

Abstract
AIM: To study the effect of sulfated cholecystokinin-octapeptide (CCK-8) on systemic hypotension, gene and protein expression of TNF-α in spleen of lipopolysaccharide (LPS) nduced endotoxic shock (ES) rats, and further investigate the signal transduction mechanism of p38 mitogen-activated protein kinase (MAPK).
METHODS: The changes of blood pressure were observed using physiological record instrument in four groups of rats: LPS (8 mg·kg- 1, iv), CCK-8 (40 μg·kg-1, iv) pretreatment 10 min before LPS (8 mg·kg-1), CCK-8 (40 μg·kg-1, iv) or normal saline (control) group. The content of TNF-α in the spleen was assayed 2 h after LPS administration using ELISA kit and the expression of TNF-α mRNA was examined 30 min, 2 h and 6 h after LPS administration by reverse transcribed polymerase chain reaction (RT-PCR). Activation of p38 MAPK was detected with Western blot 30 min after LPS administration.
RESULTS: CCK-8 reversed LPS-induced decrease of mean arterial pressure ( MAP ) in rats. The content of TNF-α in the spleen was (282 ± 30) ng·L-1 in control group, while it increased to (941 ± 149) ng·L-1 in LPS group, P < 0.01. CCK-8 significantly inhibited the LPS-induced increase of TNF-α content in spleen. It decreased to (462 ± 87) ng·L-1 in CCK-8 + LPS group, P < 0.01. The expression of TNF-α mRNA 30 min and 2 h after treatment was stronger in LPS group, while it was lowered after CCK-8 pretreatment.The p38 MAPK expression increased significantly in LPS group (5.84 times of control) and CCK-8 increased the activation of p38 MAPK in ES rats (10.74 times of control).
CONCLUSION: CCK-8 reverses the decrease of MAP in ES rats and has inhibitory effect on the gene and protein expression of TNF-α in spleen, and p38 MAPK may be involved in its signal transduction mechanisms.
INTRODUCTION
Lipopolysaccharide (LPS), main component of Gram-negative bacterial endotoxin[1], is the leading cause of sepsis or endotoxic shock (ES), and when administered experimentally to animals, mimics the same inflammatory response. LPS exerts its effects through cytokines[2]. The pathophysiological changes seen in sepsis are often not due to the infectious organism itself but instead to the uncontrolled production of pro-inflammatory cytokines, including tumor necrosis factor (TNF)-α [3]. TNF-α , which is produced by LPS-activated target cells, is thought to be LPS’s primary mediator[4]. TNF is known to have cytotoxic and cytostatic effects on certain tumor cells, and with a pivotal role in inflammatory reactions and regulation of immunological response[5,6]. TNF-α was mainly produced in the early stage of endotoxemia, and decreased obviously from 6 h to 9 h after challenge[7]. Specific intracellular signaling pathways that modulate cytokine gene expression probably exist in target cells and may represent novel targets for tomorrow’s antisepsis therapies[8]. Mitogen-activated protein kinases (MAPKs) are members of discrete signaling cascades that form focal points for diverse extracellular stimuli and function to regulate fundamental cellular processes[9]. The p38 MAPK, one class of MAPK family, is involved in intracellular signals that regulate a variety of cellular responses during inflammation[10].
CCK-8 possessed both excitatory and inhibitory action on contractile activity of different regions of stomach in guinea pigs[11]. In the spleen, CCK-8 is formed in high abundance in the white pulp where it appears to surround cell clusters. CCK-8 is a chemoattractant for human monocytes and rat macrophages, enhances human eosinophil chemotaxis induced by PAF and LTB4 in allergic patients. Our previous study demonstrated that CCK-8 could protect animals from LPS-induced ES and the protective effect of CCK-8 may be related to its modulation of cytokines [12-14]. Spleen, one of the targets stimulated by LPS, is an important immunological organ. In this study, we examined the expression of TNF-α with reverse transcribed polymerase chain reaction (RT-PCR) and enzyme linked immunoabsorbant assay (ELISA) in spleen and further investigated the mechanism involving p38 MAPK.
MATERIAL AND METHODS
Material
CCK-8 (sulfated), LPS (E.coli LPS, serotype 0111: B4), leupeptin, pepstatin A and Triton X-100 were all purchased from Sigma, and aprotinin from Boehringer. The ELISA kit was purchased from Medsystem (Austria) for assay of TNF-α. Total RNA isolation system and access RT-PCR system were purchased from Promega (USA). Monoclonal anti-p38 MAPK (diphosphorytlated p38) was purchased from Sigma (USA). All other reagents used were of analytic grade. Healthy male Sprague-Dawley rats (n = 52, weighing 150 g-200 g BW) were obtained from Experimental Animal Center of Hebei Province.
Methods
The rats were randomly assigned to four groups and injected different agents via caudal vein. A bolus dose (8 mg·kg-1, 5 g·L-1) of LPS was injected to group receiving LPS and a bolus dose (40 μg·kg-1, 0.05 g·L-1) of CCK-8 was given to the group of CCK-8 + LPS 10 min before injection of LPS. Saline or CCK-8 (40 μg·kg-1) was administered separately to the control or CCK-8 group. Catheter was inserted into femoral artery and Mean arterial pressure (MAP) was detected using physiological record instrument (RM-6000, Japan). The agents were injected through caudal vein after the MAP became steady.Animals were sacrificed at 30 min, 2 h or 6 h after treatment, spleen was rapidly excised, rinsed of blood. The samples were stored at -80 °C. The samples collected at different time points were for the assay of TNF-α mRNA using RT-PCR, samples collected at 2 h were for the assay of TNF-α protein by ELISA and at 30 min for the expression of p38 MAPK by Western blot.
TNF-α dection by ELISA
Frozen tissue samples were weighed and placed in homogenization buffer (4 °C) at a ratio of 100 mg per milliliter of buffer. Buffer contained a protease-inhibitor cocktail including 1 mmol·L-1 phenylmethylsulfonglfluoride (PMSF), 1 mg·L-1 pepstatin A, 1 mg·L-1 aprotinin, and 1 mg·L-1 leupeptin in phosphate-buffered saline solution, pH 7.2, containing 5 g·L-1 Triton X-100. Samples were homogenized and centrifuged at 18000 r·min-1, 4 °C. Tissue supernatants were analyzed for TNF-α using ELISA kit.
Analysis of TNF-α mRNA by RT-PCR
Total RNA was extracted from spleen tissues. The concentration of RNA was determined from absorption at 260 nm. The primers for TNF-α and β-actin were as follows: β-actin (420 bp), 5’-GAGACCTTCAACACCCAGCC-3’, 5’-TCGGGGCATCGGAACCGCTCA-3’; TNF-α (468 bp), 5’-GGATCATCTTCTCAAAACTCG-3’, 5’-TCACAGAGCAATGACTCCAAA-3’. Polymerase chain reactions were performed in a 50 μL reaction volume. RT-PCR reaction was run in the following procedures: 48 °C for 45 min, 1 circle; 94 °C for 2 min, 1 circle; 94 °C for 30 s, 57 °C for 30 s, 68 °C for 1 min, 30 circles; 68 °C for 7 min, 1 circle. 6 μL PCR product was placed on to 15 g·L-1 agarose gel and observed by EB staining using Gel-Pro analyzer.
Detection of p38 MAPK expression by Western blot
Spleen was rapidly excised 30 min after agents administration. Tissues were homogenized with PBS (pH 7.2) and centrifuged at 4 °C, 18000 r·min-1 for 10min.Protein contents were detected in supernatants by coomassie brilliant blue (CBB). Sample loading buffer was added, and each sample was boiled for 8 min prior to loading equal content of protein onto a 12 g·L-1 SDS-polyacrylamide gels, transferred to polyvinylidene fluoride (PVDF) membranes, and incubated with phospho-specific anti-p38 MAPK. Being washed three times in T-PBS, membranes were incubated in horseradish peroxidase-linked secondary antibody for 1 h at room temperature. Membranes were again washed three times with T-PBS and stained with diaminobenzidine (DAB).
Statistical analysis
Data were reported as -x ± s. Statistical differences between values from different groups were determined by one way ANOVA and Newman-Keuls q test. Significance was set at P < 0.05. Gel-Pro Analyzer was used to analyze the PCR and Western results.
RESULTS
Changes of MAP
There was no significant difference between groups before treatment. LPS administration resulted in a significant sustained decrease in MAP during the period of 2 h, decreased to (7.82 ± 0.43) kPa 30 min after LPS administration and restored to (9.33 ± 0.63) kPa by pretreatment with CCK-8 (Figure 1).
Figure 1.
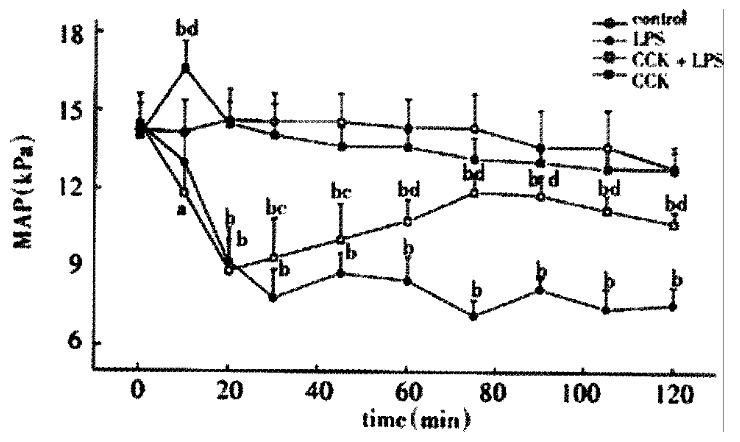
Mean arterial pressure (MAP) of animals injected normal saline, LPS, CCK + LPS and CCK, n = 6. aP < 0.05, bP < 0.01, vs Control, cP < 0.05, dP < 0.01, vs LPS
TNF-α content in spleen 2 h after LPS
Spleen TNF-α content became significantly higher 2 h after administration of LPS as compared to the control animals (941 ± 149) ng·L-1 vs (282 ± 30) ng·L-1, P < 0.01, while CCK-8 significantly inhibited the LPS induced increase of TNF-α (462 ± 87) ng·L-1,P < 0.01. No significant changes were noted in TNF-α content following CCK-8 administration compared with normal saline administration (Figure 2).
Figure 2.
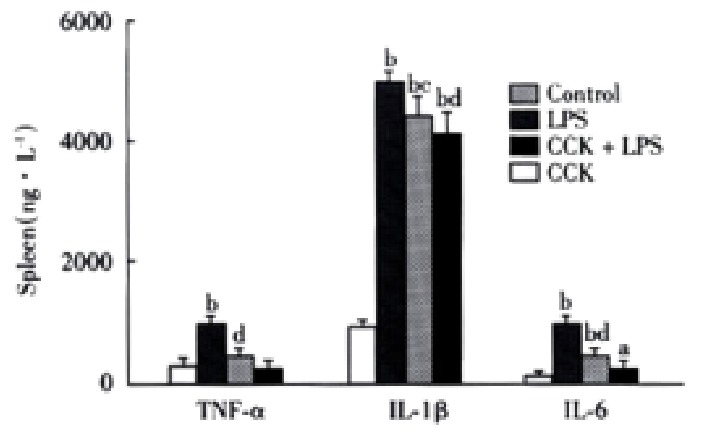
Effects of CCK on TNF-α , IL-1β and IL-6 2 h(TNF-α) or 6 h (IL-1β and IL-6) following LPS administration. n = 6. aP < 0.05, bP < 0.01, vs Control; cP < 0.05, dP < 0.01, vs LPS.
RT-PCR detection of TNF-α
TNF-α mRNA in spleen was detected by RT-PCR analysis. The results showed that the spleen of rats 30 min and 2 h after LPS administration expressed the gene coding for TNF-α because RT-PCR generated a DNA fragment corresponding to the predicted length, 468 bp, of the TNF-α amplification product. The expression of TNF-α decreased in CCK-8 + LPS group compared with LPS group. The ratios of β-actin at 30 min and 2 h were (85% ± 8%) and (57% ± 7%) in LPS group, while it decreased to (30% ± 6%) and (16% ± 2%) respectively in CCK-8 + LPS group. The TNF-α amplification product was not detected 6 h after agents administration. In each tissue sample, all β-actin amplification products were of 420 bp length (Figure 3).
Figure 3.
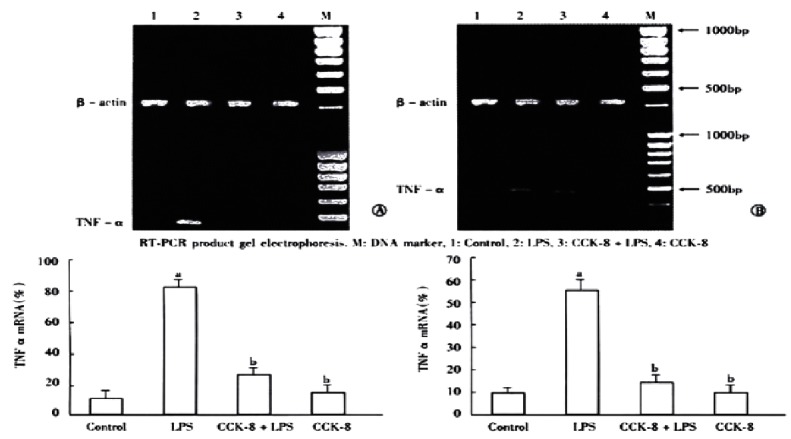
Effect of CCK-8 on LPS-induced TNF-α mRNA expression. Total RNA from the rat spleen was extrated at 30 min(A) or 2h(B) after LPS administration. n = 3. aP < 0.01 vs Control; bP < 0.01 vs LPS.
Analysis of p38 MAPK expression by Western blot
Significant phosphorylation of p38 MAPK was observed in the spleen of rats 30 min after LPS administration, the densitometry units (DU) of LPS group is 5.84 times that of control. CCK-8 can enhance LPS-induced phosphorylation of p38 MAPK significantly, the DU of CCK-8 + LPS group is 10.74 times that of control. Phosphorylation of p38 MAPK was also observed in control and CCK-8 groups (4.64 times of control)(Figure 4).
Figure 4.
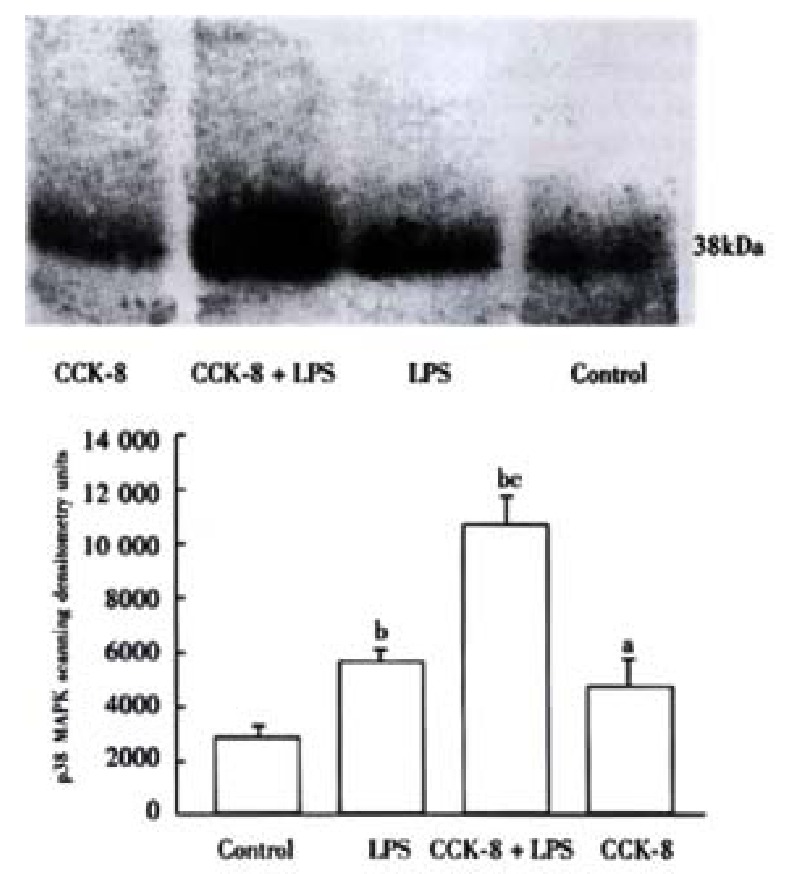
CCK-8 increases p38 MAPK activationon induced by LPSin spleen. aP < 0.05, bP < 0.01 vs Control; cP < 0.01 vs LPS.
DISCUSSION
Although macrophages are the main secretors of TNF-α, other spleen cells, such as lymphocytes, may secret low amounts of TNF-α and may influence macrophage ability to generate and secrete the cytokine[15]. This has led to the suggestion that spleen is an important organ in the production of TNF-α following LPS administration, releasing the cytokines into the circulation, thereby contributing to the elevated serum level of the cytokine. Numerous TNF-α and IL-1β mRNA positive cells were observed using in situ hybridization in the marginal zone and in the red pulp of the spleen in rats after LPS injections, whereas sections from saline-treated animals showed minimal cytokine mRNA expression[16]. LPS resulted in a greater increase in circulating levels of TNF-α which peaked at 90 min and decreased at 150 min after LPS administration[17]. We found that CCK-8 significantly inhibits LPS-induced increase of TNF-α in spleen, which agreed with the results we obtained before[14]. While Cunningham et al[18] reported that CCK-8 stimulated production of TNF-α, IL-1β and IL-6 by monocytes, but was considerably less than LPS response. Later studies[19] suggest that the increase of cytokines induced by CCK-8 may be due to the detection of endotoxin/LPS in medium.
Despite convincing data indicating the protective function of CCK-8 to organism in ES, the precise mechanism remains elusive. Our previous studies showed that CCK can protect pulmonary arterial endothelium against detrimental effects by LPS or TNF-α [20,21] The anti-inflammatory effect of CCK-8 shown in this study may mediate the cell protective function of CCK-8 in ES. The p38 MAPK pathway may be involved. Members of the MAPK cascade are considered to play key roles in signal transduction pathways activated by a wide range of stimuli[22]. CCK is known to activate MAPK signaling pathways[23]. Activated MAPK then tranduces into the nucleus and phosphorylates the ternary complex factor TCF which activates the expression of immediate-early genes, such as c-fos and egr-1[24]. The three best characterized members of this growing family of serine/threonine kinases are extracellular signal regulated kinase (ERK), c-jun N-terminal kinase (JNK) and p38. While ERK responds vigorously to growth factors and certain hormones, JNK and p38 are rather activated by stress stimuli and are widely believed to be part of the cellular stress response machinery[22,25]. The p38 MAPK is associated with immune cell activation, because this kinase is activated by a variety of inflammatory mediators. Challenge of neutrophils with LPS leads to activation of p38 but with slower kinetics than G-protein-coupled chemoattractant receptors[26]. CCK-A and CCK-B receptors are G-protein-coupled[27]. p38 is expressed in the pancreas and rapidly activated by CCK receptor agonist, cerulein[28,29]. p38 inhibition aggravated cerulein pancreatitis, indicating that p38 activation may support protection of the pancreas against damage through hyperstimulation stress. CCK activated p38 MAPK and increased the phosphorylation of HSP27[30]. The p38-MAPKAPK2-HSP27 pathway may be important for organ protection in the pancreas. Other studies indicate that p38 activation can indeed be protective[14]. Sodium salicylate (NaSal) significantly reduced TNF-alpha production in LPS-stimulated macrophages. LPS-stimulated activation of ERK and SAPK/JNK was inhibited by NaSal pretreatment. NaSal treatment of macrophages activated p38 MAPK independent of LPS stimulation[8]. Our data have raised the question of how the increase in p38 MAPK activity, a potent intermediate signal transducer involved in the production of TNF-α after LPS administration, could be associated with decreased production of TNF-α , detected by ELISA and RT-PCR analysis in the presence of CCK-8. LPS-induced production of TNF-α is regulated mainly, but not exclusively, through the p38 MAPK pathway[31]. TNF-α expression in T cells is regulated by several distinct MAPK pathways that functionally cooperate and are critical for transcriptional as well as for posttranscriptional processes[32]. Carbon oxide inhibited the LPS-induced production of TNF-α in mice, which was mediated by p38 MAPK activation[3]. Perhaps a delicate balance exists in the actions of p38, in that subtle cellular activation, as with LPS alone, is stimulatory for TNF-α synthesis, whereas hyperstimulation, as seen in spleen treated with CCK-8 and LPS, becomes inhibitory and thus results in downregulation of TNF-α .This modulation by CCK-8 of LPS-induced production of TNF-α exemplifies accumulating evidence emphasizing the complexity of the molecular regulation of TNF-α expression.
The results of the present study show that administration of CCK-8 prevents LPS-induced decrease of MAP and attenuates LPS-induced increase of TNF-α gene and protein expression in spleen. The different activation of p38 MAPK by LPS or CCK-8 may be involved in their effect on TNF-α production. CCK-8, therefore, might be used therapeutically to treat septic shock syndrome and other inflammatory disease states.
Footnotes
Supported by the Health Committee of Hebei Province (No.2k002), project supported by Science and Technology Department of Hebei Province (01276410D) and project supported by Natural Science Foundation of Hebei Province (No.302490)The paper published on World J Gastroenterol, 2001; 7 (5): 667-671 is key project supported by the Health Committee of Hebei Province (No.2k002), project supported by Science and Technology Department of Hebei Province (01276410D) and project supported by Natural Science Foundation of Hebei Province (No.302490)
Edited by Ma JY
References
- 1.Fan K. Regulatory effects of lipopolysaccharide in murine macrophage proliferation. World J Gastroenterol. 1998;4:137–139. doi: 10.3748/wjg.v4.i2.137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Turrin NP, Gayle D, Ilyin SE, Flynn MC, Langhans W, Schwartz GJ, Plata-Salamán CR. Pro-inflammatory and anti-inflammatory cytokine mRNA induction in the periphery and brain following intraperitoneal administration of bacterial lipopolysaccharide. Brain Res Bull. 2001;54:443–453. doi: 10.1016/s0361-9230(01)00445-2. [DOI] [PubMed] [Google Scholar]
- 3.Otterbein LE, Bach FH, Alam J, Soares M, Tao Lu H, Wysk M, Davis RJ, Flavell RA, Choi AM. Carbon monoxide has anti-inflammatory effects involving the mitogen-activated protein kinase pathway. Nat Med. 2000;6:422–428. doi: 10.1038/74680. [DOI] [PubMed] [Google Scholar]
- 4.Yamakawa T, Eguchi S, Matsumoto T, Yamakawa Y, Numaguchi K, Miyata I, Reynolds CM, Motley ED, Inagami T. Intracellular signaling in rat cultured vascular smooth muscle cells: roles of nuclear factor-kappaB and p38 mitogen-activated protein kinase on tumor necrosis factor-alpha production. Endocrinology. 1999;140:3562–3572. doi: 10.1210/endo.140.8.6914. [DOI] [PubMed] [Google Scholar]
- 5.Bai XY, Jia XH, Cheng LZ, Gu YD. Influence of IFN alpha-2b and BCG on the release of TNF and IL-1 by Kupffer cells in rats with hepatoma. World J Gastroenterol. 2001;7:419–421. doi: 10.3748/wjg.v7.i3.419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Wu RQ, Xu YX, Song XH, Chen LJ, Meng XJ. Adhesion molecule and proinflammatory cytokine gene expression in hepatic sinusoidal endothelial cells following cecal ligation and puncture. World J Gastroenterol. 2001;7:128–130. doi: 10.3748/wjg.v7.i1.128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Zang GQ, Zhou XQ, Yu H, Xie Q, Zhao GM, Wang B, Guo Q, Xiang YQ, Liao D. Effect of hepatocyte apoptosis induced by TNF-alpha on acute severe hepatitis in mouse models. World J Gastroenterol. 2000;6:688–692. doi: 10.3748/wjg.v6.i5.688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Vittimberga FJ, McDade TP, Perugini RA, Callery MP. Sodium salicylate inhibits macrophage TNF-alpha production and alters MAPK activation. J Surg Res. 1999;84:143–149. doi: 10.1006/jsre.1999.5630. [DOI] [PubMed] [Google Scholar]
- 9.Browning DD, Windes ND, Ye RD. Activation of p38 mitogen-activated protein kinase by lipopolysaccharide in human neutrophils requires nitric oxide-dependent cGMP accumulation. J Biol Chem. 1999;274:537–542. doi: 10.1074/jbc.274.1.537. [DOI] [PubMed] [Google Scholar]
- 10.Ohashi N, Matsumori A, Furukawa Y, Ono K, Okada M, Iwasaki A, Miyamoto T, Nakano A, Sasayama S. Role of p38 mitogen-activated protein kinase in neointimal hyperplasia after vascular injury. Arterioscler Thromb Vasc Biol. 2000;20:2521–2526. doi: 10.1161/01.atv.20.12.2521. [DOI] [PubMed] [Google Scholar]
- 11.Li W, Zheng TZ, Qu SY. Effect of cholecystokinin and secretin on contractile activity of isolated gastric muscle strips in guinea pigs. World J Gastroenterol. 2000;6:93–95. doi: 10.3748/wjg.v6.i1.93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ling YL, Huang SS, Wang LF, Zhang JL, Wan M, Hao RL. [Cholecystokinin-octapeptide (CCK-8) reverses experimental endotoxin shock] Shengli Xuebao. 1996;48:390–394. [PubMed] [Google Scholar]
- 13.Ling YL, Huang SS, Zhang JL, Wan M, Hao RL. Effect of cholecysto-kinin on SOD, MDA contents and phagocyte chemiluminescence of reversing endotoxin shock rat. Zhongguo Bingli Shengli Zazhi. 1997;13:483–486. [Google Scholar]
- 14.Ling YL, Meng AH, Zhao XY, Shan BE, Zhang JL, Zhang XP. Effect of cholecystokinin on cytokines during endotoxic shock in rats. World J Gastroenterol. 2001;7:667–671. doi: 10.3748/wjg.v7.i5.667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lahat N, Rahat MA, Brod V, Cohen S, Weber G, Kinarty A, Bitterman H. Abdominal surgery reduces the ability of rat spleen cells to synthesize and secrete active tumour necrosis factor-alpha (TNF-alpha) by a multilevel regulation. Clin Exp Immunol. 1999;115:19–25. doi: 10.1046/j.1365-2249.1999.00758.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Meltzer JC, Sanders V, Grimm PC, Stern E, Rivier C, Lee S, Rennie SL, Gietz RD, Hole AK, Watson PH, et al. Production of digoxigenin-labelled RNA probes and the detection of cytokine mRNA in rat spleen and brain by in situ hybridization. Brain Res Brain Res Protoc. 1998;2:339–351. doi: 10.1016/s1385-299x(98)00010-5. [DOI] [PubMed] [Google Scholar]
- 17.Molina PE, Abumrad NN. Differential effects of hemorrhage and LPS on tissue TNF-alpha, IL-1 and associate neuro-hormonal and opioid alterations. Life Sci. 2000;66:399–409. doi: 10.1016/s0024-3205(99)00606-2. [DOI] [PubMed] [Google Scholar]
- 18.Cunningham ME, Shaw-Stiffel TA, Bernstein LH, Tinghitella TJ, Claus RE, Brogan DA, McMillen MA. Cholecystokinin-stimulated monocytes produce inflammatory cytokines and eicosanoids. Am J Gastroenterol. 1995;90:621–626. [PubMed] [Google Scholar]
- 19.Lieb K, Fiebich BL, Busse-Grawitz M, Hüll M, Berger M, Bauer J. Effects of substance P and selected other neuropeptides on the synthesis of interleukin-1 beta and interleukin-6 in human monocytes: a re-examination. J Neuroimmunol. 1996;67:77–81. doi: 10.1016/0165-5728(96)00034-3. [DOI] [PubMed] [Google Scholar]
- 20.Gu ZY, Ling YL, Meng AH, Cong B, Huang SS. Effect of cholecys-tokinin octapeptide on the response of rabbit pulmonary artery induced by LPS in vitro. Zhongguo Bingli Shengli Zazhi. 1999;15:484–487. [Google Scholar]
- 21.Meng AH, Ling YL, Wang DH, Gu ZY, Li SJ, Zhu TN. [Cholecystokinin-octapeptide alleviates tumor necrosis factor-alpha induced changes in rabbit pulmonary arterial reactivity and injuries of endothelium in vitro] Shengli Xuebao. 2000;52:502–506. [PubMed] [Google Scholar]
- 22.Widmann C, Gibson S, Jarpe MB, Johnson GL. Mitogen-activated protein kinase: conservation of a three-kinase module from yeast to human. Physiol Rev. 1999;79:143–180. doi: 10.1152/physrev.1999.79.1.143. [DOI] [PubMed] [Google Scholar]
- 23.Tapia JA, Ferris HA, Jensen RT, García LJ. Cholecystokinin activates PYK2/CAKbeta by a phospholipase C-dependent mechanism and its association with the mitogen-activated protein kinase signaling pathway in pancreatic acinar cells. J Biol Chem. 1999;274:31261–31271. doi: 10.1074/jbc.274.44.31261. [DOI] [PubMed] [Google Scholar]
- 24.Feng DY, Zheng H, Tan Y, Cheng RX. Effect of phosphorylation of MAPK and Stat3 and expression of c-fos and c-jun proteins on hepatocarcinogenesis and their clinical significance. World J Gastroenterol. 2001;7:33–36. doi: 10.3748/wjg.v7.i1.33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Fleischer F, Dabew R, Göke B, Wagner AC. Stress kinase inhibition modulates acute experimental pancreatitis. World J Gastroenterol. 2001;7:259–265. doi: 10.3748/wjg.v7.i2.259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Browning DD, Windes ND, Ye RD. Activation of p38 mitogen-activated protein kinase by lipopolysaccharide in human neutrophils requires nitric oxide-dependent cGMP accumulation. J Biol Chem. 1999;274:537–542. doi: 10.1074/jbc.274.1.537. [DOI] [PubMed] [Google Scholar]
- 27.Monstein HJ, Nylander AG, Salehi A, Chen D, Lundquist I, Håkanson R. Cholecystokinin-A and cholecystokinin-B/gastrin receptor mRNA expression in the gastrointestinal tract and pancreas of the rat and man. A polymerase chain reaction study. Scand J Gastroenterol. 1996;31:383–390. doi: 10.3109/00365529609006415. [DOI] [PubMed] [Google Scholar]
- 28.Metzler W, Höfken T, Weber H, Printz H, Göke B, Wagner AC. Hyperthermia, inducing pancreatic heat-shock proteins, fails to prevent cerulein-induced stress kinase activation. Pancreas. 1999;19:150–157. doi: 10.1097/00006676-199908000-00008. [DOI] [PubMed] [Google Scholar]
- 29.Wagner AC, Metzler W, Höfken T, Weber H, Göke B. p38 map kinase is expressed in the pancreas and is immediately activated following cerulein hyperstimulation. Digestion. 1999;60:41–47. doi: 10.1159/000007587. [DOI] [PubMed] [Google Scholar]
- 30.Okamoto CT. HSP27 and signaling to the actin cytoskeleton focus on "HSP27 expression regulates CCK-induced changes of the actin cytoskeleton in CHO-CCK-A cells". Am J Physiol. 1999;277:C1029–C1031. doi: 10.1152/ajpcell.1999.277.6.C1029. [DOI] [PubMed] [Google Scholar]
- 31.Wysk M, Yang DD, Lu HT, Flavell RA, Davis RJ. Requirement of mitogen-activated protein kinase kinase 3 (MKK3) for tumor necrosis factor-induced cytokine expression. Proc Natl Acad Sci USA. 1999;96:3763–3768. doi: 10.1073/pnas.96.7.3763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Hoffmeyer A, Grosse-Wilde A, Flory E, Neufeld B, Kunz M, Rapp UR, Ludwig S. Different mitogen-activated protein kinase signaling pathways cooperate to regulate tumor necrosis factor alpha gene expression in T lymphocytes. J Biol Chem. 1999;274:4319–4327. doi: 10.1074/jbc.274.7.4319. [DOI] [PubMed] [Google Scholar]