

Angioimmunoblastic T-cell lymphoma (AITL) is a rare, aggressive type of T-cell lymphoma (TCL) that accounts for 18.5% of mature T-cell or natural killer (NK)-cell lymphomas.1 Mutations in the IDH2, TET2, and DNMT3A genes, frequently seen in various types of cancer, have also been linked to AITL.2,3 However, the molecular mechanisms governing AITL development are largely unknown. CD28 provides a co-stimulatory signal required for full T-cell activation.4 Although many studies have demonstrated the importance of CD28 co-stimulation in T-cell function, the molecular mechanism underlying intracellular signal transduction triggered by CD28 ligation is poorly understood. We and other groups have previously reported identification of a somatic mutation of the CD28 gene, T195P, in AITL samples.5,6 Mutations in the CD28 gene have been rarely reported in other types of tumor and may play important roles in tumorigenesis and progression of AITL considering the importance of CD28 in T-cell activation. T195P substitution was found in 3 out of 9 AITL patients we initially examined. The mutation is located in the cytoplasmic region, which indicated potential alterations in downstream T-cell receptor signaling as a consequence (Figure 1A and B). To better estimate the mutation rate and determine subtype specificity among various categories of TCL, we expanded the patient cohort to include an additional 40 AITL, 39 peripheral TCL not otherwise specified (PTCL-NOS), and 37 NK/T cell lymphoma (NK/T) cases (Figure 1C). Sanger sequencing of PCR products demonstrated a mutation rate of 10.2% (5 of 49) in the cases of AITL (Online Supplementary Figure S1). All of mutation-positive cases were heterozygotes for this mutation. Importantly, the CD28 T195P mutation was not found in PTCL-NOS or in NK/T cases, indicating that this mutation is specific to the AITL subtype. We also analyzed the F51V mutation in CD28,5 but the CD28 F51V mutation was not found in 49 AITL cases. Clinically, CD28 T195P mutation in AITL patients has no correlation with age, sex, international prognostic index, stage, performance status, LDH level, bone marrow involvement, or survival. Overall survival was 45.2±31.6 months for mutation-negative group (44 patients) and 19.2±2.7 months for mutation-positive group (5 patients) (P=0.553 by log rank) (Online Supplementary Figure S2).
Figure 1.
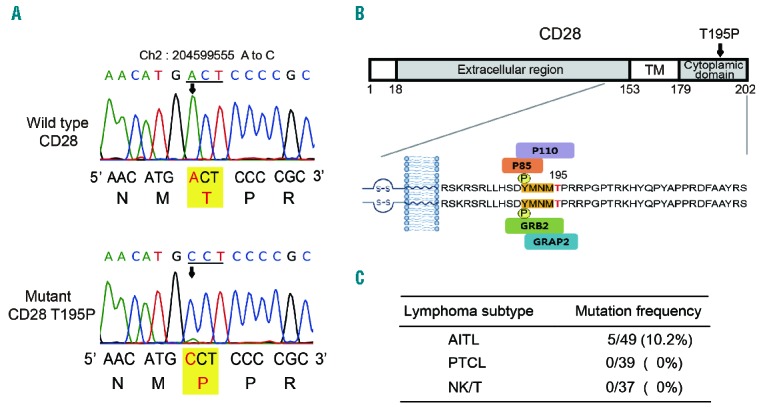
Mutation of T195P on CD28. (A) Representative sequencing traces of wild-type and T195P mutant CD28. Threonine (ACT) was changed to proline (CCT) at the 195th amino acid of CD28. (Top) Sequence from wild-type genomic DNA. (Bottom) Sequence from an AITL sample. Arrows and red letters note the location of the base change. The mutant sequence trace shows heterozygosity. (B) Mutation position and functional domains of the CD28 protein. PI3-K (p110δ/p85): GRB2 and GRAP2 bind to phosphorylated YMNM motif in cytoplasmic region of CD28. The red letter denotes the mutated threonine. TM, transmembrane domain. (C) The genomic DNA of CD28 from lymphoma patients was analyzed by PCR amplification and Sanger sequencing. The mutation of CD28 T195P was detected in 5 (10.2%) of 49 AITL patients, in 0 (0%) of 39 PTCL patients and 0 (0%) of 37 NK/T patients.
We next explored the functional significance of T195P mutation in CD28. This mutation is located in the cytoplasmic region, next to the YMNM motif that associates with signaling proteins such as phosphatidyl inositol 3 kinase (PI3K), GRB2 and GRAP27–9 (Figure 1B). PI3K regulates a number of signaling cascades through the phosphorylation of inositol lipids. GRB2 and GRAP2 are involved in CD28-mediated IL-2 promoter activation and NF-κB activation.10 CD28 becomes tyrosine-phosphorylated by Src family kinases such as Lck and Fyn and by Tec family kinases such as Itk.7,11 Phosphorylation of a tyrosine (Y191) within an YMNM motif and an PYAP motif in the cytoplasmic domain of CD28 permits recruitment of the signaling proteins, which bind to the receptor via their SH2 domains.7,12–15 We first analyzed the binding of signaling proteins to the cytoplasmic domain of CD28 (Figure 2A). GST pull-down assay showed that binding of PI3K (p85/p110) to the cytoplasmic domain with T195P mutation decreased compared to the level seen with the wild-type CD28. However, the binding of GRB2 and GRAP2 to the cytoplasmic domain containing T195P mutation increased either with or without phosphorylation of tyrosine (Y191). These results indicate that the substitution of the 195th amino acid from threonine to proline in CD28 changes the binding of multiple signaling proteins, and, therefore, probably the signaling mediated by CD28. We next analyzed the effect of the CD28 T195P mutation on cell proliferation. Jurkat (human T-cell acute lymphoblastic leukemia) cells transfected with the construct expressing CD28 T195P showed an approximately 15% higher proliferation rate than cells expressing wild-type CD28 either with or without co-stimulation (Figure 2B and Online Supplementary Figure S3). This result indicates that the T195P mutant is likely constitutively active in T-cell stimulation and promotes cell proliferation. Next, we examined the phosphorylation of AKT, mTOR and ERK. These proteins are involved in intracellular signaling pathways required for CD28-mediated co-stimulation of T cells. CD28-mediated co-stimulation led to increased AKT, mTOR and ERK phosphorylation (Figure 2C). Importantly, cells expressing CD28 T195P showed increased AKT, mTOR and ERK phosphorylation relative to cells expressing wild-type CD28, even without stimulation. These results are consistent with the concept that mutant CD28-mediated signaling is constitutive. We also analyzed the effect of CD28 T195P mutation on cytokine production of T cells (Figure 2D). Expression of CD28 T195P mutant in Jurkat cells enhanced production of interleukin 2 (IL-2), a key cytokine for T-cell proliferation, compared to wild-type CD28 after co-stimulation with anti-CD3/CD28 antibodies indicating that CD28 T195P mutant promote proliferation via increasing the expression of IL-2. Because the NF-κB pathway plays a prominent role in CD28-mediated T-cell signaling, we analyzed the effect of CD28 T195P mutation on NF-κB signaling (Figure 2E). Expression of CD28 T195P mutant in Jurkat cells enhanced luciferase activity by approximately 20% compared to wild-type CD28 after stimulation with PMA/ionomycin. We also examined the effect of mutation on NF-κB activation after stimulation by anti-CD3 and CD28 antibodies and found that activation was also obtained with antibody treatment, albeit at slightly lower levels (data not shown). This result indicates that the T195P mutant enhances NF-κB signaling. In summary, we have isolated a recurrent mutation of CD28 and explored mechanistic implications in T-cell lymphoma development. The CD28 T195P mutation was unique to AITL and led to upregulation of TCR signaling. Clearly, further detailed mechanistic analyses are required to fully understand the significance. However, our preliminary functional characterization of CD28 T195P mutation represents an important step toward understanding the molecular mechanism underlying AITL carcinogenesis. T195P mutation in the CD28 gene is oncogenic in nature, and, therefore, is a likely genetic marker for AITL and a potential target for cancer immunotherapies.
Figure 2.
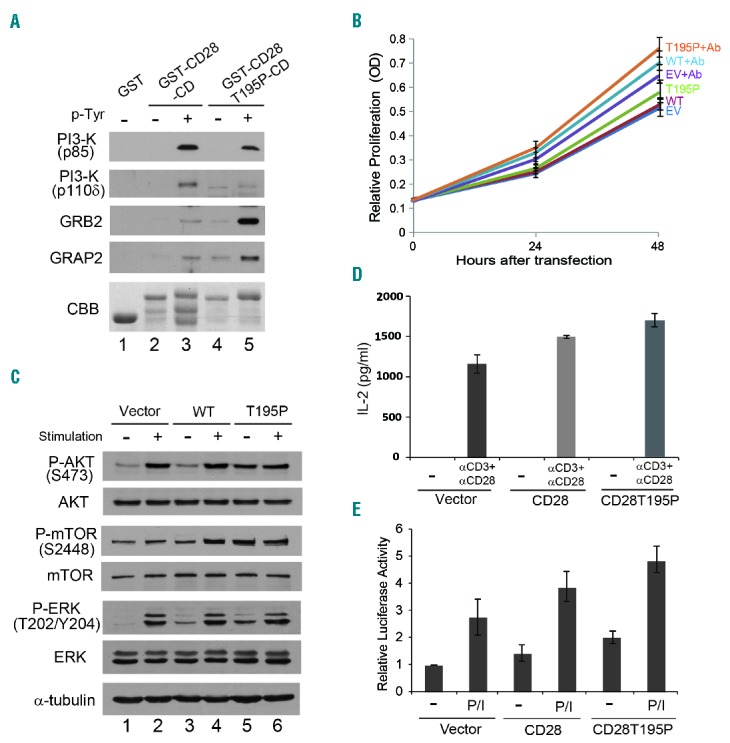
Effect of the T195P mutation on CD28 activity. (A) GST beads containing GST, GST-CD28-CD (cytoplasmic domain), and GST-CD28 T195P-CD were incubated with Jurkat cell lysate. For tyrosine phosphorylation of CD28 cytoplasmic domain, GST-fusion proteins were expressed in E. coli strain expressing tyrosine kinase. Bound proteins were retrieved and examined by immunoblotting with anti-PI3-K (p85/p110δ), anti-GRB2 and anti-GRAP2 antibodies. GST, GST-CD28-CD, and GST-CD28-T195P-CD on immunoblots were stained with Coomassie Brilliant Blue (CBB). (B) Jurkat cells expressing CD28 T195P mutant enhances cell proliferation (P<0.001 compared with cells expressing wild-type CD28) with or without co-stimulation by anti-CD3/CD28 beads. Each condition with 5 replicates was repeated three times, and data are expressed as mean ± standard deviation. OD: optical density; EV: empty vertor. (C) Expression of CD28 T195P enhanced phosphorylation of AKT, mTOR, and ERK without stimulation. (D) Expression of CD28 T195P mutant enhances interleukin 2 (IL-2) production after co-stimulation with anti-CD3/CD28 beads (P<0.001 compared with cells expressing wild-type CD28). IL-2 production was measured three times, and data are expressed as mean ± standard deviation. (E) Expression of CD28 T195P mutant activates NF-κB pathway with or without PMA/ionomycin (P/I) stimulation (P<0.001 compared with cells expressing wild-type CD28). Luciferase activity was measured three times, and data are expressed as mean ± standard deviation.
Footnotes
Funding: this work was supported by grants from the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health & Welfare, Republic of Korea (HI14C3414 and HI14C2331).
Information on authorship, contributions, and financial & other disclosures was provided by the authors and is available with the online version of this article at www.haematologica.org.
References
- 1.Vose J, Armitage J, Weisenburger D, International TCLP. International peripheral T-cell and natural killer/T-cell lymphoma study: pathology findings and clinical outcomes. J Clin Oncol. 2008;26(25):4124–4130. [DOI] [PubMed] [Google Scholar]
- 2.Cairns RA, Iqbal J, Lemonnier F, et al. IDH2 mutations are frequent in angioimmunoblastic T-cell lymphoma. Blood. 2012;119(8):1901–1903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Couronne L, Bastard C, Bernard OA. TET2 and DNMT3A mutations in human T-cell lymphoma. N Engl J Med. 2012;366(1):95–96. [DOI] [PubMed] [Google Scholar]
- 4.Lenschow DJ, Walunas TL, Bluestone JA. CD28/B7 system of T cell costimulation. Annu Rev Immunol. 1996;14:233–258. [DOI] [PubMed] [Google Scholar]
- 5.Yoo HY, Sung MK, Lee SH, et al. A recurrent inactivating mutation in RHOA GTPase in angioimmunoblastic T cell lymphoma. Nat Genet. 2014;46(4):371–375. [DOI] [PubMed] [Google Scholar]
- 6.Rohr J, Guo S, Hu D, et al. CD28 mutations in peripheral T-cell lymphomagenesis and progression. Blood. 2014; 124(21):1681 abstract. [Google Scholar]
- 7.Pages F, Ragueneau M, Rottapel R, et al. Binding of phosphatidylinositol-3-OH kinase to CD28 is required for T-cell signalling. Nature. 1994;369(6478):327–329. [DOI] [PubMed] [Google Scholar]
- 8.Schneider H, Cai YC, Prasad KV, et al. T cell antigen CD28 binds to the GRB-2/SOS complex, regulators of p21ras. Eur J Immunol. 1995;25(4):1044–1050. [DOI] [PubMed] [Google Scholar]
- 9.Ellis JH, Ashman C, Burden MN, et al. GRID: a novel Grb-2-related adapter protein that interacts with the activated T cell costimulatory receptor CD28. J Immunol. 2000;164(11):5805–5814. [DOI] [PubMed] [Google Scholar]
- 10.Watanabe R, Harada Y, Takeda K, et al. Grb2 and Gads exhibit different interactions with CD28 and play distinct roles in CD28-mediated costimulation. J Immunol. 2006;177(2):1085–1091. [DOI] [PubMed] [Google Scholar]
- 11.King PD, Sadra A, Teng JM, et al. Analysis of CD28 cytoplasmic tail tyrosine residues as regulators and substrates for the protein tyrosine kinases, EMT and LCK. J Immunol. 1997;158(2):580–590. [PubMed] [Google Scholar]
- 12.Prasad KV, Cai YC, Raab M, et al. T-cell antigen CD28 interacts with the lipid kinase phosphatidylinositol 3-kinase by a cytoplasmic Tyr(P)-Met-Xaa-Met motif. Proc Natl Acad Sci USA. 1994; 91(7):2834–2838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Stein PH, Fraser JD, Weiss A. The cytoplasmic domain of CD28 is both necessary and sufficient for costimulation of interleukin-2 secretion and association with phosphatidylinositol 3′-kinase. Mol Cell Biol. 1994;14(5):3392–3402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Okkenhaug K, Wu L, Garza KM, et al. A point mutation in CD28 distinguishes proliferative signals from survival signals. Nat Immunol. 2001;2(4):325–332. [DOI] [PubMed] [Google Scholar]
- 15.Boomer JS, Deppong CM, Shah DD, et al. Cutting edge: a double-mutant knockin of the CD28 YMNM and PYAP motifs reveals a critical role for the YMNM motif in regulation of T cell proliferation and Bcl-xL expression. J Immunol. 2014;192(8):3465–3469. [DOI] [PMC free article] [PubMed] [Google Scholar]