

Comprehension deficits are common in both Wernicke’s aphasia and semantic aphasia. Thompson et al. compare these groups of patients on a task in which related items are repeatedly presented, increasing semantic competition across repetitions. Both groups show semantic impairment, but only individuals with prefrontal damage show harmful effects of repetition.
Keywords: semantic, Wernicke, aphasia, modality, refractory
Comprehension deficits are common in both Wernicke’s aphasia and semantic aphasia. Thompson et al. compare these groups of patients on a task in which related items are repeatedly presented, increasing semantic competition across repetitions. Both groups show semantic impairment, but only individuals with prefrontal damage show harmful effects of repetition.
Abstract
Comprehension deficits are common in stroke aphasia, including in cases with (i) semantic aphasia, characterized by poor executive control of semantic processing across verbal and non-verbal modalities; and (ii) Wernicke’s aphasia, associated with poor auditory–verbal comprehension and repetition, plus fluent speech with jargon. However, the varieties of these comprehension problems, and their underlying causes, are not well understood. Both patient groups exhibit some type of semantic ‘access’ deficit, as opposed to the ‘storage’ deficits observed in semantic dementia. Nevertheless, existing descriptions suggest that these patients might have different varieties of ‘access’ impairment—related to difficulty resolving competition (in semantic aphasia) versus initial activation of concepts from sensory inputs (in Wernicke’s aphasia). We used a case series design to compare patients with Wernicke’s aphasia and those with semantic aphasia on Warrington’s paradigmatic assessment of semantic ‘access’ deficits. In these verbal and non-verbal matching tasks, a small set of semantically-related items are repeatedly presented over several cycles so that the target on one trial becomes a distractor on another (building up interference and eliciting semantic ‘blocking’ effects). Patients with Wernicke’s aphasia and semantic aphasia were distinguished according to lesion location in the temporal cortex, but in each group, some individuals had additional prefrontal damage. Both of these aspects of lesion variability—one that mapped onto classical ‘syndromes’ and one that did not—predicted aspects of the semantic ‘access’ deficit. Both semantic aphasia and Wernicke’s aphasia cases showed multimodal semantic impairment, although as expected, the Wernicke’s aphasia group showed greater deficits on auditory-verbal than picture judgements. Distribution of damage in the temporal lobe was crucial for predicting the initially ‘beneficial’ effects of stimulus repetition: cases with Wernicke’s aphasia showed initial improvement with repetition of words and pictures, while in semantic aphasia, semantic access was initially good but declined in the face of competition from previous targets. Prefrontal damage predicted the ‘harmful’ effects of repetition: the ability to reselect both word and picture targets in the face of mounting competition was linked to left prefrontal damage in both groups. Therefore, patients with semantic aphasia and Wernicke’s aphasia have partially distinct impairment of semantic ‘access’ but, across these syndromes, prefrontal lesions produce declining comprehension with repetition in both verbal and non-verbal tasks.
Introduction
Semantic cognition allows us to understand the significance of words and objects that we encounter. It encompasses several interacting components within a widely distributed neural network (Patterson et al., 2007; Jefferies, 2013), giving rise to different types of semantic impairment after brain damage (Jefferies and Lambon Ralph, 2006; Robson et al., 2012). Progressive degradation of conceptual representations is observed in semantic dementia following atrophy of the anterior temporal lobes (Snowden et al., 1989; Hodges et al., 1992; Mummery et al., 2000; Galton et al., 2001). In other patients, knowledge is preserved but deficient semantic ‘access’ prevents the effective retrieval of this information (Lambon Ralph, 2014). Patients can show difficulties mapping sensory perception into semantic knowledge, such as the disordered translation of auditory input into semantics in word deafness and Wernicke’s aphasia (Goldblum and Albert, 1972; Tanaka et al., 1987; Poeppel, 2001; Slevc et al., 2011; Robson et al., 2013). Further, there may be a deficit of controlled selection/retrieval of semantic knowledge that is sensitive to task demands, as in semantic aphasia (Jefferies and Lambon Ralph, 2006).
Wernicke’s aphasia is often thought of as the ‘quintessential’ comprehension disorder in stroke aphasia. It is characterized by impaired auditory comprehension and repetition, paired with fluent speech involving phonological paraphasias and neologisms (Goodglass et al., 2001a). Other linguistic skills such as reading and writing can be relatively preserved (Ellis et al., 1983), suggesting there is a continuum in Wernicke’s aphasia of auditory linguistic and non-auditory abilities which are partially dissociable (Robson et al., 2012). Because of this, Wernicke’s aphasia is commonly considered to be based on an auditory ‘input’ or phonological perception deficit (Harris, 1970; Luria, 1970; Goldblum and Albert, 1972). However, this account is challenged by the observation that other aphasic patients can be equally impaired at phonemic discrimination tasks without showing severe semantic access deficits at the single word level (Blumstein et al., 1977). Moreover, patients with Wernicke’s aphasia can exhibit impairment on non-verbal as well as verbal semantic tasks, although they are notably worse at understanding spoken words (De Renzi et al., 1972; Cohen et al., 1980; Gainotti et al., 1983; Ogar et al., 2011). For example, recent evidence suggests that patients with Wernicke’s aphasia can have additional multimodal semantic control impairments (Robson et al., 2012).
Although auditory–verbal comprehension in Wernicke’s aphasia has been most commonly studied, a long parallel literature has examined stroke patients with ‘multimodal’ semantic deficits (semantic aphasia; Head, 1926; Luria, 1973). Luria (1970) described patients who were unable to integrate relationships between concepts, while Head (1926) characterized semantic aphasia as a deficit in comprehending the full extent of meaning of words and pictures. Both descriptions emphasize a deficit of complex semantic processing across modalities, with difficulty drawing inferences beyond dominant or literal interpretations (see also Hier et al., 1980; Ardila et al., 2000). We have used the term ‘semantic aphasia’ to refer to a multimodal semantic deficit in which there is difficulty in the controlled ‘application’ of knowledge correlating with executive impairment (Jefferies and Lambon Ralph, 2006; Jefferies et al., 2007; Corbett et al., 2009b; Hoffman et al., 2010; Noonan et al., 2010; Jefferies, 2013; Lambon Ralph, 2014). These features share some similarities with the original descriptions of semantic aphasia (for review, see Gainotti, 2014) despite researchers emphasizing different characteristics and using different tasks. The original definitions of semantic aphasia provided by Head (1926) and Luria (1973) referred to a ‘high-level’ deficit in understanding multiple concepts with mild (if any) deficits in single object processing. In this study, and previous publications by the current authors, the majority of semantic aphasia cases had large frontoparietal lesions and comprehension problems at a single-item level, though the patients’ impairments were still more apparent with more complex stimuli. The term ‘semantic aphasia’ transcends classical ‘Boston’ aphasia classifications; however, many patients with semantic aphasia present with the profile of transcortical sensory aphasia, displaying good repetition—at least no worse than would be expected from spontaneous speech production—and speech free from jargon. Cases with semantic aphasia show deregulated verbal and non-verbal semantic behaviour, especially when they are required to use conceptual information in a flexible way, in the absence of few external constraints (Corbett et al., 2009a, b, 2011). Their semantic performance is (i) relatively good on tasks with minimal control demands, such as matching words or objects that are highly associated (e.g. ‘salt’ with ‘pepper’), but poorer for weak associations (e.g. ‘salt’ with ‘sugar’; Noonan et al., 2010); (ii) highly consistent across decisions involving words and pictures, but not across tasks with different executive demands, such as word-picture matching and association judgements (Jefferies and Lambon Ralph, 2006); and (iii) susceptible to being aided by cues and misled by miscues that are designed to activate the target or distracters in both picture naming and object use demonstrations (Jefferies et al., 2007; Soni et al., 2009; Noonan et al., 2010; Corbett et al., 2011).
Current descriptions of Wernicke’s aphasia and semantic aphasia suggest these groups may show somewhat distinct, yet partially overlapping deficits of semantic ‘access’. These problems have been previously explored (and partly defined by) the ‘classical’ paradigm of cyclical word–picture matching (Warrington and McCarthy, 1983; Forde and Humphreys, 1997; Warrington and Crutch, 2004). In semantic ‘access’ patients, including cases with semantic aphasia, accuracy declines when a small set of semantically-related items are repeatedly tested over a number of cycles, with the target on one trial becoming a distractor on the next (Forde and Humphreys, 1997; Warrington and Crutch, 2004; Jefferies et al., 2007; Gardner et al., 2012; Thompson and Jefferies, 2013). The same effect has been shown in non-repeating related items across a session (Warrington and McCarthy, 1983; Forde and Humphreys, 1997), suggesting that competition builds up between semantic associates making it increasingly difficult to select the appropriate target. However, competition in the traditional cyclical task is particularly strong since it requires: (i) inhibition of items after they have been selected; and (ii) reselection when these items subsequently become targets again in later cycles (Thompson-Schill et al., 1997; Gotts and Plaut, 2002; Badre et al., 2005; Jefferies et al., 2007; Mirman et al., 2013).
Even though cyclical word–picture matching tasks have been paradigmatic in establishing the existence of semantic ‘access’ impairment, research using these tasks has typically examined single cases or small clusters of patients selected on the basis that they show declining performance over cycles (Warrington and McCarthy, 1983; Warrington and Cipolotti, 1996; Forde and Humphreys, 1997; Warrington and Crutch, 2004; Crutch and Warrington, 2008; Thompson and Jefferies, 2013). As a consequence, not enough is known about the typical profile of comprehension deficits following stroke, and whether declining semantic ‘access’ with repetition is a common problem—yet this issue has important clinical and theoretical implications. This study tackled these questions by comparing the comprehension of two groups (Wernicke’s aphasia and semantic aphasia), with overlapping yet distinct aphasia and lesion profiles, on classical cyclical spoken word-picture and picture-picture matching for the first time. The patients were not specifically selected to show effects of cycle—instead, this was our key outcome measure. We used a comparative case series approach (Lambon Ralph et al., 2007, 2011) to investigate group-level differences and similarities, as well as the causes of individual variation within the groups, examining the impact of patient group (semantic aphasia versus Wernicke’s aphasia), modality (pictures versus words), lesion location (prefrontal versus temporoparietal lesions) and item repetition (charting improvement or decline in comprehension as concepts are repeated). Below, each of these variables is discussed in more detail.
Patient group
Clinical labels are important in aphasia research and therapy as they capture meaningful constellations of symptoms and make predictions about how individuals will perform in multiple tasks (Henseler et al., 2014); however, these classifications are graded rather than absolute, can be partially overlapping and different types of patient can show similar deficits on specific tasks (Butler et al., 2014). Deficient executive control over semantic processing is a core feature of semantic aphasia but has recently been extended to predict behaviour in Wernicke’s aphasia (Robson et al., 2012). Thus, we might anticipate overlapping deficits in patients with semantic aphasia and Wernicke’s aphasia.
Lesion location
Classifications of aphasic symptoms were devised before neuroimaging methods provided us with detailed information about the functions of specific regions of cortex. These insights suggest that aphasic symptoms should be strongly predicted by lesion location; nonetheless, individual circumstances (e.g. pre-stroke anatomical functioning, age at stroke, amount of therapy post-stroke etc.) affect functional adjustment to brain injury, meaning that lesion location is not deterministic but affects the probability of particular impairments. On average temporoparietal damage is more extrasylvian than perisylvian in semantic aphasia than Wernicke’s aphasia (Chertkow et al., 1997; Berthier, 1999; Dronkers et al., 2004; Robson et al., 2012). Wernicke’s aphasia is particularly associated with damage to the superior temporal gyrus (Eggert, 1977), an area linked to speech perception at a phonological level (Hickok and Poeppel, 2000; Buchsbaum et al., 2001; Okada and Hickok, 2006) and damage here is also thought to account for phonemic paraphasias and poor repetition/naming in Wernicke’s aphasia (Damasio, 1992; Goodglass, 1992). In contrast, the temporal lobe damage in semantic aphasia is focused on posterior middle and inferior temporal gyrus (Berthier, 1999; Noonan et al., 2010; Corbett et al., 2011; Jefferies, 2013), brain areas linked to both word and picture-based semantic tasks.
Individuals in both groups can have left inferior frontal gyrus damage (Gardner et al., 2012; Robson et al., 2012), and this is not predictive of aphasia classification yet could relate to semantic access deficits. Previous work has suggested that patients with damage to left inferior frontal gyrus show declining semantic performance over cycles, whereas lesions restricted to posterior temporal cortex do not elicit this pattern (Jefferies et al., 2007; Campanella et al., 2009; Schnur et al., 2009; Gardner et al., 2012). These findings are broadly consistent with the proposal that left inferior frontal gyrus is crucial for post-retrieval selection (Badre et al., 2005; Badre and Wagner, 2007), particularly when a concept has been activated and inhibited, and then needs to be reselected. Some researchers have explicitly linked left inferior frontal gyrus damage to lexical selection ‘for speech production’ and have suggested that patients do not show the same pattern in comprehension tasks (Schnur et al., 2006, 2009). The current study examines if this relationship with lesion location holds, irrespective of patient classification for semantic matching tasks.
Modality
Much of the previous work on semantic ‘access’ deficits has targeted the auditory–verbal domain (Warrington and McCarthy, 1983, 1987; McNeil et al., 1994; Warrington and Cipolotti, 1996; Jefferies et al., 2007). Recently, however, comparisons of verbal and non-verbal cyclical matching tasks have become a focus of debate: some individual patients appear to have an ‘access’ impairment that is limited to verbal tasks (Warrington and McCarthy, 1983; Warrington and Cipolotti, 1996; Warrington and Crutch, 2004; Crutch and Warrington, 2008), while we and others have proposed that semantic access/control mechanisms are domain-general and occur across modalities (Forde and Humphreys, 1997; Corbett et al., 2009a, 2011; Gardner et al., 2012). It is perhaps unsurprising that cases with semantic aphasia show a decline in comprehension for both words and pictures on cyclical matching tasks, since they have multimodal semantic control deficits (although the patients are not specifically recruited to show deficits in executive-semantic processing or effects of repetition/cycle). In contrast, as cases with Wernicke’s aphasia typically show poorer comprehension of spoken words than pictures, it might be that these patients have marked access deficits in the verbal domain and, as a consequence, resemble the semantic ‘access’ patients studied by Warrington and colleagues (like the single case described by Thompson and Jefferies, 2013).
Repetition
Semantic ‘access’ patients show declining performance when stimuli are repeated in the cyclical matching paradigm even though repetition in other tasks typically enhances performance. When errors arise from a failure to ‘activate’ concepts from a particular sensory input (Dell et al., 1997), repetition should ameliorate this deficit. Wernicke’s aphasics show strong facilitation with repetition priming, i.e. when a target is presented that is identical to a prime (Blumstein et al., 2000). However, patients with Wernicke’s aphasia have also shown interference effects and can be negatively affected by repetition through activation of related distractors—just like patients with semantic aphasia. For example, patients with Wernicke’s aphasia can show impairment on tasks that have phonemically overlapping primes (e.g. ‘piano-pyjamas’) or phonologically related distractors (e.g. hammer with ‘hammock’), suggesting a difficulty in deactivating competing word candidates (Weiner et al., 2004; Janse, 2006; Yee et al., 2008). Indeed, Mirman and colleagues (2013) suggested that impaired input processing (such as that observed in Wernicke’s aphasia) makes residual activation of previously processed items more difficult for the new input to overcome. Therefore, patients with Wernicke’s aphasia may show a mixture of both facilitation from repetition (due to failure to activate concepts initially), followed by interference from overactivation of competing concepts (Campanella and Shallice, 2011). Patients with semantic aphasia, on the other hand, are unlikely to show initial facilitation from repetition, due to intact input processing mechanisms.
In summary, although patients with semantic aphasia and Wernicke’s aphasia are considered to have problems ‘accessing’ conceptual representations, rather than degradation of semantic representations per se (e.g. as observed in semantic dementia: Warrington and Cipolotti, 1996; Jefferies and Lambon Ralph, 2006), these groups have not been directly compared on cyclical matching tasks designed to elicit these deficits. This study establishes (i) whether semantic ‘access’ impairment is a common problem for comprehension-impaired people with stroke aphasia (in cases not specifically selected to show the pattern of declining accuracy over repeated cycles); (ii) whether Wernicke’s aphasia and semantic aphasia cases show qualitatively the same type of semantic access disorder; (iii) to what extent access disorders are limited to the verbal domain (in either group); and (iv) what accounts for variability ‘within’ semantic aphasia and Wernicke’s aphasia, assessing effects of lesion location, modality and repetition.
Materials and methods
Participants
Semantic aphasia
Aphasia profiles and demographic information are displayed in Table 1. Thirteen patients with semantic aphasia took part in this experiment. Data are included from nine patients with semantic aphasia reported by Gardner et al. (2012), along with four additional patients. In line with previous studies on semantic aphasia, patients were selected on the basis of semantic comprehension deficits affecting both words and pictures using the Camel and Cactus task of semantic association (Bozeat et al., 2000). They were not specifically selected to show effects of cycle in matching tasks or deficient semantic control. While some of the patients had relatively selective comprehension deficits (and thus presented with a pattern of ‘transcortical sensory aphasia’), speech fluency and repetition scores were not used as selection criteria and therefore patients within the group had variable language deficits affecting these domains. All patients had chronic impairment after a cerebrovascular accident at least 1 year before testing.
Table 1.
Aphasia profiles and demographic information for patients with semantic aphasia and Wernicke’s aphasia
| Case | Age | Sex | Full-time education (leaving age) | Aphasia classification | BDAE fluency percentile | Repetition |
Cambridge comprehension – spoken | BDAE comprehension percentile | |
|---|---|---|---|---|---|---|---|---|---|
| Non-words (% correct) | Words (% correct) | ||||||||
| HN | 80 | M | 15 | Anomic/TSA | NT | 56 | 86 | 56 | NT |
| SC | 76 | M | 16 | Anomic/TSA | 90 | 87 | 98 | 89 | 37 |
| ME | 36 | F | 16 | TSA | 100 | 93 | 100 | 81 | 33 |
| KS | 59 | M | 16 | TSA | 97 | 73 | 94 | 72 | 43 |
| EW | 74 | F | 15 | TSA | NT | NT | 80 | 91 | NT |
| PG | 59 | M | 18 | TSA | 40 | 73 | 91 | 94 | 20 |
| NY | 63 | M | 15 | Mixed transcortical | 37 | 40 | 81 | 89 | 47 |
| BB | 55 | F | 16 | Mixed transcortical | 17 | 83 | 96 | 76 | 10 |
| DB | 83 | M | 16 | TSA | 90 | 70 | 85 | 73 | 13 |
| GH | 55 | F | 15 | Global | NTa | NTa | 16a | 94 | NT |
| EC | M | 70 | 16 | Global | NTa | 0a | 0a | 63 | NT |
| KA | 74 | M | 14 | Global | 23a | 0a | 0a | 49 | 0 |
| LS | 71 | M | 15 | TSA | 90 | 90 | 96 | 74 | 13 |
| EL | 62 | M | 15 | WA | 96 | 0 | 18 | 45 | 14 |
| MR | 66 | M | 15 | WA | 83 | 4 | 8 | 52 | 20 |
| CW | 71 | M | 15 | WA | 91 | 13 | 49 | 71 | 48 |
| DMC | 68 | M | 18 | WA | 80 | NT | 0 | 25 | 10 |
| DR | 77 | M | 15 | WA | 74 | NT | 1 | 14 | 5 |
| LaS | 67 | M | 15 | WA | 85 | NT | 6 | 50 | 15 |
| DL | 74 | M | 15 | WA | 90 | NT | 1 | 13 | 8 |
| CB | 61 | M | 15 | WA | 38 | NT | 4 | NT | 10 |
aLow fluency with minimal words produced on a cookie theft task (Goodglass and Kaplan, 1983).
BDAE = Boston Diagnostic Aphasia Examination (Goodglass and Kaplan, 1983).
BDAE Comprehension percentile is derived from three subtests (word discrimination, commands, complex ideational material). Cambridge comprehension refers to an average percentage score on spoken word-to-picture matching tasks found in the Cambridge Semantic Battery (Bozeat et al., 2000) and the environmental sounds task (Bozeat et al., 2003). BDAE fluency percentile is derived from phrase length, melodic line and grammatical form ratings. BDAE Repetition percentile is an average of word and sentence repetition subtests. Word/non-word repetition = Tests 8 and 9 from Psycholinguistic Assessments of Language Processing in Aphasia: PALPA (Kay et al., 1992). Aphasia classifications were based on fluency, repetition and comprehension. TSA (transcortical sensory aphasia) was defined as good or intermediate fluency/repetition and poorer comprehension. Wernicke’s aphasia (WA) was defined as relatively fluent speech with poor repetition and comprehension. NT = not tested.
Wernicke’s aphasia
Eight patients with Wernicke’s aphasia were selected after a single left hemisphere stroke to show classic symptoms of Wernicke’s aphasia: a single word comprehension impairment, fluent sentence-like speech punctuated with phonological or neologistic errors, and errors on single word repetition and naming. Participants were screened using the Boston Diagnostic Aphasia Examination (Goodglass et al., 2001b) to show comprehension impairment below the 45th percentile and sentence repetition impairment below the 65th percentile. The average phrase length in everyday speech was required to be above six words. In structured picture description, paraphasias had to occur every few utterances as a minimum. These were largely phonological (e.g. ‘papple’ for apple) or neologistic.
Patient lesion analysis
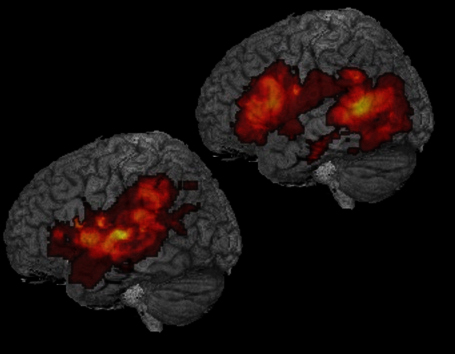
CT/MRI scans were available for all eight patients with Wernicke’s aphasia (one CT scan, seven MRI scans), and 12/13 patients with semantic aphasia (two CT scans, 10 MRI scans). A scan was not available for one semantic aphasia case (Patient PG) due to contraindications for MRI; a report of a CT scan in the acute phase indicated a left frontal lesion. An overlay of lesion maps created from automated lesion identification (Seghier et al., 2008) is displayed in Fig. 1.
Figure 1.

Lesion overlay map from automatic lesion identification. Lightest colours show areas of maximal overlap between subjects. There were eight patients with Wernicke’s aphasia, and 12 patients with semantic aphasia included in this lesion analysis (shown in A and B; a further semantic aphasia patient was not scanned). In the prefrontal group (C), there were four patients with Wernicke’s aphasia and eight patients with semantic aphasia (one semantic aphasia patient with a prefrontal cortex lesion was not scanned). In the temporoparietal group, there were four patients with Wernicke’s aphasia and five with semantic aphasia (D). To produce these images, the patients’ brains were compared to aged-matched control MRI scans, which were collected at the Universities of Manchester and York. Grey matter, white matter and CSF were segmented and changes from the healthy control brains were highlighted as ‘lesion’ (or more precisely, an unexpected tissue class) using automated methods (Seghier et al., 2008). For the Manchester patients there were 19 controls, with a mean age of 68.2 years [standard deviation (SD) = 5.99; eight female, 11 male]. For the York patients, there were 14 controls, mean age of 64.7 years (SD = 6.5, eight female, six male). The automatic lesion identification algorithm was run separately for Manchester and York cases who were examined using a different MRI scanner. CT scans (Patients MR, BB and KA) were hand-drawn onto a template (Damasio and Damasio, 1989), allowing these patients to be included in these images.
Table 2 displays details of the patients’ lesions, focusing on regions of interest in temporal, parietal and frontal cortex. The inferior anterior temporal lobe, implicated in amodal semantic representation and atrophied in semantic dementia, was spared in all cases (Binney et al., 2010; Mion et al., 2010). The main area of lesion overlap in semantic aphasia was in posterior middle temporal and inferior temporal gyrus [Brodmann area (BA) 37 and BA21]. In the Wernicke’s aphasia group, there was greater damage to the posterior superior temporal and supramarginal gyri (BA 22 and BA 40). Nearly all patients with Wernicke’s aphasia had damage to anterior-to-mid superior temporal gyrus, where almost none of the semantic aphasia cases did. Similarly, the posterior occipital-temporal region was nearly always damaged in semantic aphasia patients but not in those with Wernicke’s aphasia. The groups could be entirely separated on the basis of temporal lobe lesion location after the removal of a few cases (Patient EL from the Wernicke’s aphasia group and Patients BB, GH and EC from the semantic aphasia group). Thus, the distribution of damage in the temporal lobe was a key predictor of aphasia classification (Supplementary material and Supplementary Tables 1 and 2).
Table 2.
Details of semantic aphasia and Wernicke’s aphasia patients’ lesions
| Prefrontal |
Anterior temporal |
Posterior temporal |
Parietal |
||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Patient | Lesion group | Lesion size (%)a | DLPFC | orbIFG | trIFG | opIFG | sTP | aSTG | pSTG | pMTG | pITG | FG | POT | AG | SMG | ||
| BA9 | BA46 | BA47 | BA45 | BA44 | BA38 | BA22 | BA22 | BA21 | BA20 | BA36 | BA37 | BA39 | BA40 | ||||
| HN | SA TP-only | 6 | - | - | - | - | - | - | - | - | 2 | 1 | - | 2 | - | - | |
| SC | SA TP-only | 8 | - | - | - | - | - | - | - | - | 2 | 2 | - | 2 | 2 | 1 | |
| ME | SA TP-only | 5 | - | - | - | - | - | - | - | - | 1 | 2 | 2 | 1 | - | - | |
| KS | SA TP-only | 2 | - | - | - | - | - | - | - | 1 | 2 | - | - | 2 | - | - | |
| EW | SA TP-only | 2 | - | - | - | - | - | - | - | - | - | 1 | - | 1 | - | - | |
| NY | SA PF+ | 14 | - | 1 | 2 | 2 | 2 | - | - | 2 | - | - | - | - | 1 | 1 | |
| BB | SA PF+ | 3 | - | - | 2 | 2 | 2 | - | 1 | 1 | - | - | - | - | - | - | |
| DB | SA PF+ | 12 | 1 | 1 | 1 | 2 | 2 | - | - | 2 | 1 | - | - | - | - | 1 | |
| GH | SA PF+ | 12 | - | - | 2 | 1 | 1 | - | 1 | 2 | 1 | - | - | 2 | 1 | 1 | |
| EC | SA PF+ | 17 | - | - | 2 | 1 | 2 | 1 | 1 | 2 | 1 | - | - | 1 | - | - | |
| KA | SA PF+ | 6 | - | - | - | - | 2 | - | - | 2 | 1 | - | - | 1 | - | 1 | |
| LS | SA PF+ | 17 | - | 1 | - | 2 | 2 | - | - | - | 2 | 2 | - | 2 | 2 | 2 | |
| % SA patients with damage to that region | 8 | 25 | 42 | 50 | 58 | 8 | 25 | 58 | 75 | 42 | 8 | 75 | 33 | 50 | |||
| EL | WA TP-only | 5 | - | - | - | - | - | - | - | 1 | - | - | - | - | 1 | 2 | |
| MR | WA TP-only | 3 | - | - | - | - | - | - | 1 | 2 | 1 | - | - | - | - | 2 | |
| CW | WA TP-only | 4 | - | - | - | - | - | - | 1 | 2 | - | - | - | - | - | 2 | |
| DMC | WA TP-only | 16 | - | - | - | - | - | 1 | 1 | 2 | 1 | - | - | - | 2 | 2 | |
| DR | WA PF+ | 8 | 1 | - | - | 2 | 2 | - | 1 | 1 | - | - | - | - | 1 | 1 | |
| LaS | WA PF+ | 12 | - | - | 1 | - | 2 | 1 | 2 | 2 | 2 | - | - | 1 | 1 | 2 | |
| DL | WA PF+ | 8 | - | - | - | 2 | 1 | 2 | 2 | 1 | 2 | - | - | - | - | 2 | |
| CB | WA PF+ | 17 | - | - | 1 | 2 | 2 | 1 | 2 | 2 | - | - | - | - | - | 2 | |
| % WA patients with damage to that region | 13 | 0 | 25 | 38 | 50 | 50 | 88 | 100 | 50 | 0 | 0 | 13 | 50 | 100 | |||
Quantification of lesion: 2 = complete destruction/serious damage to cortical grey matter; 1 = partial destruction/mild damage to cortical grey matter. Anatomical abbreviations: DLPFC = dorsolateral prefrontal cortex; orbIFG = pars orbitalis in inferior frontal gyrus; trIFG = pars triangularis in inferior frontal gyrus; opIFG = pars opercularis in inferior frontal gyrus; sTP = superior temporal pole; STG = superior temporal gyrus; MTG = middle temporal gyrus; ITG = inferior temporal gyrus; FG = fusiform gyrus; POT = posterior occipitotemporal area; SMG = supramarginal gyrus; AG = angular gyrus; SA = semantic aphasia; WA = Wernicke’s aphasia; PF+ = prefrontal areas; TP-only = temporoparietal only.
aLesion size was estimated by overlaying a standardized grid of squares onto each patient’s template to determine the percentage of squares damaged relative to the complete undamaged template. Anterior superior temporal gyrus (aSTG) was obtained by assessing BA 22 on the fourth and fifth slice of the Damasio template: any damage in front of the midpoint was defined as lesioned anterior superior temporal gyrus. Posterior superior temporal gyrus (pSTG) was restricted to the back half of the superior temporal gyrus, using the fifth and sixth slide of the Damasio template. A scan for Patient PG was unavailable; a radiographer’s report identified frontal and capsular damage.
The two patient groups were further subdivided into those with damage restricted to temporoparietal regions (the ‘TP-only’ group) and those with lesions encroaching into prefrontal areas (the ‘PF+’ group), including left inferior frontal gyrus (BA 44 and BA 45). Four cases with Wernicke’s aphasia had damage restricted to posterior temporal regions and four had damage extending anteriorly to frontal regions. There were eight semantic aphasia cases with damage extending to prefrontal regions and five with damage restricted to posterior temporal regions (Table 2).
Neuropsychological assessment
Background neuropsychology
General neuropsychological testing included digit span (Wechsler, 1987), Ravens Coloured Progressive Matrices test of non-verbal reasoning (Raven, 1962), the Visual Object and Space Perception battery (VOSP, Warrington and James, 1991), Elevator Counting with and without distraction from the Test of Everyday Attention (Robertson et al., 1994), and the Brixton Spatial Rule Attainment task (Burgess and Shallice, 1997). The patients were also examined on a standard battery of semantic tests to assess their comprehension of pictures, environmental sounds and words (presented simultaneously in spoken and written forms unless otherwise stated). These included: (i) Pyramids and Palm Trees (Howard and Patterson, 1992), a two alternative-forced-choice test of semantic associations for pictures and words; (ii) three components of the Cambridge 64-item semantic test battery (Bozeat et al., 2000): spoken word–picture matching using 10 semantically-related response options, plus picture and word versions of the Camel and Cactus Test (a four alternative-forced-choice test tapping semantic associations); (iii) a three alternative-forced-choice test 96-item synonym judgement task (Jefferies et al., 2009); and (iv) an environmental sounds battery, which involved matching environmental sounds-to-pictures, spoken words-to-pictures, written words-to-pictures and sounds-to-written words (Bozeat et al., 2000). There were 10 semantically-related response options.
Cyclical matching tasks
These picture–picture matching and word–picture matching tasks were used previously by Gardner et al. (2012, Experiment 1) and Warrington and Crutch (2004). Participants selected one of four pictures matching a spoken word or picture probe (Fig. 2). Items were presented repeatedly such that the target on one trial became the distractor on another, until all items within a semantic category had been tested. This completed one cycle. There were four cycles for each set of items, which probed the items in the set in a pseudorandom order, followed by a short break. This removed the potential confound between time and cycle, as cycle four of set one was presented before cycle one of set two. Participants had 10 s to point to the target, and immediately after each response the researcher triggered the next trial. There were four practice items before the start of each block. The same probes were presented as pictures and words. Testing was carried out in four blocks using an ABBA design to control for order effects across modalities. The stimuli consisted of 40 inanimate objects. These were grouped into 10 semantic sets (tools, electrical items, drink containers, clothes, household appliances, kitchen tools × 2, furniture × 2 and vehicles). The experiment was run using E-prime 1.1
Figure 2.

Examples of trials used in the cyclical item matching task. PPM = Picture-Picture matching; WPM = Word-Picture matching.
Results
Background neuropsychology
Non-semantic tasks
Results are provided in Table 3. There was a significant difference between groups in digit span [t(17) = 2.538, P = 0.021]. Patients with Wernicke’s aphasia were at floor on this task, whereas only 7/13 semantic aphasia cases were below the normal cut-off. There were no differences between patients with semantic aphasia and Wernicke’s aphasia on the Ravens Coloured Progressive Matrices test of non-verbal reasoning (t < 1), Brixton (t < 1), the Elevator Counting task with or without distraction (t < 1), or any subscale of the Visual Object and Space Perception (VOSP) battery [t(13) ≤ 1.447, P ≥ 0.172]. Patients with semantic aphasia and and those with Wernicke’s aphasia showed evidence of impaired executive control. Some individuals in both groups were also impaired on visual tasks. Dot counting from VOSP battery is influenced by the ability to produce number words, whereas cube analysis draws on executive skills, making these scores hard to interpret. Moreover, spatial deficits are unlikely to explain the effects of cycle on our experimental tasks. Although visual search is relevant for global performance in this paradigm, visual impairment should have a relatively stable impact across trials. In addition, many of the patients with semantic aphasia reported here were previously shown to (i) have unchanging performance across cycles for blocks of semantically unrelated items in the same paradigm; and (ii) declining picture naming performance across cycles—even though visual search requirements are substantially reduced in this task (Jefferies et al., 2007).
Table 3.
Background performance: non-semantic tasks
| Patient | Group | Digit span | RCPM | VOSP |
TEA |
Brixton | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Dot counting | Position discrimination | Number location | Cube analysis | No distraction | Distraction | |||||
| Max | 36 | 10 | 20 | 10 | 10 | 7 | 10 | 54 | ||
| Control mean (SD) | 7 (0)a | 32.6 (2.3)a | 9.9 (0.3) | 19.6 (0.9) | 9.4 (1.1) | 9.2 (1.2) | 6.6 (1.2) | 8.2 (2.8) | 30 (4.8)a | |
| Normal cut-off | 5 | 28b | 9.5 | 17.8 | 7.2 | 6.8 | 4.2 | 2.6 | 28 | |
| HN | SA – TP-only | 6 | 20* | 8* | 19 | 9 | 4* | 7 | 9 | 28 |
| SC | SA – TP-only | 6 | 22* | 10 | 17* | 10 | 9 | 7 | 1* | 25* |
| ME | SA – TP-only | 6 | 13* | 3* | 15* | 2* | 4* | 7 | 9 | 11* |
| KS | SA – TP-only | 8 | 31 | NT | NT | NT | NT | 5 | 9 | 28 |
| EW | SA – TP-only | 4* | 30 | 10 | 20 | 10 | 7 | NT | NT | 33 |
| PG | SA – PF+ | 6 | 23* | 5* | 20 | 9 | 10 | 3* | 0* | 26* |
| NY | SA – PF+ | 3* | 26* | 10 | 20 | 10 | 5* | 3* | 2* | 34 |
| BB | SA – PF+ | 5 | 24* | 10 | 18 | 8 | 2* | 0* | 4 | 23* |
| DB | SA – PF+ | 4* | 31 | 6* | 0* | 10 | 3* | 2* | 2* | 31 |
| GH | SA – PF+ | 2* | 32 | 10 | 4* | 0* | 0* | 6 | 1* | 18* |
| EC | SA – PF+ | 0* | 12* | 3* | 14* | 10 | 6* | 1* | 1* | 24* |
| KA | SA – PF+ | 0* | 12* | 0* | 14* | 6* | 0* | 5 | 5 | 6* |
| LS | SA – PF+ | 4* | 6* | 6* | 16* | 8 | 4* | 2* | 3 | 14* |
| EL | WA – TP-only | 2* | 27* | 7* | 20 | 10 | 6* | 0* | 0* | 25* |
| MR | WA – TP-only | 2* | 31 | 9* | 19 | 5* | 6* | 7 | 2* | 16* |
| CW | WA – TP-only | 4* | 29 | 10 | 19 | 6* | 10 | 7 | 7 | 39 |
| DMC | WA – TP-only | 1* | 23* | NT | NT | NT | NT | NT | NT | NT |
| DR | WA – PF+ | 1* | 10* | NT | NT | NT | NT | NT | NT | NT |
| LaS | WA – PF+ | 1* | 21* | NT | NT | NT | NT | NT | NT | NT |
| DL | WA – PF+ | NT | 22* | NT | NT | NT | NT | NT | NT | NT |
| CB | WA – PF+ | 2* | 25* | NT | NT | NT | NT | NT | NT | NT |
*Denotes impaired performance. Control performance and normal cut-offs taken from the following published texts except where stated.
aNorms from 15 healthy controls tested at the University of York, average age 68, four male.
b2 SD below mean of controls tested at the University of York.
RCPM = Raven’s Coloured Progressive Matrices (Raven, 1962); VOSP = Visual Object and Space Perception battery (Warrington and James, 1991) section 5–8; TEA = Test of Everyday Attention (Robertson et al., 1994); BSRA = Brixton Spatial Rule Attainment Task (Burgess and Shallice, 1997); NT = not tested; WA = Wernicke’s aphasia; SA = semantic aphasia; TP = temporoparietal; PF = prefrontal.
Semantic tasks
Results are displayed in Table 4. All patients showed some degree of impairment in both modalities (written word or picture), except one patient with Wernicke’s aphasia who did not show a deficit for pictures (Patient CW).
Table 4.
Background performance: semantic tasks
| Patient | Group | Spoken WPM | Naming | CCTp | CCTw | PPTp | PPTw | Synonyms | Environmental sounds test |
|||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Written word- picture | Spoken word- picture | Sound- picture | Sound- written word | |||||||||
| Max | 64 | 64 | 64 | 64 | 52 | 52 | 96 | 48 | 48 | 48 | 48 | |
| Control mean (s.d.) | 63.7 (0.5) | 62.3 (1.6) | 58.9 (3.1) | 60.7 (2.06) | 51.2 (1.4) | 51.1 (1.1) | 94.4 (1.2) | NTa | 47.8 (0.6) | 41.2 (2.5) | 40.8 (3.8) | |
| Normal cut off | 63 | 59 | 53 | 57 | 49 | 49 | 92 | NT | 46.6 | 36.2 | 33.2 | |
| HN | SA – TP-only | 50*** | 51*** | 54 | 54* | 35*** | 44*** | 89** | 42*** | 16*** | 36 | NT |
| SC | SA – TP-only | 59*** | 28*** | 47** | 56 | 29*** | 39*** | 71*** | 48 | 41*** | 32** | 32 |
| ME | SA – TP-only | 50*** | 5*** | 13*** | 33*** | 29*** | 39*** | 80*** | 40*** | 40*** | 33** | 35 |
| KS | SA – TP-only | 46*** | 21*** | 44*** | NT | NT | NT | 81*** | NT | NT | NT | NT |
| EW | SA – TP-only | 57*** | 45*** | 45** | 48*** | 50 | 52 | 86*** | 38*** | 45* | 22*** | NT |
| PG | SA – PF+ | 58*** | 46*** | 44*** | 40*** | 42*** | 43*** | 69*** | 44*** | 47 | 33** | 25** |
| NY | SA – PF+ | 60*** | 55** | 36*** | 39*** | 47* | 42*** | 69*** | 47 | 40*** | 28*** | 34 |
| BB | SA – PF+ | 53*** | 10*** | 38*** | 30*** | 41*** | 35*** | 63*** | 26*** | 33*** | 26*** | 27** |
| DB | SA – PF+ | 46*** | 39*** | 51* | 46*** | NT | NT | 54*** | 38*** | 36*** | 21*** | NT |
| GH | SA – PF+ | 60*** | 19*** | 45** | 29*** | NT | NT | 71*** | NT | NT | NT | NT |
| EC | SA – PF+ | 40*** | 1*** | 32*** | 20*** | NT | NT | 41*** | NT | NT | NT | NT |
| KA | SA – PF+ | 35*** | 0*** | 46** | 36*** | 44*** | 44*** | 60*** | 36*** | 21*** | 22*** | 14*** |
| LS | SA – PF+ | 48*** | 5*** | 15*** | 16*** | 31*** | 39*** | 47*** | 33*** | 35*** | 27*** | 17*** |
| EL | WA – TP-only | 30*** | 24*** | 49* | 36*** | 48 | 36*** | 62*** | 45** | 21*** | 30** | 24** |
| MR | WA – TP-only | 32*** | 11*** | 45** | 46*** | 50 | 39*** | 66*** | 40*** | 26*** | 20*** | 17*** |
| CW | WA – TP-only | 51*** | 41*** | 55 | 55* | 51 | 52 | 89** | 47 | 30*** | 21*** | 22*** |
| DMC | WA – TP-only | 16*** | 0*** | NT | NT | 42*** | 39*** | NT | NT | NT | NT | NT |
| DR | WA – PF+ | 9*** | 3*** | NT | NT | 47* | 33*** | NT | NT | NT | NT | NT |
| LaS | WA – PF+ | 32*** | 1*** | NT | NT | 46** | 34*** | NT | NT | NT | NT | NT |
| DL | WA – PF+ | 8*** | 2*** | NT | NT | 46** | 32*** | NT | NT | NT | NT | NT |
| CB | WA – PF+ | 30*** | 0*** | NT | NT | 42*** | 43*** | NT | NT | NT | NT | NT |
*Denotes impaired performance. * ≤ 0.05, ** ≤ 0.01, *** ≤ 0.001 using a modified t-statistic to examine whether an individual is significantly below a control group, taking into account control group size, mean and standard deviation (Crawford et al., 2010). Control performance and normal cut-offs taken from the following published texts except where stated.
aNorms for analysis taken from spoken word-picture matching using the same stimuli. Spoken Word-Picture Matching (WPM) from the Cambridge Semantic Battery (Bozeat et al., 2000); Synonym judgment (Jefferies et al., 2009); Environmental Sounds Test (Bozeat et al., 2003); CCT = Camel and Cactus task in picture and written word forms (Bozeat et al., 2000); PPT = Pyramids and Palm Trees task in picture and written word forms (Howard and Patterson, 1992); NT = not tested; WA = Wernicke’s aphasia; SA = semantic aphasia; TP = temporoparietal; PF = prefrontal.
On the Pyramids and Palm Trees test, repeated-measures ANOVA revealed no main effect of group [F(1,14) = 2.858, P = 0.113] or modality [F(1,15) = 2.246, P = 0.155] but an interaction between group and modality [F(1,15) = 13.247, P = 0.002]. Wernicke’s aphasia patients were less impaired than those with semantic aphasia on picture trials, but the two groups showed similar performance for words.
The 64 item word-picture matching task showed a significant group difference between semantic aphasia and Wernicke’s aphasia [t(18) = 4.895, P < 0.001] with higher performance in patients with semantic aphasia. This identity matching task arguably has fewer semantic control demands than association matching tasks like Pyramids and Palm Trees, as there is no requirement to apply conceptual knowledge in a flexible fashion to determine the relevant relationship in each trial. However, word–picture matching is highly dependent on the ability to decode the spoken probe word and access its meaning, which is a core deficit in Wernicke’s aphasia.
The other semantic tasks were examined in fewer patients (10–16 out of 21 in total). The Camel and Cactus test showed no effect of group [F(1,11) = 1.202, P = 0.296] or interaction of modality and group (F < 1). There was no significant group effect in any condition of the environmental sounds task [t(11) ≤ 1.607, P ≥ 0.136] or in the synonym test (t < 1). The synonym judgement task additionally allowed us to assess the effect of frequency on comprehension (Supplementary material), as the absence of frequency effects is another hallmark of semantic access impairment (Warrington and Shallice, 1979; Warrington and Cipolotti, 1996; Hoffman et al., 2011a; Almaghyuli et al., 2012). Of three patients with Wernicke’s aphasia tested, none showed a significant frequency effect, whereas in the semantic aphasia group, just 2 of 13 patients showed a difference (Supplementary Table 3). Furthermore, in one of these cases, performance was significantly greater for low frequency items, potentially reflecting the greater control demands for high frequency words which have greater contextual diversity (Hoffman et al., 2011a, b). In contrast, other patients with ‘storage’ rather than ‘access’ semantic deficits show robust comprehension benefits for high frequency items on the same task (Jefferies et al., 2009).
Cyclical matching tasks
Healthy controls show ceiling level performance on the cyclical matching tasks and, if anything, a slight speeding up of reaction time with repetition (Gardner et al., 2012). Therefore, the analyses focused only on the patient data.
Omnibus ANOVA
Patients with semantic aphasia and Wernicke’s aphasia showed different effects of cycle and modality (Fig. 3A). A 2 × 4 × 2 repeated-measures ANOVA of modality (picture–picture matching or word–picture matching), cycle, and aphasia group (semantic aphasia or Wernicke’s aphasia) revealed a significant main effect of cycle [F(3,17) = 4.355, P = 0.019], which interacted with aphasia group [F(3,17) = 7.250, P = 0.002] (see below). There was also a marginally significant main effect of aphasia group [F(1,19) = 4.080, P = 0.058] a main effect of modality reflecting poorer performance for words [F(1,19) = 6.858, P = 0.017] and an interaction of aphasia group and modality [F(1,19) = 6.298, P = 0.021] (see below). There were no other significant interactions. As noted in the Supplementary material, these effects of aphasia classification are likely to reflect the distribution of temporal lobe damage in the two groups.
Figure 3.

Accuracy (%) across cycles. (A) Data from semantic aphasia (SA) and Wernicke’s aphasia (WA) patients for word-picture matching (WPM) and picture-picture matching (PPM). (B) Prefrontal (PF+) and temporoparietal (TP-only) patients, including cases from both Wernicke’s aphasia and semantic aphasia groups and combining word and picture modalities. Error bars show standard error of mean.
Cycle
Patients with Wernicke’s aphasia and semantic aphasia showed the same decline in accuracy from cycle 2 to 4, but patients with Wernicke’s aphasia showed initial improvement between cycles 1 and 2, whereas semantic aphasia cases showed a decline. Independent samples t-tests (averaging across both modalities) revealed a significant aphasia group difference at cycle 1 [Bonferroni t(19) = 3.802, P = 0.004] but no differences at other cycles [t(19) ≤ 1.360, P ≥ 0.1]. Indeed, when rerunning the ANOVA without cycle 1, there was no longer an interaction of cycle and aphasia group (F < 1) or a main effect of aphasia group [F(1,19) = 2.173, P = 0.157] although there was an effect of cycle [F(2,18) = 5.696, P = 0.012].
Modality
The interaction of modality and aphasia group reflected equivalent performance for the two groups on the picture–picture matching task, whereas the Wernicke’s aphasia cases showed poorer accuracy on word–picture matching. Independent samples t-tests (averaged across all cycles within each modality) found no significant difference between patients with semantic aphasia and Wernicke’s aphasia on picture–picture matching (t < 1). However, there was a significant difference between aphasia groups for word–picture matching [Bonferroni t(19) = 2.914, P = 0.02]. The ANOVA revealed no interaction of cycle and modality, and no three-way interaction between cycle, aphasia group and modality (F < 1), indicating that patients with Wernicke’s aphasia were consistently worse at the word–picture matching task than patients with semantic aphasia across cycles.
Left inferior frontal gyrus damage
The effect of left inferior frontal gyrus damage is shown in Fig. 3B. Repeated-measures ANOVA examined the following factors: left inferior frontal gyrus damage (TP-only or PF+ groups), modality (word–picture matching versus picture–picture matching), aphasia group (semantic aphasia, Wernicke’s aphasia) and cycle (1–4). Only main effects and interactions reflecting the presence or absence of left inferior frontal gyrus damage are reported here, since the other factors were considered above. There was a near-significant main effect of left inferior frontal gyrus damage [F(1,17) = 3.975, P = 0.062] suggesting that PF+ patients may have been slightly more impaired than TP-only patients overall. PF+ patients had a larger lesion volume [t(18) = 2.822, P = 0.011], but this did not appear to explain the differential effects of cycle in PF+ and TP-only groups. There was no correlation between lesion size and the maximal change between cycles (expressed as a single variable in each case) for either picture–picture matching: r = 0.270, P = 0.250, or word–picture matching: r = 0.304, P = 0.193. This suggests that lesion location, rather than size, predicts semantic access deficits. There was an interaction of cycle and left inferior frontal gyrus damage [F(3,15) = 4.930, P = 0.014]; PF+ patients showed a greater drop in accuracy across cycles than those with damage restricted to TP-only regions. Between-subjects t-tests comparing (i) cycles 1 to 4; and (ii) cycles 2 to 4 were computed for PF+ and TP-only patient groups (averaged across modalities). PF+ patients showed a decline in accuracy between cycles 1 and 4 [Bonferroni t(11) = 3.621, P = 0.008] and cycles 2 and 4 [Bonferroni t(11) = 3.911, P = 0.004]. TP-only patients did not show a difference between cycles 1 and 4 [t(8) = 1.484, P > 0.1] or cycles 2 and 4 [t(8) = 1.178, P > 0.2]. There were no other interactions involving the status of left inferior frontal gyrus, suggesting that both groups and modalities contributed to these effects of lesion location (although this analysis lacks statistical power to detect a subtle interaction with aphasia classification).
Analysis at the level of individual patients largely converged with these findings (Supplementary material). Patients with Wernicke’s aphasia showed consistent modality effects (pictures > words), whereas semantic aphasia cases did not. Significant effects of cycle were found in five semantic aphasia PF+ patients and effects approached significance in a further two cases (i.e. declining accuracy was seen in 7/8 individuals). Effects of cycle were not found in any of the TP-only patients from either group. This analysis also failed to detect effect of cycle in individual PF+ Wernicke’s aphasia cases (Supplementary Table 4).
Consistency across cycles and modality
A characteristic symptom of ‘access’ impairment is inconsistent retrieval of the same items when probed repeatedly (Warrington and McCarthy, 1983; Warrington and Crutch, 2004). Jefferies and Lambon Ralph (2006) found inconsistency in semantic aphasia cases across tasks with differing executive demands, and consistent performance when demands were broadly equivalent, such as when the items were presented in two modalities within the same task. We used logistic regression to assess accuracy consistency across cycles (which might have differing control demands) and between modalities (which should have similar control demands). To achieve this, we used performance on earlier cycles (cycles 1, 2 and 3) to predict scores on later cycles (cycles 2, 3 and 4, respectively), while including the additional variables of modality, lesion location, aphasia group, word frequency and individual patient ID in the model. We also used performance on picture–picture matching to predict accuracy on the word–picture matching task, including cycle and the other variables in the model.
Modality
Both Wernicke’s aphasia and semantic aphasia cases showed significant consistency across modalities. Word–picture matching accuracy was significantly predicted by picture–picture matching accuracy (W = 12.229, P < 0.001). There were also effects of lesion (W = 9.621, P = 0.002) aphasia group (W = 25.658, P < 0.001) and patient ID (W = 281.375, P < 0.001). There were no other significant factors, and no significant interactions. The reverse contrast (predicting picture–picture matching from word–picture matching) yielded similar results, but the main effect of aphasia group became non-significant.
Cycle
Item-by-item accuracy on cycles 2, 3, 4 was significantly predicted by performance on the same items in the previous cycle (1, 2 and 3, respectively) (W = 115.320, P < 0.001). There were also significant effects of modality (W = 25.684, P < 0.001) and patient ID (W = 290.510, P < 0.001). There were interactions between accuracy on the previous cycle and aphasia group (Wernicke’s aphasia versus semantic aphasia) (W = 45.628, P < 0.001) plus accuracy on the previous cycle and lesion location (PF+ versus TP-only) (W = 4.437, P = 0.035). The three-way interaction term (cycle, lesion location and aphasia group) was not significant when this was added to the model (W = 2.161, P = 0.142). By examining the same analyses for subgroups of patients in each modality separately, we were able to interpret these interactions as follows: semantic aphasia PF+ patients showed no consistency across sets of trials hypothesized to have varying control demands (i.e. between earlier and later cycles), whereas semantic aphasia TP-only patients showed some evidence of consistency across cycles on the visual task (not the verbal task). Wernicke’s aphasia cases showed significant levels of consistency, and this was again most evident on the visual task, with the Wernicke’s aphasia TP-only group showing the most consistent performance on this task. This is displayed in Supplementary Table 5.
Error analysis
A full error analysis is described in the Supplementary material. First, we analysed the proportion of errors that were perseverations and omissions across cycles (Supplementary material). Perseverations went up across cycles in both groups, but omissions were greatest at cycle 1 for the patients with Wernicke’s aphasia (and not for the semantic aphasia PF+ group). This might have reflected the initial difficulties that patients with Wernicke’s aphasia had in accessing semantics from inputs: this deficit might be ameliorated by repetition (Supplementary Table 6). We also examined whether errors for a particular target were consistent semantic confusions (i.e. the same incorrect response option was chosen across trials; Supplementary material). A deficit of semantic control was predicted to create consistent errors, since items which are most similar compete for selection (e.g. knife and fork). This prediction was confirmed by our results, displayed in Supplementary Table 7.
Discussion
Although semantic aphasia and Wernicke’s aphasia have a long-standing history of study (Head, 1926; Eggert, 1977), they have rarely been directly compared. Both disorders are considered to reflect an ‘access’ disorder rather than an impairment/degradation of semantic representations per se (Warrington and Cipolotti, 1996; Jefferies and Lambon Ralph, 2006). Yet the term ‘access disorder’ can refer to two different deficits, either impaired entry into semantics from a particular modality (e.g. spoken words), or deficient control over the retrieval of semantic information. This study was able to answer key clinical and theoretical questions with regard to semantic ‘access’ disorders, including: (i) which type of ‘access’ disorder is present in patients with Wernicke’s aphasia versus patients with semantic aphasia; (ii) whether ‘access’ disorders are limited to the verbal domain; and (iii) how the distribution of temporal and frontal lobe damage relates to access impairments. This was achieved using verbal and non-verbal versions of the cyclical matching task. Whilst this assessment has become paradigmatic of ‘access’ semantic disorders, it is typically investigated in cases specifically selected to show declining comprehension with cycle. In contrast, we examined performance on this task systematically and comparatively in case series of Wernicke’s aphasia and semantic aphasia patients.
There were clear differences in the nature of the comprehension impairment in semantic aphasia and Wernicke’s aphasia, which reflected the distribution of temporal lobe damage in these groups. Patients with Wernicke’s aphasia showed poorer comprehension of spoken words than pictures, and input processing deficits characterized by omission errors that were initially ameliorated when stimuli were repeated, following greater damage to anterior-to-mid superior temporal gyrus. In contrast, patients with semantic aphasia showed equivalent impairment of verbal and non-verbal tasks and no beneficial effects of repetition, coupled with greater damage to occipital-temporal cortex. Nevertheless, the Wernicke’s aphasia and semantic aphasia groups showed a similar decline in verbal and non-verbal matching performance as competition increased in later cycles, and in both groups, this decline in performance with repetition was associated with damage to left inferior frontal gyrus. This poorer comprehension on later cycles reflected difficulty inhibiting previous targets, resulting in more perseverations of the preceding response. Our findings confirm a dual-deficit in Wernicke’s aphasia, i.e. deficient auditory/phonological input-processing plus impaired control over conceptual activation affecting both word and picture tasks (Robson et al., 2012). This is compatible with the idea that key parts of the semantic network (both for representation and control) are multimodal (Jefferies and Lambon Ralph, 2006; Binder et al., 2009; Lambon Ralph, 2014).
The semantic aphasia and Wernicke’s aphasia cases also showed other classic features of access impairment (Warrington and Shallice, 1979), including (i) weak or absent frequency effects in a synonym judgement task; and (ii) inconsistent performance when the same items were probed repeatedly. The first of these findings is thought to reflect the higher ‘contextual diversity’ of frequent words: high frequency items occur in a wider range of contexts and are therefore more easily associated with the distracter words in a synonym judgement task, increasing control demands (Hoffman et al., 2011a; Almaghyuli et al., 2012). The consistency analyses also appeared to reflect fluctuations in control demands. Both groups showed consistent performance across modalities, as the executive demands of word and picture trials were broadly matched (Jefferies and Lambon Ralph, 2006). Patients with semantic aphasia showed more inconsistency than Wernicke’s aphasia cases across cycles, consistent with their executive-semantic deficits in the absence of input processing deficits.
The largest difference between Wernicke’s aphasia and semantic aphasia came with the first repetition of the items: Wernicke’s aphasia showed an increase in performance, where patients with semantic aphasia showed no such facilitation. Moreover, patients with Wernicke’s aphasia frequently ask for repetition of spoken input in everyday settings and formal assessments. Presumably, repetition of the stimulus boosts initial poor encoding and patients utilize this powerful effect across many different situations. Similarly, patients with Wernicke’s aphasia show repetition priming effects, absent in some other types of aphasia (Blumstein et al., 2000). These effects have been linked to deficits in activating lexical representations and in auditory working memory (Janse, 2008). For the first time, however, our findings indicate that this repetition priming effect goes beyond the verbal domain: patients with Wernicke’s aphasia found it difficult to activate the meaning of an item from a single presentation irrespective of modality. In contrast, patients with semantic aphasia performed at their best on the first cycle: they were able to maximize access to semantic information with the initial input. Cortical regions showing reduced activity with repetition (e.g. a semantic priming or adaptation effect)—including anterior parts of left inferior frontal gyrus and posterior middle temporal gyrus (Wagner et al., 1997; Gold et al., 2006; Badre and Wagner, 2007)—are more often intact in patients with Wernicke’s aphasia than in those with semantic aphasia. This may explain why patients with Wernicke’s aphasia receive benefit from repetition when semantic aphasia patients do not.
Over and above aphasia type, damage to the left inferior frontal gyrus was shown to predict negative effects of cycle in word and picture matching tasks. A similar conclusion was drawn in parallel studies of patients with prefrontal versus posterior temporal glioma (Campanella et al., 2009, 2012). Patients with left prefrontal cortex lesions have difficulty overcoming competition from previously-relevant responses and adapting their semantic processing when the target changes (Corbett et al., 2008; Jefferies et al., 2008). This function is increasingly required in cyclical semantic matching tasks, as the distracter on one trial becomes the target on the next and thus the target must be identified from a field of previously selected and highly active items (Jefferies et al., 2007). Once initial difficulties in activating concepts from inputs have been overcome, the semantic access impairments that occur in both semantic aphasia and Wernicke’s aphasia are best explained in terms of damage to this modulatory system in prefrontal cortex, as opposed to abnormalities within the representational system per se.
Previous studies have suggested that damage to left inferior frontal gyrus specifically disrupts lexical selection during speech production (Damian et al., 2001; Maess et al., 2002; Belke et al., 2005; Moss et al., 2005; Schnur et al., 2006, 2009; Hsiao et al., 2009; Robinson et al., 2010). This study provides strong evidence that this competition-related deficit is not specific to speech production or even to verbal semantic processing: instead, damage to the left inferior frontal gyrus produces parallel problems in word and picture comprehension tasks, indicating that the control system supports the retrieval and selection of amodal concepts as opposed to word forms alone (Gardner et al., 2012). There is already evidence that the left inferior frontal gyrus responds to non-verbal as well as verbal semantic tasks (Wagner et al., 1997; Chee et al., 2000; Bright et al., 2004; Visser et al., 2012; Krieger-Redwood et al., 2015) and this region is engaged by a multitude of different semantic tasks with high executive demands (Noonan et al., 2013). In particular, the mid-to-posterior part of the left inferior frontal gyrus (damaged in both semantic aphasia and Wernicke’s aphasia patients) is crucial for overcoming ‘post-retrieval selection’, i.e. interference from activated representations or responses that are no longer relevant (Thompson-Schill et al., 1997; Badre et al., 2005).
Beyond the left inferior frontal gyrus, semantic control is associated with a distributed network including left posterior middle/inferior temporal cortex, intraparietal sulcus, pre-supplementary motor area and right inferior frontal gyrus. Some of these regions support domain-general executive control, such as the inferior frontal sulcus and intraparietal sulcus (Duncan, 2010; Woolgar et al., 2011). Others appear to be more specifically semantic in their function, notably left anterior inferior frontal gyrus and posterior middle temporal gyrus (Noonan et al., 2013). Both left inferior frontal gyrus and posterior middle temporal gyrus show stronger activation in functional MRI studies when semantic control demands are maximal (Thompson-Schill et al., 1997; Badre et al., 2005; Whitney et al., 2011a; Noonan et al., 2013), and transcranial magnetic stimulation to these regions elicits equivalent disruption of high-control tasks (Whitney et al., 2011b). Moreover, patients with semantic aphasia with left prefrontal and temporoparietal-only lesions show equivalent effects of various semantic control manipulations, including ambiguity, probe-target connectedness and distractor strength (Noonan et al., 2010). This distinction between cyclical matching tasks (which specifically implicate left inferior frontal gyrus in control; Schnur et al., 2006, 2009; Campanella et al., 2009; Gardner et al., 2012) and other situations suggests that although left inferior frontal gyrus and posterior temporal areas jointly support semantic control, they may make different contributions: the left inferior frontal gyrus might be important when the goals for semantic retrieval change and previous responses are no longer relevant, whereas both structures might work together to determine the correct response when relatively automatic stimulus-driven semantic retrieval is insufficient to support understanding.
In summary, the unique contribution of this study is to show that ‘refractory’ effects (i.e. negative effects of cycle) are comparable for words and pictures, not only in patients with semantic aphasia (selected to have multimodal semantic impairment) but also in patients with Wernicke’s aphasia (selected to show poor single-word comprehension and speech punctuated with phonological or neologistic errors). These effects are linked to damage to left prefrontal cortex (left inferior frontal gyrus), which has been previously associated with the control of competition from previously relevant responses. We conclude that patients with Wernicke’s aphasia have two types of semantic access impairment—both difficulty with initial conceptual activation (ameliorated by repetition) and difficulty in the face of strong competition (increased by repetition), whereas patients with semantic aphasia show the second type of semantic access deficit in isolation.
Supplementary Material
Acknowledgements
We thank the patients and their carers for their generous assistance with this study, and Sebastian Crutch and Elizabeth Warrington for the use of their materials.
Funding
This work was partially supported by a grant from Age UK (ref. 335). H.E.T. received additional support from the Wellcome Trust [ref: 105624] through the Centre for Chronic Diseases and Disorders (C2D2) at the University of York. E.J. was supported by a grant from the European Research Council (283530-SEMBIND); H.R. by a Stroke Association Allied Health Bursary (TSAB2008/01) and M.A.L by an MRC programme grant (MR/J004146/1).
Supplementary material
Supplementary material is available at Brain online.
References
- Almaghyuli A, Thompson HE, Lambon Ralph MA, Jefferies E. Deficits of semantic control produce absent or reverse frequency effects in comprehension: evidence from neuropsychology and dual task methodology. Neuropsychologia 2012; 50: 1968–79. [DOI] [PubMed] [Google Scholar]
- Ardila A, Concha M, Rosselli M. Angular gyrus syndrome revisited: acalculia, finger agnosia, right-left disorientation and semantic aphasia. Aphasiology 2000; 14: 743–54. [Google Scholar]
- Badre D, Wagner AD. Left ventrolateral prefrontal cortex and the cognitive control of memory. Neuropsychologia 2007; 45: 2883–901. [DOI] [PubMed] [Google Scholar]
- Badre D, Poldrack RA, Paré-Blagoev EJ, Insler RZ, Wagner AD. Dissociable controlled retrieval and generalized selection mechanisms in ventrolateral prefrontal cortex. Neuron 2005; 47: 907–18. [DOI] [PubMed] [Google Scholar]
- Belke E, Meyer AS, Damian MF. Refractory effects in picture naming as assessed in a semantic blocking paradigm. Q J Exp Psychol A 2005; 58: 667–92. [DOI] [PubMed] [Google Scholar]
- Berthier ML. Transcortical aphasias. Hove: Psychology Press; 1999. [Google Scholar]
- Binder JR, Desai RH, Graves WW, Conant LL. Where is the semantic system? A critical review and meta-analysis of 120 functional neuroimaging studies. Cereb Cortex 2009; 19: 2767–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Binney RJ, Embleton KV, Jefferies E, Parker GJM, Lambon Ralph MA. The ventral and inferolateral aspects of the anterior temporal lobe are crucial in semantic memory: evidence from a novel direct comparison of distortion-corrected fMRI, rTMS, and semantic dementia. Cereb Cortex 2010; 20: 2728–38. [DOI] [PubMed] [Google Scholar]
- Blumstein SE, Baker EH, Goodglass H. Phonological factors in auditory comprehension in aphasia. Neuropsychologia 1977; 15: 19–30. [DOI] [PubMed] [Google Scholar]
- Blumstein SE, Milberg W, Brown T, Hutchinson A, Kurowski K. The mapping from sound structure to the lexicon in aphasia: evidence from rhyme and repetition priming. Brain Lang 2000; 72: 75–99. [DOI] [PubMed] [Google Scholar]
- Bozeat S, Lambon Ralph MA, Patterson K, Garrard P, Hodges JR. Non-verbal semantic impairment in semantic dementia. Neuropsychologia 2000; 38: 1207–15. [DOI] [PubMed] [Google Scholar]
- Bozeat S, Lambon Ralph MA, Graham KS, Patterson K, Wilkin H, Rowland J, et al. A duck with four legs: investigating the structure of conceptual knowledge using picture drawing in semantic dementia. Cognitive Neuropsychology 2003; 20: 27–47. [DOI] [PubMed] [Google Scholar]
- Bright P, Moss H, Tyler LK. Unitary vs multiple semantics: PET studies of word and picture processing. Brain Lang 2004; 89: 417–32. [DOI] [PubMed] [Google Scholar]
- Buchsbaum BR, Hickok G, Humphries C. Role of left posterior superior temporal gyrus in phonological processing for speech perception and production. Cogn Sci 2001; 25: 663–78. [Google Scholar]
- Burgess PW, Shallice T. The hayling and brixton tests. Bury St Edmunds: Thames Valley Test Company; 1997. [Google Scholar]
- Butler RA, Lambon Ralph MA, Woollams AM. Capturing multidimensionality in stroke aphasia: mapping principal behavioural components to neural structures. Brain 2014; 137: 3248–66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campanella F, Shallice T. Refractoriness and the healthy brain: a behavioural study on semantic access. Cognition 2011; 118: 417–31. [DOI] [PubMed] [Google Scholar]
- Campanella F, Mondani M, Skrap M, Shallice T. Semantic access dysphasia resulting from left temporal lobe tumours. Brain 2009; 132: 87–102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campanella F, Crescentini C, Mussoni A, Skrap M. Refractory semantic access dysphasia resulting from resection of a left frontal glioma. Neurocase 2012; 1–9. [DOI] [PubMed] [Google Scholar]
- Chee MW, Weekes B, Lee KM, Soon CS, Schreiber A, Hoon JJ, et al. Overlap and dissociation of semantic processing of Chinese characters, English words, and pictures: evidence from fMRI. Neuroimage 2000; 12: 392–403. [DOI] [PubMed] [Google Scholar]
- Chertkow H, Bub D, Deaudon C, Whitehead V. On the status of object concepts in aphasia. Brain Lang 1997; 58: 203–32. [DOI] [PubMed] [Google Scholar]
- Cohen R, Kelter S, Woll G. Analytical competence and language impairment in aphasia. Brain Lang 1980; 58: 203–32. [DOI] [PubMed] [Google Scholar]
- Corbett F, Jefferies E, Lambon Ralph MA. The use of cueing to alleviate recurrent verbal perseverations: evidence from transcortical sensory aphasia. Aphasiology 2008; 22: 362–82. [Google Scholar]
- Corbett F, Jefferies E, Lambon Ralph MA. Exploring multimodal semantic control impairments in semantic aphasia: evidence from naturalistic object use Neuropsychologia 2009a; 47: 2721–31. [DOI] [PubMed] [Google Scholar]
- Corbett F, Jefferies E, Lambon Ralph MA. Deregulated semantic cognition follows prefrontal and temporoparietal damage: evidence from the impact of task constraint on non-verbal object use. J Cogn Neurosci 2011; 23: 1125–35. [DOI] [PubMed] [Google Scholar]
- Corbett F, Jefferies E, Ehsan S, Lambon Ralph MA. Different impairments of semantic cognition in semantic dementia and semantic aphasia: evidence from the non-verbal domain. Brain 2009b; 132: 2593–608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crawford JR, Garthwaite PH, Porter S. Point and interval estimates of effect sizes for the case-controls design in neuropsychology: rationale, methods, implementations, and proposed reporting standards. Cogn Neuropsychol 2010; 27: 245–260. [DOI] [PubMed] [Google Scholar]
- Crutch SJ, Warrington EK. The influence of refractoriness upon comprehension of non-verbal auditory stimuli. Neurocase 2008; 14: 494–507 [DOI] [PubMed] [Google Scholar]
- Damasio AR. Aphasia. N Engl J Med 1992; 326: 531–9. [DOI] [PubMed] [Google Scholar]
- Damasio H, Damasio AR. Lesion analysis in neuropsychology. New York, NY: Oxford University Press; 1989. [Google Scholar]
- Damian MF, Vigliocco G, Levelt WJM. Effects of semantic context in the naming of pictures and words. Cognition 2001; 81: B77–86. [DOI] [PubMed] [Google Scholar]
- De Renzi E, Faglioni P, Scotti G, Spinnler H. Impairment in associating colour to form concomitant with aphasia. Brain 1972; 95: 293–304. [DOI] [PubMed] [Google Scholar]
- Dell GS, Schwartz MF, Martin N, Saffran EM, Gagnon DA. Lexical access in aphasic and nonaphasic speakers. Psychol Rev 1997; 104: 801–38. [DOI] [PubMed] [Google Scholar]
- Dronkers NF, Wilkins DP, Van Valin RD, Redfern BB, Jaeger JJ. Lesion analysis of the brain areas involved in language comprehension. Cognition 2004; 92: 145–77. [DOI] [PubMed] [Google Scholar]
- Duncan J. The multiple-demand (MD) system of the primate brain: mental programs for intelligent behaviour. Trends Cogn Sci 2010; 14: 172–9. [DOI] [PubMed] [Google Scholar]
- Eggert GH. Wernicke's work on aphasia: a sourcebook and review. The Hague: Mouton Publishers; 1977. [Google Scholar]
- Ellis AW, Miller D, Sin G. Wernicke's aphasia and normal language processing: a case study in cognitive neuropsychology. Cognition 1983; 15: 111–44. [DOI] [PubMed] [Google Scholar]
- Forde EME, Humphreys GW. A semantic locus for refractory behaviour: implications for access storage distinctions and the nature of semantic memory. Cogn Neuropsychol 1997; 14: 367–402. [Google Scholar]
- Gainotti G. Old and recent approaches to the problem of non-verbal conceptual disorders in aphasic patients. Cortex 2014; 53: 78–89. [DOI] [PubMed] [Google Scholar]
- Gainotti G, Silveri MC, Villa G, Caltagirone C. Drawing objects from memory in aphasia. Brain 1983; 106 (Pt 3): 613–22. [DOI] [PubMed] [Google Scholar]
- Galton CJ, Patterson K, Graham KS, Lambon Ralph MA. Differing patterns of temporal atrophy in Alzheimer's disease and semantic dementia. Neurology 2001; 57: 216-225. [DOI] [PubMed] [Google Scholar]
- Gardner HE, Lambon Ralph MA, Dodds N, Jones T, Eshan S, Jefferies E. The differential contributions of pFC and temporoparietal cortices to multimodal semantic control: exploring refractory effects in semantic aphasia. J Cogn Neurosci 2012; 24: 778–93. [DOI] [PubMed] [Google Scholar]
- Gold BT, Balota DA, Jones SJ, Powell DK, Smith CD, Andersen AH. Dissociation of automatic and strategic lexical-semantics: functional magnetic resonance imaging evidence for differing roles of multiple frontotemporal regions. J Neurosci 2006; 26: 6523–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldblum MC, Albert ML. Phonemic discrimination in sensory aphasia. Int J Mental Health 1972; 1: 25–9. [Google Scholar]
- Goodglass H. Diagnosis of conduction aphasia. SE Kohn. Conduction aphasia. Hillsdale, NJ: Lawrence Erlbaum Associates; 1992. 39-49. [Google Scholar]
- Goodglass H, Kaplan E. The assessment of aphasia and related disorders 2nd edn Philadelphia, PA: Lea & Febiger; 1983. [Google Scholar]
- Goodglass H, Kaplan E, Barresi B. The assessment of aphasia and related disorders. 3rd ednBaltimore, MD: Lippincott Williams & Wilkins. (BDAE) 2001a. [Google Scholar]
- Goodglass H, Kaplan E, Barresi B. Boston diagnostic aphasia examination. Baltimore, MD: Lippincott Williams & Wilkins; 2001b. [Google Scholar]
- Gotts SJ, Plaut DC. The impact of synaptic depression following brain damage: a connectionist account of “access/refractory” and “degraded-store” semantic impairments. Cogn Affect Behav Neurosci 2002; 2: 187–213. [DOI] [PubMed] [Google Scholar]
- Head H. Aphasia and kindred disorders of speech. London: CUP; 1926. [DOI] [PubMed] [Google Scholar]
- Henseler I, Regenbrecht F, Obrig H. Lesion correlates of patholinguistic profiles in chronic aphasia: comparisons of syndrome-, modality- and symptom-level assessment. Brain 2014; 137: 918–30. [DOI] [PubMed] [Google Scholar]
- Hickok G, Poeppel D. Towards a functional neuroanatomy of speech perception. Trends Cogn Sci 2000; 4: 131–8. [DOI] [PubMed] [Google Scholar]
- Hier DB, Mogil SI, Rubin NP, Komros GR. Semantic aphasia: a neglect entity. Brain Lang 1980; 10: 120–31. [DOI] [PubMed] [Google Scholar]
- Hodges JR, Patterson K, Oxbury S, Funnell E. Semantic Dementia - progressive fluent aphasia with temporal-lobe atrophy. Brain 1992; 115: 1783-1806. [DOI] [PubMed] [Google Scholar]
- Hoffman P, Jefferies E, Lambon Ralph MA. Ventrolateral prefrontal cortex plays an executive regulation role in comprehension of abstract words: convergent neuropsychological and repetitive TMS evidence. J Neurosci 2010; 30: 15450–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffman P, Rogers TT, Lambon Ralph MA. Semantic diversity accounts for the “missing” word frequency effect in stroke aphasia: insights using a novel method to quantify contextual variability in meaning. J Cogn Neurosci 2011a; 23: 2432–46. [DOI] [PubMed] [Google Scholar]
- Hoffman P, Jefferies E, Lambon Ralph MA. Remembering ‘zeal’ but not ‘thing’: reverse frequency effects as a consequence of deregulated semantic processing. Neuropsychologia 2011b; 49: 580–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howard D, Patterson K. The pyramid and palm trees test: a test of semantic access from words and pictures. Bury St Edmunds: Thames Valley Test Company; 1992. [Google Scholar]
- Hsiao EY, Schwartz MF, Schnur TT, Dell GS. Temporal characteristics of semantic perservations induced by blocked-cyclic picture naming. Brain Lang 2009; 108: 133–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janse E. Lexical competition effects in aphasia: deactivation of lexical candidates in spoken word processing. Brain Lang 2006; 97: 1–11. [DOI] [PubMed] [Google Scholar]
- Janse E. Spoken-word processing in aphasia: effects of item overlap and item repetition. Brain Lang 2008; 105: 185–98. [DOI] [PubMed] [Google Scholar]
- Jefferies E. The neural basis of semantic cognition: converging evidence from neuropsychology, neuroimaging and TMS. Cortex 2013; 49: 611–25. [DOI] [PubMed] [Google Scholar]
- Jefferies E, Lambon Ralph MA. Semantic impairment in stroke aphasia versus semantic dementia: a case-series comparison. Brain 2006; 129: 2132-2147. [DOI] [PubMed] [Google Scholar]
- Jefferies E, Baker SS, Doran M, Lambon Ralph MA. Refractory effects in stroke aphasia: a consequence of poor semantic control. Neuropsychologia 2007; 45: 1065–79 [DOI] [PubMed] [Google Scholar]
- Jefferies E, Hoffman P, Jones RW, Lambon Ralph MA. The impact of semantic impairment on verbal short-term memory in stroke aphasia and semantic dementia: a comparative study. J Mem Lang 2008; 58: 66–87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jefferies E, Patterson K, Jones RW, Lambon Ralph MA. Comprehension of concrete and abstract words in semantic dementia. Neuropsychology 2009; 23: 492–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kay J, Lesser R, Coltheart M. Psycholinguistic assessments of language processing in aphasia (PALPA). Hove: Lawrence Erlbaum Associates; 1992. [Google Scholar]
- Krieger-Redwood K, Teige C, Davey J, Hymers M, Jefferies E. Conceptual control across modalities: graded specialisation for pictures and words in inferior frontal and posterior temporal cortex. Neuropsychologia 2015, in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lambon Ralph MA. Neurocognitive insights on conceptual knowledge and its breakdown. Philos Trans R Soc B Biol Sci 2014; 369: 20120392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lambon Ralph MA, Lowe C, Rogers TT. Neural basis of category-specific semantic deficits for living things: evidence from semantic dementia, HSVE and a neural network model. Brain 2007; 130: 1127–37. [DOI] [PubMed] [Google Scholar]
- Lambon Ralph MA, Patterson K, Plaut DC. Finite case series or infinite single-case studies? Comments on “Case series investigations in cognitive neuropsychology” by Schwartz and Dell (2010). Cogn Neuropsychol 2011; 28: 466–74. [DOI] [PubMed] [Google Scholar]
- Luria AR. Traumatic Aphasia. Mouton: The Hague; 1970. [Google Scholar]
- Luria AR. The working brain: an introduction to neuropsychology. New York, NY: Basic Books; 1973. [Google Scholar]
- Maess B, Friederici AD, Damian M, Meyer AS, Levelt WJM. Semantic category interference in overt picture naming: sharpening current density localization by PCA. J Cogn Neurosci 2002; 14: 455–62. [DOI] [PubMed] [Google Scholar]
- McNeil JE, Cipolotti L, Warrington EK. The accessibility of proper names. Neuropsychologia 1994; 32: 193–208. [DOI] [PubMed] [Google Scholar]
- Mion M, Patterson K, Acosta-Cabronero J, Pengas G, Izquierdo-Garcia D, Hong YT, et al. What the left and right anterior fusiform gyri tell us about semantic memory. Brain 2010; 133: 3256–68. [DOI] [PubMed] [Google Scholar]
- Mirman D, Britt AE, Chen Q. Effects of phonological and semantic deficits on facilitative and inhibitory consequences of item repetition in spoken word comprehension. Neuropsychologia 2013; 51: 1848–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moss H, Abdallah S, Fletcher P, Bright P, Pilgrim L, Acres K, et al. Selecting among competing alternatives: selection and retrieval in the left inferior frontal gyrus. Cereb Cortex 2005; 15: 1723–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mummery CJ, Patterson K, Price CJ, Ashburner J, Frackowiak RSJ, Hodges JR. A voxel-based morphometry study of semantic dementia: relationship between temporal lobe atrophy and semantic memory. Ann Neurol 2000; 47: 36–45. [PubMed] [Google Scholar]
- Noonan KA, Jefferies E, Corbett F, Lambon Ralph MA. Elucidating the nature of deregulated semantic cognition in semantic aphasia: evidence for the roles of prefrontal and temporo-parietal cortices. J Cogn Neurosci 2010; 22: 1597–613. [DOI] [PubMed] [Google Scholar]
- Noonan KA, Jefferies E, Visser MEJ, Lambon Ralph MA. Going beyond inferior prefrontal involvement in semantic control: evidence for the additional contribution of parietal and posterior middle temporal cortex. J Cogn Neurosci 2013; 25: 1824–50. [DOI] [PubMed] [Google Scholar]
- Ogar JM, Baldo JV, Wilson SM, Brambati SM, Miller BL, Dronkers NF, et al. Semantic dementia and persisting Wernicke's aphasia: linguistic and anatomical profiles. Brain Lang 2011; 117: 28–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okada K, Hickok G. Identification of lexical-phonological networks in the superior temporal sulcus using functional magnetic resonance imaging. Neuroreport 2006; 17: 1293–6. [DOI] [PubMed] [Google Scholar]
- Patterson K, Nestor PJ, Rogers TT. Where do you know what you know? The representation of semantic knowledge in the human brain. Nat Rev Neurosci 2007; 8: 976–87. [DOI] [PubMed] [Google Scholar]
- Poeppel D. Pure word deafness and the bilateral processing of the speech code. Cogn Sci 2001; 25: 679–93. [Google Scholar]
- Raven JC. Coloured progressive matrices sets A, AB, B. London: H. K. Lewis.; 1962. [Google Scholar]
- Robertson IH, Ward T, Ridgeway V, Nimmo-Smith I. The test of everyday attention. London: Thames Valley Test Company; 1994. [Google Scholar]
- Robinson G, Shallice T, Bozzali M, Cipolotti L. Conceptual proposition selection and the LIFG: neuropsychological evidence from a focal frontal group. Neuropsychologia 2010; 48: 1652–63 [DOI] [PubMed] [Google Scholar]
- Robson H, Sage K, Lambon Ralph MA. Wernicke's aphasia reflects a combination of acoustic-phonological and semantic control deficits: a case-series comparison of Wernicke's aphasia, semantic dementia and semantia aphasia. Neuropsychologia 2012; 50: 266–75. [DOI] [PubMed] [Google Scholar]
- Robson H, Grube M, Lambon Ralph MA, Griffiths TD, Sage K. Fundamental deficits of auditory perception in Wernicke's aphasia. Cortex 2013; 49: 1808–22. [DOI] [PubMed] [Google Scholar]
- Schnur TT, Schwartz MF, Brecher A, Hodgson C. Semantic interference during blocked-cyclic naming: evidence from aphasia. J Mem Lang 2006; 54: 199–227. [Google Scholar]
- Schnur TT, Schwartz MF, Kimberg DY, Hirshorn E, Coslett HB, Thompson-Schill SL. Localizing interference during naming: convergent neuroimaging and neuropsychological evidence for the function of Broca's area. Proc Natl Acad Sci USA 2009; 106: 322–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seghier ML, Ramlackhansingh A, Crinion J, Leff AP, Price CJ. Lesion identification using unified segmentation-normalisation models and fuzzy clustering. Neuroimage 2008; 41: 1253–66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Slevc LR, Martin RC, Hamilton AC, Joanisse MF. Speech perception, rapid temporal processing, and the left hemisphere: a case study of unilateral pure word deafness. Neuropsychologia 2011; 49: 216–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Snowden JS, Goulding PJ, Neary D. Semantic dementia: a form of circumscribed cerebral atrophy. Behav Neurol 1989; 2: 167–82. [Google Scholar]
- Soni M, Lambon Ralph MA, Noonan KA, Ehsan S, Hodgson C, Woollams AM. “L” is for tiger: effects of phonological (mis)cueing on picture naming in semantic aphasia. J Neurolinguistics 2009; 22: 538–47. [Google Scholar]
- Tanaka Y, Yamadori A, Mori E. Pure word deafness following bilateral lesions: a psychophysical analysis. Brain 1987; 110: 381-403. [DOI] [PubMed] [Google Scholar]
- Thompson-Schill SL, D'Esposito M, Aguirre GK, Farah MJ. Role of left inferior prefrontal cortex in retrieval of semantic knowledge: a reevaluation. Proc Natl Acad Sci USA 1997; 94: 14792–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson HE, Jefferies E. Semantic control and modality: an input processing deficit in aphasia leading to deregulated semantic cognition in a single modality. Neuropsychologia 2013; 51: 1998–2015. [DOI] [PubMed] [Google Scholar]
- Visser M, Jefferies E, Embleton KV, Lambon Ralph MA. Both the middle temporal gyrus and the ventral anterior temporal area are crucial for multimodal semantic processing: distortion-corrected fMRI evidence for a double gradient of information convergence in the temporal lobes. J Cogn Neurosci 2012; 24: 1766–78. [DOI] [PubMed] [Google Scholar]
- Wagner AD, Desmond JE, Demb JB, Glover G, Gabrieli JD. Semantic repetition priming for verbal and pictorial knowledge: a functional MRI study of left inferior prefrontal cortex. J Cogn Neurosci 1997; 9: 714–26. [DOI] [PubMed] [Google Scholar]
- Warrington EK, Shallice T. Semantic access dyslexia. Brain 1979; 102: 43–63. [DOI] [PubMed] [Google Scholar]
- Warrington EK, McCarthy RA. Category specific access dysphasia. Brain 1983; 106: 859–78. [DOI] [PubMed] [Google Scholar]
- Warrington EK, McCarthy RA. Categories of knowledge - further fractionations and an attempted integration. Brain 1987; 110: 1273–96. [DOI] [PubMed] [Google Scholar]
- Warrington EK, James M. The visual object and space perception battery. Bury St Edmunds: Thames Valley Test Company.; 1991. [Google Scholar]
- Warrington EK, Cipolotti L. Word comprehension - The distinction between refractory and storage impairments. Brain 1996; 119: 611–25. [DOI] [PubMed] [Google Scholar]
- Warrington EK, Crutch SJ. A circumscribed refractory access disorder: a verbal semantic impairment sparing visual semantics. Cognitive Neuropsychology 2004; 21: 299–315 [DOI] [PubMed] [Google Scholar]
- Wechsler D. Wechsler memory scale - Revised (WMS-R). New York, NY: Psychological Corporation; 1987. [Google Scholar]
- Weiner DA, Connor LT, Obler LK. Inhibition and auditory comprehension in Wernicke's aphasia. Aphasiology 2004; 18: 599–609. [Google Scholar]
- Whitney C, Jefferies E, Kircher TTJ. Heterogeneity of the left temporal lobe in semantic representation and control: priming multiple vs. single meanings of ambiguous words. Cereb Cortex 2011a; 21: 831–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitney C, Kirk M, O'Sullivan J, Lambon Ralph MA, Jefferies E. The neural organization of semantic control: TMS evidence for a distributed network in left inferior frontal and posterior middle temporal gyrus. Cereb Cortex 2011b; 21: 1066–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woolgar A, Hampshire A, Thompson R, Duncan J. Adaptive coding of task-relevant information in human fronto-parietal cortex. J Neurosci 2011; 31: 14592–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yee E, Blumstein SE, Sedivy JC. Lexical-semantic activation in Broca's and Wernicke's aphasia: evidence from eye movements. J Cogn Neurosci 2008; 20: 592–612. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
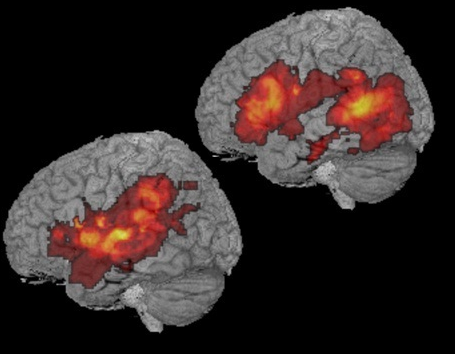{kind=link}