

Abstract
AIM: Hepatic stellate cell (HSC) plays a pivotal role in liver fibrosis and is considered as the therapeutic target for the treatment of hepatic fibrosis. Tyrosine protein kinase plays an important role in the proliferation, activation of HSC. The purpose of the study is to investigate the effects of the tyrosine protein kinase inhibitor genistein on the proliferation and activation of cultured rat HSC.
METHODS: Rat HSC were isolated from Wistar rats by in situ perfusion of collagenase and pronase and single-step density Nycodenz gradient. Culture-activated HSC were serum-starved and incubated with 10-9 to 10-5 mol/L concentration of genistein for 24, 48 or 72 h. In PDGF-induced HSC proliferation, HSC were stimulated with 10 μg·L-1 PDGF-BB for 15 min, and then treated with genistein for the same time. Cell proliferation was measured by MTT assay and based on flow cytometric analysis of cell cycle. The α-smooth muscle actin (α-SMA) expression in HSC was studied with confocal laser microscopy and flow cytometry. c-fos, c-jun and cyclin D1 expression in HSC was also detected by flow cytometry.
RESULTS: Genistein inhibited basal and PDGF-induced proliferation of HSC at the concentration of 10-8 to 10-5 mol/L, and treatment with 10-7 mol/L concentration of genistein for 48 h inhibited the HSC proliferation significantly (the inhibition rate was 70.3%, P < 0.05). Immunofluorescence detected by confocal laser microscopy and flow cytometry showed that treatment with 10-7 mol/L genistein for 48 h suppressed the expression of α-SMA significantly in HSC (the specific fluorescence intensity were 60.2 ± 21.5 vs 35.3 ± 11.6 and 12.8 ± 10.4 vs 9.54 ± 6.39, respectively, both P < 0.05). The intensity of c-fos, c-jun and cyclin D1 expression of HSCs treated with 10-7 mol/L genistein for 48 h was also significantly decreased compared with the controls.
CONCLUSION: Genistein influences proliferation of HSC, suppresses the expression of α-SMA in HSC and t inhibits the intensity of c-fos, c-jun and cyclin D1 expression of HSCs. Genistein has therapeutic potential against liver fibrosis.
INTRODUCTION
Hepatic stellate cells (HSC) are liver mesenchymal cells located in the space of Disse, in close contact with hepatocytes and sinusoidal endothelial cells. In normal liver tissue, they are responsible for vitamin A storage and metabolism. During liver fibrogenesis, HSC undergo a process of activation by acquiring a myofibroblast-like phenotype characterized by increased proliferation and extracellular matrix component synthesis[1-3]. It is known that several cytokine can stimulate HSC proliferation (such as PDGF[4], EGF and bFGF) and collagen synthesis (TGF-β1) in activated HSCs[5-7]. In addition, the activated cells express substantial amounts of α-smooth muscle actin (α-SMA) and show strong contracting activity. Taken together, HSC have been postulated to play critical roles in the development of fibrosis of the liver that was injured by viral infection, alcohol and various drugs[8-11]. Therefore, it is important to find out some agents that block HSC activation, a prerequisite to liver fibrosis.
During culture of HSC in a medium supplemented with serum, they spontaneously undergo activation. This culture-induced activation has been extensively studied as a model of the activation secondary to liver fibrogenesis. This in vitro model is useful to understand the molecular mechanism underlying the activation and/or inactivation of HSC in injured liver. In this context, interferons[12], nitrovasdilators[13], relaxin[14] and pentoxifylling[15] have been shown to inhibit the activation of cultured HSC. Molecular mechanism for the activation of HSC was found to involve intracellular signal cascades and transcriptional regulation of certain genes[16-19].
Recently, tyrosine protein kinase (TPK) signaling pathways were reported to play an important role in the activation of HSC[20]. TPK are involved in signal transduction pathways that control cell proliferation and differentiation and they may be classified into two general groups: (a) membrane receptor tyrosine protein kinases, including EGF-, PDGF-, CFS-, IGF- and insulin receptor[21], and (b) non-receptor linked and cytosolic tyrosine protein kinase[22]. Receptor tyrosine kinase that mediate HSC proliferation include the PDGF receptors[4,23].
Among many polypeptide growth factors potentially involved in chronic liver inflammation, PDGF, a dimmer of two chain referred to as A-chain and B-chain, has been shown to the most potent mitogen for cultured HSC isolated from rat, mouse or human liver[24]. Of the three possible dimeric forms of PDGF (AA, AB and BB), PDGF-BB had been shown to be the most potent in stimulating HSC growth and relative intracellular signaling, in agreement with a predominant expression of PDGF-Rβ (or type B) subunits as compared with the expression of PDGF-Rα (or type A) subunits in activated HSC. Importantly, co-distribution of PDGF with cells expressing the relative receptor subunits (α and β) has been clearly shown after chronic liver tissue damage[23], thus, confirming a functional role of this polypeptide mitogen in the development of hepatic fibrosis.
PDGF receptors are tyrosine protein kinases that are activated by dimerization and autophosphorylation after ligand stimulation. The phosphorylated tyrosine can then associate with intracellular signaling molecules including phospholipase Cγ (PLCγ), phosphatidylinositol 3'-kinase (PI-3K), Src, Grb2-Sos, and Ras-GTPase activating protein. In particular, the attention was focused on PDGF-induced activation of three major pathways recently shown to be highly relevant for transducing the mitogenic effect of this polypeptide mitogen, namely PI-3K, extracellular signal regulated kinase (ERK) and changed in intracellular calcium concentration ([Ca2+]i)[25,26].
In the past decade many efforts were made to develop drugs that inhibit the tyrosine kinase receptors and their intracellular signal transduction pathway[26]. Inhibition of PDGF-R receptor tyrosine kinase activity has been reported for tyrphostin classes of compounds[27]. Tyrphostins are small molecules designed to inhibit tyrosine kinases either through competition for substrates or through adenosine triphosphate binding. Iwamoto et al[28] had shown that Tyrphostin AG1295 reduced the proliferation response of HSC, and their findings suggested a new strategy for the prevention of liver fibrosis with inhibitors of tyrosine kinase receptors. However, whether TPK inhibitors other than tyrphostin also have such effects on HSC is not known. Furthermore, the molecular mechanism by which TPK inhibitors suppress the activation of HSC also remains uncovered.
Genistein (4,5,7-trihydroxyisoflavone), a soybean-derived isoflavone, is a TPK inhibitor that attenuates growth factor- and cytokine- stimulated proliferation of both normal and cancer cells. Extensive epidemiological, in vitro, and animal studies have been performed, and most studies indicated that genistein has beneficial effects on a multitude of human disorders, including cancers, cardiovascular diseases, osteoporosis, and postmenopausal symptoms[29-31]. Although genistein has been shown to dose-dependently inhibit natural and PDGF-BB-induced proliferation and DNA synthesis of aortic smooth muscle cells from stroke-prone spontaneously hypertensive rats[32], it is unknown whether genistein could inhibit the proliferation of activated HSC. The aim of the present study was to investigate the effects of genistein on basal and PDGF-induced HSC proliferation and activation of HSC.
MATERIALS AND METHODS
Materials
Male Wister rats were obtained from Experimental Animal Center of West China Medical Center of Sichuan University (West China University of Medical Sciences, Chengdu, Sichuan). Genistein and Nycodenz were obtained from Sigma (ST. Louis, USA). Dulbecco's modified medium (DMEM), trypsin-EDTA and new-born calf serum (CS) were from GibcoBRL (Maryland, USA). Pronase, Collagenase B and DNAase I were from Roche Molecular Biochemicals, (Mannhein, Germany). Monoclonal Antibodies to Desmin, μ-smooth muscle actin (α-SMA) were obtained from Dako (Glostrup, Denmark). Monoclonal antibody to phosphotyrosine containing protein (p-Tyr), polyclonal antibodies to cyclinD1, c-fos and c-jun were purchased from Santa Cruz Biotechnology (Santa Cruz, USA).
Methods
HSC isolation and culture HSC were isolated from male Wister rats by in situ pronase, collagenase perfusion and single-step Nycodenz gradient according to our previous report[33]. The cells were seeded at a density of 1.5 × 105/cm2 on glass coverslips in 6-well culture plate or 100-mm dishes (Falcon) and maintained in DMEM containing 20 mL·L-1 heat-inactivated new-born calf serum (CS). The purity of HSC preparations was assessed by intrinsic vitamin A autofluorescence and immuocytochemistry with antibody against desmin. The viability of the cells was evaluated by the Trypan blue dye exclusion test. The purity and viability of the primary cells exceeded 90% and 95%, respectively. Therefore, HSC were cultured on uncoated plastic dishes where they spontaneously acquired an activated phenotype, characterized by expression of α-SMA and by loss of vitamin A droplets[34,35]. After reaching confluency (about 10-14 d after plating), activated HSC were detached by incubation with trypsin, and split in a 1:2 ratio. Experiments were performed on cells between the second and 5th passages using 3 independent cell lines, and the purity of activated HSC exceeded 98%.
Cell vialility and proliferation assay The HSC were plated at densities of 2 × 104/mL in 24-well plates and incubated for various time periods and various concentrations with genistein. The viability of cells was analyzed by light microscope, Trypan blue staining[35]. The proliferation of HSC was assessed with 3-(4,5-dimethylthiazol-2-yi)-2,5-diphenyltetrazolium (MTT), as described[36], using cell cultured in 96-well plates. The HSC were plated at a density of 2000 cells/well in 96-well tissue culture dishes and allowed to grow for 24 hrs in DMEM containing 20 mL·L-1 CS under standard tissue culture conditions. After three cell washes with phosphate-buffered saline (PBS), the growth of HSCs was arrested by adding DMEM medium without serum for 24 h. First, HSC were incubated with a concentration of 0 (control) or 10-9 to 10-5 mol/L genistein in fresh DMEM medium without serum and allowed to grow for 24, 48 or 72 h. Second, in PDGF-BB (10 μg·L-1), proliferation of HSC was induced for 15 min and cells were incubated with a concentration of 0 (control) or 10-9 to 10-5 mol/L genistein for 24, 48 or 72 h. Four replicates were used for each experimental point. Absorbance values for cell-free wells were subtracted from all values.
Cell cycle analyze by flow cytometry After 24 h of serum deprivation, HSC were stimulated with or without 10 μg·L-1 PDGF-BB for 15 min and treated with genistein. At the indicated time, HSC were harvested by trypsin-EDTA, washed twice with PBS, and fixed in 700 mL·L-1 ethanol overnight at 4 °C. After washing once with PBS, fixed cells were stained with propidium iodide (50 mg·L-1) in the presence of RNase A (100 mg·L-1), and the cell cycle distribution was analyzed using the FACScan (Coulter, EPICS ELITE ESP model), with a cell cycle analysis program.
Immunoflurescence and confocal laser microscopy Rat HSC were cultured on glass cover slips in DMEM medium containing 20 mL·L-1 CS for 24 h, left in serum-free DMEM medium for an additional 24 h, and then incubated with genistein (10-9 - 19-5 mol/L) for 24, 48 or 72 h. In PDGF treatment, genestein was added after incubated with PDGF-BB (10 μg·L-1) for 15 min. The cells were fixed with 4 mg·L-1 buffered paraformaldehyde with a pH of 7.4 for 20 min and then permeabilized for 10 min with PBS containing 0.1 mg·L-1 Triton X-100. The cells were then stained for indirect immunoflurescence using either a monoclonal antibody against α-SMA diluted 1:100 in PBS or a monoclonal antibody against phospho-tyrosine containing protein diluted 1:50 in PBS as primary antibodies. A goat anti-mouse fluorescein isothiocyanate (FITC)-conjugated affinity-purified antibody was used as secondary antibodies at 1:200 dilution in PBS. Cells were analyzed with a laser scanner confocal microscope Bio-Rad MRC 1024ES equipped with a Nikon (Tokyo, Japan) Diaphot inverted microscope.
Immunoflurescent detection of α-SMA, cyclinD1, c-fos and c-jun by flow cytometry Rat HSC were cultured in plastic 100 mm dishes (Nunc) in DMEM medium containing 20 mL·L-1 CS for 24 h, left in serum-free DMEM medium for an additional 24 h, and then incubated with Genistein (10-9-10-5 mol/L) for 24, 48 or 72 h. After treatment, the cells were detached by trypsin and fixed with 4 mg·L-1 buffered paraformaldehyde with a pH of 7.4 for 20 min and then permeabilized for 10 min with PBS containing 0.1 mg·L-1 Trixon X-100. The cells were then stained for indirect immunoflurescence using either a monoclonal antibody against α-SMA diluted 1:50 in PBS or a polyclonal antibody against cyclin D1, c-fos and c-jun diluted 1:50 in PBS as primary antibodies. A goat anti-mouse fluorescein isothiocyanate (FITC)-conjugated affinity-purified antibody and a goat anti-rabbit FITC-conjugated antibody were used as secondary antibodies at 1:200 dilution in PBS. Cells were analyzed on a FACScan (Coulter, EPICS ELITE ESP model) to determine the expression of α-SMA, cyclin D1, c-fos and c-jun.
Statistical analysis
Results of cell cycle analyze were expressed as percentage of total examined cells and statistical analysis was performed by χ² test. Other results were expressed as ¯x ± s. Differences between means were analyzed with student t test for paired samples. A value of P < 0.05 was considered statistically significant.
RESULTS
Effects of genistein on the viability of HSC
In order to determine toxic effects of genistein on rat HSC, they were treated for 3 d with concentration from 10-9 to 10-5 mol/L of genistein and analyzed by light microscope and trypan blue staining. Genistein concentration of 1 × 10-5 mol/L or lower concentrations had no detectable effects on the cellular morphology or Trypan blue staining.
Effects of Genistein on the proliferation of HSC
First, we observed effects of genistein on the proliferation of cultured HSC. As shown in Figure 1, genistein inhibited proliferation of HSC at the concentration of 10-8 to 10-5 mol/L, and treatment with 10-7 mol/L concentration of genistein for 48 h inhibited the HSC proliferation significantly (the inhibition rate was 70.3%, and the absorbance value was 0.042 ± 0.014 vs 0.151 ± 0.055, t = 3.51, P < 0.05).
Figure 1.
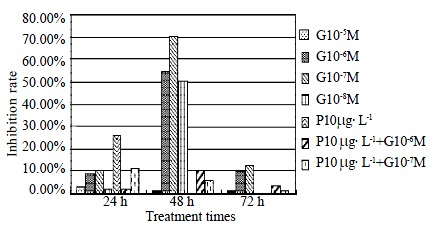
Genistein inhibited the basal and PDGF-BB induced proliferation of HSCs. Cell proliferation was measured by MTT incorporation.
Second, in PDGF-BB-induced proliferation of HSC, the effective concentration for significant inhibition was 10-7 mol/L as shown in Figure 1.
Similar results were obtained with the cell cycle analyze in HSC. As shown in Figure 2, A and B, the percentages of S phase in HSCs of control or PDGF- stimulated groups were 24% and 40.1% respectively. In contrast, genistein treatment led to a significant inhibition of new DNA synthesis, and the percentages of S phase in HSCs of 10-7 mol/L concentration of genistein treated for 48 h were 16.3% and 19.9% respectively (Figure 2, C and D). (24% vs 16.3%, χ² = 53.01, P < 0.01; 40.1% vs19.9%, χ² = 240.1, P < 0.01).
Figure 2.
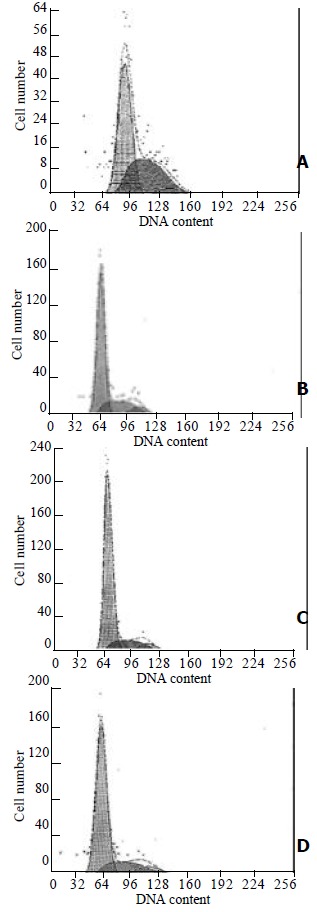
Flow cytometric analysis of cell cycle of HSC. (A) Control HSC; (B) HSCs stimulated with 10 μg·L-1 PDGF-BB for 15 min; (C) HSC treated with 10-7 mol/L genistein for 48 h; (D) HSC stimulated with 10 μg·L-1 PDGF-BB for 15 min and then incubated with 10-7 mol/L genistein for 48 h.
Effects of genistein of the expression of α-SMA in HSC
Smooth muscle α-actin has been accepted for use as one of the indicator of activated HSCs. Immunofluorescence detecting by confocal laser microscopy showed that treatment with 10-7 mol/L genistein for 48 h suppressed the expression of α-SMA significantly (the specific flurescence intensity was 60.2 ± 21.5 vs 35.3 ± 11.6, t = 12.45, P < 0.01, Figure 3). And this observation was also confirmed by detecting the immunofluorescence expression of α-SMA in HSC by flow cytometey, as shown in Figure 4 and Table 1.
Figure 3.
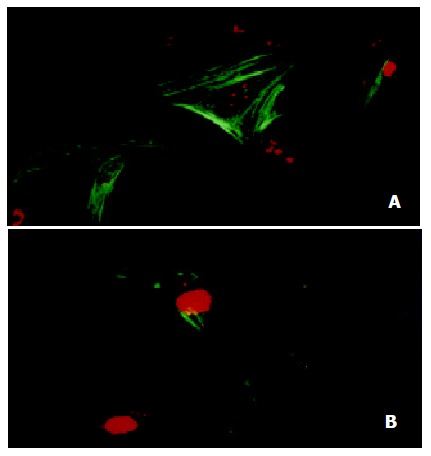
Immunoflurescence detection of α-SMA in HSC by confocal laser microscopy (× 600). (A) Strong staining was observed in control HSC; (B) only a few cells were stained when HSC treated with 10-7 mol/L genistein for 48 h.
Figure 4.
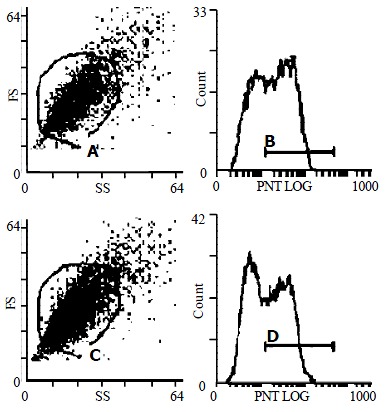
Flow cytometric analysis of α-SMA in HSC. (A, B) control HSC (C, D) HSC treated with 10-7 mol/L genistein for 48 h. Showed significant decreased fluorescence intensity (P < 0.05).
Table 1.
Flow cytometric analysis of α-SMA, c-fos, c-jun and cyclin D1 expression in HSCs (the specific fluorescence intensity)
| Groups | HSC control for 24 h | 10-7 mol/Lgenistein treated for 24 hrs | t test | HSC control for 48 hrs | 10-7 mol/Lgenistein treated for 48 hrs | t test |
| α-SMA Detected | 6.15 ± 4.03 | 5.01 ± 3.30 | t = 15.39 | 12.8 ± 10.4 | 9.54 ± 6.39 | t = 15.03 |
| cell numbers (n) | 4857 | 5086 | P < 0.05 | 2940 | 3985 | P < 0.05 |
| c-fos Detected | 19.4 ± 7.55 | 16.3 ± 5.74 | t = 20.64 | 14.7 ± 5.68 | 12.1 ± 4.18 | t = 18.68 |
| cell numbers (n) | 3122 | 7649 | P < 0.05 | 2412 | 2917 | P < 0.05 |
| c-jun Detected | 17.22 ± 5.49 | 15.69 ± 4.77 | t = 13.32 | 7.99 ± 4.11 | 6.74 ± 2.05 | t = 3.20 |
| cell numbers (n) | 3454 | 5100 | P < 0.05 | 6589 | 4194 | P < 0.05 |
| Cyclin D1 Detected | 25.6 ± 11.1 | 24.0 ± 9.98 | t = 10.23 | 5.76 ± 2.05 | 3.00 ± 1.05 | t = 66.32 |
| cell numbers (n) | 8604 | 9569 | P < 0.05 | 5615 | 5584 | P < 0.05 |
Effects of genistein on tyrosine phosphorylation in HSCs
Immunoflurescence detecting by confocal laser microscopy showed that PDGF-BB stimulated the tyrosine phosphorylation significantly after 10 μg·L-1 PDGF-BB incubated with HSC for 15 min (the specific fluorescence intensity was 27.1 ± 18.8 vs 66.5 ± 36.3, t = 14.65, P < 0.05); but treatment with 10-7 mol/L genistein for 48 h suppressed the expression of tyrosine phosphorylation significantly (the specific fluorescence intensity was 66.5 ± 36.3 vs 46.6 ± 16.4, t = 7.39, P < 0.05, Figure 5).
Figure 5.
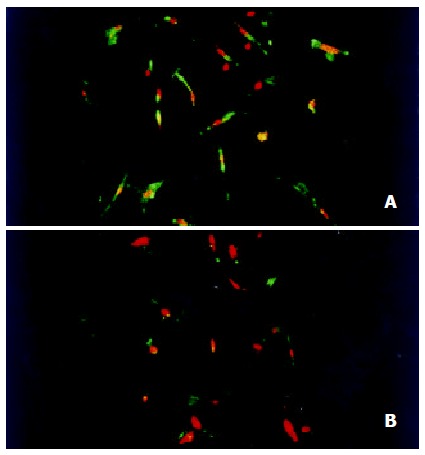
Immunoflurescence detection of the expression of tyrosine phosphorylation in HSC by confocal laser microscopy (× 200). (A) HSC stimulated with 10 μg·L-1 PDGF-BB for 15 min showed a strong staining for phosphoyrosine containing protein; (B) HSCs stimulated with 10 μg·L-1 PDGF-BB for 15 min and then incubated with 10-7 mol/L genistein for 48 h showed significant decreased fluorescence intensity for phosphoyrosine containing protein.
Effects of genistein on expression of c-fos, c-jun and cyclin D1
The expression of c-fos, c-jun and cyclin D1 were remarkably lower in HSCs that incubated with 10-7 mol/L genistein for 48 hrs than in the controls, as shown in Table 1 and Figure 6.
Figure 6.
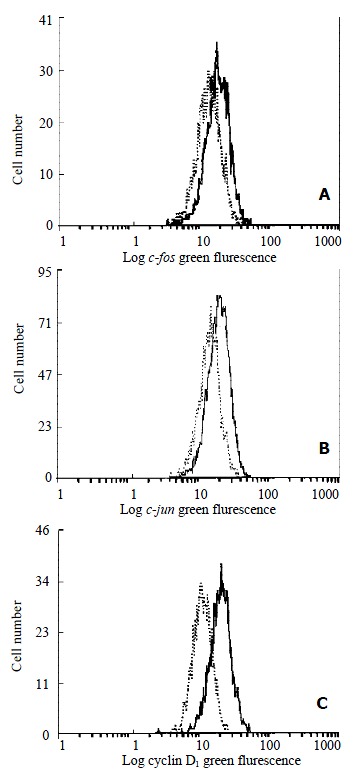
Flow cytometric analysis of c-fos (A), c-jun (B) and cyclin D1 (C) expression in HSC. -indicated the control HSC and -indicated HSC treated with 10-7 mol/L genistein for 48 h.
DISCUSSION
Genistein-a soy derived isoflavone has recently attracted much attention of the medical scientific community. This compound was found to be a potent agent in both prophylaxis and treatment of cancer as well as other chronic diseases. In addition to its proposed actions on prevention cancer, genistein has recently been shown to have antitumor, anti-oxidant and anti-inflammatory effects[29,30]. The great interest that has focused on genistein led to the identification of numerous intracellular targets of its action in the live cells. It was reported[37] that six flavonoids (fisetin, quercetin, apigenin, phloretin, hesperatin and chalcone) inhibited the proliferation of HSCs, but the underlying mechanisms responsible for theirs effects are not yet completely understood. And little is known about the effects of genistein on the HSCs, the cell type primarily involved in hepatic fibrogenesis. Therefore we used an in vitro model in which the activation and proliferation of HSCs were induced by growth on plastic dishes to study the effects of genistein on HSCs and its possible mechanisms.
This culture-induced activation process showed, as far as known, all features of the in vivo process. The present report describes that genistein influences several aspects of the HSC activation. It diminishes proliferation, decreases the expression of α-SMA, c-fos, c-jun and cyclin D1 and these effects of genistein are not due to a toxic cell damage.
In this study, we observed the effect of genistein on the proliferation of activated HSCs. We found that 10-8 to 10-5 mol/L concentration of genistein not only inhibited the basal proliferation of activated HSC, but also inhibited the proliferation of HSCs induced by PDGF-BB. Inhibition of proliferation of HSCs by genistein is not surprising, because reduced cell growth by genistein has been extensively reported. The activated HSCs express receptors for various growth factors, such as PDGF[23,38], IGF[39], and bFGF[40], which are receptor tyrosine protein kinases (RTKs) that have been implicated in HSCs proliferation. One mechanism how genistein might influence the proliferation of HSCs might be the down-regulation of the RTKs in the membrane of HSCs.
In mammalian cells, the Ras/ERK pathway directs signals to the immediate-early genes, such as c-fos, c-jun and other co-regulated genes. c-fos has been reported to function by froming homodimer alone or heterodimer with c-jun, which forms the dimeric transcription factor complex activator protein-1 (AP-1)[41,42]. Recently, it has been demonstrated that the motigen actvited protein kinase (MAPK) family, including c-jun NH2-terminal kinase (JNK) and ERK, plays an important role in the HSC cellular response to stress[43]. Acetadehyde, the major active metabolite of alcohol, is known to stimulate alpha I (I) collagen production in HSC in a JNK-dependent pathway[44]. C-fos and c-jun, two important terminals of various signal transduction pathways and important nuclear factors involved in regulating cytokine gene expression, have been reported to express in activated human HSCs[45]. Recent evidence showed that genistein significantly attenuated the hydrogen peroxide-induced proliferation in aortic smooth muscle cells and c-jun gene expression[46], and the inhibition of c-fos expression, AP-1 transactivation and ERK phosphorylation may contribute to the growth-inhibitory effect of genistein in some breast cell types[47]. The results of our experiments showing an inhibition of c-fos and c-jun expression by genistein further support an effect of this drug on the activated Ras/ERK pathway, including downstream nuclear events leading to HSC cell proliferation.
Another interesting finding is that genistein reduced the protein level of cyclin D1, a cell cycle related protein. Proliferation of eukaryotic cells is generally controlled at stages from G1 to S transition phases. G1 cyclins include three forms of cyclin D (D1, D2 and D3) and cyclin E that associate with either cdk2, cdk4 or cdk6, and cyclin D1, D2, and D3 are markers for G1 phase[48]. It was reported that administration of genistein to rats with a acute renal injury decreased the activation of ERK, and the induction of cyclin D1 was also prevented by this treatment[49]. The present study showed that genistein decreased the cellular level of cyclin D1, and this notion was consistent with the results obtained by cell cycle analyze and MTT assay.
Genistein examined in the present study also inhibited the expression of α-SMA in HSCs. Expression of α-SMA is reported to be regulated by the discoidin domain receptor 2 (DDR2), a tyrosine kinase receptor expressed in mesenchymal tissue whose ligand is fibrillar collagen[50]. In general, HSC undergoing proliferation concomitantly express α-SMA[1-3]. Inhibitors for HSC activation, such as candesartan and perindopril[51], the semisynthetic analogue of fumagillin TNP-470[52], interferon[12] and a selective ROCK inhibitor, Y27632[53], block both cellular proliferation and expression of α-SMA. Therefore, proliferation and expression of α-SMA in HSC seem to be triggered by the same signal transduction pathway, although so far a detailed analysis has been lacking. Perhaps, growth-stimulatory signals involve the activation of positive transcription factors in the 5'-flanking region of α-SMA gene. However, the molecular mechanism by which genestein inhibits the expression of α-SMA in HSC should be further studied.
In summary, the present report showed that genistein regulated the proliferation and the expression of α-SMA, c-fos, c-jun and the cell cycle protein D1, thereby modulating functions of HSC. This effect appears partially mediated by a reduction of PDGF-stimulated ERK activity as well as of other intracellular pathways. The mechanisms of action of genistein include its role as weak estrogens, inhibitor of TPK-dependent signal transduction processes and as cellular antioxidants[30,54]. Findings[55,56] suggest that estradiol is a potent inhibitor of HSC activation and transformation, and it suppressed the induction of hepatic fibrosis. So the effects of genistein to HSCs may also include its weak estrogens role. Because HSCs play pathological roles in inflammation and fibrosis of the liver, therapeutic potential of genistein to hepatic fibrosis should be studied further.
Footnotes
Edited by Pagliarini R
Supported by the National Natural Science Foundation of China, No.39800054
References
- 1.Eng FJ, Friedman SL. Fibrogenesis I. New insights into hepatic stellate cell activation: the simple becomes complex. Am J Physiol Gastrointest Liver Physiol. 2000;279:G7–G11. doi: 10.1152/ajpgi.2000.279.1.G7. [DOI] [PubMed] [Google Scholar]
- 2.Friedman SL. Molecular regulation of hepatic fibrosis, an integrated cellular response to tissue injury. J Biol Chem. 2000;275:2247–2250. doi: 10.1074/jbc.275.4.2247. [DOI] [PubMed] [Google Scholar]
- 3.Albanis E, Friedman SL. Hepatic fibrosis. Pathogenesis and principles of therapy. Clin Liver Dis. 2001;5:315–34, v-vi. doi: 10.1016/s1089-3261(05)70168-9. [DOI] [PubMed] [Google Scholar]
- 4.Kinnman N, Goria O, Wendum D, Gendron MC, Rey C, Poupon R, Housset C. Hepatic stellate cell proliferation is an early platelet-derived growth factor-mediated cellular event in rat cholestatic liver injury. Lab Invest. 2001;81:1709–1716. doi: 10.1038/labinvest.3780384. [DOI] [PubMed] [Google Scholar]
- 5.Xiang DD, Wei YL, Li QF. Molecular mechanism of transforming growth factor β1 on Ito cell. Shijie Huaren Xiaohua Zazhi. 1999;7:980–981. [Google Scholar]
- 6.Liu F, Liu JX. Role of transforming growth factor beta1 in hepatic fibrosis. Shijie Huaren Xiaohua Zazhi. 2000;8:86–88. [Google Scholar]
- 7.Liu F, Wang XM, Liu JX, Wei MX. Relationship between serum TGF-β1 of chronic hepatitis B and hepatic tissue pathology and hepatic fibrosis quantity. Shijie Huaren Xiaohua Zazhi. 2000;8:528–531. [Google Scholar]
- 8.Bataller R, Brenner DA. Hepatic stellate cells as a target for the treatment of liver fibrosis. Semin Liver Dis. 2001;21:437–451. doi: 10.1055/s-2001-17558. [DOI] [PubMed] [Google Scholar]
- 9.Wang FS, Wu ZZ. Current situation in studies of gene therapy for liver cirrhosis and liver fibrosis. Shijie Huaren Xiaohua Zazhi. 2000;8:371–373. [Google Scholar]
- 10.Dai WJ, Jiang HC. Advances in gene therapy of liver cirrhosis: a review. World J Gastroenterol. 2001;7:1–8. doi: 10.3748/wjg.v7.i1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Yao XX. Diagnosis and treatment of liver fibrosis. Shijie Huaren Xiaohua Zazhi. 2000;8:681–689. [Google Scholar]
- 12.Cheng ML, Wu YY, Huang KF, Luo TY, Ding YS, Lu YY, Liu RC, Wu J. Clinical study on the treatment of liver fibrosis due to hepatitis B by IFN-alpha (1) and traditional medicine preparation. World J Gastroenterol. 1999;5:267–269. doi: 10.3748/wjg.v5.i3.267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Failli P, DeFRANCO RM, Caligiuri A, Gentilini A, Romanelli RG, Marra F, Batignani G, Guerra CT, Laffi G, Gentilini P, et al. Nitrovasodilators inhibit platelet-derived growth factor-induced proliferation and migration of activated human hepatic stellate cells. Gastroenterology. 2000;119:479–492. doi: 10.1053/gast.2000.9354. [DOI] [PubMed] [Google Scholar]
- 14.Williams EJ, Benyon RC, Trim N, Hadwin R, Grove BH, Arthur MJ, Unemori EN, Iredale JP. Relaxin inhibits effective collagen deposition by cultured hepatic stellate cells and decreases rat liver fibrosis in vivo. Gut. 2001;49:577–583. doi: 10.1136/gut.49.4.577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wu YA, Kong XT. Anti-hepatic fibrosis effect of pentoxifylling. Shijie Huaren Xiaohua Zazhi. 1999;7:265–266. [Google Scholar]
- 16.Huang GC, Zhang JS. Intercellular signal transduction of activated hepatic stellate cells. Shijie Huaren Xiaohua Zazhi. 2001;9:1056–1060. [Google Scholar]
- 17.Svegliati-Baroni G, Ridolfi F, Di Sario A, Saccomanno S, Bendia E, Benedetti A, Greenwel P. Intracellular signaling pathways involved in acetaldehyde-induced collagen and fibronectin gene expression in human hepatic stellate cells. Hepatology. 2001;33:1130–1140. doi: 10.1053/jhep.2001.23788. [DOI] [PubMed] [Google Scholar]
- 18.Lang A, Brenner DA. Gene regulation in hepatic stellate cell. Ital J Gastroenterol Hepatol. 1999;31:173–179. [PubMed] [Google Scholar]
- 19.Pinzani M, Marra F. Cytokine receptors and signaling in hepatic stellate cells. Semin Liver Dis. 2001;21:397–416. doi: 10.1055/s-2001-17554. [DOI] [PubMed] [Google Scholar]
- 20.Carloni V, Pinzani M, Giusti S, Romanelli RG, Parola M, Bellomo G, Failli P, Hamilton AD, Sebti SM, Laffi G, et al. Tyrosine phosphorylation of focal adhesion kinase by PDGF is dependent on ras in human hepatic stellate cells. Hepatology. 2000;31:131–140. doi: 10.1002/hep.510310121. [DOI] [PubMed] [Google Scholar]
- 21.Lamorte L, Park M. The receptor tyrosine kinases: role in cancer progression. Surg Oncol Clin N Am. 2001;10:271–288, viii. [PubMed] [Google Scholar]
- 22.Martin GS. The hunting of the Src. Nat Rev Mol Cell Biol. 2001;2:467–475. doi: 10.1038/35073094. [DOI] [PubMed] [Google Scholar]
- 23.Robino G, Parola M, Marra F, Caligiuri A, De Franco RM, Zamara E, Bellomo G, Gentilini P, Pinzani M, Dianzani MU. Interaction between 4-hydroxy-2,3-alkenals and the platelet-derived growth factor-beta receptor. Reduced tyrosine phosphorylation and downstream signaling in hepatic stellate cells. J Biol Chem. 2000;275:40561–40567. doi: 10.1074/jbc.M007694200. [DOI] [PubMed] [Google Scholar]
- 24.Kinnman N, Hultcrantz R, Barbu V, Rey C, Wendum D, Poupon R, Housset C. PDGF-mediated chemoattraction of hepatic stellate cells by bile duct segments in cholestatic liver injury. Lab Invest. 2000;80:697–707. doi: 10.1038/labinvest.3780073. [DOI] [PubMed] [Google Scholar]
- 25.Betsholtz C, Karlsson L, Lindahl P. Developmental roles of platelet-derived growth factors. Bioessays. 2001;23:494–507. doi: 10.1002/bies.1069. [DOI] [PubMed] [Google Scholar]
- 26.Ostman A, Heldin CH. Involvement of platelet-derived growth factor in disease: development of specific antagonists. Adv Cancer Res. 2001;80:1–38. doi: 10.1016/s0065-230x(01)80010-5. [DOI] [PubMed] [Google Scholar]
- 27.Morin MJ. From oncogene to drug: development of small molecule tyrosine kinase inhibitors as anti-tumor and anti-angiogenic agents. Oncogene. 2000;19:6574–6583. doi: 10.1038/sj.onc.1204102. [DOI] [PubMed] [Google Scholar]
- 28.Iwamoto H, Nakamuta M, Tada S, Sugimoto R, Enjoji M, Nawata H. Platelet-derived growth factor receptor tyrosine kinase inhibitor AG1295 attenuates rat hepatic stellate cell growth. J Lab Clin Med. 2000;135:406–412. doi: 10.1067/mlc.2000.105974. [DOI] [PubMed] [Google Scholar]
- 29.Goldwyn S, Lazinsky A, Wei H. Promotion of health by soy isoflavones: efficacy, benefit and safety concerns. Drug Metabol Drug Interact. 2000;17:261–289. doi: 10.1515/dmdi.2000.17.1-4.261. [DOI] [PubMed] [Google Scholar]
- 30.Polkowski K, Mazurek AP. Biological properties of genistein. A review of in vitro and in vivo data. Acta Pol Pharm. 2000;57:135–155. [PubMed] [Google Scholar]
- 31.Wang HK. The therapeutic potential of flavonoids. Expert Opin Investig Drugs. 2000;9:2103–2119. doi: 10.1517/13543784.9.9.2103. [DOI] [PubMed] [Google Scholar]
- 32.Pan W, Ikeda K, Takebe M, Yamori Y. Genistein, daidzein and glycitein inhibit growth and DNA synthesis of aortic smooth muscle cells from stroke-prone spontaneously hypertensive rats. J Nutr. 2001;131:1154–1158. doi: 10.1093/jn/131.4.1154. [DOI] [PubMed] [Google Scholar]
- 33.Liu XJ, Yang L, Wu HB, Qiang O, Huang MH, Wang YP. Apoptosis of rat hepatic stellate cells induced by anti-focal adhesion kinase antibody. World J Gastroenterol. 2002;8:734–738. doi: 10.3748/wjg.v8.i4.734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Huang GC, Zhang JS, Zhang YE. Effects of retinoic acid on proliferation, phenotype and expression of cyclin-dependent kinase inhibitors in TGF-beta1-stimulated rat hepatic stellate cells. World J Gastroenterol. 2000;6:819–823. doi: 10.3748/wjg.v6.i6.819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wang JY, Zhang QS, Guo JS, Hu MY. Effects of glycyrrhetinic acid on collagen metabolism of hepatic stellate cells at different stages of liver fibrosis in rats. World J Gastroenterol. 2001;7:115–119. doi: 10.3748/wjg.v7.i1.115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Faouzi S, Lepreux S, Bedin C, Dubuisson L, Balabaud C, Bioulac-Sage P, Desmoulière A, Rosenbaum J. Activation of cultured rat hepatic stellate cells by tumoral hepatocytes. Lab Invest. 1999;79:485–493. [PubMed] [Google Scholar]
- 37.Zhang M, Zhang JP, Ji HT, Wang JS, Qian DH. Effect of six flavonoids on proliferation of hepatic stellate cells in vitro. Acta Pharmacol Sin. 2000;21:253–256. [PubMed] [Google Scholar]
- 38.Di Sario A, Svegliati Baroni G, Bendia E, Ridolfi F, Saccomanno S, Ugili L, Trozzi L, Marzioni M, Jezequel AM, Macarri G, et al. Intracellular pH regulation and Na+/H+ exchange activity in human hepatic stellate cells: effect of platelet-derived growth factor, insulin-like growth factor 1 and insulin. J Hepatol. 2001;34:378–385. doi: 10.1016/s0168-8278(00)00062-3. [DOI] [PubMed] [Google Scholar]
- 39.Svegliati-Baroni G, Ridolfi F, Di Sario A, Casini A, Marucci L, Gaggiotti G, Orlandoni P, Macarri G, Perego L, Benedetti A, et al. Insulin and insulin-like growth factor-1 stimulate proliferation and type I collagen accumulation by human hepatic stellate cells: differential effects on signal transduction pathways. Hepatology. 1999;29:1743–1751. doi: 10.1002/hep.510290632. [DOI] [PubMed] [Google Scholar]
- 40.Fibbi G, Pucci M, Grappone C, Pellegrini G, Salzano R, Casini A, Milani S, Del Rosso M. Functions of the fibrinolytic system in human Ito cells and its control by basic fibroblast and platelet-derived growth factor. Hepatology. 1999;29:868–878. doi: 10.1002/hep.510290343. [DOI] [PubMed] [Google Scholar]
- 41.Thomson S, Mahadevan LC, Clayton AL. MAP kinase-mediated signalling to nucleosomes and immediate-early gene induction. Semin Cell Dev Biol. 1999;10:205–214. doi: 10.1006/scdb.1999.0302. [DOI] [PubMed] [Google Scholar]
- 42.Schinelli S, Zanassi P, Paolillo M, Wang H, Feliciello A, Gallo V. Stimulation of endothelin B receptors in astrocytes induces cAMP response element-binding protein phosphorylation and c-fos expression via multiple mitogen-activated protein kinase signaling pathways. J Neurosci. 2001;21:8842–8853. doi: 10.1523/JNEUROSCI.21-22-08842.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Reeves HL, Dack CL, Peak M, Burt AD, Day CP. Stress-activated protein kinases in the activation of rat hepatic stellate cells in culture. J Hepatol. 2000;32:465–472. doi: 10.1016/s0168-8278(00)80398-0. [DOI] [PubMed] [Google Scholar]
- 44.Chen A, Davis BH. The DNA binding protein BTEB mediates acetaldehyde-induced, jun N-terminal kinase-dependent alphaI (I) collagen gene expression in rat hepatic stellate cells. Mol Cell Biol. 2000;20:2818–2826. doi: 10.1128/mcb.20.8.2818-2826.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Zhu YH, Hu DR, Nie QH, Liu GD, Tan ZX. Study on activation and c-fos, c-jun expression of in vitro cultured human hepatic stellate cells. Shijie Huaren Xiaohua Zazhi. 2000;8:299–302. [Google Scholar]
- 46.Jin N, Hatton ND, Harrington MA, Xia X, Larsen SH, Rhoades RA. H (2)O (2)-induced egr-1, fra-1, and c-jun gene expression is mediated by tyrosine kinase in aortic smooth muscle cells. Free Radic Biol Med. 2000;29:736–746. doi: 10.1016/s0891-5849(00)00376-2. [DOI] [PubMed] [Google Scholar]
- 47.Dampier K, Hudson EA, Howells LM, Manson MM, Walker RA, Gescher A. Differences between human breast cell lines in susceptibility towards growth inhibition by genistein. Br J Cancer. 2001;85:618–624. doi: 10.1054/bjoc.2001.1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Lu B, Dai YM. Abnormal cycle regulation of cells in the HCC. Shijie Huaren Xiaohua Zazhi. 2001;9:205–208. [Google Scholar]
- 49.Ishizuka S, Yano T, Hagiwara K, Sone M, Nihei H, Ozasa H, Horikawa S. Extracellular signal-regulated kinase mediates renal regeneration in rats with myoglobinuric acute renal injury. Biochem Biophys Res Commun. 1999;254:88–92. doi: 10.1006/bbrc.1998.9902. [DOI] [PubMed] [Google Scholar]
- 50.Olaso E, Labrador JP, Wang L, Ikeda K, Eng FJ, Klein R, Lovett DH, Lin HC, Friedman SL. Discoidin domain receptor 2 regulates fibroblast proliferation and migration through the extracellular matrix in association with transcriptional activation of matrix metalloproteinase-2. J Biol Chem. 2002;277:3606–3613. doi: 10.1074/jbc.M107571200. [DOI] [PubMed] [Google Scholar]
- 51.Yoshiji H, Kuriyama S, Yoshii J, Ikenaka Y, Noguchi R, Nakatani T, Tsujinoue H, Fukui H. Angiotensin-II type 1 receptor interaction is a major regulator for liver fibrosis development in rats. Hepatology. 2001;34:745–750. doi: 10.1053/jhep.2001.28231. [DOI] [PubMed] [Google Scholar]
- 52.Wang YQ, Ikeda K, Ikebe T, Hirakawa K, Sowa M, Nakatani K, Kawada N, Kaneda K. Inhibition of hepatic stellate cell proliferation and activation by the semisynthetic analogue of fumagillin TNP-470 in rats. Hepatology. 2000;32:980–989. doi: 10.1053/jhep.2000.18658. [DOI] [PubMed] [Google Scholar]
- 53.Tada S, Iwamoto H, Nakamuta M, Sugimoto R, Enjoji M, Nakashima Y, Nawata H. A selective ROCK inhibitor, Y27632, prevents dimethylnitrosamine-induced hepatic fibrosis in rats. J Hepatol. 2001;34:529–536. doi: 10.1016/s0168-8278(00)00059-3. [DOI] [PubMed] [Google Scholar]
- 54.Barnes S, Boersma B, Patel R, Kirk M, Darley-Usmar VM, Kim H, Xu J. Isoflavonoids and chronic disease: mechanisms of action. Biofactors. 2000;12:209–215. doi: 10.1002/biof.5520120133. [DOI] [PubMed] [Google Scholar]
- 55.Shimizu I, Mizobuchi Y, Yasuda M, Shiba M, Ma YR, Horie T, Liu F, Ito S. Inhibitory effect of oestradiol on activation of rat hepatic stellate cells in vivo and in vitro. Gut. 1999;44:127–136. doi: 10.1136/gut.44.1.127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Yasuda M, Shimizu I, Shiba M, Ito S. Suppressive effects of estradiol on dimethylnitrosamine-induced fibrosis of the liver in rats. Hepatology. 1999;29:719–727. doi: 10.1002/hep.510290307. [DOI] [PubMed] [Google Scholar]