

Abstract
Background
Protein kinase A (PKA) is a holoenzyme that consists of a dimer of regulatory subunits and two inactive catalytic subunits that bind to the regulatory subunit dimer. Four regulatory subunits (RIα, RIβ, RIIα, RIIβ) and four catalytic subunits (Cα, Cβ, Cγ, Prkx) have been described in the human and mouse genomes. Previous studies showed that complete inactivation of the Prkar1a subunit (coding for RIα) in the germline leads to embryonic lethality, while Prkar1a–deficient mice are viable and develop schwannomas, thyroid, and bone neoplasms, and rarely lymphomas and sarcomas. Mice with inactivation of the Prkar2a and Prkar2b genes (coding for RIIα and RIIβ, respectively) are also viable but have not been studied for their susceptibility to any tumors.
Methods
Cohorts of Prkar1a+/−, Prkar2a+/−, Prkar2a−/−, Prkar2b+/− and wild type (WT) mice have been observed between 5 and 25 months of age for the development of hematologic malignancies. Tissues were studied by immunohistochemistry; tumor-specific markers were also used as indicated. Cell sorting and protein studies were also performed.
Results
Both Prkar2a−/− and Prkar2a+/− mice frequently developed hematopoietic neoplasms dominated by histiocytic sarcomas (HS) with rare diffuse large B cell lymphomas (DLBCL). Southern blot analysis confirmed that the tumors diagnosed histologically as DLBCL were clonal B cell neoplasms. Mice with other genotypes did not develop a significant number of similar neoplasms.
Conclusions
Prkar2a deficiency predisposes to hematopoietic malignancies in vivo. RIIα’s likely association with HS and DLBCL was hitherto unrecognized and may lead to better understanding of these rare neoplasms.
Keywords: Protein kinase A, Histiocytic sarcoma, Cyclic AMP, Diffuse large B cell lymphoma, Carney complex
Background
Protein Kinase A (PKA) is the main mediator of cyclic adenosine mono-phosphate (cAMP)-dependent signaling [1, 2]. In humans and rodents, PKA is a heterotetramer formed by two regulatory (RIα, RIβ, RIIα, RIIβ) and two catalytic subunits (Cα, Cβ, Cγ, Cx) [1, 3]. The PKA subunits have distinct expression patterns: α-subunits are expressed ubiquitously, whereas β-subunits are expressed in a more tissue-specific fashion. There are two types of PKA holoenzymes, type I and type II, depending on the identity of the regulatory subunit [4]. The cAMP binds to the PKA regulatory subunits (two cAMP molecules per subunit), which leads to dissolution of the tetramer, allowing for the catalytic subunits to act as serine-threonine kinases, phosphorylating numerous targets [1, 2].
Among the regulatory subunits, the PKA regulatory subunit type 1A (PRKAR1A or RIα) has the highest affinity for cAMP and is therefore more sensitive to cAMP, which makes RIα essential for maintaining regulated PKA activity [5]. The PRKAR1A gene on human chromosome 17q22-24 is the gene encoding for RIα; mutations in this gene are responsible for the multiple tumor syndrome Carney complex (CNC, Online Mendelian Inheritance in Man #160980) [6, 7]. CNC patients develop myxomas, skin lesions, schwannomas, bone and endocrine tumors, and a variety of cancers but they are not known to be predisposed to hematologic malignancies. Mice haploinsufficient for a null allele of Prkar1a (Prkar1a+/−) developed tumors, mainly bone tail and thyroid tumors and schwannomas, but the spectrum was somewhat different from that seen in humans [8]. Mice bearing a transgene expressing an anti-sense Prkar1a construct (AS-Prkar1a) developed more tumors than the Prkar1a+/− mice, including adrenal hyperplasia, histiocytic sarcomas (HS) and lymphomas [9]. Taken together, these studies clearly identified PRKAR1A as a tumor suppressor gene affecting several cell types in both humans and mice but left it unclear as to whether PKA defects might lead to a predisposition to hematologic malignancies.
Previous studies showed that Prkar2a knockout (KO) mice are viable, but a predisposition to tumor development has not been reported [10–12]. We studied Prkar2a- haploinsufficient (Prkar2a+/−), Prkar2b- haploinsufficient (Prkar2b+/−), Prkar2a−/− KO, as well as double-heterozygote (Prkar1a+/−x Prkar2a+/-) F1 mice for the development of hematologic malignancies. We found that Prkar2a animals developed a spectrum of B cell lineage-derived and histiocytic hematopoietic neoplasms (as well as lung and liver tumors) with an incidence significantly higher than those previously reported for Prkar1a KO or Prkar1a-haploinsufficient mice.
Methods
Animal protocol
Mice deficient for Prkar1a and Prkar2a alleles that have been described previously [8, 12], were maintained on a mixed genetic background (C57BL/6/129Sv) and were crossed to generate Prkar2a+/− and Prkar2a−/− mice. We also studied Prkar2b+/− mice on the same mixed genetic background. All studies were performed under animal protocol 12–033 and were approved by and conducted in accordance with the Eunice Kennedy Shriver National Institute for Child Health and Human Development Institutional Animal Care and Use Committee. Mice have been observed between the ages of 5 and 25 months and were necropsied when found to have splenomegaly, lymphadenopathy, hepatomegaly, labored breathing, ruffled fur, or other signs of significant morbidity. At necropsy, harvested tissues were fixed in 10 % neutral buffered formalin for histopathologic study and other samples were frozen for subsequent DNA, RNA and protein studies.
Flow Cytometry Analysis (FACS)
Single-cell suspensions were stained with mAb conjugated to FITC, PE, PerCP, APC, or biotin (BD Biosciences) assayed on a FACSCalibur (Becton Dickinson). The data were analyzed with FlowJo (TreeStar Inc) or WinMDI (The Scripps Institute) software. The following antibodies were used: B220-PercP (BD553093), IgM-APC (BD550676), CD43-APC (BD560663), CD24 (HSA)-APC (BD562349), Ly-1 (BP-1)-PE (BD496578), CD3e-PE-Cy™7 (BD552774), CD4-PE (BD553730), CD8-APC (BD553035).
Immunohistochemistry
Formalin fixed, paraffin embedded sections were stained with hematoxylin and eosin (H&E) and antibodies (B220 antibody (dilution 1:50), BD Pharmingen, 550286 and Mac-2 (Galectin-3) antibody (dilution 1:500), Biolegend, 810801). Histologic diagnoses of hematopoietic neoplasms were made using established criteria [13–16].
Southern blot hybridization
DNA prepared from frozen samples of spleen obtained at necropsy was processed for Southern blot hybridization using established procedures and hybridized with an IgH JH probe [15, 17–19].
Results
Tumor development in Prkar1a-, Prkar2a- and Prkar2b-deficient mice
Prkar1a+/−, Prkar2a+/−, Prkar2a−/−, Prkar2b+/− and wild type (WT) mice were necropsied when they developed splenomegaly, lymphadenopathy (at any age) or when moribund between 5 and 25 months of age. Tissues from a total of 31 mice were studied.
Prkar2a-deficienct mice developed hematopoietic tumors with higher frequency than Prkar1a-deficient and WT mice (Fig. 1). Remarkably, a total of 16 cases (51.6 %), including 11/19 cases from the Prkar2a+/− mice (58 %), were diagnosed as histiocytic sarcomas (HS) [11] (Table 1). Seven others cases were tumors of B cell lineage origin, including diffuse large B cell lymphomas (DLBCL), a plasmocytoma and marginal zone lymphomas (MZL) (Table 1) [13, 20–22].
Fig. 1.
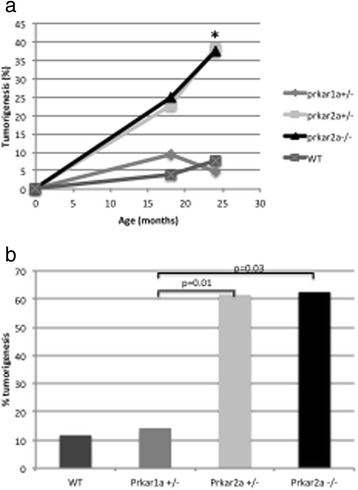
Incidence and time course of tumor development in Prkar1a +/− (n = 28), Prkar2a +/− (n = 21), Prkar2a −/− (n = 5) and wild type (WT) (n = 25) mice. All mice were on a mixed (C57BL/6/129Sv) genetic background. a Total tumor incidence among mice followed for 25 months; * = p < 0.02. b Frequency of tumors among mice necropsied at 9 and 25 months of age
Table 1.
Summary of all tumors developed in the animals during the course of the study
| Mouse ID | Sex | Age | Genotype | Diagnosis | Additional findings |
|---|---|---|---|---|---|
| 3209 | male | 18 mon | Prkar2a +/− | DLBCL and HS rich | |
| 3150 | female | 18 mon | Prkar2a +/− | DLBCL | |
| 3345 | male | 20 mon | Prkar2a +/− | MZL | |
| 3485 | male | 22 mon | Prkar2a +/− | HS | |
| 2378 | male | 18 mon | Prkar2a +/− | HS | Lung adenoma |
| 2379 | male | 18 mon | Prkar2a +/− | HS | |
| 2552 | female | 25 mon | Prkar2a +/− | HS | |
| 3078 | female | 23 mon | Prkar2a +/− | HS | |
| 3305 | female | 24 mon | Prkar2a +/− | Liver, Lung, Pancreas: lymphoid infiltrates | Liver: lymphoplasmatic infiltrates |
| 3210 | male | 24 mon | Prkar2a +/− | HS | |
| 3211 | male | 24 mon | Prkar2a +/− | Liver hemangiosarcoma | |
| 3217 | male | 16 mon | Prkar2a +/− | DLBCL and HS | |
| 2622 | male | 17 mon | Prkar2a +/− | HS | |
| MS1200161 | male | 16 mon | Prkar2a +/− | HS | |
| 3306 | female | 19 mon | Prkar2a +/− | HS | |
| 2494 | female | 18 mon | Prkar2a +/− | HS | |
| 3794 | male | 6 mon | Prkar2a +/− | Spleen Lymphoid Hyperplasia | |
| 2545 | male | 20 mon | Prkar2a +/− | Plasmacytoma | |
| 3753 | female | 6 mon | Prkar2a +/− | Spleen Lymphoid Hyperplasia | |
| 4071 | female | 18 mon | Prkar2a −/− | HS | |
| 4072 | female | 18 mon | Prkar2a −/− | HS | |
| 2537 | female | 18 mon | Prkar2a −/− | DLBCL and HS rich | |
| 3988 | female | 15 mon | Prkar1a +/−; Prkar2a +/− | Liver - Early adenoma | |
| 2383 | female | 24 mon | WT | HS | |
| 3146 | female | 18 mon | WT | EMZ | |
| 3852 | male | 17 mon | WT | MZL | |
| 3437 | male | 18 mon | Prkar2b +/− | Lymph nodes: plasma cell tumor | |
| 3436 | female | 18 mon | Prkar2b +/− | Lung: adenoma | Spleen plasma cells |
| 3147 | female | 18 mon | Prkar2b +/− | EMZ | |
| 2860 | female | 25 mon | Prkar2b +/− | HS | |
| 1941 | female | 16 mon | Prkar2b +/− | DLBCL | |
| 2204 | female | 24 mon | Prkar2b +/− | HS | |
| 2939 | female | 24 mon | Prkar1a +/− | MZL | Lymph nodes plasmacytosis |
| 3069 | male | 24 mon | Prkar1a +/− | Lung: fibrosarcoma; adenoma | |
| 3087 | male | 22 mon | Prkar1a +/− | HS | |
| 1354 | male | 23 mon | Prkar1a +/− | Early HS | Trachea lymphoplasmacytic |
HS Histiocytic sarcoma, DLBCL Diffuse large B cell lymphoma, EMZ Expanded marginal zone, MZL Marginal zone lymphoma
Overall, the most frequent neoplasms were hematopoietic in origin and included diffuse large B cell lymphomas (DLBCL) with 3 of 4 of these cases (Table 1, mice 3209, 3217, 2537, 3150) being histiocyte-rich. In one of these cases, the mouse appeared to have a coexisting DLBCL and a HS (Fig. 2, Table 1 mouse #3217). The spleen of this mouse was infiltrated by DLBCL that stained intensely with antibody to the B cell marker, B220 (Fig. 2a) and had metastasized to the liver. The liver mass stained uniformly with B220 (Fig. 2b) but was negative for the macrophage marker, Mac-2 (data not shown). In another part, the same spleen was essentially replaced with a HS associated with the presence of large multinucleate cells (Fig. 2c) that are often seen in mouse as well as human HS [23].
Fig. 2.
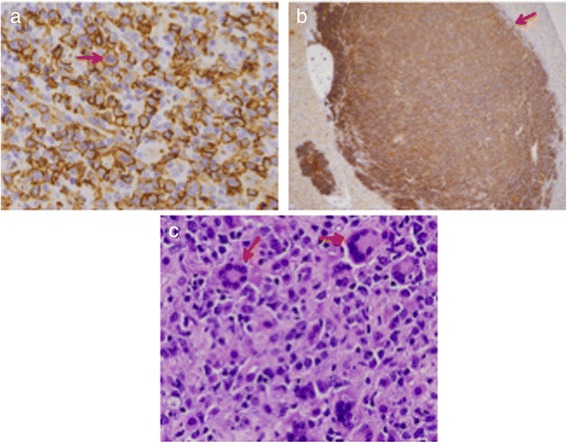
Histopathology and immunohistochemistry of coexisting DLBCL and HS. Sections of spleen (magnification 40x) (a) and liver (magnification 4x) (b) from the same mouse were stained with antibody to B220 (red arrows). a section from another portion of the same spleen stained with H&E reveals features consistent with the diagnosis of HS including many multinucleate giant cells (red arrows) (magnification 40x) (c)
Other mice developed malignancies of plasma cells involving the spleen and lymph nodes (Table 1, mice #2545, 3437, 3436, 2939, 3305, 1354). One case (#3437) had sheets of easily recognizable fully mature plasma cells with few mitotic figures (Fig. 3a). A second case (#2545) had numerous plasmablasts with round nuclei containing a large central nucleolus and thick nuclear membranes (Fig. 3b).
Fig. 3.
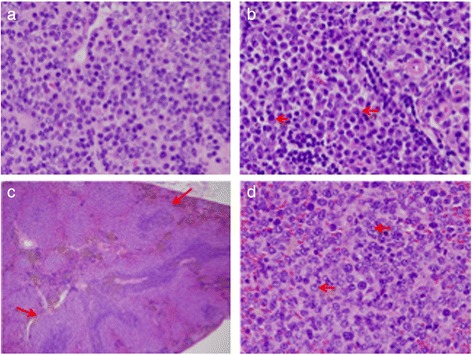
Plasma cell tumors and high-grade splenic marginal zone lymphoma. a, b Lymph node sections from two mice (from different litters) stained with H&E (magnification 40x). a. Low-grade tumor with mature plasma cells. b Higher grade tumor dominated by plasmablasts (red arrows) (c) Low power view of a spleen stained with H&E showing small follicles surrounded by multiple layers of pale-staining marginal zone cells (red arrows) (magnification 4x). d Cells in the marginal zone have acquired centroblastic cytology (red arrows) (magnification 40x)
We identified a case of high-grade splenic marginal zone lymphoma [24] (Table 1, mouse #3345). At low power, sheets of pale staining cells can be seen, surrounding the follicles and invading both the follicles and the red pulp (Fig. 3c). At high power, the marginal zone is seen to be populated by centroblasts with one or more prominent nucleoli at the nuclear membrane and some immunoblasts with a large magenta bar-shaped nucleolus appended to one side of the nuclear membrane (Fig. 3d).
DNA studies
To determine if the B cell lineage tumors were clonal, we performed DNA extraction from 4 cases of DLBCL (Table 1, mice # 3217, 3150, 3209, 2537), 1 HS (Table 1, mouse #2622) and a normal spleen and analyzed them by Southern blot hybridization for organization of the IgH locus (Fig. 4). All the DLBCL cases exhibited clonal or oligoclonal rearrangements of IgH while the HS and normal spleen, as expected, had only a 6.8 kb band characteristic of the germline sequence. It is noteworthy that the intensity of the germline band was reduced in only one of the cases of DLBCL indicating that the tumors were not the dominant cell types in the other cases. We concluded from these studies, that the tumors diagnosed histologically as DLBCL were clonal B cell neoplasms.
Fig. 4.
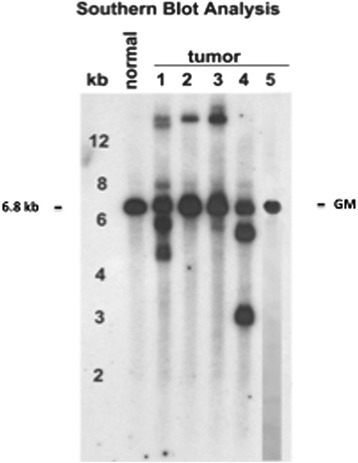
Southern blot hybridization of DNA prepared from 4 DLBCL (Lanes 1–4), a HS (Lane 5) and a normal spleen hybridized with a JH probe. Germline band of 6.8 kb is indicated
Cell sorting
To determine if altered PRKAR2A expression might affect early events in hematopoietic differentiation in the BM or the development and distribution of cell subsets in peripheral lymphoid tissues, we performed extensive multiparameter FACS analyses of single cell suspensions prepared from bone marrow (BM), spleen, lymph nodes and thymi from 3–6 month old Prkar2a+/− and Prkar2a−/− mice. The analyses were for B cells, T cells and their subsets, macrophages, dendritic cells and granulocytes. We found no significant changes in any cell subset in any tissue from data obtained with WT mice (data not shown). These results indicate that if aberrant PRKAR2A expression affected hematopoietic differentiation, the effects only became evident well into adult life.
Non-hematopoietic tumors
We identified mice with tumors of the lung (Table 1, mice # 2378, 3436, 3069). These included a pulmonary adenoma (Fig. 5a, b mouse #2378) and a pulmonary fibrosarcoma [21] (Fig. 5c, mouse #3069). The adenoma, which presented as a round mass lesion, was comprised of monomorphic well-differentiated cells with complete loss of normal alveolar architecture. In contrast, the fibrosarcoma presented as a large mass lesion displacing all normal lung elements and was associated with a large necrotic area. It is not known if the lung was where this tumor originated from, but sarcomatous lesions were not identified in other tissues of this mouse.
Fig. 5.
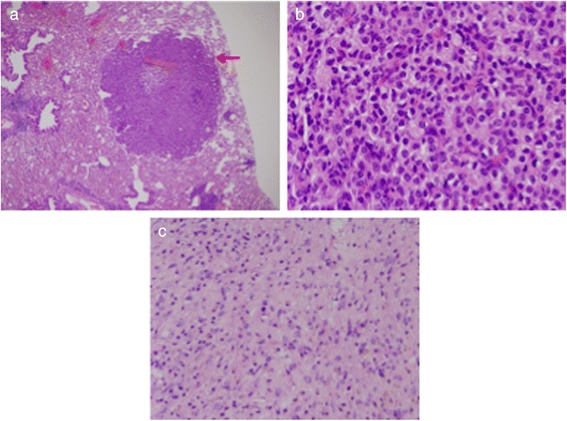
Lung tumors. Pulmonary adenoma (a), (magnification 4x) and same pulmonary adenoma at higher magnification (b) (magnification 40x). c fibrosarcoma (magnification 20x) (c)
One mouse (#3211) developed an uncommon liver vascular lesion readily evident at necropsy (Fig. 6a). Histologic sections of liver revealed features consistent with a diagnosis of malignant hemangiosarcoma but that also resembled peliosis hepatis, a non-malignant lesion in which multiple blood filled cavities can be found throughout the liver [25] (Fig. 6b, c).
Fig. 6.
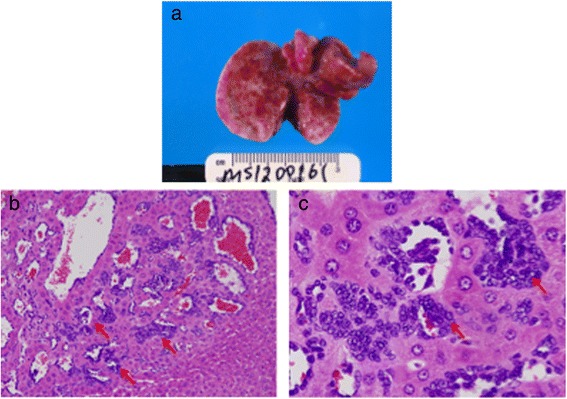
Gross and microscopic presentation of a hepatic hemangiosarcoma. a Multiple liver hemangiomas were evident at necropsy. b, c Liver sections showing small vascular lesions (red arrows) (magnification 10x) (b) associated with malignant endothelial cells (red arrows) (magnification 40x) (c)
Discussion
Our findings in this study identify Pkrar2a as a gene whose deficiency predisposes mice to a spectrum of mostly hematopoietic neoplasms. This phenotype has some similarities but also differences from the spectrum of tumors exhibited by humans and mice with RIα defects [6, 9, 26]. The major difference is that Prkar2a+/− and Prkar2a−/− mice develop with a high frequency hematopoietic neoplasms, particularly macrophage-derived HS tumors, as well as neoplasms derived from mature cells of the B cell lineage. Lymphomas or HS have not been reported in patients with CNC but were among the tumors found relatively rarely in Prkar1a-deficient mice [8, 9]. This suggests that alterations in the RIα and RIIα signaling in different cell types may preferentially predispose them to transformation. It has been shown, for example, that PRKAR1A inactivation promotes the proliferation of human B cells in association with reduced apoptosis [27], both of which may enhance B cell sensitivity to malignant transformation.
HS is an uncommon lesion in humans and mice and the etiology in either species remains unknown [16, 23]. Studies of Prkar2a-deficient mice revealed that many animals without clear histologic evidence of HS exhibited histiocytic proliferation in association with DLBCL, a condition we refer to as histiocyte-rich DLBCL [13, 28] or, isolated, in the absence of other tumors. It is not clear if such proliferation might be the precursor to tumor development. However, such progression appears possible since in a few animals distinct areas of the spleen were occupied by HS with other areas showing classic features of histiocyte-rich DLBCL. Rarely, HS have been found to harbor IgH or TCR rearrangements, likely as the result of trans-differentiation [29], but this was not found to be the case with the single HS tested here for IgH organization.
It remains to be determined how dysregulation of PRKAR2A expression contributes to transformation of these cells. There is a report of a single in-frame expressed fusion gene resulting from a chromosomal translocation in breast cancer between PRKAR2A and SLC26A6, which encodes an anion transporter [30], but the oncogenetic significance of this event has not been determined. A comprehensive review of the cancer genome atlas (TCGA) did not reveal PRKAR2A mutations in any human tumors but several studies show copy number changes of PRKAR2A’s chromosomal locus at 3p21 which, however, is gene-rich and is frequently lost in a variety of neoplasms [31–35].
In conclusion, the present study supports the notion that alterations in PKA signaling in a variety of cell types, including those of the hematologic lineage, may lead to neoplastic transformation. Prkar2a- deficiency, in particular, predisposes mice to a spectrum of mostly hematopoietic neoplasms, which may lead to better understanding of the molecular pathology of these lesions in both mice and humans.
Acknowledgements
This work was supported by the Intramural Research Program (IRP) of the Eunice Kennedy Shriver National Institute of Child Health & Human Development (NICHD),and the National Institute of Allergy and Infectious Diseases (HCM, CQI), National Institutes of Health (NIH), Bethesda, MD 20892 and by a research grant from Hellenic Society of Medical Oncology (awardee: Emmanouil Saloustros).
Abbreviations
- PKA
Protein Kinase A
- WT
Wild Type
- DLBCL
Diffuse Large B Cell Lymphoma
- CNC
Carney Complex
- HS
Histiocytic Sarcoma
- KO
Knockout
- H&E
Hematoxylin and Eosin
- MZL
Marginal Zone Lymphoma
- HR
Histiocytic Rich
- FACS
Fluorescence Activated Cell Sorting
- BM
Bone Marrow
- TCR
T Cell Receptor
Footnotes
Emmanouil Saloustros and Paraskevi Salpea contributed equally to this work.
Competing interests
None of the authors have any competing interests in this manuscript.
Author’s contributions
Drs. E. Saloustros and P. Salpea have contributed equally to the work described and the writing of this manuscript and they share first authorship.
Drs. Constantine A. Stratakis and Herbert C. Morse III share senior authorship.
Contributor Information
Emmanouil Saloustros, Email: esaloustros@yahoo.gr.
Paraskevi Salpea, Email: salpeapa@mail.nih.gov.
Chen-Feng Qi, Email: cqi@niaid.nih.gov.
Lina A. Gugliotti, Email: laguglio@gmail.com
Kitman Tsang, Email: kitman1214@gmail.com.
Sisi Liu, Email: sisi.liu@nih.gov.
Matthew F. Starost, Email: starostm@ors.od.nih.gov
Herbert C. Morse, III, Phone: 001-301-761-5006, Email: hmorse@niaid.nih.gov.
Constantine A. Stratakis, Phone: 001-301-496-4686, Email: stratakc@mail.nih.gov
References
- 1.Skalhegg BS, Tasken K. Specificity in the cAMP/PKA signaling pathway. Differential expression, regulation, and subcellular localization of subunits of PKA. Front Bioscience. 2000;5:D678–693. doi: 10.2741/Skalhegg. [DOI] [PubMed] [Google Scholar]
- 2.Das R, Esposito V, Abu-Abed M, Anand GS, Taylor SS, Melacini G. cAMP activation of PKA defines an ancient signaling mechanism. Proc Natl Acad Sci U S A. 2007;104:93–8. doi: 10.1073/pnas.0609033103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Gamm DM, Baude EJ, Uhler MD. The major catalytic subunit isoforms of cAMP-dependent protein kinase have distinct biochemical properties in vitro and in vivo. J Biol Chem. 1996;271:15736–15742. doi: 10.1074/jbc.271.26.15736. [DOI] [PubMed] [Google Scholar]
- 4.Di Benedetto G, Zoccarato A, Lissandron V, Terrin A, Li X, Houslay MD, et al. Protein kinase A type I and type II define distinct intracellular signaling compartments. Circ Res. 2008;103:836–44. doi: 10.1161/CIRCRESAHA.108.174813. [DOI] [PubMed] [Google Scholar]
- 5.Amieux PS, McKnight GS. The essential role of RI alpha in the maintenance of regulated PKA activity. Ann N Y Acad Sci. 2002;968:75–95. doi: 10.1111/j.1749-6632.2002.tb04328.x. [DOI] [PubMed] [Google Scholar]
- 6.Kirschner LS, Carney JA, Pack SD, Taymans SE, Giatzakis C, Cho YS, et al. Mutations of the gene encoding the protein kinase A type I-alpha regulatory subunit in patients with the Carney complex. Nat Genet. 2000;26:89–92. doi: 10.1038/79238. [DOI] [PubMed] [Google Scholar]
- 7.Carney JA, Boccon-Gibod L, Jarka DE, Tanaka Y, Swee RG, Unni KK, et al. Osteochondromyxoma of bone: a congenital tumor associated with lentigines and other unusual disorders. Am J Surg Pathol. 2001;25:164–76. doi: 10.1097/00000478-200102000-00004. [DOI] [PubMed] [Google Scholar]
- 8.Kirschner LS, Kusewitt DF, Matyakhina L, Towns WH, 2nd, Carney JA, Westphal H, et al. A mouse model for the Carney complex tumor syndrome develops neoplasia in cyclic AMP-responsive tissues. Cancer Res. 2005;65:4506–14. doi: 10.1158/0008-5472.CAN-05-0580. [DOI] [PubMed] [Google Scholar]
- 9.Griffin KJ, Kirschner LS, Matyakhina L, Towns WH, 2nd, Carney JA, Westphal H, et al. Down-regulation of regulatory subunit type 1A of protein kinase A leads to endocrine and other tumors. Cancer Res. 2004;64:8811–5. doi: 10.1158/0008-5472.CAN-04-3620. [DOI] [PubMed] [Google Scholar]
- 10.Scott JD, Glaccum MB, Zoller MJ, Uhler MD, Helfman DM, McKnight GS, et al. The molecular cloning of a type II regulatory subunit of the cAMP-dependent protein kinase from rat skeletal muscle and mouse brain. Proc Natl Acad Sci U S A. 1987;84:5192–6. doi: 10.1073/pnas.84.15.5192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Burton KA, Treash-Osio B, Muller CH, Dunphy EL, McKnight GS. Deletion of type IIalpha regulatory subunit delocalizes protein kinase A in mouse sperm without affecting motility or fertilization. J Biol Chem. 1999;274:24131–6. doi: 10.1074/jbc.274.34.24131. [DOI] [PubMed] [Google Scholar]
- 12.Burton KA, Johnson BD, Hausken ZE, Westenbroek RE, Idzerda RL, Scheuer T, et al. Type II regulatory subunits are not required for the anchoring-dependent modulation of Ca2+ channel activity by cAMP-dependent protein kinase. Proc Natl Acad Sci U S A. 1997;94:11067–72. doi: 10.1073/pnas.94.20.11067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Morse HC, 3rd, Anver MR, Fredrickson TN, Haines DC, Harris AW, Harris NL, et al. Bethesda proposals for classification of lymphoid neoplasms in mice. Blood. 2002;100:246–58. doi: 10.1182/blood.V100.1.246. [DOI] [PubMed] [Google Scholar]
- 14.Kogan SC, Ward JM, Anver MR, Berman JJ, Brayton C, Cardiff RD, et al. Bethesda proposals for classification of nonlymphoid hematopoietic neoplasms in mice. Blood. 2002;100:238–45. doi: 10.1182/blood.V100.1.238. [DOI] [PubMed] [Google Scholar]
- 15.Hartley JW, Chattopadhyay SK, Lander MR, Taddesse-Heath L, Naghashfar Z, Morse HC, 3rd, et al. Accelerated appearance of multiple B cell lymphoma types in NFS/N mice congenic for ecotropic murine leukemia viruses. Lab Invest. 2000;80:159–69. doi: 10.1038/labinvest.3780020. [DOI] [PubMed] [Google Scholar]
- 16.Hao X, Fredrickson TN, Chattopadhyay SK, Han W, Qi CF, Wang Z, et al. The histopathologic and molecular basis for the diagnosis of histiocytic sarcoma and histiocyte-associated lymphoma of mice. Vet Pathol. 2010;47:434–45. doi: 10.1177/0300985810363705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Qi CF, Zhou JX, Lee CH, Naghashfar Z, Xiang S, Kovalchuk AL, et al. Anaplastic, plasmablastic, and plasmacytic plasmacytomas of mice: relationships to human plasma cell neoplasms and late-stage differentiation of normal B cells. Cancer Res. 2007;67:2439–47. doi: 10.1158/0008-5472.CAN-06-1561. [DOI] [PubMed] [Google Scholar]
- 18.Butenko ZA, Smirnova IA, Zak KP, Kishinskaja EG, Janok EA. Leukemia-associated gene rearrangements in blood mononuclears of subjects in long terms after radiation exposure. J Exp Clin Cancer Res. 2000;19:57–9. [PubMed] [Google Scholar]
- 19.Moriyama H, Kawahara K, Noguchi T, Kikuchi R, Wada S, Takeno S, et al. Primary extramedullary plasmacytoma of the small intestine. A case report and review of the literature. J Exp Clin Cancer Res. 2006;25:129–34. [PubMed] [Google Scholar]
- 20.Ward JM, Rehg JE, Morse HC., 3rd Differentiation of rodent immune and hematopoietic system reactive lesions from neoplasias. Toxicol Pathol. 2012;40:425–434. doi: 10.1177/0192623311431467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Algarra I, Collado A, Garrido F. Protein bound polysaccharide PSK abrogates more efficiently experimental metastases derived from H-2 negative than from H-2 positive fibrosarcoma tumor clones. J Exp Clin Cancer Res. 1997;16:373–380. [PubMed] [Google Scholar]
- 22.Pisani F, Romano A, Anticoli Borza P, Marino M, Micheli A, Botti C, et al. Diffuse large B-cell lymphoma involving the breast. A report of four cases. J Exp Clin Cancer Res. 2006;25:277–81. [PubMed] [Google Scholar]
- 23.Campo E, Swerdlow SH, Harris NL, Pileri S, Stein H, Jaffe ES. The 2008 WHO classification of lymphoid neoplasms and beyond: evolving concepts and practical applications. Blood. 2011;117:5019–32. doi: 10.1182/blood-2011-01-293050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Fredrickson TN, Lennert K, Chattopadhyay SK, Morse HC, 3rd, Hartley JW. Splenic marginal zone lymphomas of mice. Am J Pathol. 1999;154:805–12. doi: 10.1016/S0002-9440(10)65327-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Yanoff M, Rawson AJ. Peliosis Hepatis. An Anatomic Study with Demonstration of Two Varieties. Arch Pathol. 1964;77:159–165. [PubMed] [Google Scholar]
- 26.Veugelers M, Wilkes D, Burton K, McDermott DA, Song Y, Goldstein MM, et al. Comparative PRKAR1A genotype-phenotype analyses in humans with Carney complex and prkar1a haploinsufficient mice. Proc Natl Acad Sci U S A. 2004;101:14222–7. doi: 10.1073/pnas.0405535101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Robinson-White AJ, Leitner WW, Aleem E, Kaldis P, Bossis I, Stratakis CA. PRKAR1A inactivation leads to increased proliferation and decreased apoptosis in human B lymphocytes. Cancer Res. 2006;66:10603–12. doi: 10.1158/0008-5472.CAN-06-2200. [DOI] [PubMed] [Google Scholar]
- 28.Terada Y, Nakamae H, Aimoto R, Kanashima H, Sakamoto E, Aimoto M, et al. Impact of relative dose intensity (RDI) in CHOP combined with rituximab (R-CHOP) on survival in diffuse large B-cell lymphoma. J Exp Clin Cancer Res. 2009;28:116. doi: 10.1186/1756-9966-28-116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Hanson CA, Jaszcz W, Kersey JH, Astorga MG, Peterson BA, Gajl-Peczalska KJ, et al. True histiocytic lymphoma: histopathologic, immunophenotypic and genotypic analysis. Br J Haematol. 1989;73:187–98. doi: 10.1111/j.1365-2141.1989.tb00251.x. [DOI] [PubMed] [Google Scholar]
- 30.Stephens PJ, McBride DJ, Lin ML, Varela I, Pleasance ED, Simpson JT, et al. Complex landscapes of somatic rearrangement in human breast cancer genomes. Nature. 2009;462:1005–10. doi: 10.1038/nature08645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Xiong W, Zeng ZY, Xia JH, Xia K, Shen SR, Li XL, et al. A susceptibility locus at chromosome 3p21 linked to familial nasopharyngeal carcinoma. Cancer Res. 2004;64:1972–4. doi: 10.1158/0008-5472.CAN-03-3253. [DOI] [PubMed] [Google Scholar]
- 32.Naylor SL, Marshall A, Hensel C, Martinez PF, Holley B, Sakaguchi AY. The DNF15S2 locus at 3p21 is transcribed in normal lung and small cell lung cancer. Genomics. 1989;4:355–61. doi: 10.1016/0888-7543(89)90342-X. [DOI] [PubMed] [Google Scholar]
- 33.Zeng Z, Zhou Y, Zhang W, Li X, Xiong W, Liu H, et al. Family-based association analysis validates chromosome 3p21 as a putative nasopharyngeal carcinoma susceptibility locus. Genet Med. 2006;8:156–60. doi: 10.1097/01.gim.0000196821.87655.d0. [DOI] [PubMed] [Google Scholar]
- 34.Bokemeyer A, Eckert C, Meyr F, Koerner G, von Stackelberg A, Ullmann R, et al. Copy number genome alterations are associated with treatment response and outcome in relapsed childhood ETV6/RUNX1-positive acute lymphoblastic leukemia. Haematologica. 2014;99:706–14. doi: 10.3324/haematol.2012.072470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kreisel F, Kulkarni S, Kerns RT, Hassan A, Deshmukh H, Nagarajan R, et al. High resolution array comparative genomic hybridization identifies copy number alterations in diffuse large B-cell lymphoma that predict response to immuno-chemotherapy. Cancer Genet. 2011;204:129–37. doi: 10.1016/j.cancergen.2010.12.010. [DOI] [PMC free article] [PubMed] [Google Scholar]