

Abstract
High rosmarinic acid (RA) productivity has been achieved by applying jasmonic acid and yeast extract elicitors to the in vitro sprout culture of Orthosiphon aritatus (IOSC). The highest RA accumulation from three solvents was detected in IOSC after treatment with yeast extract (5 g/L). HPLC analysis clearly confirmed a drastic increase in RA subjected to yeast extract elicitation. Therefore, this yeast extract elicited IOSC was chosen for a lactic acid bacteria (LAB) fermentation study as a model system. This selected IOSC was subjected to different types of LAB fermentations (Lactobacillus plantarum ATCC 8014 and Lactobacillus acidophilus NCFM) for different periods of time 24, 48 and 72 h. The LAB fermentations consisted of solid state fermentations (SSF) and liquid state fermentations (LSF) in a Digital Control Unit (DCU) fermenter system. The aim was to determine the effect of fermentation on the antioxidant properties of the plant extract. Results indicated that all types of LAB fermentation decreased the level of RA and total phenolics, however, a slight increase in total flavonoids and flavonols was observed in SSF samples. HPLC results confirmed that the longer the fermentation, the greater the reduction in RA content. The highest reduction was obtained in the sample of LSF inoculated with L. plantarum for a period of 72 h. The temperature of fermentation (37 °C) was predicted as contributing to the declining level in RA content. The loss in RA was concomitant with a loss of total antioxidant activity (1,1-diphenyl-2-picrylhydrazyl (DPPH) scavenging activity, Trolox Equivalent Antioxidant Capacity (TEAC), and Superoxide Dismutase (SOD)-like activity). These results indicate that RA is the major contributor to the antioxidant activity of this plant.
Keywords: antioxidant, Orthosiphon aristatus, fermentation, in vitro sprout culture
Abbreviations
- RA
Rosmarinic Acid
- IOSC
in vitro Sprout Culture of Orthosiphon aristatus
- LAB
Lactic Acid Bacteria
- SSF
Solid State Fermentations
- LSF
Liquid State Fermentations
- LP
Lactobacillus plantarum
- LA
Lactobacillus acidophilus
- OD
Optical Density
- LP-SSF
Solid State Fermentations inoculated with Lactobacillus plantarum
- LA-SSF
Solid State Fermentations inoculated with Lactobacillus acidophilus
- LP-LSF
Liquid State Fermentations inoculated with Lactobacillus plantarum
- LA-LSF
Liquid State Fermentations inoculated with Lactobacillus acidophilus
- DPPH
1,1-diphenyl-2- picrylhydrazyl
- TEAC
Trolox Equivalent Antioxidant Capacity
- SOD
Superoxide Dismutase
- JA
MS medium with Jasmonic Acid
- YE
MS medium with Yeast Eextract
- TE
Trolox Equivalent
- GAE
Gallic Acid Equivalent
- QE
Quercetin Equivalent
- DW
Dry Weight
1. Introduction
Rosmarinic acid (RA) is an ester of caffeic acid and 3, 4-dihydroxyphenyllacticacid. It is commonly found in plants from the Lamiaceae family, e.g., Orthosiphon aristatus, and Coleus blumei. This caffeic acid is one of the most important and targeted phenolic compounds in formulating functional food and supplements due to its interesting biological activities, such as antiviral, antibacterial, anti-inflammatory and antioxidant [1].
RA from medicinal plants such as Orthosiphon aritatus that contain naturally occurring antioxidants is preferable [2]. Orthosiphon aristatus is a native tropic-medicinal plant from Southeast Asia. It belongs to the Lamiaceae family. RA was identified as the most abundant polyphenol (caffeic acid derivative) in O. aristatus and it was thought to be the main contributor for the therapeutic effects and antioxidant activity of this plant [3]. The escalating demand of RA led us to conduct this research in order to increase the production of RA in Orthosiphon aristatus and to characterize the antioxidant properties of this plant extract, which may provide useful information for better formulation of the application of functional foods.
Indeed, plant cell cultures that accumulate RA have been proposed for production of this phenolic compound [2]. Plant tissue cultures using plantlets that were obtained from direct shoot organogenesis are generally preferred for mass production of the targeted compound as they are more genetically stable [4]. Moreover, as a native tropic-medicinal plant, to grow Orthosiphon aristatus in a sub-tropic climate area has been considered to be relatively difficult. Therefore, in vitro sprout culture of Orthosiphon aristatus (IOSC) was used as the media for this study.
Other reasons for using IOSC are (1) this in vitro system may easily control biotic and abiotic stresses on the cultivar, (2) this system allows the harvest of this plant under any conditions regardless of climate condition and seasons, (3) and also through this system, rapid growth mass production can be achieved as well as the combination with sustainable secondary metabolites production [4,5]. Furthermore, this system can also be used to facilitate the fermentation study of in vitro plant culture.
To improve the production of secondary metabolites of in vitro plant cultures, several strategies such as supplementation and elicitation have been employed [6], for instance, the use of yeast extract [7] and jasmonic acid [8]. However, there has been no report on the use of these elicitors in in vitro sprout culture of Orthosiphon aristatus to enhance RA production.
Fermentation, as a biotechnological process, is considered one of the oldest technologies, which is simple, natural and valuable. It can maintain and improve the safety, nutritional, sensory and shelf-life properties of many food products including plant food products [9]. Lactic acid bacteria (LAB) are the main bacteria responsible for the fermentation of plant food products. Most plant products can be lactic-acid-fermented, yet cucumber, cabbage and olives are the only plant products that are fermented in large volumes for human consumption [10].
It has been generally known that plants produce secondary metabolites that are of importance in the healthcare, food, flavor and cosmetics industries. Many biochemical changes occur during fermentation, leading to alteration in the composition of plant secondary metabolites which affects product properties such as antioxidant activity [11]. Phenolics are among the secondary metabolites of many plants that have been studied including the changes subjected to the LAB fermentation process [11,12,13,14,15]. However, most of the published reports used a terrestrial plants system as a media for studying the effect of LAB fermentation on their antioxidant properties [16,17,18,19,20,21,22]. We have not yet come across any published report describing the effect of LAB fermentation on antioxidant properties using an in vitro plant system. The advantage of using terrestrial plants in an LAB fermentation study is that the plant is ready to use for fermentation. Conversely, the disadvantage of using a terrestrial plant in the raw material form is that the effect of the available natural microorganisms in the plant is unavoidable. Therefore, IOSC was established to facilitate this LAB fermentation study.
There are two main objectives for using IOSC as a model in this study, namely: (1) to find the high RA line of IOSC for LAB fermentation, and (2) to study the effect of LAB fermentation on RA content and antioxidant properties of selected IOSCs. These two objectives result in several target approaches as follows: (a) to select the high RA line and high antioxidant properties of IOSC by elicitation; (b) to characterize the antioxidant properties of IOSC using three different solvents after elicitation treatment; (c) to characterize the effect of LAB fermentation on the antioxidant properties of selected IOSCs; and (d) to identify the individual phenolic acids of IOSCs and their quantification changing with elicitation and LAB fermentation.
2. Experimental Section
The flowchart of the current study is illustrated in Figure 1.
Figure 1.

Flowchart of the effect of LAB fermentation on antioxidant properties of IOSC.
2.1. Materials
2.1.1. Plant Materials
Orthosiphon aristatus with article number 600646 was purchased online from Pflanzenversand Hans-Günter Röpke (refer to: http://www.pflanzenkindergarten.de). This was used to establish the in vitro sprout culture.
2.1.2. Culture Media
Agar Cell culture media and MS medium (Merck, Darmstadt, Germany), Oxoid Yeast Extract (Code L21) (Unipath LTD, Hampshire, England), (±) Jasmonic acid (Product of Israel, Plant cell culture tested, C12H18O3) (Sigma-Aldrich Chemie GmbH, Steinheim, Germany), and other materials were analytical and plant cell culture tested grades.
2.1.3. Chemicals for Analysis
Gallic acid, ABTS (2,2'-azino-bis (3-ethylbenzthiazoline-6-sulphonic acid)) and standards of phenolic acids were obtained from Sigma-Aldrich Chemie GmbH (Steinheim, Germany); Quercetin Dihydrate (C15H10O7·2H2O ~99% HPLC) was from Fluka, Biochemika (Neu-Ulm, Germany). All used chemicals were analytical HPLC grade.
2.2. Methods
2.2.1. Establishment of IOSC
The established procedure of IOSC was based on the previous report of Shevchenko et al. [5] with liquid MS medium modification as follows: MS medium (control), MS with jasmonic acid (100 mM) (JA), and MS medium with yeast extract (5 g/L) (YE).
2.2.2. Fermentation Study of Selected IOSCs
The highest RA content in IOSC samples was chosen and subjected to fermentation study.
2.2.3. LAB Fermentation of IOSC
Lactobacillus plantarum ATCC 8014 (supplied by the Department of Food Biotechnology and Food Process Engineering, Institute of Food Technology and Food Chemistry, TU Berlin, Germany) and Lactobacillus acidophilus NCFM, Danisco (a donation from Institute of Biotechnologie, TU Berlin) were used in this LAB fermentation study of IOSC.
2.2.4. Inoculums Preparation
For preparation of inoculums, cells from stock culture were transferred to 25 mL sterile de Man, Rogosa and Sharpe (MRS) broth and incubated for 24 h at 37 °C. The growth density was measured at a wavelength of 620 nm using a spectrophotometer (PerkinElmer Precisely Lambda 25 UV/VIS Spectrometer, PerkinElmer, MA, USA). The bacteria were kept under stationary conditions until they reached an absorbance of 5.0. One ml of inoculums contained around 109 colony forming units (cfu). Dilutions were made with MRS broth to bring the lactobacillus to an optical density at 620 nm of 0.1 ± 0.02 corresponding to a cell density of 105–107 cfu/mL.
2.2.5. The Effect of IOSC Extract on Viable Cell LAB Strains Growth
The selected HPLC IOSC extract (10 mg/mL) (HPLC extraction explained below) with the highest concentration of RA was further evaluated for its inhibitory effect on the growth of LAB strains. The method of Alberto et al. [23] was used with modification as follows: 15% (0.75 mL) of IOSC extract was aseptically filtered with a 0.2 µm sterile syringe (VWR International Gmbh, Langenfeld, Germany) and added to 4.25 mL inoculated LAB in MRS broth (OD620 0.1) in tubes and the total concentration of extract was 1.5 mg/mL. The samples tubes were incubated as explained above. The LAB growth was evaluated by direct measurement of OD620 every 4 h for a period of 24 h comparing the growth density with those obtained from inoculated LAB controls (OD620 0.1) without IOSC extract.
2.2.6. Solid State Fermentations (SSF)
This was carried out using 10 g of three-week freshly harvested IOSC inoculated with 5% (v/w) of the above culture and aseptically placed in Petri dishes in a climatic incubator controlled at 37 °C and 90% relative humidity for 24, 48 and 72 h. After fermentation, the samples were dipped in sterile water and then immersed in liquid nitrogen and freeze-dried. SSF were performed in duplicate.
2.2.7. Total LAB
Viable cells of LAB were quantified at the end of fermentation (24, 48 and 72 h) using the plate enumeration method [24]. A total of 1 g of SSF was blended in 9 mL of Ringer’s solution (No. 15525, Merck, Darmstadt, Germany) and serially diluted for plate counting on MRS agar. The viable cell numbers were determined after 48 h of incubation at 37 °C under anaerobic condition produced by anaerobic kits (Anaerocult® A, Merck, Darmstadt, Germany).
2.2.8. Liquid State Fermentations (LSF)
LSF were performed under aseptic conditions for 24, 48 and 72 h at a temperature of 37 °C in a 1 L DCU system with digital measurement and control system for fermenter (B. Braun Biotech International, Melsungen, Germany) by aseptically placing 7.5 g IOSC (1.5% g/v) in MRS broth (500 mL) with adjusted initial OD620 of 0.1. Anaerobic fermentation experiments were conducted without aeration at an agitation speed of 150 rpm. LSF samples were cleaned with sterile water and dipped in liquid nitrogen and then freeze-dried.
2.2.9. Viable Cells LAB Growth
LSF were started at OD620 0.1 and by the end of fermentation the growth of the cells was recorded by taking OD620 measurements against the control without IOSC material. The growth density (OD620) determination was done in duplicate.
2.2.10. Sample Preparation and Extraction of the IOSC
After two weeks the IOSC and LAB fermented samples were harvested, weighed and immersed in liquid nitrogen to prevent volatilization of phenolic compounds and then were freeze-dried. The lyophilized samples were ground with a flint mill (Retsch GmBH, Haan, Germany) (20,000 rpm, 2 min) to a fine powder. The dried powder of IOSC was extracted using three different solvents: 70% methanol, 80% ethanol and distilled water. The dried powders of IOSC samples were weighed accurately at 100 mg. They were treated with 5 mL of each solvent then placed in an ultrasonic water bath at 75 °C (Bandelin Sonorex Digital 10 P, Bandelin electronic GmbH & Co. KG, Berlin, Germany) for 2 h. The mixtures were centrifuged at 13,000 rpm for 5 min and the supernatants were collected as different samples. The extraction was repeated twice until the final concentration of the IOSC extract was 10 mg/mL. The extract was stored at −20 °C until further use. All samples and determinations were prepared and measured in triplicate.
2.2.11. Antioxidant Properties of the IOSC
The antioxidant properties consist of characterizing the antioxidant activity and components by rapid reliable spectrophotometric measurement. The characterization of antioxidant activity was estimated based on DPPH, TEAC methods, and SOD-like activity followed by measurements of the antioxidant components: total phenolics, flavonoids, flavonols and RA. The DPPH free radical scavenging activity of the sample extract was conducted as described by Mohdaly et al. [25]. Then the percentage inhibition activity as relative antioxidant activity was calculated based on the following formula:
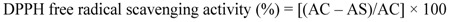 |
where AC is the absorbance of the control, and AS is the absorbance of the samples.
Meanwhile, the TEAC assay was performed by following standard operation procedures (SOP) of Re et al. [26]. A Trolox® standard calibration curve of OD734nm versus concentration of Trolox in mmol·L−1 was constructed. The final results were expressed as mM/g DW Trolox Equivalent (TE). The antioxidant activity of the freeze-dried colorants was calculated using the following equation:
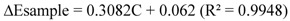 |
where ∆E sample is the difference in absorbance of the samples through inhibition/decolorization of the ABTS●+ radical scavenging activity and C is the concentration (mM/g·DW·TE).
SOD-like activity was measured according to the previously described method by Debnath et al. [27]. The percentage of SOD-like antioxidant activity was calculated as follow:
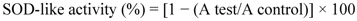 |
The total phenolic content of the IOSC extract was determined using the Folin-Ciocalteu method described by Mohdaly et al. [25]. Gallic acid was used as the standard for the calibration curve. Total phenolic content was expressed as gallic acid equivalent (GAE), calculated by plotting against the standard calibration curve using the following linear equation:
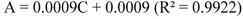 |
where A is the absorbance and C is the concentration (mg·GAE/g·DW).
The total flavonoids content of the IOSC extract was determined using methods previously described by Mohdaly et al. [25]. Freshly prepared quercetin was used as standard. The total flavonoids content was then further expressed as mg quercetin equivalent (QE) per gram dry weight (mg·QE/g·DW) and was calculated according to the following linear equation based on the calibration curve:
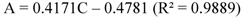 |
where A is the absorbance and C is the concentration (mg QE/g DW).
The concentration of total flavonols was measured according to the method of Mohdaly et al. [25]. The total flavonols were calculated based on the regression equation between quercetin standard and absorbance:
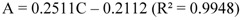 |
where A is the absorbance and C is the concentration (mg·QE/g·DW).
Quantification of the RA content was calculated based on a spectrophotometric method described by Kiong et al. [28].
2.2.12. HPLC Phenolic acids in IOSC Extract
Powdered samples were weighed (20 mg accurately) and extracted within 15 min using 750 µL 70% methanol (v/v, pH 4.0, phosphoric acid) in an ultrasonic water bath with ice. The samples were centrifuged for 5 min at 6,000 rpm. The supernatants were collected and the pellets were re-extracted twice with 500 µL 70% methanol. As internal standard 40 µL of 1 mM cinnamic acid was added to the first extraction step (the recovery rate of cinnamic acid was above 90%). The combined supernatants from each sample were kept in a rotary evaporator (SPD 111V Speed Vac. Concentrator, Thermo Scientific, USA; CVC 3,000V, Vacuubrand GmbH, Wertheim, Germany) at 25 °C under vacuum to remove the solvent completely. The residues were re-dissolved in 1 mL MilliQ water (HPLC grade). The samples were filtered using a 0.22 µm filter (SPIN-X centrifuge tube filter) and then analyzed with HPLC.
The separation of phenolic compounds was performed by HPLC (UltiMate SR-3000, Dionex, Idstein, Germany), equipped with LPG-3400SD pump, WPS-3000SL automated sample injector, AcclaimPA C16-column (3 lm, 2.1 9 150 mm, Dionex) and DAD-3000 diode array detector (Dionex) and software Chromeleon 6.8. The column was operated at a temperature of 35 °C. The mobile phase consisted of 0.1% (v/v) phosphoric acid in water (eluent A) and of 40% (v/v) acetonitrile (eluent B). A multistep gradient was used for all separations with an initial injection volume of 40 µL and a flow rate of 0.4 mL·min−1. The multistep gradient was used as follows: 0–1 min: 0.5% (v/v) B; 1–10 min: 0.5–40% B; 10–12 min: 40% B; 12–18 min: 40–80% B; 18–20 min: 80% B; 20–24 min: 80–99% B; 24–30 min: 99–100% B; 30–34 min: 100–0.5% B; 34–39 min: 0.5% B. Simultaneous monitoring was performed at 290, 330 and 254 nm. Diode array detection was used for the identification of the compounds. Retention times and UV/visible absorption spectra of the peaks were compared with those of the authentic standards.
2.3. Statistical Analysis
Experimental results in this study are reported as means ± standard deviation (S.D.) of five replicates. The data were subjected to analysis of variance (ANOVA) and post-hoc Tukey’s Honestly Significant Different (HSD) tests. The level of statistical significance was set at p-values <0.05 using SPSS version 16.0 (SPSS Inc. Chicago, IL, USA). Correlation analyses between the parameters were carried out using the “ProcCorr” procedure in SAS (2002-10) for Windows, version 9.3 (SAS Institute, Cary, NC, USA).
3. Results and Discussion
3.1. Selection of High RA Line and Antioxidant Properties of IOSCs by Elicitation for the LAB Fermentation Study
The main difference between in vitro sprout culture and in vitro tissue culture of O. aristatus is the aggregate state of the nutrient medium. The nutrient medium of the sprout culture, regardless of nutrient content and sucrose concentration on it, is always liquid. Another difference in sprout culture is that it is always agitated. Further information about in vitro sprout culture was explained by Sevchenko et al. [5]. Preliminary research showed that Orthosiphon aristatus in in vitro sprout culture was capable of developing faster and had higher biomass compared to that of in vitro tissue culture which is important in producing tea leaves. Data of two-week IOSC behavior subjected to elicitation are depicted in Table 1.
Table 1.
Data of two-week IOSC behavior subjected to elicitation.
| Sample group | IOSC Length | Number of leaves | Number of roots | Fresh weight (FW) (g) |
|---|---|---|---|---|
| Control | 5.02 ± 0.58 | 9.40 ± 2.88 | 4.40 ± 1.14 | 6.29 ± 1.44 |
| JA | 3.88 ± 0.94 | 5.80 ± 1.30 | 3.80 ± 2.17 | 4.23 ± 0.86 |
| YE | 3.10 ± 0.80 | 5.20 ± 1.30 | 3.60 ± 1.95 | 5.40 ± 1.18 |
Data reported as average ± SD with n = 5.
Control = MS medium; JA = MS medium with jasmonic acid; YE = MS medium with yeast extract.
3.1.1. Antioxidant Activity of IOSC after Elicitation
In response to elicitation treatment, an increase in the percentage of antioxidant activity was noticed for all samples compared to the control (Figure 2). Yeast extract elicitation resulted in the highest percentage of DPPH scavenging activity (32.21 ± 0.69% for the 70% methanol extract as seen in Figure 2(a). Similarly, the highest TEAC antioxidant activity was recorded for IOSC samples induced with yeast extract elicitor and corresponded to 1.51; 1.99; and 1.65 mM/g TE for the IOSC; WEO, MEO and EEO respectively (Figure 2(b)). Furthermore, in terms of SOD-like activity attained (Figure 2(c)), elicitation showed a significant increase in the percentage of SOD-like antioxidant activity. Yeast extract elicited IOSC with 70% methanol as solvent had the highest SOD-like activity among all treatments.
Figure 2.

Antioxidant activity of the IOSC subjected to elicitation using three different solvents (a) DPPH scavenging activity; (b) TEAC antioxidant activity; and (c) SOD-like activity.
3.1.2. Antioxidant Components of IOSC after Elicitation
In accordance with the results of antioxidant activity, the total phenolics, flavonoids, flavonols, and RA concentration responded positively to elicitation with increases in content, suggesting that the elicitors stimulated the synthesis of these secondary metabolites (Figure 3). Among all the samples, yeast extract IOSC had the highest amount of total phenolic compounds, flavonoids, flavonols, and RA content for all three tested solvents. The total phenolics from the ethanol extraction of IOSC with yeast extract used as elicitation were two fold (324.56 ± 3.73 mg·GAE/g·DW) compared to the IOSC control (151.22 ± 2.41 mg GAE/g DW) (Figure 3(a)). A similar pattern was also observed for total flavonoids, flavonols, and RA level from IOSC with yeast extract elicitation, which had the highest amount (Figure 3(b–d)). The total amount of flavonoids was 2.96; 4.67 and 3.95 mg·QE/g·DW and total flavonols was 2.47; 3.85 and 3.42 mg·QE/g·DW for WEO, MEO and EEO respectively as illustrated in Figure 3(c,d). These results indicated that the increase in antioxidant activity was in conjunction with increases in antioxidant components in the elicited samples.
Figure 3.

Antioxidant components of the IOSC subjected to elicitation using three different solvents for extraction (a) total phenolics (Phe); (b) flavonoids (FD); (c). flavonols (FL); (d) RA.
3.1.3. Solvent for Extraction
In response to the different solvents used for extraction, 70% methanol exhibited the highest yield of total phenolics, flavonoids, flavonols and RA. It consequently resulted in the strongest antioxidant activity. This finding corresponded with previous research on plant extraction carried out by Lee et al. [29] and Sun et al. [13], where they investigated the influence of different solvents on extraction. They found that the methanol extract showed the highest yield of phenolic compounds and the strongest antioxidant activity. A similar result was also reported by Azlim Almey et al. [30] confirming methanol as the most efficient solvent for the extraction of phenolic compounds in comparison to ethanol and water. They further explained that the highest yield of phenolics in the methanol extract was due to the ability of methanol to inhibit polyphenol oxidase (PPO) that causes oxidation of phenolics and lessens its evaporation compared to ethanol and water.
3.1.4. HPLC Phenolic Acids of IOSC after Elicitation
From the HPLC analysis of phenolic acids as shown in Figure 4(a), the elicitation confirmed a significant increase in RA content compared to the control. The basal level of RA content in the control sample was 2.89 ± 0.86 mg/g·DW (Figure 4(b)), meanwhile in IOSC samples elicited with JA it was 3.75-fold higher (10.83 ± 0.27 mg/g·DW). Nevertheless, the highest elicitation effect was seen in IOSC samples treated with yeast extract resulting in 4.95-fold RA content (14.28 ± 0.64 mg/g·DW) in comparison to that of the control. These results showed that jasmonic acid and the yeast extract of the elicitors were very effective in inducing the production of RA in IOSC. These results were consistent with those previously described by Ogata et al. [31] who found that the addition of yeast extract and methyl-jasmonate rapidly enhanced the accumulation of RA in the culture of Lithosermum erythrorhizon.
Figure 4.

(a) Typical HPLC chromatogram of IOSC extract subjected to elicitation: 1: vanillic acid; 2: chlorogenic acid; 3: caffeic acid; 4: p-coumaric acid; 5: sinapic acid; 6: rosmarinic acid; and 7: cinnamic acid as internal standard (peak identification based on standard compounds); and (b) HPLC-RA quantification of IOSC after elicitation.
Moreover, Kim et al. [32] reported that the stimulation of RA biosynthesis in Agastache rugosa O. Kuntze was very successful in response to the addition of yeast extract which could raise RA content up to 5.7-fold compared to the non-elicited suspension cells. Previous reports suggested that the increase of RA content by yeast extract treatment coincided with the increase in phenylalanine ammonia-lyase (PAL) activity, indicating the important role of PAL activity in the regulation of RA biosynthesis [7,33]. The increase of the level of RA can be explained by the fact that IOSCs, like other plants, carry out an elicitor-mediated defense response by inducing phenylpropanoid pathways resulting in production of more RA. As a result, yeast extract elicitor was found to be the best strategy for RA production in comparison to jasmonic acid as equally reported in other studies [7,33]. Therefore, IOSC with yeast extract elicitor was used to study the effect of LAB fermentation in IOSC.
In addition, the responses of other phenolic acids are provided in Table 2. Yeast extract elicitation, however, caused an increased in vanillic, caffeic acid and p-coumaric acid by 3.72, 3.46 and 3.50 fold of the control, respectively. In other plant species, Mallus domestica cell culture, p-coumaric acid and chlorogenic acid were more responsive to the yeast extract elicitation and enhanced more dramatically with 5.1 fold and 2.7 fold of that of the control level [6]. These observations indicated that yeast extract elicitation also enhanced the production of other phenolic acids and strengthened the choice of yeast extract-IOSC for the LAB fermentation study.
Table 2.
Individual response of phenolic acids subjected to elicitation.
| Other phenolic acids | Control(mg/g·DW) | Fold of control | |
|---|---|---|---|
| JA | YE | ||
| Vanillic acid | 2.28 ± 0.27 | 2.80 | 3.72 |
| Chlorogenic acid | 0.49 ± 0.07 | 1.10 | 1.90 |
| Caffeic acid | 4.62 ± 0.15 | 1.30 | 3.46 |
| p-Coumaric acid | 0.27 ± 0.03 | 2.40 | 3.50 |
| Sinapic acid | 1.12 ± 0.04 | 2.5 | 1.82 |
Control = MS medium; JA = MS medium with jasmonic acid; YE = MS medium with yeast extract.
3.2. LAB Fermentation Study of Selected IOSC
To the best of our knowledge, this is one of the first studies using in vitro plants as a media for LAB fermentation focusing on rosmarinic acid and related antioxidant properties.
3.2.1. The Effects of IOSC Extract on Viable Cell LAB Growth
There are several methods to determine LAB growth rates. Plate count is by far the most reliable method to establish growth curve, but it is extremely laborious. An alternative is the measurement of optical density (OD). OD measurement is adequate for modeling the growth of food-fermenting organisms, where the emphasis is exclusively on conditions that allow growth to a high population density. However, it is not reliable enough when it is use to assess the growth of food pathogens or spoilage organisms [34,35]. Therefore, in this fermentation study, the antibacterial effect of selected extracts on the growth of LAB strains was further evaluated by determining their growth density OD620.
In general, based on absorption value increase measured at 620 nm, the added IOSC extract resulted in insignificant difference (p > 0.05) compared to the growth density of the pure culture (Figure 5). These results mean that the IOSC extract at the concentration 1.5 mg/mL used had no inhibitory effect on the growth of LAB strains used in this present study. The only report found on the O. aristatus terrestrial plant showed that the aqueous extract of O. Aristatus demonstrated antibacterial activity against two serotypes of Streptococcus mutans with minimal inhibition concentration (MIC) of 7.8–23.4 mg/mL [36] which was much higher than 1.5 mg/mL. These results were in accordance with the previous investigation by Puupponen-Pimiä et al. [37] who studied the antimicrobial effect of the extract of several berries using 70% acetone as solvent on the growth of LAB strains (including L. plantarum). They reported that the berry extract at low concentration of 1 mg/mL had no antimicrobial effect on the LAB strains used. A similar finding was recently reported by He et al. [21] that the Codonopsis lanceolata water extract (0.25 g/mL) provided a good substrate for LAB growth.
Figure 5.

Growth density of Lactobacillus plantarum ATCC 8014 and Lactobacillus acidophilus NCFM with/without IOSC extract.
3.2.2. Viable Cell Growth of SSF and LSF
The growth of LAB in line with the production of organic acids as well as other possible antimicrobial substances restricted the growth of aerobic heterotrophic bacteria [11]. The growth of LAB in SSF samples remained unchanged for 24 h samples for both LP and LA inoculated SSF. The total viable count of inoculated LAB strains OD620 0.1 was 3.2 × 105 cfu/g for L. plantarum and 5.8 × 105 cfu/g for L. acidophilus and after 24 h fermentation for SSF samples, the viable cells of LAB were 3.2 and 8.1 × 105 cfu/g for SSF-LA and LP, respectively (Table 3(a)). A one log decrease to 5.2 × 104 cfu/g was observed after SSF treatment with LP for 48 h. A similar trend was observed with SSF LA-48. By the end of fermentation, the total viable cells remained unchanged at 104 cfu/g for both SSF samples.
Table 3(a).
Total LAB of SSF.
| Strains | Time (h) | Total LAB * |
|---|---|---|
| L. plantarum | Inoculated OD620 0.1 = 3.2 × 105 cfu/g | |
| 24 | 8.1 × 105 cfu/g | |
| 48 | 5.2 × 104 cfu/g | |
| 72 | 2.5 × 104 cfu/g | |
| L. acidophilus | Inoculated OD620 0.1 = 5.8 × 105 cfu/g | |
| 24 | 3.2 × 105 cfu/g | |
| 48 | 1.8 × 104 cfu/g | |
| 72 | 3.0 × 104 cfu/g | |
* Values are results of at least two independent experiments.
Meanwhile, the changing of the viable cell of LAB strains in LSF after 24, 48 and 72 h was evaluated by OD620 determination. In general, the addition of sterile IOSC (1.5% g/v) in growth density of LAB strains (L. plantarum and L. acidophilus) in a DCU fermenter resulted in an insignificant difference compared to the control without IOSC (Table 3(b)).
Table 3(b).
The growth density OD620 of LSF.
| MRS broths | Time (h) | OD620 |
|---|---|---|
| L. plantarum (without IOSC; control) | 24 | 2.221 ± 0.010 |
| 48 | 1.661 ± 0.009 | |
| 72 | 1.777 ± 0.013 | |
| L. plantarum with IOSC | 24 | 2.225 ± 0.021 |
| 48 | 1.664 ± 0.011 | |
| 72 | 1.871 ± 0.011 | |
| L. acidophilus (without IOSC; control) | 24 | 2.151 ± 0.008 |
| 48 | 1.558 ± 0.015 | |
| 72 | 1.831 ± 0.007 | |
| L. acidophilus with IOSC | 24 | 2.226 ± 0.009 |
| 48 | 1.560 ± 0.012 | |
| 72 | 1.839 ± 0.014 |
3.2.3. Solid State Fermentation (SSF) of IOSC
Figure 6(a) illustrates the percentage DPPH scavenging activity of SSF-IOSC (unfermented (control) and fermented) using three different solvents. As is shown, SSF significantly influences the DPPH radical scavenging activity of IOSC. Unlike most of the latest published reports on LAB plant fermentation studies [16,1,2,3,4,5,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21] which resulted in an increase in antioxidant activity, SSF with LAB caused a weakening percentage of DPPH radical scavenging activity of IOSC compared to the control for all three solvents tested. The longer period for SSF caused more losses in the DPPH radical scavenging activity. The percentage for 30 min tested DPPH radical scavenging activity of UF water extract was 25.98 ± 0.23% which decreased considerablyy to the level of 10.41 ± 0.21% after 72 h SSF treatment inoculated with LA. A higher decrease was observed in SSF with LP inoculation and the remaining percentage of DPPH radical scavenging activity was only 7.79 ± 0.23% after a 72 h period of fermentation.
Figure 6.

Antioxidant activity of IOSC subjected to SSF using three different solvents (a) DPPH scavenging activity; (b) TEAC antioxidant activity; and (c) SOD-like activity.
In comparison to the DPPH free radical scavenging method, the TEAC antioxidant activity method was also applied. A similar trend was observed (Figure 6(b)), a significant reduction (p < 0.05) of TEAC antioxidant activity was noted as a response to SSF treatment either LP or LA inoculation (Figure 6(b)). The rate of TEAC value reduction after 72 h SSF fermentation with LA is as follows: 50.28 ± 1.56%, 45.59 ± 2.02% and 43.47 ± 0.37% for water, 70% methanol and 80% ethanol extract, respectively. A higher reduction rate of the TEAC value was obtained in SSF for 72 h with LP inoculation, 59.63 ± 3.00%, 54.65 ± 7.83% and 54.63 ± 4.08% for water, 70% methanol and 80% ethanol extract, respectively.
In terms of the SOD-like activity results attained, LAB fermentation caused significant depletion (p < 0.05) of its activity (Figure 6(c)). Similarly, the diminishing result of antioxidant activity using SOD-like methods in Vigna sinensis var. carilla due to natural fermentation was also reported by Doblado et al. [38] and this decreasing trend of SOD-like activity was confirmed with our result.
The losses of antioxidant activity of LAB fermented SSF can be linked to the sharp decline in the total phenolics and RA during SSF. The data are presented in Figure 7.
Figure 7.

Antioxidant components of IOSC subjected to SSF using three different solvents for extraction (a) total phenolics (Phe); (b) flavonoids (FD); (c) flavonols (FL); (d) RA.
The knowledge that phenolic degradation is mainly caused by polyphenol oxidase (PPO) is well known. This phenomenon clearly indicated that the degradation of phenolic compounds occurred during the SSF process by an enzymatic process involving PPO. The obtained results were in agreement with a previous study on L. plantarum inoculation of potherb mustard (Brassica juncea Coss.) pickling fermentation by Fang et al. [39] that resulted in a decrease in antioxidant activity associated with the reduction of total phenolics during fermentation. Here it was further explained that the decrease of total phenolics indicated that the phenolics degradation by enzymatic and non-enzymatic processes occurred during fermentation. A similar declining antioxidant properties trend was also reported on the fermentation study of Chetoui olives (black, green and varicolored olive) using L. plantarum with spontaneous fermentation by Othman et al. [40]. They reported that the antioxidant activity of Chetoui olives after L. plantarum and spontaneous fermentation decreased by 50–72%. In our study, the drastic decrease of antioxidant activity was connected with an important loss of total phenolics and rosmarinic acid.
Figure 7(b) shows the effect of LAB SSF on total flavonoids and flavonols of IOSC. The flavonoid and flavonol content in response to L. plantarum SSF showed a modest decrease after 24 h fermentation for all solvents used for extraction. A higher rate of depletion in total flavonoids and flavonols occurred after 24 h of L. acidophilus SSF. Interestingly, both LAB SSFs gradually increased for longer fermentation (48 and 72 h). An increase in total flavonoids including flavonols during plant fermentation has been reported by many researchers [13,17,18,20,41,42,43]. Cho et al. [44] showed that the increase in flavonols in fermented Soybean by Bacillus pumilus HY1 was the result of the esterase and tannase activities of LAB during fermentation. This phenomenon indicated a possible conversion of fermentation-stimulated metabolites. These results suggested that some flavonoids could be degraded during fermentation and/or may be produced from the degradation of phenolic compounds [45].
In response to the different solvents used for extraction, similar findings were in agreement with the elicitation result treatment. Results showed that 70% methanol, regardless of LAB fermentation, produced the highest yield of total phenolics, flavonoids, flavonols and RA and consequently exhibited the strongest antioxidant activity (DPPH, TEAC and SOD-like activity). This finding corresponded with previous plant fermentation studies by Lee et al. [29] and Sun et al. [13] employing different solvents for extractions of which the methanol extract showed the highest yield of phenolic compounds as well as the strongest antioxidant activity.
3.2.4. Liquid State Fermentation (LSF) of IOSC
In contrast to the SSF method which utilized 5% of LAB starters, a further change in antioxidant properties of IOSC was observed in LSF predominated by LAB starters in MRS medium. The data are shown in Figure 8. Many studies have shown that the utilization of LAB in plant fermentations can increase antioxidant properties [16,19], however, such an effect was not observed in the LAB fermentation of IOSC as examined in this study. There was a remarkable decrease in the detectable antioxidant activity (DPPH, TEAC, and SOD-like activity Figure 8 (a,b,c respectively)) during the LSF process for 24, 48 and 72 h regardless of the starter organisms employed for fermentation.
Figure 8.

Antioxidant activity of IOSC subjected to LSF using three different solvents (a) DPPH scavenging activity; (b) TEAC antioxidant activity; and (c) SOD-like activity.
In general, a higher rate of reduction in antioxidant activity was observed in LSF than in SSF and as observed in SSF the L. plantarum fermentation caused greater losses of antioxidant activity in comparison to L. acidophilus. This lessening antioxidant activity of plant extracts by fermentation has been reported by several investigators [46,47,48,49]. Natural fermentation caused a reduction of TEAC value of 23% in Lupinus albus var. multolupa [47], 4% in Cajanus cajan [48], and 9–29% in soybean [49], while that with A. oryzae or L. plantarum in Lupinus angustifolius cv. Zapaton resulted in a decrease of 18% and 24% of the TEAC values, respectively [46]. Inoculated fermentation with A. oryzae, R. oryzae, or L. plantarum in soybean caused a decrease of TEAC by 40, 26, and 9–13%, respectively [49]. Similarly, Chang et al. [50] reported that the loss in the antioxidant activity was caused by the microbial activity during fermentation which was confirmed by our result.
In terms of decreasing SOD-like activity obtained due to LSF in Figure 8(c), the result was inconsistent with the report by Fernandez-Orozco et al. [46] on Lupinus angustifolius cv. Zapaton, fermented by using A. oryzae, R. oryzae, or B. subtilis where the fermentation process resulted in a drastic decrease (98–100%) of SOD-like activity. A large reduction in SOD-like activitywas also observed in fermented soybean [49]. Additionally, using a different antioxidant activity method of Peroxyl Radical-Trapping Capacity (PRTC), Doblado et al. [38] reported that the application of liquid fermentation on Vigna sinensis var. carilla caused the reduction of PRTC antioxidant activity.
The gradually weakening antioxidant activity corresponded to the sharp depletion in antioxidant components (total phenolics and RA). Figure 9 shows the components of the antioxidant levels of control (unfermented) and LSF of IOSC using LAB. The total phenolics decreased considerably in both LSF inoculated with L. acidophilus and L. plantarum (Figure 9(a,b)), and a lower level of these antioxidant components was obtained in LP-LSF samples compared to LA-LSF samples. Concerning the effect of fermentation on RA level, a similar reduction trend was also found (Figure 9(d)).
Figure 9.

Antioxidant components of IOSC subjected to LSF using three different solvents for extraction (a) total phenolics (Phe); (b) flavonoids (FD); (c) flavonols (FL); (d) RA.
This result was in agreement with the previous report by Frias et al. [47] who reported a decrease in antioxidant components of Lupinus albus var. multolupa carried out by natural fermentation or inoculated with L. plantarum. A similar observation was also reported by Landete et al. [51] who showed that the growth of L. plantarum led to the depolymerization of high-molecular-weight phenolic compounds, with a resultant decolorization of the fresh olive mill wastewater and a significant reduction in total phenolics. A decrease in the total phenolics with LSF as reported here showed that phenolics were degraded and utilized by fermented microorganisms as reported by other researchers [52,53]. Moreover, in terms of decreasing flavonoids by fermentation, it has been reported that flavonoids glycosides were metabolized within a 72 h fermentation period using an in vitro fermentation human fecal micro-flora system [54].
Interestingly, as found in SSF, an increase in total flavonols in LSF was also observed (Figure 9(c)). Fermentation led to increased total flavonols reported by Cho et al. [18] and Cho et al [44]. They explained that the esterase activity and total flavonols increased significantly with a corresponding decrease in flavonol gallates during the fermentation [44].
Turning to the solvent used for extraction, a similar pattern to elicitation and SSF in that the highest antioxidant activity and components were observed in the LSF-IOSC extract with methanol, while the lowest levels were found in a water extract. The results supported and confirmed that 70% methanol was also the most suitable for extraction of antioxidant component, bringing about a higher antioxidant activity, owing to its high polarity and good solubility for both flavanoids and components of the plant materials.
3.2.5. HPLC Analysis of IOSC after LAB Fermentation
Individual phenolic compounds were analyzed by HPLC to further examine the effect of LAB fermentation on IOSC and in particular RA as the compound of interest in this study. The determination of the phenolic profile in plants, in particular fermented plants is difficult. The reason being that the majority of the reference compounds are not commercially available and in order to carry out quantitative analysis of phenolics in plant extracts, the hydrolysis procedure, particularly the acid hydrolysis has been used very often to simplify the analysis and thus reduce the number of derivatives [55]. In our study, hydrolysis of phenolic acids occurred by the addition of acid to methanol for extraction and the phenolic profiles of IOSC were determined using the HPLC-DAD method proceeded by simultaneous monitoring at 290, 330 and 254 nm. The typical HPLC phenolic acids chromatograms of IOSC extract are presented in Figure 10.
Figure 10.

Typical HPLC chromatogram of IOSC extract subjected toLAB fermentation: 1. vanillic acid; 2. chlorogenic acid; 3. caffeic acid; 4. p-coumaric acid; 5. sinapic acid; 6. rosmarinic acid; and 7. cinnamic acid as internal standard (peak identification based on standard compounds).
Several identified phenolic acids, vanillic, chlorogenic, caffeic, p-coumaric, sinapic and RA, were eluted in the chromatograms (Figure 10). IOSCs were found to accumulate RA as the major phenolic of all identified phenolic constituents by HPLC analysis followed by caffeic acid as the second major phenolic acid in IOSC. This finding was in agreement with the report on the analysis of this plant by Sumaryano et al. [56] who equally identified RA and caffeic acid as the major phenolic constituents in the methanol extract. There are several major unknown peaks predicted to be degradation products of LAB fermentation. The changing phenolic acids subjected to LAB fermentation, SSF and LSF, are depicted in Table 4.
Table 4.
Individual response of phenolic acids to the SSF and LSF LAB fermentation of IOSC for 48 h.
| Phenolic acids | Control (un-fermented) | % decrease L. plantarum ATCC 8014 | % decrease L. acidophilus NCFM | ||
|---|---|---|---|---|---|
| (mg/g·DW) | SSF | LSF | SSF | LSF | |
| Rosmarinic acid | 14.28 ± 0.64 | 55.7 | 72.3 | 46.8 | 56.2 |
| vanillic acid | 8.48 ± 0.06 | 23.5 | 43.2 | 20.4 | 24.2 |
| chlorogenic acid | 0.93 ± 0.11 | 27.6 | 40.1 | 23.6 | 35.6 |
| caffeic acid | 9.06 ± 0.33 | 42.4 | 49.5 | 37.3 | 44.9 |
| p-coumaric acid | 0.95 ± 0.07 | 29.6 | 37.6 | 28.6 | 32.8 |
| sinapic acid | 2.04 ± 0.03 | 26.8 | 33.9 | 21.6 | 29.8 |
As observed in our HPLC results, both fermentations, SSF and LSF, caused a significant decrease of individual specific phenolic acids which has also been reported by several researchers [52,53,57,58]. Hegde, et al. [53] reported the drastic decrease in feruclic, coumaric and syringic acids of wheat and rice brans respectively after A. niger fermentation at 96 h, while gallic and protocatechuic acids were completely degraded in 96 h. Degradation of these phenolic acids clearly indicated the induction of various phenolic acid esterases. It was associated perhaps with the oxidation of those phenolic acids or even utilized by fermented microbes for various end uses [53]. Similarly, the concentration of the hydroxybenzoic acids fell slightly at the end of alcoholic fermentation while the hydroxycinnamic acids (caffeic, coumaric and ferulic acids) decreased due to dilution effects in wine fermentation [57]. Also, the content of 4-hydroxybenzoic and syringic acid decreased five- and two-fold respectively in palm oil fermented with A. niger [52]. The degradation and formation of phenolic acids clearly indicates the action of various enzymes especially the esterases and cell-wall degrading enzymes secreted by the fermented microorganism [52].
Moreover, the S. Cerevisiae fermentation of Rubus coreanus Miq. using different fractions; the soluble phenolic fraction (Fr. A), the soluble ester-type phenolic fraction (Fr. B) and the insoluble ester-type phenolic fraction (Fr. C), resulted in a decrease of gallic and ferulic acid in Fr. A and of salicylic, caffeic, and cinnamic acid contents in Fr. C [58]. These decreased phenolic acid contents could be attributed to either microbial oxidation, reduction, or the degradation of the phenolic compounds by the fermenting microbes [58]. In wine fermentation, it has been suggested that the decrease in the concentration of trans-caftaric and trans-p-coutaric acids until disappearance could be linked to lactic acid bacteria metabolism [59].
It has been generally known that L. plantarum has the ability to degrade many types of phenolic acids [60]. From the hydroxycinnamic group, L. plantarum is able to metabolize p-coumaric, caffeic, ferulic, and m-coumaric acid and from hydroxybenzoic acids both gallic and protocatechuic acid. Decarboxylation and reduction of the phenolic acid reactions were involved in their metabolism. The formation of ethyl derivatives from the degradation of the hydroxycinnamic group and from some compounds like pyrogallol and catechol, which might be beneficial for the growth and metabolism of this bacterium, were detected [60]. Another type of LAB, L. brevis was shown to degrade p-coumaric acid and caffeic acids via decarboxylation and their vinyl derivatives were detected [61]. However, there was no report on the metabolism of phenolic acids in L. acidophilus. Unfortunately, with our HPLC methods, the degradation products could not be identified. The above mentioned fermentation reactions can significantly influence food quality through the development and degradation of polyphenols. Therefore, further work on the identification of the degradation products and enzymes involved is required to improve our understanding of phenolic acid degradation by LAB.
As for the effect of LAB fermentation on RA in IOSC leaves, the HPLC analysis of phenolic acids confirmed that RA was predominant in soluble phenolics and was sharply reduced in both SSF and LSF compared to UFT (unfermented). LSF suffered a higher rate of decreasing RA content than SSF. In terms of type LAB inoculums, L. plantarum caused a more dramatic degradation compared to L. acidophilus. The relatively high temperature of 37 °C for LAB growth was predicted as contributing to the great losses of RA. This finding was in agreement with the report by Fletcher et al. [62] who showed that heat treatment at only 30 °C could significantly reduce the content of RA and phenolic compounds. Similarly, tea fermentation of C. asiatica caused a reduction in RA [63]. The RA level decreased significantly from 282 ± 42 µg/g in the unfermented to 158 ± 40 µg/g in partially fermented C. asiatica leaves and there was a further reduction to 58 ± 1 µg/g in the fully fermented C. asiatica tea leaves [63].
The fact both LAB fermentations SSF and LSF inoculated with L. plantarum and L. acidophilus using sterilized-in vitro sprout culture of Orthosiphon aristatus as model system had a lower level of phenolics acids measured by HPLC analysis, clearly supports and confirms the result of spectrophotometric measurement that fermentation not only depromotes the antioxidant activity with all mechanisms of antioxidant measurements, but also is accompanied by a notable decrease of antioxidant components.
3.2.6. Correlation Analysis between the Experimental Variables
In regard of acquiring a holistic approach to the IOSC antioxidant properties and respective of their partial chemical constituents, a Pearson correlation matrix between all these variables was performed. It has been suggested that the antioxidant activity of plant extracts is mainly due to phenolic compounds in the plant extract [25], although Azlim Almey et al. [30] found only a weak correlation. Additionally, antioxidant potency of the extract is not proportional to the total phenolic content, suggesting that the antioxidant activity is not simply a function of phenolic concentrations [64].
However, from our experimental results, a positive correlation was found. In response to elicitation, an increasing antioxidant activity was also followed by an increase in total amount of phenolics and RA and the reverse was true for the LAB fermentation results. It has been reported that not all antioxidant components have the same antioxidant activity, and gallic acid and rosmarinic acid were found to be the most potent antioxidants among the simple phenolic and hydroxycinnamic acids, respectively [65]. The highest correlation was found between TEAC and RA content (r = 0.92). This correlation result suggests that the RA content in the IOSC extract has a high contribution and is more likely to be responsible for the antioxidant activity of IOSC extract for both elicitation or LAB fermentation treatments as was reported by Tepe [66]. It was found that RA was as expected topmost on the antioxidant activity of several Salvia plants. Additionally, Park et al. [1] highlighted that RA had a stronger antioxidant activity than that of vitamin E.
4. Conclusions
To conclude, from the selection of the high RA line, it was proven that the addition of yeast extract elicitor to the IOSC enhanced RA and the synthesis of phenolic compounds resulting in relatively higher antioxidant activity. A 70% methanol extract exhibited the highest extraction ability for antioxidant activity assays, total phenolics, flavonoids and flavonols. HPLC analysis of phenolic acids clearly confirmed a drastic increase in RA subjected to yeast extract elicitation. Therefore, this yeast extract elicited line of IOSC was used for the LAB fermentation study.
From the LAB fermentation study, the results clearly demonstrate that both types of fermentation, solid and liquid state fermentation, are deleterious to antioxidant properties and counterproductive for the retention of a high level of RA from harvested IOSC. The loss of antioxidant properties was more pronounced in the liquid state fermentation compared to the solid state one. Longer fermentation resulted in a greater reduction of the antioxidant properties. Results also reveal that the fermentation with L. plantarum resulted in the highest reduction rate of antioxidant properties. HPLC analysis confirmed that LAB fermentation of IOSC caused a decreased level of phenolic acids. Lastly, correlation analysis indicated that RA is the major contributor to the antioxidant activity of this plant.
Acknowledgments
Financial support of this project from the Institute of Food Technology and Food Chemistry, TU Berlin and TU Berlin and by a DAAD Scholarship to the first author is gratefully acknowledged. The authors give many thanks to other colleagues from the Berlin University of Technology for their technical assistance. The authors also thank Istanti Hermagustiana from the English Department of the University of Mulawarman, East Borneo, Indonesia for linguistic-help and proof reading of the manuscript.
Supplementary Files
References
- 1.Park S.U., Uddin M.R., Xu H., Kim Y.K., Lee S.Y. Biotechnological applications for rosmarinic acid production in plant. Afr. J. Biotechnol. 2008;7:4959–4965. [Google Scholar]
- 2.Petersen M., Simmonds M.S.J. Rosmarinic acid. Phytochemistry. 2003;62:121–125. doi: 10.1016/s0031-9422(02)00513-7. [DOI] [PubMed] [Google Scholar]
- 3.Akowuah G.A., Ismail Z., Norhayati I., Sadikun A. The effects of different extraction solvents of varying polarities on polyphenols of Orthosiphon stamineus and evaluation of the free radical-scavenging activity. Food Chem. 2005;93:311–317. doi: 10.1016/j.foodchem.2004.09.028. [DOI] [Google Scholar]
- 4.Fadhlina A. The Effects of Temperature and Fermentation Time on Selected Cchemical Composition, Antioxidant and Quality of Cat Whiskers (Orthosiphon stamineus) Tea. University Sains Malaysia; Pulau Pinang, Malaysia: 2008. [Google Scholar]
- 5.Shevchenko Y., Smetanska I., Wendt A. Stevia Science. Euprint; Heverlee, Belgium: 2010. Sprout culture of Stevia rebaudiana Bertoni; pp. 5–26. [Google Scholar]
- 6.Cai Z. Effects Elicitors on Secondary Metabolism of Plant Suspension Cultures of Vitis vinifera and Malus domestica and their Exudates. Berlin University of Technology; Berlin, Germany: 2012. [Google Scholar]
- 7.Sumaryono W., Proksch P., Hartmann T., Nimtz M., Wray V. Induction of rosmarinic acid accumulation in cell suspension cultures of Orthosiphon aristatus after treatment with yeast extract. Phytochemistry. 1991;30:3267–3271. doi: 10.1016/0031-9422(91)83190-V. [DOI] [Google Scholar]
- 8.Mewis I., Smetanska I., Müller C., Ulrichs C. Specific poly-phenolic compounds in cell culture of Vitis vinifera L. cv. Gamay Fréaux. Appl. Biochem. Biotechnol. 2011;164:148–161. doi: 10.1007/s12010-010-9122-x. [DOI] [PubMed] [Google Scholar]
- 9.Dordevic T.M., Siler-Marinkovic S.S., Dimitrijevic-Brankovic S.I. Effect of fermentation on antioxidant properties of some cereals and pseudo cereals. Food Chem. 2010;119:957–963. doi: 10.1016/j.foodchem.2009.07.049. [DOI] [Google Scholar]
- 10.Maifreni M., Marino M., Conte L. Lactic acid fermentation of Brassica rapa: Chemical and microbial evaluation of a typical Italian product (brovada) Eur. Food Res. Technol. 2004;218:469–473. doi: 10.1007/s00217-004-0877-6. [DOI] [Google Scholar]
- 11.Katina K., Laitila A., Juvonen R., Liukkonen K.H., Kariluoto S., Piironen V., Landberg R., Aman P., Poutanen K. Bran fermentation as a means to enhance technological properties and bioactivity of rye. Food Microbiol. 2007;24:175–186. doi: 10.1016/j.fm.2006.07.012. [DOI] [PubMed] [Google Scholar]
- 12.Katina K., Liukkonen K.H., Kaukovirta-Norja A., Adlercreutz H., Heinonen S.M., Lampi A.M., Pihlava J.M., Poutanen K. Fermentation-induced changes in the nutritional value of native or germinated rye. J. Cereal Sci. 2007;46:348–355. doi: 10.1016/j.jcs.2007.07.006. [DOI] [Google Scholar]
- 13.Sun Y.-P., Chou C.-C., Yu R.-C. Antioxidant activity of lactic-fermented Chinese cabbage. Food Chem. 2009;115:912–917. doi: 10.1016/j.foodchem.2008.12.097. [DOI] [Google Scholar]
- 14.Zhao D., Ding X. Studies on the low-salt Chinese potherb mustard (Brassica juncea, Coss.) pickle. I-The effect of a homofermentative L(+)-lactic acid producer Bacillus coagulans on starter culture in the low-salt Chinese potherb mustard pickle fermentation. LWT-Food Sci. Technol. 2008;41:474–482. doi: 10.1016/j.lwt.2007.03.023. [DOI] [Google Scholar]
- 15.Liu J.-G., Hou C.-W., Lee S.-Y., Chuang Y., Lin C.-C. Antioxidant effects and UVB protective activity of Spirulina (Arthrospira platensis) products fermented with lactic acid bacteria. Process Biochem. 2011;46:1405–1410. doi: 10.1016/j.procbio.2011.03.010. [DOI] [Google Scholar]
- 16.Martinez-Villaluenga C., Peñas E., Sidro B., Ullate M., Frias J., Vidal-Valverde C. White cabbage fermentation improves ascorbigen content, antioxidant and nitric oxide production inhibitory activity in LPS-induced macrophages. LWT-Food Sci. Technol. 2012;46:77–83. doi: 10.1016/j.lwt.2011.10.023. [DOI] [Google Scholar]
- 17.Cai S., Wang O., Wu W., Zhu S., Zhou F., Ji B., Gao F., Zhang D., Liu J., Cheng Q. Comparative study of the effects of solid-state fermentation with three filamentous fungi on the Total Phenolics Content (TPC), flavonoids, and antioxidant activities of subfractions from Oats (Avena sativa L.) J. Agr. Food Chem. 2011;60:507–513. doi: 10.1021/jf204163a. [DOI] [PubMed] [Google Scholar]
- 18.Cho K.M., Lee J.H., Yun H.D., Ahn B.Y., Kim H., Seo W.T. Changes of phytochemical constituents (isoflavones, flavanols, and phenolic acids) during cheonggukjang soybeans fermentation using potential probiotics Bacillus subtilis CS90. J. Food Compos. Anal. 2011;24:402–410. doi: 10.1016/j.jfca.2010.12.015. [DOI] [Google Scholar]
- 19.Ng C.-C., Wang C.-Y., Wang Y.-P., Tzeng W.-S., Shyu Y.-T. Lactic acid bacterial fermentation on the production of functional antioxidant herbal Anoectochilus formosanus Hayata. J. Biosci. Bioeng. 2011;111:289–293. doi: 10.1016/j.jbiosc.2010.11.011. [DOI] [PubMed] [Google Scholar]
- 20.Wu S.-C., Su Y.-S., Cheng H.-Y. Antioxidant properties of Lactobacillus-fermented and non-fermented Graptopetalum paraguayense E. Walther at different stages of maturity. Food Chem. 2011;129:804–809. doi: 10.1016/j.foodchem.2011.05.025. [DOI] [PubMed] [Google Scholar]
- 21.He X., Zou Y., Yoon W.-B., Park S.-J., Park D.-S., Ahn J. Effects of probiotic fermentation on the enhancement of biological and pharmacological activities of Codonopsis lanceolata extracted by high pressure treatment. J. Biosci. Bioeng. 2011;112:188–193. doi: 10.1016/j.jbiosc.2011.04.003. [DOI] [PubMed] [Google Scholar]
- 22.Wang N.-F., Yan Z., Li C.-Y., Jiang N., Liu H.-J. Antioxidant activity of peanut flour fermented with Lactic Acid Bacteria. J. Food Biochem. 2011;35:1514–1521. doi: 10.1111/j.1745-4514.2010.00473.x. [DOI] [Google Scholar]
- 23.Alberto M.R., Farías M.E., Manca de Nadra M.C. Effect of gallic acid and catechin on Lactobacillus hilgardii 5w growth and metabolism of organic compounds. J. Agr. Food Chem. 2001;49:4359–4363. doi: 10.1021/jf0101915. [DOI] [PubMed] [Google Scholar]
- 24.Sunny-Roberts E.O. Application of Disaccarides Pre-Treatment in Improving Tolerances of Lactobacillus rhamnosus Strains to Environmental Stresses or during Vacuum- and Spray Drying Processes. Berlin University of Technology; Berlin, Germany: 2009. [Google Scholar]
- 25.Mohdaly A.A., Sarhan M.A., Smetanska I., Mahmoud A. Antioxidant properties of various solvent extracts of potato peel, sugar beet pulp and sesame cake. J. Sci. Food Agr. 2010;90:218–226. doi: 10.1002/jsfa.3796. [DOI] [PubMed] [Google Scholar]
- 26.Re R., Pellegrini N., Proteggente A., Pannala A., Yang M., Rice-Evans C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radical Biol. Med. 1999;26:1231–1237. doi: 10.1016/S0891-5849(98)00315-3. [DOI] [PubMed] [Google Scholar]
- 27.Debnath T., Park P.-J., Deb Nath N.C., Samad N.B., Park H.W., Lim B.O. Antioxidant activity of Gardenia jasminoides Ellis fruit extracts. Food Chem. 2011;128:697–703. doi: 10.1016/j.foodchem.2011.03.090. [DOI] [Google Scholar]
- 28.Kiong A.L.P., Lai A.G., Hussein S., Harun A.R. Physiological responses of Orthosiphon stamineus plantles to Gamma Irradiation. AEJSA. 2008;2:135–149. [Google Scholar]
- 29.Lee I.H., Hung Y.-H., Chou C.-C. Solid-state fermentation with fungi to enhance the antioxidative activity, total phenolic and anthocyanin contents of black bean. Int. J. Food Microbiol. 2008;121:150–156. doi: 10.1016/j.ijfoodmicro.2007.09.008. [DOI] [PubMed] [Google Scholar]
- 30.Azlim Almey A.A., Ahmed Jalal Khan C., Syed Zahir I., Mustapha Suleiman K., Aisyah M.R., Kamarul Rahim K. Total phenolic content and primary antioxidant activity of methanolic and ethanolic extracts of aromatic plants’ leaves. Int. Food Res. J. 2010;17:1077–1084. [Google Scholar]
- 31.Ogata A., Tsuruga A., Matsuno M., Mizukami H. Elicitor-induced rosmarinic acid biosynthesis in Lithospermum erythrorhizon cell suspension cultures: Activities of rosmarinic acid synthase and the final two cytochrome P450-catalyzed hydroxylations. Plant Biotechnol. 2004;21:393–396. doi: 10.5511/plantbiotechnology.21.393. [DOI] [Google Scholar]
- 32.Kim H.K., Oh S.-R., Lee H.-K., Huh H. Benzothiadiazole enhances the elicitation of rosmarinic acid production in a suspension culture of Agastache rugosa O. Kuntze. Biotechnol. Lett. 2001;23:55–60. doi: 10.1023/A:1026738409671. [DOI] [Google Scholar]
- 33.Mizukami H., Ogawa T., Ohashi H., Ellis B.E. Induction of rosmarinic acid biosynthesis in Lithospermum erythrorhizon cell suspension cultures by yeast extract. Plant Cell Rep. 1992;11:480–483. doi: 10.1007/BF00232695. [DOI] [PubMed] [Google Scholar]
- 34.Gänzle M.G., Ehmann M., Hammes W.P. Modeling of growth of Lactobacillus sanfranciscensis and Candida milleri in response to process parameters of sourdough fermentation. Appl. Environ. Microbiol. 1998;64:2616–2623. doi: 10.1128/aem.64.7.2616-2623.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Cuppers H.G.A.M., Smelt J.P.P.M. Time to turbidity measurement as a tool for modeling spoilage by Lactobacillus. J. Ind. Microbiol. Biotechnol. 1993;12:168–171. [Google Scholar]
- 36.Chen C.-P., Lin C.-C., Tsuneo N. Screening of Taiwanese crude drugs for antibacterial activity against Streptococcus mutans. J. Ethnopharmacol. 1989;27:285–295. doi: 10.1016/0378-8741(89)90003-2. [DOI] [PubMed] [Google Scholar]
- 37.Puupponen-Pimiä R., Nohynek L., Meier C., Kähkönen M., Heinonen M., Hopia A., Oksman-Caldentey K.M. Antimicrobial properties of phenolic compounds from berries. J. Appl. Microbiol. 2001;90:494–507. doi: 10.1046/j.1365-2672.2001.01271.x. [DOI] [PubMed] [Google Scholar]
- 38.Doblado R., Zielinski H., Piskula M., Kozlowska H., Munoz R., Frias J., Vidal-Valverde C. Effect of processing on the antioxidant vitamins and antioxidant capacity of Vigna sinensis Var. Carilla. J. Agr. Food Chem. 2005;53:1215–1222. doi: 10.1021/jf0492971. [DOI] [PubMed] [Google Scholar]
- 39.Fang Z., Hu Y., Liu D., Chen J., Ye X. Changes of phenolic acids and antioxidant activities during potherb mustard (Brassica juncea, Coss.) pickling. Food Chem. 2008;108:811–817. doi: 10.1016/j.foodchem.2007.11.033. [DOI] [PubMed] [Google Scholar]
- 40.Othman N.B., Roblain D., Chammen N., Thonart P., Hamdi M. Antioxidant phenolic compounds loss during the fermentation of Chétoui olives. Food Chem. 2009;116:662–669. doi: 10.1016/j.foodchem.2009.02.084. [DOI] [Google Scholar]
- 41.Juan M.-Y., Chou C.-C. Enhancement of antioxidant activity, total phenolic and flavonoid content of black soybeans by solid state fermentation with Bacillus subtilis BCRC 14715. Food Microbiol. 2010;27:586–591. doi: 10.1016/j.fm.2009.11.002. [DOI] [PubMed] [Google Scholar]
- 42.Chandrasekara A., Shahidi F. Bioaccessibility and antioxidant potential of millet grain phenolics as affected by simulated in vitro digestion and microbial fermentation. J. Funct. Foods. 2012;4:226–237. doi: 10.1016/j.jff.2011.11.001. [DOI] [Google Scholar]
- 43.Singh H.B., Singh B.N., Singh S.P., Nautiyal C.S. Solid-state cultivation of Trichoderma harzianum NBRI-1055 for modulating natural antioxidants in soybean seed matrix. Bioresource Technol. 2010;101:6444–6453. doi: 10.1016/j.biortech.2010.03.057. [DOI] [PubMed] [Google Scholar]
- 44.Cho K.M., Hong S.Y., Math R.K., Lee J.H., Kambiranda D.M., Kim J.M., Islam S.M.A., Yun M.G., Cho J.J., Lim W.J., Yun H.D. Biotransformation of phenolics (isoflavones, flavanols and phenolic acids) during the fermentation of cheonggukjang by Bacillus pumilus HY1. Food Chem. 2009;114:413–419. doi: 10.1016/j.foodchem.2008.09.056. [DOI] [Google Scholar]
- 45.Rodriguez H., Curiel J.A., Landete J.M., de las Rivas B., de Felipe F.L., Gomez-Cordoves C., Mancheno J.M., Munoz R. Food phenolics and lactic acid bacteria. Int. J. Food Microbiol. 2009;132:79–90. doi: 10.1016/j.ijfoodmicro.2009.03.025. [DOI] [PubMed] [Google Scholar]
- 46.Fernandez-Orozco R., Frias J., Muñoz R., Zielinski H., Piskula M., Kozlowska H., Vidal-Valverde C. Effect of fermentation conditions on the antioxidant compounds and antioxidant capacity of Lupinus angustifolius cv. zapaton. Eur. Food Res. Technol. 2008;227:979–988. doi: 10.1007/s00217-007-0809-3. [DOI] [Google Scholar]
- 47.Frias J., Miranda M.L., Doblado R., Vidal-Valverde C. Effect of germination and fermentation on the antioxidant vitamin content and antioxidant capacity of Lupinus albus L. var. Multolupa. Food Chem. 2005;92:211–220. doi: 10.1016/j.foodchem.2004.06.049. [DOI] [Google Scholar]
- 48.Torres A., Frias J., Granito M., Vidal-Valverde C. Fermented pigeon pea (Cajanus cajan) ingredients in pasta products. J. Agr. Food Chem. 2006;54:6685–6691. doi: 10.1021/jf0606095. [DOI] [PubMed] [Google Scholar]
- 49.Fernandez-Orozco R., Frias J., Muñoz R., Zielinski H., Piskula M.K., Kozlowska H., Vidal-Valverde C. Fermentation as a bio-process to obtain functional soybean flours. J. Agr. Food Chem. 2007;55:8972–8979. doi: 10.1021/jf071823b. [DOI] [PubMed] [Google Scholar]
- 50.Chang C.T., Hsu C.K., Chou S.T., Chen Y.C., Huang F.S., Chung Y.C. Effect of fermentation time on the antioxidant activities of tempeh prepared from fermented soybean using Rhizopus oligosporus. Int. J. Food Sci. Technol. 2009;44:799–806. doi: 10.1111/j.1365-2621.2009.01907.x. [DOI] [Google Scholar]
- 51.Landete J.M., Rodríguez H., De Las Rivas B., Muñoz R. High-added-value antioxidants obtained from the degradation of wine phenolics by Lactobacillus plantarum. J. Food Protect. 2007;70:2670–2675. doi: 10.4315/0362-028x-70.11.2670. [DOI] [PubMed] [Google Scholar]
- 52.Jamal P., Idris Z.M., Alam M.Z. Effects of physicochemical parameters on the production of phenolic acids from palm oil mill effluent under liquid-state fermentation by Aspergillus niger IBS-103ZA. Food Chem. 2011;124:1595–1602. doi: 10.1016/j.foodchem.2010.08.022. [DOI] [Google Scholar]
- 53.Hegde S., Kavitha S., Varadaraj M.C., Muralikrishna G. Degradation of cereal bran polysaccharide-phenolic acid complexes by Aspergillus niger CFR 1105. Food Chem. 2006;96:14–19. doi: 10.1016/j.foodchem.2005.01.050. [DOI] [Google Scholar]
- 54.Justesen U., Arrigoni E., Larsen B.R., Amado R. Degradation of flavonoid glycosides and aglycones during in vitro fermentation with human faecal flora. LWT-Food Sci. Technol. 2000;33:424–430. doi: 10.1006/fstl.2000.0681. [DOI] [Google Scholar]
- 55.Podsedek A., Sosnowska D., Redzynia M., Anders B. Antioxidant capacity and content of Brassica oleracea dietary antioxidants. Int. J. Food Sci. Technol. 2006;41:49–58. doi: 10.1111/j.1365-2621.2006.01260.x. [DOI] [Google Scholar]
- 56.Sumaryono W., Proksch P., Wray V., Witte L., Hartmann T. Qualitative and quantitative analysis of the phenolic constituents from Orthosiphon aristatus. Planta Med. 1991;57:176–180. doi: 10.1055/s-2006-960060. [DOI] [PubMed] [Google Scholar]
- 57.Gil-Muñoz R., Gómez-Plaza E., Martínez A., López-Roca J.M. Evolution of phenolic compounds during wine fermentation and post-fermentation: Influence of grape temperature. J. Food Compos. Anal. 1999;12:259–272. doi: 10.1006/jfca.1999.0834. [DOI] [Google Scholar]
- 58.Ju H.K., Cho E.J., Jang M.H., Lee Y.Y., Hong S.S., Park J.H., Kwon S.W. Characterization of increased phenolic compounds from fermented Bokbunja (Rubus coreanus Miq.) and related antioxidant activity. J. Pharmaceut. Biomed. Anal. 2009;49:820–827. doi: 10.1016/j.jpba.2008.12.024. [DOI] [PubMed] [Google Scholar]
- 59.Hernández T., Estrella I., Carlavilla D., Martín-Álvarez P.J., Moreno-Arribas M.V. Phenolic compounds in red wine subjected to industrial malolactic fermentation and ageing on lees. Anal. Chim. Acta. 2006;563:116–125. doi: 10.1016/j.aca.2005.10.061. [DOI] [Google Scholar]
- 60.Rodríguez H., Landete J.M., Rivas B.D.L., Muñoz R. Metabolism of food phenolic acids by Lactobacillus plantarum CECT 748T. Food Chem. 2008;107:1393–1398. doi: 10.1016/j.foodchem.2007.09.067. [DOI] [Google Scholar]
- 61.Curiel J.A., Rodríguez H., Landete J.M., de las Rivas B., Muñoz R. Ability of Lactobacillus brevis strains to degrade food phenolic acids. Food Chem. 2010;120:225–229. doi: 10.1016/j.foodchem.2009.10.012. [DOI] [Google Scholar]
- 62.Fletcher R.S., Slimmon T., McAuley C.Y., Kott L.S. Heat stress reduces the accumulation of rosmarinic acid and the total antioxidant capacity in spearmint (Mentha spicata L) J. Sci. Food Agr. 2005;85:2429–2436. doi: 10.1002/jsfa.2270. [DOI] [Google Scholar]
- 63.Ariffin F., Heong Chew S., Bhupinder K., Karim A.A., Huda N. Antioxidant capacity and phenolic composition of fermented Centella asiatica herbal teas. J. Sci. Food Agr. 2011;91:2731–2739. doi: 10.1002/jsfa.4454. [DOI] [PubMed] [Google Scholar]
- 64.Coinu R., Carta S., Urgeghe P.P., Mulinacci N., Pinelli P., Franconi F., Romani A. Dose-effect study on the antioxidant properties of leaves and outer bracts of extracts obtained from Violetto di Toscana artichoke. Food Chem. 2007;101:524–531. doi: 10.1016/j.foodchem.2006.02.009. [DOI] [Google Scholar]
- 65.Soobrattee M.A., Neergheen V.S., Luximon-Ramma A., Aruoma O.I., Bahorun T. Phenolics as potential antioxidant therapeutic agents: Mechanism and actions. Mutat. Res. Fund. Mol. Mech. Mutagen. 2005;579:200–213. doi: 10.1016/j.mrfmmm.2005.03.023. [DOI] [PubMed] [Google Scholar]
- 66.Tepe B. Antioxidant potentials and rosmarinic acid levels of the methanolic extracts of Salvia virgata (Jacq), Salvia staminea (Montbret & Aucher ex Bentham) and Salvia verbenaca (L.) from Turkey. Bioresource Technol. 2008;99:1584–1588. doi: 10.1016/j.biortech.2007.04.008. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.