 29.Zhang C, Lu Y, Tao L, Tao X, Su X, Wei D. Tyrosinase inhibitory effects and inhibition mechanisms of nobiletin and hesperidin from citrus peel crude extracts. J Enzyme Inhib Med Chem. 2007;22:91–8. doi: 10.1080/14756360600988989. [DOI] [PubMed] [Google Scholar]