

Abstract
Prostate cancer (PC) is a serious health problem all over the world. Cell proliferation plays a major role in the tumorigenesis of PC. It is reported that microRNAs (miRNAs) played crucial roles in the regulation of cell proliferation. However, the underlying mechanism of miRNAs in PC has not been intensively investigated. In the present study, the effect of miR-592 on the cell proliferation of PC was investigated. The results showed that miR-592 was significantly upregulated in PC cell and PC tissues. To investigate the biological roles of miR-592, we induced either the up- or downregulation of miR-592 expression by transfecting DU145 PC cells with miR-592 mimics or miR-592 inhibitor. Our results demonstrated that the upregulation of miR-592promoted cell growth, while miR-592 inhibitor showed the opposite effect. Further experiment revealed that miR-592 repressed the expression of FOXO3 by directly targeting the 3’UTR of the FOXO3 transcript, which resulted in upregulating of the expression of cyclin D1 and downregulating of the expression of p21. In sum, our data indicated a novel aspect of the miR-592 in the molecular etiology of PC.
Keywords: miR-592, prostate cancer, FOXO3, cell proliferation
Introduction
Prostate cancer (PC) is one of the most prevalent types of malignant disease and the second leading cause of cancer-related deaths among men [1]. In recent years, the incidence of PC is increasing while the overall fiver-year survival rate is decreasing [2]. Thus, it is essential to understand the mechanism of PC development for finding better treatment.
A growing body of evidence indicated that microRNAs (miRNAs) were non-coding RNA molecules (21-23 nucleotides in length), contributed to cell proliferation, metastasis, invasion, angiogenesis and apoptosis of various types of cancers [3-7]. Aberrant expression of miRNAs functioned as either tumor suppressors or oncogenes by regulating targeted gene expression at the transcriptional or posttranscriptional level [8-10]. For example, miR-490-5p was markedly down-regulated and acted as a tumor suppressor in human bladder cancer [11]. Finding by Fang Y et al. indicated that MiR-744 functioned as a proto-oncogene in nasopharyngeal carcinoma progression and metastasis via transcriptional control of ARHGAP5 [12]. In the current study, we investigated the biological effects and the potential mechanisms of miR-592 on cell proliferation in PC and identified miR-592 as a tumor promoter to induce cell proliferation of PC cells by targeting FOXO3.
Materials and methods
Clinical specimens
Eight human PC clinical tissues and their matched adjacent normal tissues (ANT) were obtained from PC patients at Department of Urology, the second hospital of Jiangxi province Nanchang City (Jiangxi, People’s Republic of China). The study was approved by the ethics committee of the second hospital of Jiangxi province Nanchang City (Jiangxi, People’s Republic of China). All patients gave informed consent in written. Tissue samples were snapped into liquid nitrogen and then stored at -80°C.
Cell culture
Human PC cell lines (M12, Tsu-Pr1, PC3, DU145, 22RV1 and LNCAP) and a non-malignant epithelial prostate cell line (RWPE-1 as N) were purchased from the Shanghai Bioleaf Biotech Co., Ltd (Shanghai, People’s Republic of China). All Prostate cancer cell lines were grown in DulbeccoE-1 as N) were purchased from the Shanghai Bioleaf3.ll proliferation in bovine serum (FBS, Sigma-Aldrich, USA), 100 units/ml of penicillin-streptomycin (Invitrogen, Carlsbad, CA), and RWPE-1 cells (as control) were maintained in keratinocyte serum-free medium (KSFM; GIBCO Laboratories, Grand Island, NY, USA) supplemented with 0.05 mg/ml bovine pituitary extract, 5% L-glutamine and 5 mg/ml EGF. Cells were cultured at 37°C in a humidified atmosphere containing 5% CO2.
Plasmids, small interfering RNA and transfection
The miR-592 mimics, miR-592 inhibitor and the relative negative controls were purchased from Shanghai GenePharma (Shanghai, Peopleai, Peopleinhibitor and the relative negative controls werLipofectamine 2000 reagent (Invitrogen) as recommended by the manufacturer. Four μg of plasmids were transfected to the cells with Lipofectamine 2000 (Invitrogen, Carlsbad, CA) as recommended by the manufacturer’s protocol.
For depletion of FOXO3, small interfering RNA-FOXO3 (siRNA-FOXO3#1, HSH005759; siRNA-FOXO3#2, HSH061727) was synthesized and purified by GeneCopoeia Co. (Guangzhou, People Carlsbad, CA) as recommended by the manufactureleai, Peopleinhibitor and the re 2000 (Invitrogen, Carlsbad, CA) as recommended by the manufacturer’s protocol.
RNA extraction and real-time quantitative PCR
Total RNA was extracted from culture cells and patient samples using Trizol reagent (Invitrogen) according to the manufacturer’s protocol. The PCR assay was performed using the SYBR Premix Ex Taq system (TaKaRa, Madison, WI, USA) according to the manufacturer’s instructions. The relative miR-592 expression levels after normalization to U6 small nuclear RNA were calculated using 2-[(Ct of miR-592)-(Ct of U6)].
For analysis of protein coding genes, real-time PCR was performed using the Applied Biosystems 7500 Sequence Detection system. The following PCR primers were synthesized by GeneCopoeiaTM as followed: Cyclin D1 (HQP016204) and p21 (HQP000331). Expression data were normalized to the geometric mean of GAPDH (HQP064347) to control the variability in expression levels and calculated as 2-[(Ct of CyclinD1 and p21)-(Ct of GAPDH)].
MTT assays and colony formation
For the cell proliferation assay, 5000 DU145 cells were seeded in triplicate in 96-well plates for each transfection group: miR-592, miR-592-in and the relative control mimics, and incubated under conditions of 37°C at 5% CO2. After 1, 2, 3, 4 and 5 day, 20 μL of MTT solution (5 mg/mL, Sigma-Aldrich, USA) was added to each well. The generated formazan was dissolved in DMSO, and the absorbance was recorded.
For the colony formation assay, 500 cells were plated into 6-cm plates and after 14 days, the cells were washed with phosphate-buffered saline (PBS), and stained with 0.1% crystal violet (Sigma-Aldrich) for 1 min after fixed with frozen methanol. The number of positive-staining colonies was counted.
Anchorage-independent growth assay
Cells were trypsinized, and 1000 cells were resuspended in 2 ml complete medium plus 0.3% agar (Sigma-Aldrich). The agar-cell mixture was plated on top of a bottom layer consisting of 1% agar in complete medium. Cells were incubated for two weeks at 37°C until colony formation and then stained with 1% Crystal Violet for counting under microscope and cell colonies were photographed at an original magnification of 100×. Only cell colonies containing more than 50 cells were counted.
Luciferase assays
The pGL3-luciferase reporter gene plasmids pGL3-FOXO3-3’-UTR were co-transfected into the cells with miR-592, miR-592-in or miR-592-mut using Lipofectamine 2000 Reagent (Invitrogen). Firefly and renilla luciferase activities were measured using the Dual-Luciferase Reporter Kit (Promega) according to the manufacturer’s instructions were assayed 48 hours after transfection.
Western blotting
Cell after transfection and the protein were lysed, the equivalent aliquots (30 μg) of proteins were electrophoresed on a 10% SDS-PAGE and transferred onto nitrocellulose membranes. The membrane was incubated overnight at 4°C with anti-FOXO3, anti-Cyclin D1 and anti-p27 (1:1000; Cell Signaling Technology) and anti-α-tubulin antibody (Sigma-Aldrich-Aldrich) acted as the control sample loading and followed by secondary anti-rabbit or anti-mouse. Protein expression was assessed by chemiluminescence (Beyotime Institute of Biotechnology, China).
Statistical analysis
Statistical analysis was performed using the SPSS 17.0 (SPSS Inc, Chicago, IL, USA). Statistical analyses were done by analysis of variance (ANOVA) or Student’s t test. Statistical significance was defined as a value of P < 0.05.
Result
MiR-592 expression was upregulated in PC cell lines and PC tissues
Real-time PCR analysis revealed that miR-592 expression was markedly upregulated in all 6 tested PC cell lines (M12, Tsu-Pr1, PC3, DU145, 22RV1 and LNCAP) compared to the non-malignant epithelial RWPE-1 prostate cell line (Figure 1A) The expression levels of miR-592 were further determined in human PC clinical tissues. As shown in Figure 1B, in comparison with the matched adjacent normal tissues (ANT), PC clinical tissues showed significantly higher miR-592 expression.
Figure 1.
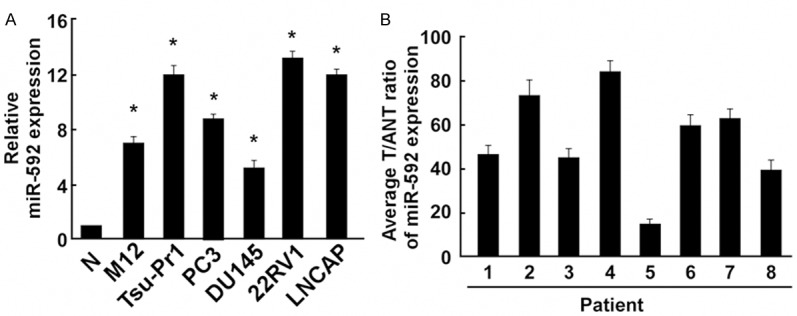
Expression of miR-592 in human Prostate cancer (PC) cell lines and tissues. A: Real-time PCR analysis of miR-592 expression in human non-malignant epithelial prostate cell line (RWPE-1 as N) and PC cell lines, including M12, Tsu-Pr1, PC3, DU145, 22RV1 and LNCAP. B: Relative miR-592 expression levels in 8 paired primary OS tissues (T) and the tumor adjacent normal tissues (ANT) from the same patient were detected by PCR analysis. Each bar represents the mean of three independent experiments. *P < 0.05.
MiR-592 promoted PC cell proliferation, miR-592-in inhibited PC cell proliferation
To gain insight into the functional role of miR-592 in PC cell growth, we transfected with miR-592, miR-592-in and the respective controls into DU145 cells, the result of PCR indicated that both of them showed great transfection efficiency (Figures 2A and 3A). MTT assay and colony formation assays showed miR-592 could increase cell growth compared with the control miRNA (Figure 2B and 2C). Consistent with the effects on cell proliferation, miR-529 markedly increased the anchorage-independent growth of DU145 cells in soft agar compared with the negative control (Figure 2D). Moreover, using the MTT and colony formation assays, compared to NC transfected cells, we discovered that miR-592-in reduced the growth of DU145 PC cells (Figure 3B and 3C). Additionally, miR-592-in also significantly reduced the anchorage-independent growth ability of DU145 PC cells (Figure 3D).
Figure 2.
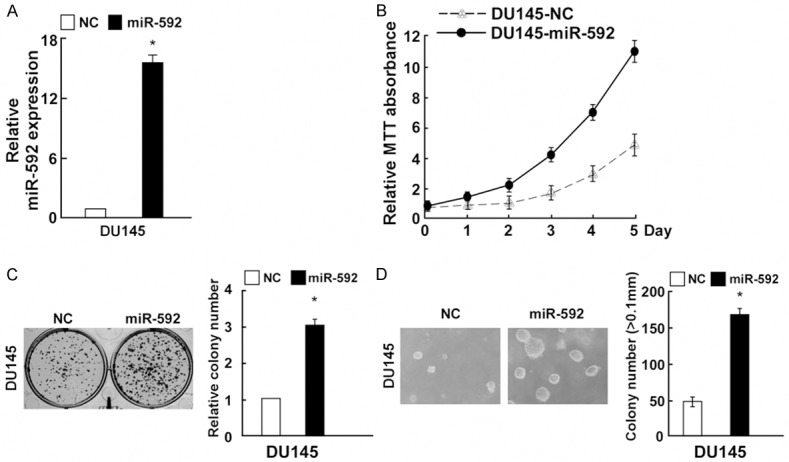
miR-592 upregulation promoted PC cell proliferation. A: Validation of miR-592 expression levels after transfection by PCR analysis. B: MTT assays revealed that inhibition of miR-592 promoted growth of DU145 PC cell line. C: Representative quantification of crystal violet-stained cell colonies. D: Upregulation of miR-592 promoted the anchorage-independent growth of DU145 cells. Representative micrographs (left) and quantification of colonies that were > 0.1 mm (right). Each bar represents the mean of three independent experiments. *P < 0.05.
Figure 3.
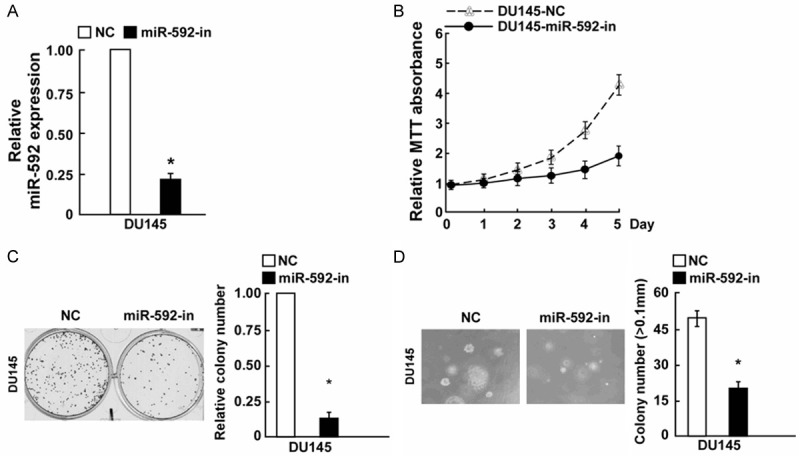
Inhibition of miR-592 inhibited PC cell proliferation. A: Validation of miR-592 expression levels after transfection by PCR analysis. B: MTT assays revealed that upregulation of miR-592 inhibited growth of DU145 PC cell line. C: Representative quantification of crystal violet-stained cell colonies. D: Inhibition of miR-592 inhibited the anchorage-independent growth of DU145 cells. Representative micrographs (left) and quantification of colonies that were > 0.1 mm (right). Each bar represents the mean of three independent experiments. *P < 0.05.
MiR-592 directly targets FOXO3 by binding to its 3’-UTR and altered levels of proteins related to cell proliferation in PC
Using publicly available algorithm TargetScan, we identified FOXO3 as a potential target of miR-592 (Figure 4A). To determine whether miR-592 affects FOXO3 expression, we transfected miR-592 mimics, miR-592-in or the respective controls into DU145 cells. Western blotting analysis showed that compared to the relative control groups, FOXO3 expression was down-regulated in miR-592 groups in DU145 cells, while the protein expression of FOXO3 was up-regulated in miR-592-in group in DU145 cells (Figure 4B). To verify the effect of miR-592 on the inhibition of FOXO3 expression, we examined whether FOXO3 is regulated by miR-592 through direct binding to its 3’UTR. We co-transfected with 3’ UTR of FOXO3 and miR-592, miR-592-in, miR-592-mut or the relative controls. Results of luciferase activity assays revealed that miR-592 significantly suppressed the luciferase activity of reporter genes containing wild-type FOXO3 3’-UTR, while miR-592 showed the opposite effect. Meanwhile, miR-592-mut had no effect on the luciferase activity of FOXO3 3’-UTR, demonstrating that miR-592-mut could not combine to the FOXO3 3’-UTR (Figure 4C). These results indicated that miR-592 directly targeted FOXO3 in PC cells.
Figure 4.
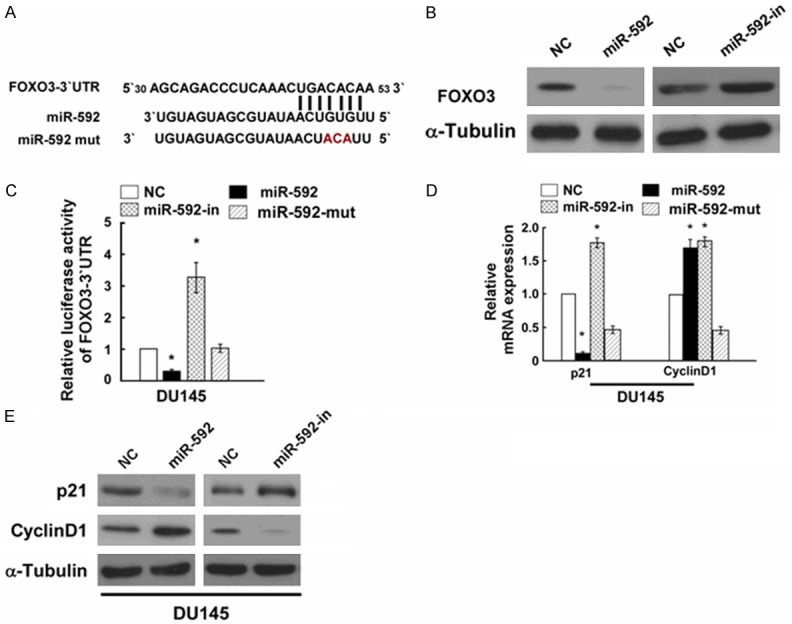
miR-592 suppresses FOXO3 expression by directly targeting the FOXO3 3’-UTR and altered levels of proteins related to cell proliferation. A: Predicted miR-592 target sequence in the 3’-UTR of FOXO3 (FOXO3-3’-UTR) and positions of three mutated nucleotides (red) in the 3’-UTR of miR-592 (miR-592-mut). B: Western blotting analysis of FOXO3 expression in DU145 cells transfected with miR-592 or the miR-592 inhibitor. α-Tubulin served as the loading control. C: Luciferase reporter assay of the indicated cells transfected with the pGL3-FOXO3-3’-UTR reporter and miR-592 or miR-592-in or miR-592 with oligonucleotides. D: Real-time PCR analysis of expression of p21 and Cyclin D1 in indicated DU145 cells. E: Western blotting analysis of expression of p21 and Cyclin D1 protein in DU145 cells. α-Tubulin served as the loading control. *P < 0.05.
As miR-592 promoted cell proliferation of DU145 PC cells, we explored the effect of miR-592 on FOXO3’s down-stream protein p21 and cyclin D1 [13,14], which regulate the cell proliferation. Using Western blotting, we observed that p21 expression was down-regulated, while cyclin D1 was up-regulated by miR-592, further confirming that miR-592 can influence the proliferation of DU145 PC cells (Figure 4E).
FOXO3 downregulation counteracted the proliferation arrest by miR-592-in
To further confirm the role of FOXO3 in DU145 PC cell proliferation, as showed in Figure 5A, we observed that FOXO3 was upregulated in DU145 PC cells after transfected with miR-592-in, but FOXO3-siRNA effectively decreased the expression of FOXO3 in miR-592-in-transfected DU145 PC cells. Furthermore, further experiment (colony formation and anchorage-independent growth assays) showed knockdown of FOXO3 significantly counteracted the proliferation arrest by miR-592-in (Figure 5B and 5C).
Figure 5.
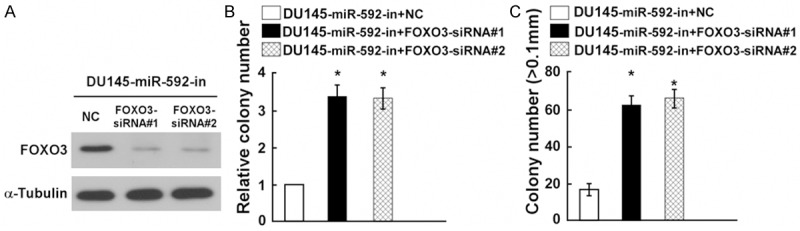
FOXO3 downregulation counteracted the proliferation arrest by miR-592-in. A: Western blot analysis verified that silencing FOXO3 effectively decreased the expression of FOXO3 in miR-592-in-transfected DU145 cells. B: miR-592-in-transfected DU145 cells after transfection with FOXO3-siRNAs promoted cell colonies formation. C: miR-592-in-transfected DU145 cells after transfection with FOXO3-siRNAs promoted the anchorage-independent growth. Representative quantification of colonies that were > 0.1 mm. Each bar represents the mean of three independent experiments. *P < 0.05.
Discussion
Lots of reports indicated that miRNAs were important regulators involved in tumor formation and progression [15-18]. Accumulating evidence has demonstrated that miRNAs acted as either tumor oncogenes or as tumor suppressors [19-21]. Previously, miR-592 was markedly down-regulated in colorectal cancer tissues and cancer cell lines, and overexpression of miR-592 reduced cell proliferation of colorectal cancer cells by directly targeting CCND3 [22]. Furthermore, finding by Liu M. et al indicated that miR-592 functioned as a novel carcinogen-initiated and metastasis-related biomarker in colorectal cancer [23]. In this present study, the key finding was that miR-592 was upregulated in PC cell lines and PC tissues, compared with the non-malignant epithelial RWPE-1 prostate cells and the tumor adjacent normal tissues. Furthermore, overexpression of miR-592 increased cell proliferation and anchorage-independent growth of DU145 PC cells, while miR-592-in showed the opposite effect, indicating that alterations of miR-592 could be involved in PC progression.
Then, we found that miR-592 acted as a tumor promoter gene by targeting FOXO3, which was a member of the forkhead transcriptional factor family, was involved in cell growth, proliferation, and development of cancer [24,25]. This is the first study to show that the FOXO3 is negatively regulated by miR-592 through a specific target site within the 3’-UTR. The down regulation of FOXO3 further affected its down-stream protein p21 and cyclin D1 [13,14], our data suggested that miR-592 promoted cell proliferation of DU145 PC cells by down-regulating p21 expression and up-regulating cyclin D1 expression. Further experiment showed that knockdown of FOXO3 caused the down regulation of FOXO3 in miR-592-in transfected DU145 PC cells, which is much lower than that in scramble. This might due to that the knockdown of FOXO3 reversed the protein level miR-592 promoted. More importantly, result of colony formation and anchorage-independent growth assays revealed that the downregulated FOXO3 regulated the cell proliferation of DU145 PC cells. Collectively, our findings suggested that miR-592 promoted the initiation and progression of PC by regulating FOXO3 expression.
In conclusion, the current study suggested that the miR-592 is frequently up-regulated in PC, which acted as an onco-miRNA in PC cells, and overexpression of miR-592 promoted PC cell proliferation by directly targeting FOXO3. These findings collectively implicated that miR-592 might serve as an important molecular biomarker for the PC patients.
Acknowledgements
This work was supported by Department of Urology, the second hospital of Jiangxi province Nanchang City and Department of Urology, Shandong Jining first people’s Hospital. All authors designed the study together and performed the experiment together; all authors analyzed the data and wrote the paper; all authors approved the final manuscript.
Disclosure of conflict of interest
None.
References
- 1.Siegel R, Ma J, Zou Z, Jemal A. Cancer statistics, 2014. CA Cancer J Clin. 2014;64:9–29. doi: 10.3322/caac.21208. [DOI] [PubMed] [Google Scholar]
- 2.DeSantis CE, Lin CC, Mariotto AB, Siegel RL, Stein KD, Kramer JL, Alteri R, Robbins AS, Jemal A. Cancer treatment and survivorship statistics, 2014. CA Cancer J Clin. 2014;64:252–271. doi: 10.3322/caac.21235. [DOI] [PubMed] [Google Scholar]
- 3.Esquela-Kerscher A, Slack FJ. Oncomirs - microRNAs with a role in cancer. Nat Rev Cancer. 2006;6:259–269. doi: 10.1038/nrc1840. [DOI] [PubMed] [Google Scholar]
- 4.Calin GA, Croce CM. MicroRNA signatures in human cancers. Nat Rev Cancer. 2006;6:857–866. doi: 10.1038/nrc1997. [DOI] [PubMed] [Google Scholar]
- 5.Sun K, Lai EC. Adult-specific functions of animal microRNAs. Nat Rev Genet. 2013;14:535–548. doi: 10.1038/nrg3471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Cao L, Xie B, Yang X, Liang H, Jiang X, Zhang D, Xue P, Chen D, Shao Z. MiR-324-5p Suppresses Hepatocellular Carcinoma Cell Invasion by Counteracting ECM Degradation through Post-Transcriptionally Downregulating ETS1 and SP1. PLoS One. 2015;10:e0133074. doi: 10.1371/journal.pone.0133074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Yang F, Wang W, Zhou C, Xi W, Yuan L, Chen X, Li Y, Yang A, Zhang J, Wang T. MiR-221/222 promote human glioma cell invasion and angiogenesis by targeting TIMP2. Tumour Biol. 2015;36:3763–3773. doi: 10.1007/s13277-014-3017-3. [DOI] [PubMed] [Google Scholar]
- 8.Deng L, Liu H. MicroRNA-506 suppresses growth and metastasis of oral squamous cell carcinoma via targeting GATA6. Int J Clin Exp Med. 2015;8:1862–1870. [PMC free article] [PubMed] [Google Scholar]
- 9.Kong XJ, Duan LJ, Qian XQ, Xu D, Liu HL, Zhu YJ, Qi J. Tumor-suppressive microRNA-497 targets IKKbeta to regulate NF-kappaB signaling pathway in human prostate cancer cells. Am J Cancer Res. 2015;5:1795–1804. [PMC free article] [PubMed] [Google Scholar]
- 10.Wang LQ, Zhang Y, Yan H, Liu KJ, Zhang S. MicroRNA-373 functions as an oncogene and targets YOD1 gene in cervical cancer. Biochem Biophys Res Commun. 2015;459:515–520. doi: 10.1016/j.bbrc.2015.02.138. [DOI] [PubMed] [Google Scholar]
- 11.Lan G, Yang L, Xie X, Peng L, Wang Y. MicroRNA-490-5p is a novel tumor suppressor targeting c-FOS in human bladder cancer. Arch Med Sci. 2015;11:561–569. doi: 10.5114/aoms.2015.52359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Fang Y, Zhu X, Wang J, Li N, Li D, Sakib N, Sha Z, Song W. MiR-744 functions as a protooncogene in nasopharyngeal carcinoma progression and metastasis via transcriptional control of ARHGAP5. Oncotarget. 2015;6:13164–13175. doi: 10.18632/oncotarget.3754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Nho RS, Im J, Ho YY, Hergert P. MicroRNA-96 inhibits FoxO3a function in IPF fibroblasts on type I collagen matrix. Am J Physiol Lung Cell Mol Physiol. 2014;307:L632–642. doi: 10.1152/ajplung.00127.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wang D, Zhang H, Li M, Frid MG, Flockton AR, McKeon BA, Yeager ME, Fini MA, Morrell NW, Pullamsetti SS, Velegala S, Seeger W, McKinsey TA, Sucharov CC, Stenmark KR. MicroRNA-124 controls the proliferative, migratory, and inflammatory phenotype of pulmonary vascular fibroblasts. Circ Res. 2014;114:67–78. doi: 10.1161/CIRCRESAHA.114.301633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Liang WL, Cao J, Xu B, Yang P, Shen F, Sun Z, Li WL, Wang Q, Liu F. miR-892a regulated PPP2R2A expression and promoted cell proliferation of human colorectal cancer cells. Biomed Pharmacother. 2015;72:119–124. doi: 10.1016/j.biopha.2015.04.015. [DOI] [PubMed] [Google Scholar]
- 16.Jiang C, Chen X, Alattar M, Wei J, Liu H. MicroRNAs in tumorigenesis, metastasis, diagnosis and prognosis of gastric cancer. Cancer Gene Ther. 2015;22:291–301. doi: 10.1038/cgt.2015.19. [DOI] [PubMed] [Google Scholar]
- 17.Chen S, Chen X, Xiu YL, Sun KX, Zhao Y. MicroRNA-490-3P targets CDK1 and inhibits ovarian epithelial carcinoma tumorigenesis and progression. Cancer Lett. 2015;362:122–130. doi: 10.1016/j.canlet.2015.03.029. [DOI] [PubMed] [Google Scholar]
- 18.He D, Wang J, Zhang C, Shan B, Deng X, Li B, Zhou Y, Chen W, Hong J, Gao Y, Chen Z, Duan C. Down-regulation of miR-675-5p contributes to tumor progression and development by targeting pro-tumorigenic GPR55 in non-small cell lung cancer. Mol Cancer. 2015;14:73. doi: 10.1186/s12943-015-0342-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Smith AR, Marquez RT, Tsao WC, Pathak S, Roy A, Ping J, Wilkerson B, Lan L, Meng W, Neufeld KL, Sun XF, Xu L. Tumor suppressive microRNA-137 negatively regulates Musashi-1 and colorectal cancer progression. Oncotarget. 2015;6:12558–12573. doi: 10.18632/oncotarget.3726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Shen F, Cai WS, Feng Z, Li JL, Chen JW, Cao J, Xu B. MiR-492 contributes to cell proliferation and cell cycle of human breast cancer cells by suppressing SOX7 expression. Tumour Biol. 2015;36:1913–1921. doi: 10.1007/s13277-014-2794-z. [DOI] [PubMed] [Google Scholar]
- 21.Hui Z, Yiling C, Wenting Y, XuQun H, ChuanYi Z, Hui L. miR-491-5p functions as a tumor suppressor by targeting JMJD2B in ERalpha-positive breast cancer. FEBS Lett. 2015;589:812–821. doi: 10.1016/j.febslet.2015.02.014. [DOI] [PubMed] [Google Scholar]
- 22.Liu Z, Wu R, Li G, Sun P, Xu Q, Liu Z. MiR-592 inhibited cell proliferation of human colorectal cancer cells by suppressing of CCND3 expression. Int J Clin Exp Med. 2015;8:3490–3497. [PMC free article] [PubMed] [Google Scholar]
- 23.Liu M, Zhi Q, Wang W, Zhang Q, Fang T, Ma Q. Up-regulation of miR-592 correlates with tumor progression and poor prognosis in patients with colorectal cancer. Biomed Pharmacother. 2015;69:214–220. doi: 10.1016/j.biopha.2014.12.001. [DOI] [PubMed] [Google Scholar]
- 24.Zhao X, Liu Y, Du L, He L, Ni B, Hu J, Zhu D, Chen Q. Threonine 32 (Thr32) of FoxO3 is critical for TGF-beta-induced apoptosis via Bim in hepatocarcinoma cells. Protein Cell. 2015;6:127–138. doi: 10.1007/s13238-014-0121-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Li J, Li P, Chen T, Gao G, Chen X, Du Y, Zhang R, Yang R, Zhao W, Dun S, Gao F, Zhang G. Expression of microRNA-96 and its potential functions by targeting FOXO3 in non-small cell lung cancer. Tumour Biol. 2015;36:685–692. doi: 10.1007/s13277-014-2698-y. [DOI] [PubMed] [Google Scholar]