

Abstract
Regulation of appropriate osmolality of body fluid is critical for survival, yet there are sex differences in compensatory responses to osmotic challenges. Few studies have focused on the role of sex hormones such as estradiol in behavioral responses to increases or decreases in systemic osmolality, and even fewer studies have investigated whether central actions of estrogens contribute to these responses. This overview integrates findings from a series of ongoing and completed experiments conducted in my laboratory to assess estradiol effects on water and NaCl intake in response to osmotic challenges, and on activity in central pathways that mediate such responses.
Body fluid osmolality is tightly regulated, with an array of integrated mechanisms targeted toward maintaining osmolality at levels optimal for physiological function. Although much research focuses on hormonal and renal mechanisms that support osmotic regulation [1, 4–5, 18–19, 38, 50, 54], behaviors such as sodium ingestion also play a critical role. Sodium is the primary determinant of plasma osmolality, and the importance of sodium intake in osmotic regulation is readily apparent in the ‘salt blocks’ that frequently are provided to domestic animals. In humans, however, the ingestion of pure salt is less obvious; certainly, with the exception of disease states such as the classic case described by Wilkins and Richter in 1940 [56] it seems to be less common. This discrepancy may be attributable to the amount of sodium present in our diets and, in particular, in processed food. Thus, the need to seek out and consume sodium in order to maintain optimum body fluid osmolality is less of an issue for humans. In fact, dietary sodium is not only sufficient, but typically in excess of need [55, 59]. This excess sodium consumption, and the potential for negative health consequences, raises the question of whether sodium salts are consumed because of a preference for the taste, rather than to regulate body fluid osmolality. This question has been the subject of much debate (e.g., [3, 24]), with some suggesting that evolutionary underpinnings explain the innate human preference for sodium salts. The opposing viewpoint holds that salt taste preferences in human and animal models are labile and subject to factors ranging from prenatal influences to adult experiences.
One factor that has received scant attention in studies of salt taste preferences is sex differences, though some evidence [17, 22, 53] suggests that women exhibit a preference for salty foods that is related to the stage of their reproductive cycle or to reproductive hormones, more generally. This preference also has been reported in animal models [8, 21], with some evidence for hormonally-mediated changes in taste processing of salt taste—either centrally [15–16] or peripherally [11]—as the underlying mechanism. Indeed, our work showed that chorda tympani nerve responses to a concentrated NaCl solution in intact male rats, and in ovariectomized rats with or without estradiol (Figure 1; [11]) are inversely related to NaCl intake (Figure 2, left; [12]). Taken together, these findings suggest that estradiol treatment results in less intense perception of NaCl taste and, thereby, in greater NaCl intake.
Figure 1.

Chorda tympani responses to concentrated NaCl solution in male rats (hatched bar), oil vehicle (OIL) treated ovariectomized rats (OVX; white bar), and estradiol benzoate (EB) treated OVX rats (black bar). c = significantly greater than EB; for male, p<0.01, for OIL, p<0.05 Adapted from Curtis and Contreras; Behavioral Neuroscience, 2006
Figure 2.

Intake of concentrated NaCl solution (left) and of water (right) by male rats (hatched bars), OIL-treated OVX rats (white bars), and EB-treated OVX rats (black bars). a = significantly greater than males (p<0.05)
Adapted from Curtis et al., Physiology and Behavior, 2004
While intuitively appealing, this explanation overlooks the osmoregulatory consequences of increased NaCl intake. In other words, do females maintain sodium balance despite the ‘self-administered salt load’ resulting from increased NaCl intake that follows changes in salt taste preferences? We addressed this question by simply assessing concurrent water intake and found that the amount of water ingested by estradiol-treated ovariectomized rats when NaCl also is consumed water (Figure 2; [12]) is insufficient for osmotic regulation.
Our laboratory has been following up on these studies to more fully examine the influence of estradiol on behavioral mechanisms of osmoregulation. This overview integrates findings from a series of published and ongoing experiments designed to assess the effects of estradiol on water and NaCl intake in response to osmotic challenges, as well as on activity in central pathways that mediate such responses. All the procedures described were approved by the Oklahoma State Institutional Care and Use Committee and were conducted in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
In these experiments, we use adult female Sprague-Dawley rats that are ovariectomized and, after 7–10 days of recovery, are injected with estradiol benzoate (EB; 10 ug/0.1 ml oil, s.c.) or the oil vehicle (OIL; 0.1 ml, s.c.) on two consecutive days. Water and/or NaCl intake were evaluated using the 4-day hormone treatment-testing protocol shown in Figure 3. We and others have used this protocol in studies assessing behavioral effects of EB related to body fluid balance in rats [25, 32–35, 57]. The primary advantages are that it produces fluctuations in estradiol levels (vs. chronic elevations with methods such as daily injections or implantation of estradiol-filled silastic capsules), while allowing us to control the timing of the fluctuations and to examine the effects of estradiol independent of those attributable to progesterone. We have taken a three-pronged approach to verify the effectiveness of EB treatment using this protocol. In all studies, we weigh rats over the course of the experiments and reliably observe an EB-mediated decrease in body weight (Figure 4a), as expected [23, 25, 35]. In addition, uterine weights assessed at the time of sacrifice on day 4 (Figure 4b) showed the expected hypertrophy [9, 25, 36]. Finally, in a subset of rats, circulating estradiol was measured using a commercially available ELISA kit (Calbiotech #ES180s-100), and showed the expected pattern [58] over the course of the treatment-testing protocol (Figure 4c).
Figure 3.
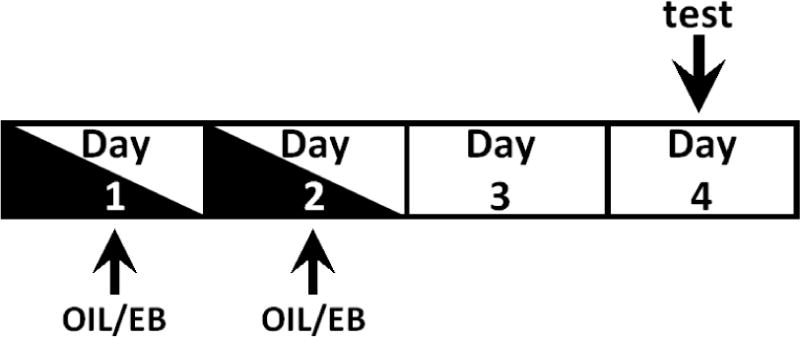
Diagram illustrating hormone treatment-testing protocol. OIL or EB injections were given on day 1 and day 2; testing was conducted on day 4.
Figure 4.

Assessment of the effectiveness of EB treatment.
(A) T-tests of the change in body weight from day 1 to day 4 revealed that OIL-treated gained more weight than did EB-treated rats, which lost weight during the protocol.
(B) T-tests showed that uterine weights in EB-treated rats were greater than those in OIL-treated rats.
(C) One-way ANOVA of plasma estradiol levels revealed a significant difference (F2,17 = 61.34, p<0.001) among the groups. Pairwise comparisons revealed that, 24 hours after the first EB injection (i.e., on the second day of the protocol; EB-d2), circulating estradiol levels were greater than those on the fourth day (EB-d4), as well as those in OIL-treated OVX rats. Estradiol levels in the EB-d4 group remained elevated and were greater than those in OIL-treated rats.
** = significantly greater than EB-d4, OIL (ps<0.001); * = significantly greater than OIL (p<0.05)
Estradiol, water intake, and hyperosmolality
In one series of studies, we assessed water intake in response to hyperosmolality produced by systemic administration of a concentrated NaCl solution. Previous studies, including our own, showed no effect of estradiol on water intake stimulated by hypertonic NaCl treatment [6, 33, 51]; however, those studies used large bolus injections that produced rapid and pronounced hypernatremia [33]. It is known that estradiol sensitizes vasopressin release in response to hypernatremia [1, 26, 44–45]. Thus, it seemed possible that a more subtle effect of estradiol on behavioral responses to hypernatremia might be revealed with a gradual increase in plasma osmolality. Accordingly, in these studies we infused a hypertonic NaCl solution (HS) via indwelling femoral venous catheters to produce a slower, more modest increase in plasma osmolality. The gradual increase in plasma osmolality also facilitated detailed assessment of the HS-stimulated water intake, including latency to begin drinking water, duration of the first water drinking bout, and water intake during the first bout.
As shown in Figure 5 (top), EB treatment substantially reduced the latency to begin drinking in response to slow intravenous (i.v.) HS infusion [31]. In fact, EB-treated OVX rats began drinking water in approximately half the time required by OIL-treated OVX rats. Surprisingly, neither the duration of the first water drinking bout (Figure 5, bottom left) nor the water intake during the first bout (Figure 5, bottom right) was affected by EB treatment. These findings suggest that EB enhances the sensitivity to increased plasma osmolality/plasma sodium, but this idea rests on the assumption that plasma osmolality in OIL- and EB-treated rats had increased comparably at the onset of water drinking by EB-treated rats. Subsequent studies bore out this assumption: at the time point at which EB-treated rats had been observed to drink water in response to i.v. HS infusion, plasma osmolality was comparable in EB- and OIL-treated rats (310.8 + 1.6 vs. 309.1 + 1.4 mOsm/L, respectively; [31]). However, more than 15 additional minutes of HS infusion elapsed before OIL-treated rats began to drink.
Figure 5.

Latency to begin water intake (top), duration of first water intake bout (bottom left) and water consumed during first bout (bottom right) during i.v. hypertonic saline (HS) infusion by OIL-treated rats (white bars) or EB-treated rats (black bars). Symbols indicate values from individual rats; T-test revealed that EB-treated rats began to drink water in significantly less time (p<0.05) than did OIL-treated rats, whereas no differences were observed in duration of first bout or water consumed then.
From Jones and Curtis, Physiology and Behavior, 2009
Thus, the more rapid onset of water drinking by EB-treated OVX rats in response to increased plasma osmolality was not attributable to differences in renal handling of the systemic salt load. Nor did EB seem to influence how accurately the induced hyperosmolality was detected, as evidenced by the bout analyses and examination of total water intake. Rather, it seemed likely that estradiol effects on water intake stimulated by i.v. HS infusion reflected differences in the sensitivity of the central nervous system (CNS) to the induced hyperosmolality. We therefore used immunohistochemical methods to evaluate HS-mediated activation in CNS areas known to play a role in osmoregulation via detection of changes in plasma sodium, or in downstream processing of signals related to such changes.
Assessment of fos immunolabeling as a marker of neuronal activation in the CNS in response to slow, modest increases in plasma osmolality via i.v. HS infusion revealed selective EB effects [30]. More specifically, EB treatment reduced HS-induced neuronal activation in the hindbrain circumventricular organ, area postrema (AP), and in the preautonomic area of the forebrain paraventricular nucleus (PVN). Neuronal activation also was blunted in the sympathoexcitatory area in the rostral ventrolateral medulla (RVLM) of EB-treated rats, and co-localization of the fos protein with dopamine-β-hydroxylase, a marker of norepinephrine neurons, suggested that the effect of EB on activity in the RVLM is general, rather than specific to catecholaminergic neurons. In contrast, we saw no effect of EB on HS-induced neuronal activity in the supraoptic nucleus of the hypothalamus (SON), whereas i.v. HS infusion was ineffective to increase activity in other CNS areas examined, including the forebrain circumventricular organs subfornical organ (SFO) and organum vasculosum of the lamina terminalis (OVLT), the sympathoinhibitory area in the caudal ventrolateral medulla (CVLM), and the nucleus of the solitary tract (NTS), the terminal site of vagal afferents.
As summarized in Figure 6, EB effects on neuronal activation in response to i.v. HS infusion were selective to a hindbrain area important for detecting changes in plasma osmolality (AP; [20, 37]), and to areas important in the control of sympathetic nerve activity (PVN, RVLM; [46, 58]). These areas share reciprocal connections [10, 14, 41–42, 49], so decreased activity in one may reduce the drive on others, and experiments to address this possibility are ongoing. In the short term, however, the answer to the question of whether estradiol influences behavioral responses and activity in CNS pathways related to hyperosmolality seems to be yes: despite comparable hyperosmolality, EB results in more rapid onset of water intake after i.v. HS infusion [31], suggesting that the CNS is more sensitive to hyperosmolality. On the surface, then, the observed decrease in neuronal activation [30] may seem to be surprising; however, our observations that the effects are selective to areas associated with sympathetic nerve activity are potentially instructive. It is known that hyperosmolality increases blood pressure by activating the sympathetic nervous system [2, 27, 47] and that increased blood pressure blunts stimulated water intake [48, 52]. Therefore, if EB treatment attenuates sympathetic nervous system activation in response to hyperosmolality as suggested by the decrease in neuronal activation in the RVLM and PVN, the associated increase in blood pressure also may be attenuated, thereby allowing a more rapid onset of water intake by EB-treated rats. In contrast, the increased blood pressure in OIL-treated OVX rats suggested by the neuronal activation in the RVLM and PVN may delay water intake until the induced hyperosmolality has reached levels sufficient to overcome this inhibition and thereby stimulate drinking.
Figure 6.

Reciprocally connected CNS areas are differentially activated by i.v. HS infusion in EB-treated rats. Left panels show schematics of CNS areas (adapted from [39]); middle panels show representative digital photomicrographs of fos immunolabeling in those areas.
Estradiol, salt and water intake, and hypo-osmolality
The studies described above suggesting that enhanced CNS sensitivity to increased osmolality underlies estradiol effects on water intake in response to hyperosmolality raise the obvious questions of whether behavioral responses to decreased osmolality and activity in central pathways that mediate such responses also are affected by estradiol. To address these questions, our laboratory is conducting studies utilizing the diuretic/natriuretic drug, Furosemide. In previous studies, sex differences in NaCl intake have been reported after Furosemide ([7] but see [43]). We opted to employ a single episode of Furosemide-induced sodium depletion in order to assess the possibility of estradiol effects on osmotic regulation prior to the establishment of behavioral changes that appear to involve long-term modulation of central pathways [29, 40].
Accordingly, in these studies, which also included male rats, rats were given s.c. injection of Furosemide (Intervet; 5 mg/kg body wt, 2X) or the 0.15 M NaCl vehicle (1.0 ml/kg body wt, 2X) on day 3 (see Figure 3) and then returned to their home cages where they had access to water, but not to food. Eighteen hours later (i.e., on the morning of day 4), one group of male rats, one group of OIL-treated OVX rats, and one group of EB-treated OVX rats were deeply anesthetized with sodium pentobarbital (Sigma; 0.5 ml). Immediately prior to perfusion, blood samples were taken from the heart into heparinized tubes and stored on ice until centrifuged for determination of plasma Na+ concentration (using ISE; Easylyte). Additional aliquots of blood were collected into microcapillary tubes and later centrifuged for determination of hematocrit.
Rats were perfused through the heart with 0.15 M NaCl followed by 4% paraformaldehyde. Brains were removed, postfixed in 4% paraformaldehyde overnight, and then placed in a 30% sucrose solution. Forebrains and hindbrains were cut into 1:3 series of 40-μm coronal sections using a cryostat (Leica), and stored in a cryoprotectant solution [28] at −20° C until processed.
These analyses focused on forebrain areas involved in the detection of osmolytes and hormones associated with hypo-osmolality (the circumventricular organs, SFO and OVLT) and on areas involved in sympathetic and/or hormonal responses to hypo-osmolality (PVN and SON). Thus, one series of free-floating forebrain sections from each rat was processed for fos immunolabeling using the avidin-biotin-peroxidase technique, as described in our previous publications [30, 34]. Briefly, after being rinsed in 0.05 M Tris-NaCl, soaked in 0.5% H2O2, rinsed again, and then soaked in 10% normal goat serum (NGS), sections were incubated in primary antibody (Santa Cruz SC-52, rabbit anti c-fos; 1:30,000 in 2% NGS) at 4°C overnight. Sections then were rinsed and incubated in the secondary antibody (biotinylated goat anti-rabbit; Vector Laboratories BA 1000; 1:300 in 2% NGS), before being rinsed again and then soaked in an avidin-biotin solution (Vectastain Elite ABC kit). Fos immunolabeling was visualized with nickel- intensified diaminobenzidine (Peroxidase substrate kit, SK-4100; Vector Laboratories). Sections were mounted on gel-coated slides before being dehydrated in an ascending series of alcohols (70%, 95%, and 100% EtOH) and xylenes and then coverslipped.
A Nikon microscope equipped with a camera and NIS-Elements AR 2.30 software was used to assess Fos immunolabeling in 2–3 matched, representative sections from the SFO, OVLT, SON, and PVN from each rat, identified based on neuroanatomical landmarks and coordinates [39]. The area of interest was outlined, and numbers of Fos-positive neurons were counted manually by observers blind to experimental conditions. Counts from SON and PVN were taken from one side; counts from the midline structures SFO and OVLT were taken in their entirety on each section. For each rat, average counts for each of the four areas were calculated; group means for each area were calculated from the averaged counts.
Additional groups of male, OIL-treated, and EB-treated rats were treated with Furosemide and, on day 4, 3-hr, 2-bottle (water and 0.5 M NaCl) intake tests were conducted immediately prior to blood collection, perfusion, brain collection and sectioning, and then immunolabeling as described. In these intake tests, we observed no hormone effects on NaCl intake stimulated by Furosemide (Figure 7). These results are consistent with findings by Scheidler and colleagues [43], but conflict with early reports by Chow and colleagues [7], who reported greater NaCl intake by female rats. While we cannot overlook methodological differences, including the route of administration or dose of EB, and/or differences in the NaCl concentration in the contradictory findings, our observation of differences in the associated water intake (Figure 7) provides additional information about a role for estradiol in body sodium balance. Specifically, significantly less water was consumed by EB-treated rats compared to both male rats and OIL-treated rats (both ps<0.001), whereas OIL-treated rats consumed more water than did male rats (p<0.05). These differences in water intake occurred despite similar NaCl intakes, and explain the difference in the calculated concentration of the fluid ingested (Figure 7, inset), with greater overall sodium concentration of the fluid ingested by EB-treated rats compared to that by OIL-treated rats (p<0.05).
Figure 7.
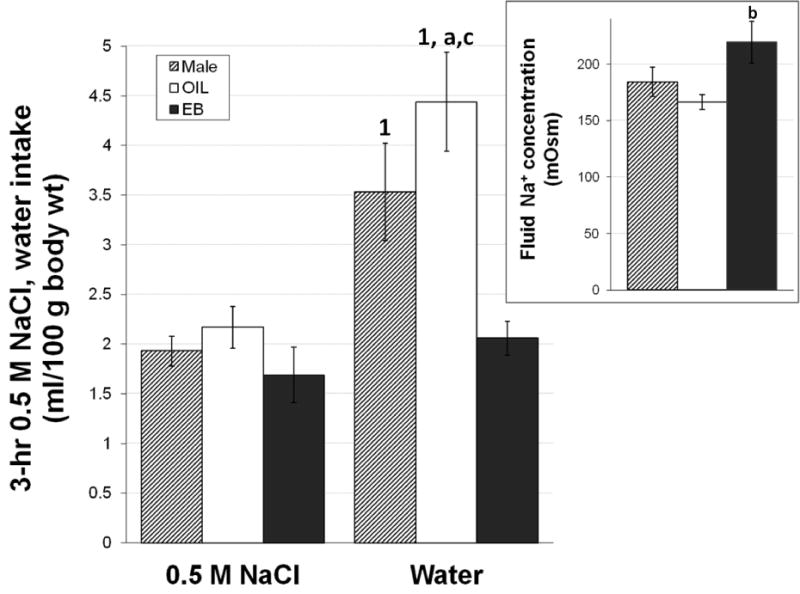
Intake of 0.5 M NaCl solution and water by male rats (hatched bars), OIL-treated OVX rats (white bars), and EB-treated OVX rats (black bars) in response to Furosemide-induced sodium depletion (Furosemide). Two-way RM ANOVA on solution intake revealed significant main effects of hormone (F2,20 = 6.46, p<0.01) and solution (F1,20 = 51.50, p<0.001), and a 2,20 = 7.87, p<0.05). Pairwise of the interaction showed that male rats and OIL-treated OVX rats consumed greater volumes of water than of 0.5 M NaCl, whereas there were no differences in water and NaCl intake by EB-treated rats. 0.5 M NaCl intake did not differ between the groups; however, both male rats and OIL-treated OVX rats drank more water than did EB-treated rats. OIL-treated rats also consumed more water than did male rats.
Inset. Na+ concentration of the fluid ingested in response to Furosemide by male rats (hatched bar), OIL-treated rats (white bar), and EB-treated rats (black bars). One-way ANOVA revealed significant differences that depended on hormone condition (F2,20) = 3.69, p<0.05. 1 = significantly greater than 0.5 M NaCl intake within hormone treatment (ps<0.001); a = significantly greater than water intake by male rats (p<0.05); b = significantly greater than fluid concentration consumed by OIL-treated rats (p<0.05); c = significantly greater than water intake by EB-treated rats (ps<0.001);
Assessment of plasma sodium concentration and hematocrit (Table 1) provides some insights into the underlying stimuli for NaCl and water intake in response to Furosemide treatment. Hyponatremia occurred in all groups after Furosemide, and this likely contributed to the comparable NaCl intake by all rats. Nevertheless, it should be noted that the induced hyponatremia was blunted in EB-treated rats, albeit not significantly. Furosemide treatment also produced a modest hypovolemia, as indicated by the elevated hematocrit, but no differences related to hormone condition were observed. Regardless of induced hyponatremia and hypovolemia, ingestion of water in combination with NaCl intake was sufficient to restore plasma sodium concentration and plasma volume in all groups, despite differences in water consumed and in the concentration of the ingested fluid. Overall, then, these findings suggest that the effects of Furosemide varied depending on hormone status. There were striking differences in behavioral responses that were not predicted from the subtle differences in renal effects, as evidenced by plasma sodium concentration and hematocrit. Ultimately, however, sodium regulation was achieved in the face of hyponatremia produced by Furosemide.
Table 1.
Plasma Na+ concentration and hematocrit in male rats and in OIL- and EB-treated OVX rats 18 hours after Vehicle treatment or Furosemide treatment with or without 3-hour, 2-bottle intake tests (0.5 M NaCl and water; Furosemide + salt intake). Two-way ANOVA revealed no effect of hormone, but a main effect of depletion condition for both plasma Na1 concentration (F2,59 = 12.76, p<0.001) and hematocrit (F2,61 = 16.74, p<0.001). Pairwise comparisons showed that, independent of hormone, plasma Na+ concentration after both Vehicle and Furosemide + salt intake were greater than that after Furosemide alone. In contrast, hematocrit after Furosemide alone was greater than that after Vehicle and Furosemide + salt intake.
| Plasma Na+ concentration (mOsm/L plasma water)
|
|||
|---|---|---|---|
| Vehicle2 | Furosemide | Furosemide + salt intake2 | |
| Male | 145.5 ± 1.3 | 139.2 ± 1.4 | 146.9 ± 1.6 |
| OIL | 143.5 ± 1.6 | 139.9 ± 1.9 | 146.9 ± 1.6 |
| EB | 146.1 ± 1.5 | 143.7 ± 1.1 | 147.8 ± 1.8 |
|
Hematocrit (%)
|
|||
|---|---|---|---|
| Vehicle | Furosemide1,3 | Furosemide + salt intake | |
| Male | 44.6 ± 1.1 | 46.8 ± 0.7 | 42.4 ± 1.1 |
| OIL | 41.8 ± 1.0 | 47.2 ± 1.4 | 39.6 ± 1.1 |
| EB | 42.8 + 1.4 | 46.4 + 1.3 | 42.4 + 1.0 |
= significantly greater than Vehicle (p<0.001);
= significantly greater than Furosemide (for Vehicle, p<0.01, for Furosemide + salt intake, p<0.001);
= significantly greater than Furosemide + salt intake (p<0.001)
Based on these estradiol-mediated differences in behavioral responses to Furosemide, we expected that CNS activity also would be affected by estradiol, as had been reported in a previous study that used multiple Furosemide depletions [13]. This expectation was not met. Fos immunolabeling in the circumventricular organ, SFO (Figure 8), increased after Furosemide whether or not the rats consumed 0.5 M NaCl and water; planned comparisons revealed that Furosemide alone stimulated greater fos labeling in the SFO of male rats compared to that in OVX rats with or without EB treatment (ps<0.05; Figure 9, top left). In contrast, labeling in the OVLT (Figure 9, top right), another circumventricular organ, was increased after Furosemide, whether or not rats consumed 0.5 M NaCl and water, and whether or not rats were treated with EB. Neuronal activation in the SON (Figure 9, bottom left) was not influenced by hormone status and appeared to depend on the behaviors stimulated by Furosemide–most likely the 0.5 M NaCl intake–rather than by Furosemide per se. Finally, fos immunolabeling in the PVN (Figure 9, bottom right) also appeared to depend on behavioral responses to Furosemide, but the difference was driven primarily by the greater labeling in OIL-treated rats, which was significantly greater than all other conditions.
Figure 8.

Schematic of the subfornical organ (SFO; adapted from [39]) and representative digital photomicrographs of fos immunolabeling in the SFO in male rats, OIL-treated OVX rats, and EB-treated OVX rats. Left panels show fos immunolabeling 18 hours after Furosemide; right panels show fos immunolabeling after a 3-hour salt and water intake test conducted 18 hours after Furosemide (Furosemide + salt intake).
Figure 9.

Fos immunolabeling in the SFO (top left), organum vasculosum of the lamina terminalis (top right), Supraoptic (bottom left) and Paraventricular (bottom right) nuclei of male rats (hatched bars), OIL-treated OVX rats (white bars), and EB-treated OVX rats (black bars) after Furosemide or Furosemide + salt intake.
SFO. Two-way ANOVA on fos immunolabeling revealed no effect of hormone, but a main effect of depletion condition (F2,62 = 28.11, p<0.001). Pairwise comparisons showed that, independent of hormone, fos labeling after Furosemide was greater than that after Vehicle. In addition, fos labeling after Furosemide + salt intake was greater than that after Furosemide and after Vehicle. Planned comparisons of fos labeling after Furosemide alone showed that labeling in the SFO of male rats was greater than that in both OIL- and EB-treated OVX rats.
OVLT. Two-way ANOVA on fos immunolabeling also revealed a main effect of depletion condition (F2,62 = 25.38, p<0.001), with pairwise comparisons showing significantly greater labeling after Furosemide and Furosemide + salt intake compared to that after Vehicle (both ps<0.001). There was no effect of hormone and no interaction between hormone and depletion condition.
1 = significantly greater than Vehicle (ps<0.001); 2 = significantly greater than Furosemide (p<0.05); b = significantly greater than OIL within a depletion condition; c = significantly greater than EB within a depletion condition (ps<0.05)
SON. Two-way ANOVA on fos immunolabeling revealed a main effect of depletion condition (F2,62 = 33.05, p<0.001), with pairwise comparisons showing greater labeling after Furosemide + salt intake compared to that after both Vehicle and Furosemide. There was no effect of hormone and no interaction between hormone and depletion condition.
PVN. Two-way ANOVA on fos immunolabeling showed a main effect of depletion condition (F2,62 = 4.17, p<0.05) and a significant interaction between hormone and depletion condition (F2,62 = 2.87, p<0.05). Pairwise comparisons revealed that, independent of hormone, fos labeling was greater after Furosemide + salt intake compared to both vehicle and Furosemide (p<0.05). Moreover, fos labeling in OIL-treated rats after Furosemide + salt intake was greater than that in all other conditions.
1, 1′ = significantly greater than Vehicle (p<0.001, 0.01); 2, 2′ = significantly greater than Furosemide (p<0.01, 0.05); * = significantly greater than all other conditions (ps<0.05–0.001)
The answer to whether behavioral responses to decreased osmolality and activity in central pathways that mediate such responses are affected by estradiol remains unclear. Although EB treatment had no effect on 0.5 M NaCl stimulated by Furosemide, the associated water intake was significantly reduced in EB-treated rats, such that the concentration of the fluid ingested was greater. These observations suggest that estradiol also increases sensitivity to hypo- osmolality; however, the sex differences observed in activation of some CNS areas after Furosemide were not estradiol-dependent. Rather, independent of estradiol status at the time of testing, females differed from males. In contrast, activation of other CNS areas was attributable to the consequences of the behavior, alone or in conjunction with estradiol-independent sex differences. It is possible that additional studies employing multiple Furosemide treatments may reveal, not only differences in 0.5 M NaCl intake (e.g., [7]), but also in neuronal activation in areas of the CNS involved in the response (e.g., [13]), as plasticity in these neural pathways seems to be an important determinant of the magnitude of the behavioral responses elicited by Furosemide-induced sodium depletion [29, 40]. Clearly, additional studies will be necessary to more fully elaborate the effects of estradiol on behavioral and central responses to hypo-osmolality, and these studies are the focus of ongoing research in our lab. Of particular interest in these studies is whether hindbrain pathways also are important in responses to hypo-osmolality as they are in responses to hyperosmolality, and whether the role of various CNS areas in Furosemide-induced hypo-osmolality change with time and/or experience with sodium depletion.
Conclusions
Findings from a series of studies from our lab show that, in EB-treated rats, hypernatremia produces more rapid water intake that is not attributable to differences in plasma osmolality. Rather, this enhanced sensitivity to hyperosmolality may involve selective effects on CNS areas, possibly those with reciprocal connections. Future studies will determine whether these connections play a functional role in modulating behavioral responses to hypernatremia and, in so doing, determine whether estradiol-mediated effects on blood pressure alter the latency to drink water. More work remains to better understand the relationship between behaviors and CNS activity during hypo-osmolality. It is clear, however, that behavioral differences observed in EB-treated rats are not attributable to the detection of decreased osmolality or in response to Na+-regulating hormones by the SFO. An interesting possibility is suggested by an integration of the findings: estradiol-independent sex differences (e.g., Figure 9) suggest that the effects of estrogens early in development may set responding, but responses may also be altered by adult patterns of estradiol fluctuations (e.g., Figure 5, 7).
In a broad sense, the goal of our lab is to understand how estradiol affects CNS pathways to elicit behavioral responses to osmotic/sodium challenges. Some progress has been made but, clearly, many questions remain. Do the CNS circuits that underlie behavioral responses to hypo- vs. hyper-osmolality overlap or are they separable? Are central pathways controlling water and salt intake both influenced by estradiol and, if so, under what conditions? What are the signals that activate these pathways, and what neurotransmitters are involved in conveying those signals? What estrogen receptor mechanism/subtype is important in the responses? How might the organizational vs. activational effects of estradiol determine the behavioral and CNS responses? Answering these questions has far-reaching implications–not only for addressing the central effects of estrogen in body sodium regulation, but also for advancing our understanding of sex hormone-dependent differences in physiological processes that support healthy functioning. With the aging of our population, and the increase in the incidence of hypertension in postmenopausal women, better understanding of the effects of estrogen in body fluid balance—and, therefore, of its absence [e.g., 44, 55]—cannot be disregarded.
Highlights.
EB treatment of OVX rats affects behavioral and CNS responses to osmotic challenges
Hyperosmolality: EB elicits rapid-onset water drinking and decreases CNS activity
Hypo-osmolality: EB does not affect salt intake but blunts associated water intake
Hypo-osmolality: activation in SFO is blunted in females but independent of EB
EB effects on responses to osmotic challenges depend on challenge and on CNS area
Acknowledgments
Grants from the National Institute on Deafness and Communications Disorders (DC-06360) and the Oklahoma Center for the Advancement of Science and Technology Health Research Program (HR-09-123) supported this research.
These studies would not have been possible without undergraduate students Megan Buford, Minh Ngo, and Jennifer Hackett; graduate students Alexis Jones, Eryn Bass, and Eric Krause; and Dr. Robert C. Contreras. Their contributions are gratefully acknowledged.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Literature Cited
- 1.Barron WM, Schreiber J, Lindheimer MD. Effect of ovarian sex steroids on osmoregulation and vasopressin secretion in the rat. Am J Physiol. 1986;250:E352–61. doi: 10.1152/ajpendo.1986.250.4.E352. [DOI] [PubMed] [Google Scholar]
- 2.Bealer SL. Central control of cardiac baroreflex responses during peripheral hyperosmolality. Am J Physiol Regul Integr Comp Physiol. 2000;278:R1157–63. doi: 10.1152/ajpregu.2000.278.5.R1157. [DOI] [PubMed] [Google Scholar]
- 3.Beauchamp GK. The human preference for excess salt. Amer Scientist. 1987;75:27–33. [Google Scholar]
- 4.Bossmar T, Forsling M, Akerlund M. Circulating oxytocin and vasopressin is influenced by ovarian steroid replacement in women. Acta Obstet Gynecol Scand. 1995;74:544–8. doi: 10.3109/00016349509024387. [DOI] [PubMed] [Google Scholar]
- 5.Calzone WL, Silva C, Keefe DL, Stachenfeld NS. Progesterone does not alter osmotic regulation of AVP. Am J Physiol Regul Integr Comp Physiol. 2001;281:R2011–20. doi: 10.1152/ajpregu.2001.281.6.R2011. [DOI] [PubMed] [Google Scholar]
- 6.Carlberg KA, Fregly MJ, Fahey M. Effects of chronic estrogen treatment on water exchange in rats. Am J Physiol. 1984;247:E101–10. doi: 10.1152/ajpendo.1984.247.1.E101. [DOI] [PubMed] [Google Scholar]
- 7.Chow SY, Sakai RR, Witcher JA, Adler NT, Epstein AN. Sex and sodium intake in the rat. Behav Neurosci. 1992;106:172–80. doi: 10.1037//0735-7044.106.1.172. [DOI] [PubMed] [Google Scholar]
- 8.Clarke SNDA, Ossenkopp K-P. Taste reactivity responses in rats: influence of sex and the estrous cycle. Am J Physiol Regul Integr Comp Physiol. 1998;274:R718–724. doi: 10.1152/ajpregu.1998.274.3.R718. [DOI] [PubMed] [Google Scholar]
- 9.Cooke PS, Buchanan DL, Young P, Setiawan T, Brody J, Korach KS, Taylor J, Lubahn DB, Cunha GR. Stromal estrogen receptors mediate mitogenic effects of estradiol on uterine epithelium. Proc Natl Acad Sci U S A. 1997;94:6535–40. doi: 10.1073/pnas.94.12.6535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cunningham ET, Jr, Miselis RR, Sawchenko PE. The relationship of efferent projections from the area postrema to vagal motor and brain stem catecholamine-containing cell groups: an axonal transport and immunohistochemical study in the rat. Neuroscience. 1994;58:635–48. doi: 10.1016/0306-4522(94)90087-6. [DOI] [PubMed] [Google Scholar]
- 11.Curtis KS, Contreras RJ. Sex differences in electrophysiological and behavioral responses to NaCl taste. Behav Neurosci. 2006;120:917–24. doi: 10.1037/0735-7044.120.4.917. [DOI] [PubMed] [Google Scholar]
- 12.Curtis KS, Davis LM, Johnson AL, Therrien KL, Contreras RJ. Sex differences in behavioral taste responses to and ingestion of sucrose and NaCl solutions by rats. Physiol Behav. 2004;80:657–64. doi: 10.1016/j.physbeh.2003.11.007. [DOI] [PubMed] [Google Scholar]
- 13.Dalmasso C, Amigone JL, Vivas L. Serotonergic system involvement in the inhibitory action of estrogen on induced sodium appetite in female rats. Physiol Behav. 2011;104:398–407. doi: 10.1016/j.physbeh.2011.04.029. [DOI] [PubMed] [Google Scholar]
- 14.Dampney RA, Coleman MJ, Fontes MA, Hirooka Y, Horiuchi J, Li YW, Polson JW, Potts PD, Tagawa T. Central mechanisms underlying short- and long-term regulation of the cardiovascular system. Clin Exp Pharmacol Physiol. 2002;29:261–8. doi: 10.1046/j.1440-1681.2002.03640.x. [DOI] [PubMed] [Google Scholar]
- 15.Di Lorenzo PM, Monroe S. Taste responses in the parabrachial pons of male, female and pregnant rats. Brain Res Bull. 1989;23:219–27. doi: 10.1016/0361-9230(89)90151-2. [DOI] [PubMed] [Google Scholar]
- 16.Di Lorenzo PM, Monro S. e Taste responses in the parabrachial pons of ovariectomized rats. Brain Res Bull. 1990;25:741–8. doi: 10.1016/0361-9230(90)90052-2. [DOI] [PubMed] [Google Scholar]
- 17.Duffy VB, Bartoshuk LM, Striegel-Moore R, Rodin J. Taste changes across pregnancy. Ann N Y Acad Sci. 1998;855:805–9. doi: 10.1111/j.1749-6632.1998.tb10663.x. [DOI] [PubMed] [Google Scholar]
- 18.Eckert T, Forsling ML, Schwarzberg H. The effect of combined oestrogen and progesterone replacement on the renal responses to oxytocin and vasopressin in ovariectomized rats. Eur J Endocrinol. 1999;141:297–302. doi: 10.1530/eje.0.1410297. [DOI] [PubMed] [Google Scholar]
- 19.Forsling ML, Peysner K. Pituitary and plasma vasopressin concentrations and fluid balance throughout the oestrous cycle of the rat. J Endocrinol. 1988;117:397–402. doi: 10.1677/joe.0.1170397. [DOI] [PubMed] [Google Scholar]
- 20.Freece JA, Van Bebber JE, Zierath DK, Fitts DA. Subfornical organ disconnection alters Fos expression in the lamina terminalis supraoptic nucleus, and area postrema after intragastric hypertonic NaCl. Am J Physiol Regul Integr Comp Physiol. 2005;288:R947–55. doi: 10.1152/ajpregu.00570.2004. [DOI] [PubMed] [Google Scholar]
- 21.Fregly MJ. Effect of an oral contraceptive on NaCl appetite and preference threshold in rats. Pharmacol Biochem Behav. 1973;1:61–5. doi: 10.1016/0091-3057(73)90056-7. [DOI] [PubMed] [Google Scholar]
- 22.Frye CA, Demolar GL. Menstrual cycle and sex differences influence salt preference. Physiol Behav. 1994;55:193–7. doi: 10.1016/0031-9384(94)90031-0. [DOI] [PubMed] [Google Scholar]
- 23.Geary N, Asarian L. Cyclic estradiol treatment normalizes body weight and test meal size in ovariectomized rats. Physiol Behav. 1999;67:141–7. doi: 10.1016/s0031-9384(99)00060-8. [DOI] [PubMed] [Google Scholar]
- 24.Goldstein P, Leshem M. Dietary sodium, added salt, and serum sodium associations with growth and depression in the U.S. general population Appetite. 2014;79:83–90. doi: 10.1016/j.appet.2014.04.008. [DOI] [PubMed] [Google Scholar]
- 25.Graves NS, Hayes H, Fan L, Curtis KS. Time course of behavioral physiological, and morphological changes after estradiol treatment of ovariectomized rats. Physiol Behav. 2011;103:261–267. doi: 10.1016/j.physbeh.2011.02.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hartley DE, Dickson SL, Forsling ML. Plasma vasopressin concentrations and Fos protein expression in the supraoptic nucleus following osmotic stimulation or hypovolaemia in the ovariectomized rat: effect of oestradiol replacement. J Neuroendocrinol. 2004;16:191–7. doi: 10.1111/j.0953-8194.2004.01150.x. [DOI] [PubMed] [Google Scholar]
- 27.Haywood JR, Hinojosa-Laborde C. Sexual dimorphism of sodium-sensitive renal-wrap hypertension. Hypertension. 1997;30:667–71. doi: 10.1161/01.hyp.30.3.667. [DOI] [PubMed] [Google Scholar]
- 28.Hoffman GE, Le WW. Just cool it! Cryoprotectant anti-freeze in immunocytochemistry and in situ hybridization. Peptides. 2004;25:425–31. doi: 10.1016/j.peptides.2004.02.004. [DOI] [PubMed] [Google Scholar]
- 29.Hurley SW, Zhang Z, Beltz TG, Xue B, Johnson AK. Sensitization of sodium appetite: evidence for sustained molecular changes in the lamina terminalis. Am J Physiol Regul Integr Comp Physiol. 2014;307:R1405–12. doi: 10.1152/ajpregu.00210.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Jones AB, Bass EE, Fan L, Curtis KS. Estradiol selectively reduces central neural activation induced by hypertonic NaCl infusion in ovariectomized rats. Physiol Behav. 2012;107:192–200. doi: 10.1016/j.physbeh.2012.06.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Jones AB, Curtis KS. Differential effects of estradiol on drinking by ovariectomized rats in response to hypertonic NaCl or isoproterenol: Implications for hyper- vs. hypo-osmotic stimuli for water intake. Physiol Behav. 2009;98:421–426. doi: 10.1016/j.physbeh.2009.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kisley LR, Sakai RR, Fluharty SJ. Estrogen decreases hypothalamic angiotensin II AT1 receptor binding and mRNA in the female rat. Brain Res. 1999;844:34–42. doi: 10.1016/s0006-8993(99)01815-6. [DOI] [PubMed] [Google Scholar]
- 33.Krause EG, Curtis KS, Davis LM, Stowe JR, Contreras RJ. Estrogen influences stimulated water intake by ovariectomized female rats. Physiol Behav. 2003;79:267–74. doi: 10.1016/s0031-9384(03)00095-7. [DOI] [PubMed] [Google Scholar]
- 34.Krause EG, Curtis KS, Markle JP, Contreras RJ. Oestrogen affects the cardiovascular and central responses to isoproterenol of female rats. J Physiol. 2007;582:435–447. doi: 10.1113/jphysiol.2007.131151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Krause EG, Curtis KS, Stincic TL, Markle JP, Contreras RJ. Oestrogen and weight loss decrease isoproterenol-induced Fos immunoreactivity and angiotensin type 1 mRNA in the subfornical organ of female rats. J Physiol. 2006;573:251–62. doi: 10.1113/jphysiol.2006.106740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Miura S, Tsong YY, Koide SS. Hormonal effects of estrogen-protein conjugates on rat uterus. Biol Reprod. 1971;5:340–2. doi: 10.1093/biolreprod/5.3.340. [DOI] [PubMed] [Google Scholar]
- 37.Osborn JW, Collister JP, Carlson SH. Angiotensin and osmoreceptor inputs to the Area Postrema: role in long-term control of fluid homeostasis and arterial pressure. Clin Exper Pharmacol Physiol. 2000;27:443–449. doi: 10.1046/j.1440-1681.2000.03263.x. [DOI] [PubMed] [Google Scholar]
- 38.Ota M, Crofton JT, Liu H, Festavan G, Share L. Increased plasma osmolality stimulates peripheral and central vasopressin release in male and female rats. Am J Physiol. 1994;267:R923–8. doi: 10.1152/ajpregu.1994.267.4.R923. [DOI] [PubMed] [Google Scholar]
- 39.Paxinos G, Watson C. The rat brain in stereotaxic coordinates. San Diego: Academic Press; 1998. [Google Scholar]
- 40.Roitman MF, Na E, Anderson G, Jones TA, Bernstein IL. Induction of a salt appetite alters dendritic morphology in nucleus accumbens and sensitizes rats to amphetamine. J Neurosci. 2002;22:RC225 1–5. doi: 10.1523/JNEUROSCI.22-11-j0001.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Sawchenko PE, Swanson LW. Immunohistochemical identification of neurons in the paraventricular nucleus of the hypothalamus that project to the medulla or to the spinal cord in the rat. J Comp Neurol. 1982;205:260–72. doi: 10.1002/cne.902050306. [DOI] [PubMed] [Google Scholar]
- 42.Sawchenko PE, Swanson LW. The organization of noradrenergic pathways from the brainstem to the paraventricular and supraoptic nuclei in the rat. Brain Res. 1982;257:275–325. doi: 10.1016/0165-0173(82)90010-8. [DOI] [PubMed] [Google Scholar]
- 43.Scheidler MG, Verbalis JG, Stricker EM. Inhibitory effects of estrogen on stimulated salt appetite in rats. Behav Neurosci. 1994;108:141–50. [PubMed] [Google Scholar]
- 44.Stachenfeld NS, DiPietro L, Palter SF, Nadel ER. Estrogen influences osmotic secretion of AVP and body water balance in postmenopausal women. Am J Physiol. 1998;274:R187–95. doi: 10.1152/ajpregu.1998.274.1.R187. [DOI] [PubMed] [Google Scholar]
- 45.Stachenfeld NS, Keefe DL. Estrogen effects on osmotic regulation of AVP and fluid balance. Am J Physiol Endocrinol Metab. 2002;283:E711–21. doi: 10.1152/ajpendo.00192.2002. [DOI] [PubMed] [Google Scholar]
- 46.Stern JE, Zhang W. Preautonomic neurons in the paraventricular nucleus of the hypothalamus contain estrogen receptor beta. Brain Res. 2003;975:99–109. doi: 10.1016/s0006-8993(03)02594-0. [DOI] [PubMed] [Google Scholar]
- 47.Stocker SD, Madden CJ, Sved AF. Excess dietary salt intake alters the excitability of central sympathetic networks. Physiol Behav. 2010;100:519–524. doi: 10.1016/j.physbeh.2010.04.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Stocker SD, Stricker EM, Sved AF. Acute hypertension inhibits thirst stimulated by ANG II, hyperosmolality, or hypovolemia in rats. Am J Physiol Regul Integr Comp Physiol. 2001;280:R214–24. doi: 10.1152/ajpregu.2001.280.1.R214. [DOI] [PubMed] [Google Scholar]
- 49.Swanson LW, Sawchenko PE, Berod A, Hartman BK, Helle KB, Vanorden DE. An immunohistochemical study of the organization of catecholaminergic cells and terminal fields in the paraventricular and supraoptic nuclei of the hypothalamus. J Comp Neurol. 1981;196:271–85. doi: 10.1002/cne.901960207. [DOI] [PubMed] [Google Scholar]
- 50.Swenson KL, Sladek CD. Gonadal steroid modulation of vasopressin secretion in response to osmotic stimulation. Endocrinology. 1997;138:2089–2097. doi: 10.1210/endo.138.5.5142. [DOI] [PubMed] [Google Scholar]
- 51.Thrasher TN, Fregly MJ. Responsiveness to various dipsogenic stimuli in rats treated chronically with norethynodrel, ethinyl estradiol and both combined. J Pharmacol Exp Ther. 1977;201:84–91. [PubMed] [Google Scholar]
- 52.Thunhorst RL, Johnson AK. Effects of arterial pressure on drinking and urinary responses to intracerebroventricular angiotensin II. Am J Physiol. 1993;264:R211–7. doi: 10.1152/ajpregu.1993.264.1.R211. [DOI] [PubMed] [Google Scholar]
- 53.Verma P, Mahajan KK, Mittal S. A Ghildiyal Salt preference across different phases of menstrual cycle. Indian J Physiol Pharmacol. 2005;49:99–102. [PubMed] [Google Scholar]
- 54.Wang YX, Crofton JT, Liu H, Sato K, Brooks DP, Share L. Estradiol attenuates the antidiuretic action of vasopressin in ovariectomized rats. Am J Physiol. 1995;268:R951–7. doi: 10.1152/ajpregu.1995.268.4.R951. [DOI] [PubMed] [Google Scholar]
- 55.Whelton PK, Appel LJ, Sacco RL, Anderson CA, Antman EM, Campbell N, Dunbar SB, Frohlich ED, Hall JE, Jessup M, Labarthe DR, MacGregor GA, Sacks FM, Stamler J, Vafiadis DK, Van Horn LV. Sodium, blood pressure, and cardiovascular disease: further evidence supporting the American Heart Association sodium reduction recommendations. Circulation. 2012;126:2880–9. doi: 10.1161/CIR.0b013e318279acbf. [DOI] [PubMed] [Google Scholar]
- 56.Wilkins L, Richter CP. A great craving for salt by a child with cortico-adrenal insufficiency. JAMA. 1940;114:3. [Google Scholar]
- 57.Woolley CS, McEwen BS. Roles of estradiol and progesterone in regulation of hippocampal dendritic spine density during the estrous cycle in the rat. J Comp Neurol. 1993;336:293–306. doi: 10.1002/cne.903360210. [DOI] [PubMed] [Google Scholar]
- 58.Yang Z, Han D, Coote JH. Cardiac sympatho-excitatory action of PVN-spinal oxytocin neurones. Auton Neurosci. 2009;147:80–85. doi: 10.1016/j.autneu.2009.01.013. [DOI] [PubMed] [Google Scholar]
- 59.http://www.heart.org/HEARTORG/GettingHealthy/NutritionCenter/HealthyEating/Reducing-Sodium-in-a-Salty-World_UCM_457519_Article.jsp; Reducing Sodium in a Salty World; updated January 26, 2015; downloaded February 28, 2015.