

Abstract
Activation of taste buds triggers the release of several neurotransmitters, including ATP and serotonin (5-hydroxytryptamine; 5-HT). Type III taste cells release 5-HT directly in response to acidic (sour) stimuli and indirectly in response to bitter and sweet tasting stimuli. Although ATP is necessary for activation of nerve fibers for all taste stimuli, the role of 5-HT is unclear. We investigated whether gustatory afferents express functional 5-HT3 receptors and, if so, whether these receptors play a role in transmission of taste information from taste buds to nerves. In mice expressing GFP under the control of the 5-HT3A promoter, a subset of cells in the geniculate ganglion and nerve fibers in taste buds are GFP-positive. RT-PCR and in situ hybridization confirmed the presence of 5-HT3A mRNA in the geniculate ganglion. Functional studies show that only those geniculate ganglion cells expressing 5-HT3A-driven GFP respond to 10 μm 5-HT and this response is blocked by 1 μm ondansetron, a 5-HT3 antagonist, and mimicked by application of 10 μm m-chlorophenylbiguanide, a 5-HT3 agonist. Pharmacological blockade of 5-HT3 receptors in vivo or genetic deletion of the 5-HT3 receptors reduces taste nerve responses to acids and other taste stimuli compared with controls, but only when urethane was used as the anesthetic. We find that anesthetic levels of pentobarbital reduce taste nerve responses apparently by blocking the 5-HT3 receptors. Our results suggest that 5-HT released from type III cells activates gustatory nerve fibers via 5-HT3 receptors, accounting for a significant proportion of the neural taste response.
SIGNIFICANCE STATEMENT Historically, serotonin (5-hydroxytryptamine; 5-HT) has been described as a candidate neurotransmitter in the gustatory system and recent studies show that type III taste receptor cells release 5-HT in response to various taste stimuli. In the present study, we demonstrate that a subset of gustatory sensory neurons express functional 5-HT3 receptors that play a significant role in the neurotransmission of taste information from taste buds to nerves. In addition, we show that the anesthetic pentobarbital, widely used in taste nerve recordings, blocks 5-HT3 signaling. Therefore, many conclusions drawn from those data need to be reexamined in light of this anesthetic effect.
Keywords: 5ht3, barbiturate, calcium, geniculate ganglion, serotonin, taste signaling
Introduction
Taste buds contain several types of chemosensory cells and are innervated by multiple nerve fibers. All taste fibers express ionotropic purinergic (P2X) receptors (Bo et al., 1999; Ishida et al., 2009) that are required for transmission from taste receptor cells to the afferent nerves; genetic deletion or pharmacological blockade of the P2X2 and P2X3 receptors expressed by the gustatory nerve fibers eliminates chorda tympani nerve responses to all taste stimuli (Finger et al., 2005; Ohkuri et al., 2012; Jaber et al., 2014; Vandenbeuch et al., 2015). Sweet, umami, and bitter tastants stimulate type II receptor cells to release ATP (Huang et al., 2005, 2009), thereby activating sensory afferents. Sour tastants (acids) and high concentrations of NaCl stimulate type III taste cells (Huang et al., 2006, 2008), yet no one has succeeded in measuring ATP release from these cells. Rather, sour tastants evoke release of serotonin (5-hydroxytryptamine; 5-HT) directly from type III cells (Huang et al., 2005, 2009). However, indirect release of 5-HT has been reported with sweet- and bitter-tasting stimuli (Meredith et al., 2015). Herein, we test whether 5-HT released by taste cells plays a role in the transmission of taste information by activation of 5-HT3 receptors on afferent nerve fibers.
Only two 5-HT receptors have been identified in taste tissue: 5-HT1 and 5-HT3 (Herness and Chen, 2000; Kaya et al., 2004). 5-HT1 receptors are expressed by type II taste cells and 5-HT3 receptors are hypothesized to be expressed in gustatory nerve fibers (Jackson and Yakel, 1995; Wang et al., 2002). Whereas 5-HT1 receptors are inhibitory G-protein-coupled receptors (Hoyer and Schoeffter, 1991), 5-HT3 receptors are excitatory-ligand-gated ion channels that form pentameric combinations of the 5-HT3 isoforms (5-HT3A–5-HT3E). Every functional 5-HT3 receptor must contain at least one 5-HT3A subunit (Jackson and Yakel, 1995; Hassaine et al., 2014).
5-HT released by type III taste cells activates adjacent type II taste cells that express the 5-HT1A receptor, resulting in inhibition of type II cells (Herness and Chen, 2000; Huang et al., 2005; Jaber et al., 2014). The 5-HT released by type III cells could also activate 5-HT3 receptors on afferent nerve fibers. Type III cells (but not type II cells) form traditional synapses with afferent nerve fibers (Kinnamon et al., 1985; Yee et al., 2001; DeFazio et al., 2006) and synthesize and store 5-HT (Dvoryanchikov et al., 2007) in dense-core synaptic vesicles located at these synaptic sites (Fujimoto et al., 1987). Therefore, 5-HT signaling to afferent nerve fibers could participate in the transmission of sour or some components of salty taste by type III cells.
In the present study, we examine the role of 5-HT3 signaling in transmission of taste information. We used anatomical and molecular approaches to demonstrate expression of the 5-HT3A subunit in taste ganglia and taste buds and then used physiological and pharmacological approaches to determine whether these receptors are functional and crucial in the taste system.
Materials and Methods
Animals.
Mice were housed at the University of Colorado Anschutz Medical Campus or at the German Institute of Human Nutrition in ventilated cages on a 12 h/12 h light/dark cycle and fed standard chow ad libitum. All experimental procedures were approved by the Animal Care and Use Committee at the University of Colorado School of Medicine or the German Institute of Human Nutrition. Calcium imaging and anatomical experiments were conducted on tissue from 2- to 6-month-old male and female 5-HT3AGFP mice originally on FVB/N-Swiss Webster background (STOCK Tg(Htr3a-EGFP)DH30Gsat/Mmnc; RRID:IMSR_MMRRC:000273) but crossed for 2–4 generations with the C57BL/6 line. Nerve recordings were conducted on 2- to 6-month-old male and female 5-HT3AKO (Zeitz et al., 2002) mice bred on a C57BL/6J background (B6.129X1-Htr3atm1Jul/J; RRID:IMSR_JAX:005251; Jackson Laboratories) and male C57BL/6J WT (RRID:IMSR_JAX:000664) controls. RT-PCR and in situ hybridization experiments were conducted on tissue from 2- to 6-month-old male C57BL/6J mice. No differences due to sex were observed in any experiments. Genotyping of 5-HT3AGFP and 5-HT3AKO mice were done per distributor recommendations (MMRRC and Jackson Laboratory, respectively).
RT-PCR.
RNA was extracted from geniculate ganglia of 5-HT3AKO and WT mice (3 mice each) according to manufacturer's instructions using the RNeasy Micro kit (Qiagen), including a 30 min DNase I treatment at room temperature for removal of genomic DNA. Reverse transcription of 250 ng of RNA was performed using the iScript cDNA Synthesis kit (Bio-Rad). For every experiment, parallel reactions were set up in which the reverse transcriptase enzyme was omitted as a control to detect for DNA contamination. Ten percent (2 μl) of the RT-PCR product was added to the PCR (Qiagen TaqPCR Core kit).
Primer sequences for each PCR are described in Table 1. PCR primers for 5-HT3A were designed in accordance with information from Jackson Laboratories and anneal in exon 7 and 8 (mutation location of 5-HT3AKO; Table 1). PCR conditions for detection of both 5-HT3A and β-actin included an initial 5 min denaturation step, followed by 35 cycles of 30 s denaturation at 95°C, 30 s annealing at 63°C, and 45 s extension at 72°C; concluding with a 7 min final extension step. We included cDNA from mouse whole brain (Clontech) and a no template control (water). Amplified sequences were visualized by gel electrophoresis in 2% agarose gels stained with GelRed (Biotium).
Table 1.
List of primers used for riboprobe generation (in situ hybridization, ISH) and in RT-PCR experiments
| Gene | Primer sequence | Size | Accession no. |
|---|---|---|---|
| β-actin (PCR) | F: 5′-CACCCTGTGCTGCTCACC-3′ | 328 bp | NM_007393 |
| R: 5′-GCACGATTTCCCTCTCAG-3′ | |||
| 5-HT3A (PCR) | F: 5′-AACAGCTATGCAGAAATGAAGTT-3′ | 66 bp | NM_013561 |
| R: 5′-GGCTGACTGCGTAGAATAAAGG-3′ | |||
| 5-HT3A (for ISH) | F: 5′-GCCTTGACATCTACAACTTCCC-3′ | 660 bp | NM_013561 |
| R: 5′-GAGCAGTCATCAGTCTTGTTGG-3′ |
In situ hybridization.
Using cDNA from brain and oligonucleotides shown in Table 1, we performed PCRs to generate probes for in situ hybridization. PCR conditions included an initial 10 min denaturation step, followed by 39 cycles of 1 min denaturation at 95°C, 30 s annealing at 64°C, and 1 min extension at 68°C, concluding with a 10 min final extension step. PCR products were sequenced and cloned into the transcription vector pBluescriptKS (Stratagene). Before in vitro transcription, we linearized the plasmids with appropriate restriction endonucleases and generated sense and antisense riboprobes using in vitro transcriptions (Roche Applied Science) with T7 polymerase and the DIG RNA labeling kit (Roche Diagnostics).
Frozen sections (14 μm) of mouse ganglia were cut (Microm), thaw mounted onto positively charged glass slides (Menzel), and stored at −80°C. Before hybridization, the sections were fixed with 4% paraformaldehyde in PBS (0.1 m phosphate buffer, pH 7.2, 0.9% saline) and then permeabilized with 0.2 m hydrochloric acid for 10 min and 1% Triton X-100 in PBS for 2 min. After acetylation by treatment with 0.1 m triethanolamine 0.25% acetic anhydride, pH 8.0, the tissue was prehybridized (prehybridization solution: 0.75 m NaCl, 25 mm PIPES, 25 mm EDTA, 5× Denhardt's reagent, 0.2% SDS, 250 μg/ml Escherichia coli tRNA, and 500 μg/ml salmon testis DNA, pH 6.8) at room temperature for 5 h. Riboprobes were incubated for 10 min at 85°C before application onto the sections and used for hybridization at a final concentration of 500 ng/ml. Hybridization was performed overnight at 56°C in a chamber humidified with 50% formamide. After hybridization, the slides were washed several times, followed by 30 min RNAase treatment (1 μg/ml RNase A in 0.5 m NaCl, 10 mm Tris-HCl, and 1.0 mm EDTA, pH 7.5) and additional washes with 1× SSC buffer at 45°C. To detect hybridized riboprobes, sections were incubated with an alkaline-phosphatase-conjugated anti-digoxigenin antibody (1:750; Roche Applied Science) for 1 h at room temperature. Colorimetric detection was accomplished with color substrate (0.175 mg/ml 5-bromo-4-chlor-indolyl-phosphate and 0.25 mg/ml nitroblue-tetrazolium-chloride) applied to the sections and incubated overnight at room temperature in darkness. On the following day, color reaction was stopped by a 5 min incubation in Tris-EDTA buffer. Sections were mounted with glass coverslips and images were taken using a microscope (Axioplan; Zeiss) connected to a CCD camera (RT slider; Diagnostic Instruments).
Geniculate ganglion retrograde label.
The geniculate ganglion houses the cell bodies of nerve fibers innervating taste buds of the anterior tongue and soft palate, as well as general cutaneous fibers innervating part of the external ear. The majority of cells of the geniculate ganglion are gustatory, contributing to the chorda tympani and greater superficial petrosal nerves (Foley and DuBois, 1943; van Buskirk, 1945) innervating, respectively, fungiform papillae and palatal taste fields. Somatosensory innervation of these taste fields arises from the trigeminal nerve; therefore, cells of the geniculate ganglion that innervate taste fields are gustatory rather than somatosensory in function.
To identify ganglion cells innervating palatal or fungiform taste buds, the retrograde tracer FluoroGold (Fluorochrome) was injected into the anterior tongue and soft palate using a protocol modified from King and Bradley (2000). Mice were anesthetized with intramuscular injections of dexmedetomidine hydrochloride (0.4 mg/kg; Pfizer), followed by ketamine hydrochloride (40 mg/kg; Bioniche Pharma). After a surgical level of anesthesia was obtained, the mouth was gently opened and 10–20 μl of a 5% FluoroGold solution in water was injected under taste epithelium using a Hamilton microsyringe (#701; Hamilton). Injection sites included the most anterior end of the tongue (fungiform papillae region), the lateral tongue (foliate papillae region), and the soft palate. Anesthesia was reversed with atipamezole (2 mg/kg; Pfizer) and mice were treated with 0.5% Marcaine (Hospira) during their postsurgical recovery. Five days after injection, the mice were reanesthetized with pentobarbital (FatalPlus 0.02 ml, i.p; Vortech) and then perfused transcardially with 4% paraformaldehyde in phosphate buffer (in mM/L: 29 NaH2PO4, 75 Na2HPO4, pH 7.2–7.4).
Immunohistochemistry.
Tissues were harvested immediately after perfusion fixation with buffered 4% paraformaldehyde and postfixed in the same fixative for 2–4 h. All tissue was cryoprotected overnight with 20% sucrose in phosphate buffer and then cryostat sectioned at 12–16 μm. Some animals were injected intraperitoneally with 5-hydroxy-l-tryptophan (50 mg/kg; Sigma-Aldrich) 1 h before killing to increase 5-HT levels. The cryostat sections were immunoreacted overnight at 4°C with primary antibodies to label specific cell types and nerve fibers: GFP to label 5-HT3AGFP containing nerve fibers, GNAT3 (gustducin) to label a subpopulation of type II taste cells, P2X3 to label nerve fibers, and 5-HT to label serotonin-accumulating type III taste cells. See Table 2 for details of antisera. The humanized monoclonal P2X3 antibody was prepared using a lipoparticle approach (Banik et al., 2011) incorporating full-length human P2X3 protein. Preparations lacking primary antisera were routinely run in parallel to test for specificity of secondary antisera. Specificity of the gustducin antiserum was tested on gustducin-knock-out mice and showed no staining. Similarly, the P2X3 antibody was tested on tissue from P2X3-knock-out mice and no signal was observed. Fluorescent secondary antibodies (Table 3) were reacted with tissue sections for 2 h at room temperature before mounting with Fluoromount G (Southern Biotech).
Table 2.
List of primary antisera
| Target protein | Host | Dilution | Manufacturer | Catalog no. | RRID | Lot |
|---|---|---|---|---|---|---|
| GFP | Chicken | 1:2000 | Aves | GFP-1010 | AB_2307313 | 0511FP12 |
| GNAT3 | Goat | 1:500 | Aviva System Biology | OAEB00418 | AB_10882823 | G2 E060911 |
| Gustducin | Rabbit | 1:5000 | Santa Cruz | SC-395 | AB_673678 | H3112 |
| P2X3 | Guinea pig | 1:750 | Neuromics | GP10108 | AB_2283325 | 400911 |
| P2X3 | Human | 5 μg/ml | Integral Molecular | n/a | n/a | LM-021 |
| Serotonin | Rabbit | 1:5000 | Immunostar | 20080 | AB_572263 | 924005 |
Both P2X3 antisera were validated by lack of staining of tissue from P2X3-KO mice. LM-021 is a humanized monoclonal antibody directed against the full-length protein using lipoparticle-based immunization (Banik et al., 2011).
Table 3.
List of commercially available fluorescent secondary antisera used in this study
| Target species | Host | Dilution | Manufacturer | Catalog no. | RRID | Wavelength |
|---|---|---|---|---|---|---|
| Chicken | Donkey | 1:800 | Jackson ImmunoResearch | 703-475-155 | AB_2340373 | 488 |
| Chicken | Goat | 1:800 | Invitrogen | A11039 | AB_142924 | 488 |
| Goat | Donkey | 1:800 | Invitrogen | A21447 | AB_141844 | 647 |
| Guinea Pig | Donkey | 1:800 | Jackson ImmunoResearch | 706-605-148 | AB_2340476 | 647 |
| Guinea Pig | Goat | 1:800 | Invitrogen | A11075 | AB_141954 | 568 |
| Human | Donkey | 1:800 | Jackson ImmunoResearch | 709-295-149 | AB_2340547 | Rho-Red-X |
| Rabbit | Donkey | 1:800 | Invitrogen | A10042 | AB_11180183 | 568 |
| Rabbit | Goat | 1:800 | Invitrogen | A21245 | AB_141775 | 647 |
Z-stack images of taste buds and geniculate ganglia were collected on an Olympus Fluoview FV300 laser scanning confocal microscope with a 60× oil-immersion objective [numerical aperture (NA) 1.3] and 20× oil-immersion objective (NA 0.7) or on a Leica TCS SP5 laser scanning confocal microscope with a 63× oil-immersion objective (NA 1.4).
Quantification of geniculate ganglion cell labeling.
Regions of interest in the perikaryon of individual ganglion cells (as identified by P2X3 immunoreactivity) were outlined using ImageJ (RRID: nif-0000-30467). Average fluorescence intensity for each region of interest was measured in all channels. Background fluorescence, measured in a region absent of tissue, was subtracted from the measured value of each cell and these background-subtracted values were normalized to the maximum value of its respective channel. For quantification of FluoroGold labeling, cells were scored in a binary fashion (yes or no) for presence of FluoroGold and/or GFP. After fluorescence intensity normalization (where 0 is no expression and 1 is maximum), individual cells were scored in each channel (FluoroGold and GFP) as above or below a predetermined threshold. Threshold was determined by fitting bimodal distributions of intensity values with two peak Gaussian functions. The threshold for expression was defined as any value >2 SDs above the mean of the lower, background peak. Cells above threshold were considered positive for label. For quantitative analysis, only raw images were used. Images for manuscript display were adjusted with Photoshop (Adobe Systems) using only levels, brightness, and contrast modifications to the entire image.
Geniculate ganglion cell isolation.
Geniculate ganglia were rapidly excised from CO2-euthanized mice and placed in enzyme solution [minimum essential medium with Earle's balanced salts (MEM/EBSS; Hyclone) containing 1.25 mg/ml trypsin (Sigma-Aldrich) and 2.5 mg/ml collagenase A (Roche Diagnostics)] for 30 min. Digested ganglia were washed three times with MEM/EBSS before gentle trituration with a fire-polished glass pipette. Cells were resuspended in HEPES buffer containing the following (in mm): 136 NaCl, 5.6 KCl, 1 MgCl2, 2.2 CaCl2, 11 glucose, 10 HEPES; pH adjusted to 7.4 with NaOH before plating on poly-d-lysine (0.02 mg/ml; BD Bioscience) and laminin (0.02 mg/ml; Sigma-Aldrich)-coated coverslips.
Calcium imaging.
Fura-2-AM (50 μm, Invitrogen) stock was prepared in HEPES buffer containing 2.5% DMSO and 0.01% Pluronic F-127 (Invitrogen) and then pipetted into the medium bathing the coverslips to a final concentration of 2 μm Fura-2-Am. Cells were loaded for 20 min before washing with HEPES buffer. The test compounds (all from Sigma-Aldrich), ATP, 5-HT, m-chlorophenylbiguanide (CPG), ondansetron (ODS), and 55 mm KCl (with KCl replacing equimolar NaCl) were added to HEPES buffer. For experiments testing effects of pentobarbital on the ganglion cells, nembutal sodium solution (pentobarbital sodium; Oak Pharmaceuticals) was added to HEPES buffer at a final concentration of 150 or 800 μm. Fura-2-AM-loaded cells were imaged through a 40× oil-immersion objective lens of an inverted microscope and acquired with a Sensicam QE CCD camera (PCO-Tech). Emission at ∼510 nm was collected from excitations at 350 and 380 nm. Images were collected every 3 s using Imaging Workbench 5 software (Indec Biosystems). Raw data are represented as change in fluorescence ratio normalized to baseline fluorescence ratio. Dose–response curves were fit with a four parameter logistic function using Sigmaplot (Systat Software; SciRes_000184).
Nerve recording.
Two different anesthetics were tested in these experiments. Some mice were anesthetized with sodium pentobarbital, a common anesthetic used for taste nerve recording (Horio et al., 2011) and used in a recent study on the possible role of 5-HT1 and 5-HT3 in taste function (Jaber et al., 2014). Because barbiturate anesthetics are reported to interfere with functionality of 5-HT3 receptors in several in vitro systems (Jenkins et al., 1996; Barann et al., 1997; Barann et al., 2000; Rüsch et al., 2007), we also tested urethane, which gave significantly different results (described below). Mice were anesthetized using injections of sodium pentobarbital (50 mg/kg, i.p.) or urethane (2 g/kg, i.p.; Sigma Chemical) and tracheotomized to facilitate breathing. The chorda tympani nerve was exposed using a ventral approach and cut near the tympanic bulla. The whole dissected nerve was placed on a silver electrode and a reference electrode was placed in the nearby tissue. Taste responses were amplified (P511; Grass Instruments), integrated (time constant 0.5 s) and collected using Acknowledge software (Biopac). The anterior tongue was stimulated with a continuous flow (Mini-pump; Fisher Scientific) and different tastants were injected into this flow: NH4Cl 100 mm, sucrose 500 mm, NaCl 100 mm, monosodium glutamate (MSG) 300 mm with amiloride 100 μm, quinine 10 mm, citric acid (3, 5, 10, and 20 mm), and HCl (3, 5, 10, and 20 mm). The tastants were applied for 30 s and rinsed with water for 40 s. In some experiments, a specific antagonist of 5-HT3 receptors (ODS dissolved in 0.9% NaCl; 1 mg/kg) or a vehicle control (0.9% NaCl) was injected intraperitoneally after all stimuli had been successively applied to the tongue. Fifteen minutes after the injection, the same stimuli were reapplied and responses compared before and after injection.
To analyze the data, the amplitude of the integrated responses was averaged for 30 s using Acknowledge software (Biopac) and normalized to the baseline (averaged for 5 s before each taste stimulation). We chose to normalize to the baseline activity as we have done in the past (Vandenbeuch et al., 2013) because serotonergic transmission could affect responses to most taste stimuli. Small drifts in baseline were observed over the course of the experiments, but these changes had little effect on the interpretation of the data because response magnitude was proportional to baseline activity. Individual mice with large drifts in baseline were excluded from analysis. The normalized responses then were compared using a two-way ANOVA with Tukey's post hoc test using Statistica software (verseion 10; Statsoft) for grouped tastants [3–20 mm citric acid, 3–20 mm HCl, and “others” (NH4Cl, NaCl, sucrose, MSG with amiloride, and quinine)]. A statistical summary for nerve recording data is provided in Table 4.
Table 4.
Statistical comparisons for nerve-recording data
| Df | F | p | |
|---|---|---|---|
| WT (7) vs KO (8) (urethane) | |||
| Citric acid | 52 | 5.22 | 0.017 |
| HCl | 39 | 3.29 | 0.009 |
| Others | 39 | 4.44 | 0.001 |
| WT (6), pre vs post ODS injection (urethane) | |||
| Citric acid | 40 | 0.81 | 0.004 |
| HCl | 30 | 5.45 | 0.011 |
| Others | 30 | 2.14 | 0.525 |
| WT (4), pre vs post NaCl injection (urethane) | |||
| Citric acid | 20 | 0.11 | 0.845 |
| HCl | 18 | 0.27 | 0.959 |
| Others | 18 | 0.12 | 0.979 |
| WT (8) vs KO (7) (pentobarbital) | |||
| Citric acid | 39 | 2.09 | 0.240 |
| HCl | 39 | 1.89 | 0.147 |
| Others | 52 | 1.52 | 0.209 |
| WT, urethane (7) vs pentobarbital (8) | |||
| Citric acid | 52 | 6.09 | <0.001 |
| HCl | 39 | 6.74 | <0.001 |
| Others | 39 | 10.19 | <0.001 |
| KO, urethane (8) vs pentobarbital (7) | |||
| Citric acid | 42 | 0.58 | 0.632 |
| HCl | 42 | 0.44 | 0.728 |
| Others | 56 | 3.78 | 0.008 |
| WT, pentobarbital (8) vs post-ODS (6) | |||
| Citric acid | 36 | 2.30 | 0.094 |
| HCl | 26 | 0.73 | 0.540 |
| Others | 48 | 2.43 | 0.060 |
Degrees of freedom (Df), F-statistic, and p-values are for two-way ANOVAs comparing nerve recording data. Number of mice are in parentheses. For post hoc test results, see Figures 6 and 7. Citric acid and HCl, 3–20 mm. “Others,” 100 mm NH4Cl, 500 mm sucrose, 100 mm NaCl, 300 mm MSG plus 100 μm ammiloride, and 10 mm quinine.
Statistics.
All data are represented as normalized raw data or as mean ± SEM. All statistical analyses were performed using Sigmaplot (Systat Software; SciRes_000184) or Statistica (version 10; Statsoft). p < 0.05 was considered statistically significant.
Results
5-HT3A mRNA is expressed in ganglion cells
RT-PCR shows 5-HT3A transcripts in geniculate ganglia as well as in brain (Fig. 1A). Negative control experiments using the same primers on ganglia of 5-HT3AKO mice show no detectable 5-HT3A mRNA. In situ hybridization on sections of gustatory ganglia (geniculate, petrosal/nodose) and a somatosensory ganglion (trigeminal) were performed to reveal serotonin receptor gene expression at the cellular level. In all cases, a subset of ganglion cells displayed robust label for 5-HT3A mRNA (Fig. 1B–G).
Figure 1.
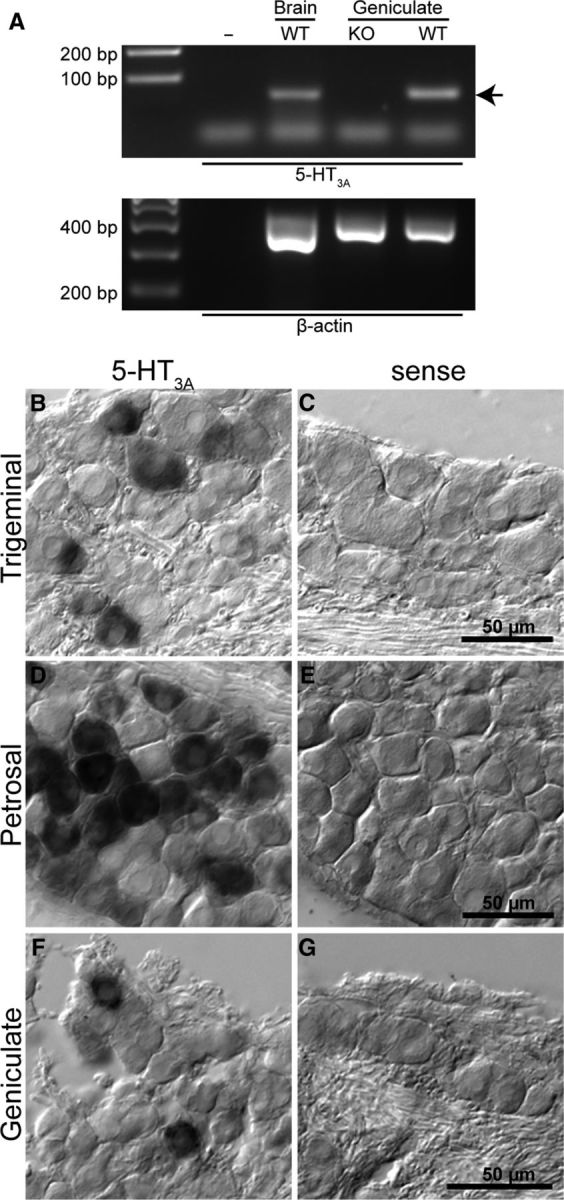
Expression of 5-HT3A mRNA in geniculate ganglia as demonstrated by RT-PCR and in situ hybridization. A, RT-PCR showing the presence of 5-HT3A transcripts in brain and geniculate ganglion of WT mice, but the lack of 5-HT3A mRNA in geniculate ganglion of 5-HT3AKO mice. Arrow indicates expected size of 66 bp 5-HT3A band. Primer dimers likely constitute the smallest band seen in both control and experimental lanes. PCR of β-actin was used as a loading control (lower gel). B–G, In situ hybridization for 5-HT3A in three cranial ganglia and the respective sense controls. B, C, Trigeminal ganglion. D, E, Petrosal ganglion. F, G, Geniculate ganglion. In each ganglion, a subpopulation of ganglion cells show robust signal for 5-HT3A mRNA.
5-HT3A-driven GFP is expressed in a subset of geniculate ganglion cells innervating the tongue and intragemmal fibers of taste buds
To first determine whether gustatory afferents express GFP in 5-HT3AGFP mice, we examined the geniculate ganglion and taste buds. Similar to the results from in situ hybridization, a subset of geniculate ganglion cells displayed bright GFP fluorescence (Fig. 2A). Because all taste fibers express the P2X3 purinergic receptor (Ishida et al., 2009; Vandenbeuch et al., 2015), we immunoreacted geniculate ganglion sections from 5-HT3AGFP mice with antibodies against GFP (to amplify 5-HT3AGFP signal) and against P2X3. GFP immunoreactivity was detected in a subset of geniculate ganglion neurons (169/682, 24.8%; Fig. 2B,C), whereas the P2X3 signal was present in nearly all ganglion cells (664/682, 97.4%; Fig. 2B,C).
Figure 2.

5-HT3A-driven GFP is expressed in geniculate ganglion neurons that innervate the tongue. A, Confocal Z-projection of 10 0.7 μm optical slices of geniculate ganglion from a 5-HT3AGFP mouse showing expression of GFP (green) and P2X3 (magenta). Scale bar, 100 μm. B, Histograms of GFP and P2X3 normalized immunoreactivity of individual cells showing that whereas the P2X3 staining shows a unimodal normal distribution (mean 0.58), the GFP population is bimodal suggestive of high-expressing (mean = 0.69) and low- or nonexpressing populations (mean 0.12). Insert at upper right shows the higher expressing population rescaled along the abscissa. C, Scatterplot comparing relative fluorescence intensity for normalized P2X3 and GFP immunoreactivity. No correlation exists between the two factors (r = 0.16, Pearson's correlation; n = 5 mice, 9 ganglia, 682 cells). Dotted lines (B, C) indicated threshold for each channel. D, Confocal Z-projections of geniculate ganglion from 5-HT3AGFP mice 5 d after FluoroGold injection showing expression of GFP (green) and FluoroGold label (FG; magenta). Arrows indicate double-labeled cells (above threshold in each channel). E, Histograms of GFP and FG normalized fluorescence intensity of individual cells showing bimodal distributions suggestive of a high-expressing population (mean GFP: 0.59, FG: 0.62) and a nonexpressing population (mean GFP: 0.08, FG: 0.08). F, Scatterplot comparing the relative fluorescence intensity for normalized GFP and FG fluorescence. No correlation exists between the two factors (r = 0.35 Pearson's correlation; n = 3 mice, 6 ganglia, 468 cells). Dotted lines (E, F) indicate threshold for each channel. G, Table showing relative number of cells showing suprathreshold expression of GFP and FG label.
To determine whether any 5-HT3AGFP-positive ganglion cells of the geniculate ganglion innervate taste buds, as opposed to the external ear, we injected the retrograde tracer FluoroGold into the anterior tongue and palatal taste fields. After 5 d, many cells in the geniculate ganglion were labeled with FluoroGold. Semiquantitative analysis revealed that approximately half (73/138) of FluoroGold-labeled cell bodies exhibited GFP label, whereas in the ganglion as a whole, including both gustatory and nongustatory innervation, a significantly lower proportion of ganglion cells exhibit GFP fluorescence [139/468; χ2(1,n = 606) = 25.21, p < 0.0001 Pearson's χ2 test; Fig. 2D–G]. Therefore, GFP-expressing ganglion cells preferentially innervate taste buds.
Consistent with GFP expression in the geniculate ganglion, most taste buds innervated by the geniculate ganglion contained some GFP-labeled intragemmal nerve fibers. The GFP-expressing nerve fibers often were closely associated with serotonergic type III cells (Fig. 3A–D), suggesting the possibility of serotonergic transmission between serotonergic type III taste cells and 5-HT3AGFP-expressing nerve fibers. 5-HT3A-driven GFP was not expressed in taste receptor cells. These GFP-labeled fibers were, however, only a subset of the total gustatory innervation of the taste buds, as revealed by immunoreactivity for P2X3 (Fig. 3E).
Figure 3.

5-HT3A-driven GFP is expressed in a subset of nerve fibers innervating taste buds. A–D, Confocal Z-projections of taste buds of a 5-HT3AGFP mouse showing expression of GFP (green), 5-HT (serotonin, type III taste cell marker, red), and GNAT3 (gustducin, type II taste cell marker; blue). The GFP-labeled fibers are closely associated with 5-HT labeled type III cells (white arrows) in all taste fields. A, Eight 0.7 μm optical slices of a taste bud from a fungiform papilla. B, Thirteen 0.7 μm optical slices of a palatal taste bud. C, Ten 0.7 μm optical slices of a taste bud from a foliate papilla. D, Eleven 0.7 μm optical slices of a taste bud from a circumvallate papilla. Arrows highlight sites of convergence between 5-HT-expressing type III cells and 5-HT3AGFP-expressing nerve fibers. Scale bars, 10 μm. E, Longitudinal section through a fungiform papilla from a 5-HT3AGFP mouse stained for P2X3 (red) and gustducin (blue). Intragemmal (inside the taste bud) taste fibers are stained with the P2X3 antibody, whereas GFP expression occurs in a subset of the intragemmal fibers as well as perigemmal fibers innervating the surrounding epithelium. The perigemmal fibers (GFP-only in E), presumably of trigeminal origin, exhibit only 5-HT3A-driven GFP, whereas intragemmal GFP-labeled fibers are also immunoreactive for P2X3. Some intragemmal fibers only show P2X3 immunoreactivity (arrows in E′). Confocal Z-stack 9.6 μm. Off-tissue noise was removed digitally. Blue hues were modified using Adobe Photoshop to enhance visibility.
Geniculate ganglion neurons expressing 5-HT3A-driven GFP have functional 5-HT3 receptors
We next tested the responsiveness of GFP-expressing and GFP-nonexpressing geniculate ganglion neurons to exogenously applied 5-HT. Ganglion cells were isolated from 5-HT3AGFP mice and loaded with the membrane-permeable, ratiometric calcium indicator dye Fura-2-AM. We identified “healthy” cells by a response to 10 μm ATP because all geniculate ganglion neurons express P2X-type purinergic receptors (Ishida et al., 2009; Vandenbeuch et al., 2015). We observed similar robust increases of intracellular calcium concentrations in response to KCl and ATP in both GFP-expressing (n = 28) and GFP-nonexpressing cells (n = 12), whereas 5-HT elicited an increase in intracellular calcium only in GFP-expressing cells (KCl: t(40) = 0.98, p = 0.33; ATP t(38) = 1.04, p = 0.30; 5-HT: t(38) = 5.24, p < 0.0001 Student's t tests; Fig. 4A,B,F). In GFP-expressing cells, dose–response curves revealed EC50 values of 1.2 ± 0.1 μm (n = 9) for ATP and 1.5 ± 0.3 μm (n = 5) for 5-HT (Fig. 4E). Next, to test whether the response to 5-HT was specific to 5-HT3 receptors, we blocked 5-HT3 receptors with ODS and coapplied 5-HT. Preapplication of 1 μm ODS blocked the response to 5-HT (n = 12, t(11) = 7.09, p < 0.0001 paired Student's t test), but not to ATP (n = 6, t(5) = 0.66, p = 0.55 paired Student's t test; Fig. 4C,G,H). In most cases, the antagonistic effects of ODS were reversible. The ability of the specific 5-HT3 antagonist to block Ca2+ responses indicates that 5-HT3 is the only 5-HT receptor capable of generating a Ca2+ signal in the ganglion cells. As an alternative test for receptor specificity, we applied the 5-HT3 receptor agonist CPG, which induced an increase in intracellular calcium in 5-HT3AGFP cells; the agonistic effect was blocked by ODS (n = 5, t(4) = 20.55, p < 0.0001 paired Student's t test; Fig. 4D,H). From these data, we conclude that activation of geniculate ganglion neurons by 5-HT, as measured by cytosolic Ca2+, is mediated only by 5-HT3 receptors.
Figure 4.
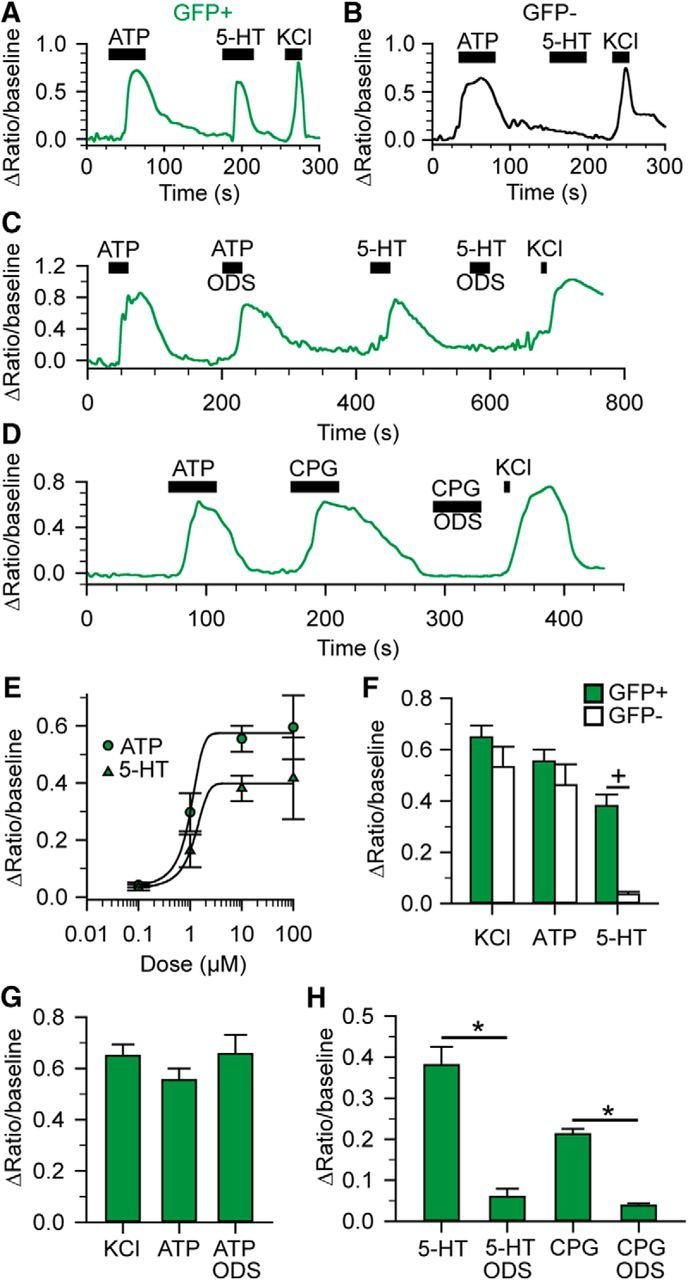
5-HT3AGFP-expressing cells show increased intracellular Ca2+ in response to 5-HT3 activation. A, B, Ca2+ signals from GFP+ (A) and GFP− (B) geniculate ganglion cells in response to 10 μm ATP, 10 μm 5-HT, and 55 mm KCl. C, D, Ca2+ signals from GFP+ geniculate ganglion cells in response to 10 μm ATP, 10 μm ATP + 1 μm ODS, 10 μm 5-HT, 10 μm 5-HT + 1 μm ODS, and 55 mm KCl (C) or 10 μm ATP, 10 μm CPG, 10 μm CPG + 1 μm ODS, and 55 mm KCl (D). E, ATP (circles) and 5-HT (triangles) dose–response curves for 5-HT3AGFP-expressing geniculate ganglion cells. F, Average responses to KCl, ATP, and 5-HT in GFP+ and GFP− ganglion cells. Cross indicates p < 0.05, Student's t test. G, Average responses of 5-HT3AGFP-expressing cells to 55 mm KCl, 10 μm ATP and ATP + 1 μm ODS. All conditions were measured in each cell (n = 5, p = 0.399, one-way ANOVA). H, Average responses of 5-HT3AGFP-expressing cells. Responses to both 5-HT and CPG are blocked by the 5-HT3-antagonist ODS. Asterisks indicate p < 0.0001, paired Student's t tests.
Ganglion cell responses to 5-HT are lacking in 5-HT3AKO mice
Our data demonstrate that a subset of geniculate ganglion neurons expresses 5-HT3 receptors and innervates taste buds, including serotonergic type III cells. These findings suggest that 5-HT released from type III taste cells might activate gustatory sensory terminals in taste buds. Given these conditions, we hypothesized that 5-HT3 receptors would be implicated in the transmission of type III cell-mediated taste information from taste buds to the nerve fibers. To test this, we used mice lacking functional 5-HT3 receptors (5-HT3AKO mice). Successful genetic deletion of 5-HT3 was confirmed by RT-PCR (Fig. 1A). In addition, we tested whether other 5-HT receptors were upregulated after genetic deletion of 5-HT3 by measuring 5-HT responses in isolated geniculate ganglion neurons of WT and 5-HT3AKO mice. Approximately 25% of all WT geniculate neurons (5/19; Fig. 5) responded to exogenous 5-HT with an increase in intracellular calcium, whereas no neurons from 5-HT3AKO mice responded (0/25, p = 0.011, Fisher's exact test; Fig. 5). Therefore, even after genetic elimination of the 5-HT3 receptor, no other serotonin receptors were capable of generating a calcium signal in the ganglion cells.
Figure 5.

Geniculate ganglion neurons of 5-HT3AKO mice do not respond to serotonin. Normalized responses of Fura-2-AM loaded geniculate ganglion neurons revealed that a subset (solid circles; 5/19) of WT neurons were activated by 10 μm 5-HT; no neurons from 5-HT3AKO animals (open circles; 0/25) responded to 5-HT, yet all responded to 10 μm ATP and 55 mm KCl. Symbols represent individual cells. Data are normalized to the response to KCl. Asterisk indicates p < 0.05, Fisher's exact test.
Contribution of 5-HT3 receptors to transmission of taste information
Under urethane anesthesia, the chorda tympani nerves of both WT and KO animals responded robustly to all classes of taste stimuli. Nonetheless, responses in KO animals to most stimuli (100 mm NH4Cl, 500 mm sucrose, 300 mm MSG with amiloride, 5–20 mm citric acid, and 5–20 mm HCl) were reduced compared with WT (Fig. 6B, Table 4), suggesting a role for 5-HT3 signaling in more than just acid and salt transmission; that is, tastants likely to evoke direct type III cell-mediated signaling.
Figure 6.

Role of 5-HT3A signaling in taste transmission. A, Example of integrated chorda tympani responses of a single WT mouse under urethane anesthesia to multiple stimuli before and after ODS injection (1 mg/kg, i.p). Scale bar, 30 s. B, Average chorda tympani responses normalized to baseline of WT and 5-HT3AKO mice. Responses to multiple stimuli were significantly larger in WT mice. n = 7 WT mice (6–7 trials per tastant), 8 KO mice (6–8 trials per tastant). C, D, Average chorda tympani responses in WT mice before and after injection of the 5-HT3 antagonist ODS (C) or vehicle (D). The responses to acids were significantly smaller after ODS treatment, but not vehicle injection. n = 6 mice (6 trials per tastants) ODS, 4 mice (4 trials per tastants) vehicle. Data are presented as mean ± SEM. Asterisks indicate p < 0.05, two-way ANOVA with Tukey's post hoc test.
To further examine the role of 5-HT3 receptors in taste transmission, we tested whether the specific 5-HT3 antagonist ODS decreased taste responses in WT mice under urethane anesthesia. Intraperitoneal injection of ODS significantly decreased chorda tympani nerve response magnitude to acids (sour tastants), as well as NaCl (Fig. 6C, Table 4). Injection of vehicle had no effect on the magnitude of any responses, indicating that reductions measured with ODS were not the result of general loss of nerve activity over the course of the experiment (Fig. 6D, Table 4). Although the effects of ODS on acid and salt responses closely phenocopied 5-HT3AKO, the effects of ODS on other stimuli were less robust but trended in the same direction.
Effects of anesthesia on 5-HT3A signaling
We find that pharmacological blockade of 5-HT3 receptors with ODS, similar to the 5-HT3KO, significantly reduces chorda tympani responses to multiple stimuli, including acids and salt. In contrast, Jaber et al. (2014), using pentobarbital anesthetic, reported that ODS injection in rats had no effect on chorda tympani responses. Because barbiturate anesthetics (e.g., pentobarbital) are reported to block 5-HT3 signaling in heterologous expression systems (Barann et al., 2000; Rüsch et al., 2007), we tested the effects of different anesthetics (pentobarbital vs urethane) on chorda tympani responses. Similar to the findings reported by Jaber et al. (2014) for rats, chorda tympani responses to all tastants were similar for WT and KO mice under pentobarbital anesthesia (Fig. 7A, Table 4). Next, we compared the difference between urethane and pentobarbital anesthesia in WT mice. In WT animals, responses to all tastants except 5 mm quinine were larger under urethane anesthesia than with pentobarbital (Fig. 7B, Table 4), but this difference in response magnitude with anesthetic did not occur in the KO mice (Fig. 7C, Table 4), with the exception of sucrose. The lack of effect of pentobarbital in 5-HT3KO mice suggests an interaction between the drug and 5-HT3 receptors in vivo. To further understand the mechanism by which pentobarbital affects chorda tympani responses, we applied pentobarbital to isolated 5-HT3AGFP-expressing geniculate ganglion neurons. Similar to other in vitro studies (Barann et al., 1997; Barann et al., 2000; Rüsch et al., 2007), pentobarbital inhibited 5-HT induced calcium responses in geniculate ganglion neurons [n = 10 cells (n = 5 for 150 μm pentobarbital), F(1,24) = 105.66, p < 0.001 one-way repeated-measures ANOVA; Figure 7D,E]. The block of serotonin signaling in ganglion cells by pentobarbital could explain the reduced chorda tympani responses in WT mice under pentobarbital anesthesia (Fig. 7B).
Figure 7.

Effect of pentobarbital anesthesia on 5-HT3A signaling. A, Average chorda tympani responses of WT and 5-HT3AKO mice under pentobarbital anesthesia. Responses to all stimuli were similar between WT and KO mice. n = 8 WT mice (7–8 trials per tastant), 7 KO mice (6–7 trials per tastant). B, Average chorda tympani responses of WT mice under different anesthetic conditions. Responses to most stimuli were significantly larger under urethane anesthesia. n = 7 mice urethane (6–7 trials per tastant), 8 mice pentobarbital (7–8 trials per tastant). C, Average chorda tympani responses of 5-HT3AKO mice under different anesthetic conditions. There was no significant difference in response to acid between pentobarbital and urethane anesthesia. The response to sucrose was significantly different under urethane anesthesia. n = 8 mice (7–8 trials per tastant) for urethane, 7 mice (6–7 trials per tastants) for pentobarbital. D, Example Fura-2-loaded geniculate ganglion neuron response to 10 μm ATP, 10 μm 5-HT, 10 μm 5-HT + 800 μm pentobarbital, and 55 mm KCl. E, Summary data of all ganglion cells in response to 5-HT and 5-HT + pentobarbital. n = 10 cells for 0 and 800 μm pentobarbital, n = 5 cells 150 μm pentobarbital. Nerve-recording summary data are presented as mean ± SEM. Asterisks indicate p < 0.05, two-way ANOVA with Tukey's post hoc test. Triple asterisks indicate p < 0.001, one-way repeated-measures ANOVA with Tukey's post hoc test.
In summary, both genetic deletion and pharmacological blockade of the 5-HT3 receptor affect transmission of multiple taste modalities. Neither manipulation of 5-HT3 entirely eliminates the responses, suggesting that other neurotransmitters such as ATP participate in neurotransmission in this system.
Discussion
Taste stimuli activate taste receptor cells, which release one or more neurotransmitters to excite gustatory afferent nerve fibers (Roper, 2013). These nerve fibers encode taste quality information, which is transmitted to the brainstem gustatory complex. However, a paucity of information exists on the steps between taste receptor cell activation and afferent nerve fiber firing. Although ATP acting on neural P2X receptors is crucial for transmission of all taste qualities (Finger et al., 2005; Vandenbeuch et al., 2015), the role of other substances acting as neurotransmitters, cotransmitters, or modulators of neural activity is less clear (Kaya et al., 2004; Huang et al., 2005; Jaber et al., 2014; Takai et al., 2015). Further, whereas ATP release from type II taste cells has been observed in response to bitter-, sweet-, and umami-tasting stimuli, ATP release has not been detected from type III taste cells (Huang et al., 2007; Romanov et al., 2007; Murata et al., 2010). These data suggest that one or more other transmitters may play a role in afferent signaling. In the present study, we have investigated the role of 5-HT in the communication between taste receptor cells and afferent nerve fibers.
Using a mouse expressing GFP under the control of the 5-HT3A promoter, we found that a subset of geniculate ganglion neurons innervating taste buds expresses GFP, suggesting the presence of 5-HT3 receptors. Our physiological analysis established that functional 5-HT3 receptors are present only in these GFP-labeled ganglion cells and that 5-HT3 is required for ganglion cell responses to 5-HT. Although other 5-HT receptors may exist in the ganglion, 5-HT3 is the only ionotropic 5-HT receptor and is normally involved in excitatory signaling in the nervous system (Galligan et al., 2000; Galligan, 2002). Further, in the absence of 5-HT3 signaling, ganglion cells do not respond to 5-HT, as assessed by Ca2+ imaging. It is possible that the 5-HT3 receptor that we demonstrate in the ganglion cell somata is not transported to the distal processes that innervate taste buds but other receptors present in cell bodies of taste ganglia are, including P2X receptors (Bo et al., 1999) and GLP-1 receptors (Shin et al., 2008). Furthermore, injection of 5-HT3 antagonist in vivo materially affects taste responses recorded from the afferent nerves distal to the ganglion, suggesting a peripheral site of action.
Although 5-HT3 receptors play a role in afferent signaling in taste buds, the full neural taste response clearly involves other neurotransmitter receptors. ATP acting on P2X2 and P2X3 receptors is required for transmission of all taste qualities (Finger et al., 2005; Vandenbeuch et al., 2015). The source of ATP for transmission of bitter, sweet, and umami tastants is evidently the type II taste cells, which release ATP with gustatory activation (Huang et al., 2007; Murata et al., 2010). For acids, the taste of which is mediated by type III cells, one possibility is that ATP is coreleased from vesicles along with 5-HT. However, type III cells do not express the vesicular nucleotide transporter VNUT, which is reportedly required to load ATP into vesicles (Iwatsuki et al., 2009), and ATP release has not been detected from type III cells, so the source of ATP with sour stimulation is unclear (Huang et al., 2007; Romanov et al., 2007). One possibility is that tonic release of ATP, possibly from type II cells or nongustatory tissue, may be required to set a basal tone of afferent nerve depolarization upon which another type III cell transmitter such as 5-HT provides additional depolarization to excite gustatory afferents in response to acids. In the absence of basal P2X activation, the gustatory afferents may simply be too hyperpolarized to permit generation of action potentials by other transmitters and modulators. Nonetheless, the persistence of residual taste-induced neural activity in the absence of 5-HT3 signaling strongly suggests that ATP, possibly along with other neurotransmitters, provides sufficient signaling to generate a neural response to taste.
A role for 5-HT in taste signaling has been described in humans in that 5-HT reuptake inhibitors decrease the threshold for quinine and sucrose (Heath et al., 2006), but whether this may be a peripheral or central effect is unclear. In rats, however, the same 5-HT reuptake inhibitors had no effect on the behavioral taste thresholds to sucrose, NaCl, or citric acid, thus confounding the role of 5-HT in peripheral taste signaling (Mathes and Spector, 2014). Whether these confounds represent differences in methodology or possibly species differences remains to be determined.
In taste buds, 5-HT is synthesized, stored, and ultimately released by type III taste cells that respond to acidification (Huang et al., 2009, 2011) or high levels of NaCl (Oka et al., 2013). The serotonergic type III taste cells appear to contact the 5-HT3AGFP-expressing taste nerve fibers, which then are positioned appropriately to respond to a 5-HT signal. Whole taste nerve recordings in WT mice show that 5-HT3 antagonists (both ODS and pentobarbital) significantly decrease responses to most tastants, suggesting that 5-HT3 receptors have a role in communication between taste receptor cells and afferent nerve fibers. Superficially, our findings seem discrepant with the lack of effect of intravascular injection of ODS on chorda tympani taste responses in rats (Jaber et al., 2014), but this apparent discrepancy is likely due to the use of barbiturate anesthetics in that study, which by itself in our experiments in mice blocks 5-HT3 signaling.
The pronounced effect of barbiturate anesthetics on 5-HT3 function as reported in our study in vivo mirrors previous findings in heterologous, in vitro systems (Jenkins et al., 1996; Barann et al., 1997; Barann et al., 2000; Rüsch et al., 2007), in which barbiturates act via an allosteric interaction with the receptor (Davies, 2011). In our study, pentobarbital inhibited 5-HT evoked calcium signals in geniculate ganglion neurons and, at anesthetic levels in mice, significantly inhibits 5-HT3 function in vivo. For the anesthetic doses used in our experiments, plasma and tissue concentrations of intraperitoneally injected pentobarbital in mice are likely to be at or above the reported IC50 values for 5-HT3 receptors (Nelson and Halberg, 1973). Other investigators working with human 5-HT3A suggest that barbiturates only affect these receptors at doses considerably higher than the anesthesic levels used in our study (Rüsch et al., 2007).
We note that the effects of blocking 5-HT3 or knocking out 5-HT3A were not restricted to established type III cell-mediated modalities—that is, acids (Huang et al., 2008) and high concentrations of NaCl (Oka et al., 2013)—but also affected responses to other tastants, including NH4Cl and sucrose. Although the cell type mediating NH4Cl responses is unclear, nonserotonergic, type II taste cells are responsible for transmission of sucrose. The unexpected involvement of 5-HT3 in sucrose responses may be attributable to the secondary release of 5-HT from type III cells in response to the ATP released from type II taste cells during stimulation with sweeteners (Meredith et al., 2015). In lingual slices, type III cells show intracellular calcium responses to multiple stimuli, including sweeteners (Tomchik et al., 2007), likely due to secondary activation via sweet-responsive type II cells. The 5-HT thus released has been hypothesized to be involved in a negative feedback onto type II cells via 5-HT1A receptors (Huang et al., 2009). Nonetheless, some of the released 5-HT will likely activate the neural 5-HT3 receptors. If a significant stimulation of 5-HT3 expressing afferents occurs with sweet tastant stimulation, then one would expect sweet-tasting stimuli to evoke indirect side band excitation of sour-best afferent fibers. This has not been observed in single-fiber studies of taste specificity in mice (Ninomiya et al., 1982, 1984a,b); however, those studies were performed using pentobarbital as an anesthetic, so the 5-HT3 receptors would have been blocked under those conditions.
The widespread use of pentobarbital as an anesthetic in studies entailing peripheral nerve recordings (Ninomiya et al., 1982; Dahl et al., 1997; Nelson et al., 2002; Horio et al., 2011; Oka et al., 2013; Jaber et al., 2014; Vandenbeuch et al., 2015; Barretto et al., 2015) calls into question the degree to which these studies reflect the native condition of the system. Because type III cells appear to integrate information from other receptor cells (Tomchik et al., 2007) and use the 5-HT3 system to transmit information, these previous nerve recording studies may have inhibited transmission of integrated cross-modal information from type III cells, leading to the appearance of strict labeled-line, modality-specific transmission of information. Interestingly, when nonbarbiturate anesthetics are used, a concentration-dependent increase in sideband excitation is observed in a subset of geniculate ganglion neurons (Wu et al., 2015). Perhaps this sideband excitation is due to intrabud signaling from type II to type III cells, thus increasing serotonin release and activation of 5-HT3 receptors with increased stimulus concentration.
In summary, our results show that serotonin, released by taste buds and activating neural 5-HT3 receptors, plays a significant but nonessential role in transmission of taste information from the tongue to the nervous system. Further, we show that pentobarbital and possibly other barbiturate anesthetics inhibit 5-HT3-mediated responses in this system, affecting some taste modalities more than others. Because the large majority of recordings from taste nerves in the past have relied on such anesthetics, reexamination of single-fiber specificity in the system should be undertaken with nonbarbiturate anesthetics.
Footnotes
This work was supported by the National Institutes of Health [Grant R01DC012931 to T.E.F., Grant R01DC012555 to S.C.K., and Grant P30DC004657 to Dr. Diego Restrepo (University of Colorado, School of Medicine)], the Federal Ministry for Education and Research, and the Ministry for Science, Research and Culture of the State of Brandenburg, Germany (W.M.). We thank Nicole Schultz and Jason Parnes for help with tissue preparation for immunohistochemistry, Kyndal Davis for genotyping and maintaining the mouse colony, Catherine Anderson for assistance with RT-PCR experiments, Jennifer Stratford for assistance with statistical analysis, and Integral Molecular for the kind gift of anti-P2X3 LM-021.
The authors declare no competing financial interests.
References
- Banik Soma SR, Berdougo E, Doranz BJ. Antibodies against membrane protein targets. Genet Eng Biotechnol News. 2011;31:18. [Google Scholar]
- Barann M, Göthert M, Bönisch H, Dybek A, Urban BW. 5-HT3 receptors in outside-out patches of N1E-115 neuroblastoma cells: basic properties and effects of pentobarbital. Neuropharmacology. 1997;36:655–664. doi: 10.1016/S0028-3908(97)00059-2. [DOI] [PubMed] [Google Scholar]
- Barann M, Meder W, Dorner Z, Brüss M, Bönisch H, Göthert M, Urban BW. Recombinant human 5-HT3A receptors in outside-out patches of HEK 293 cells: basic properties and barbiturate effects. Naunyn Schmiedebergs Arch Pharmacol. 2000;362:255–265. doi: 10.1007/s002100000288. [DOI] [PubMed] [Google Scholar]
- Barretto RP, Gillis-Smith S, Chandrashekar J, Yarmolinsky DA, Schnitzer MJ, Ryba NJ, Zuker CS. The neural representation of taste quality at the periphery. Nature. 2015;517:373–376. doi: 10.1038/nature13873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bo X, Alavi A, Xiang Z, Oglesby I, Ford A, Burnstock G. Localization of ATP-gated P2X2 and P2X3 receptor immunoreactive nerves in rat taste buds. Neuroreport. 1999;10:1107–1111. doi: 10.1097/00001756-199904060-00037. [DOI] [PubMed] [Google Scholar]
- Dahl M, Erickson RP, Simon SA. Neural responses to bitter compounds in rats. Brain Res. 1997;756:22–34. doi: 10.1016/S0006-8993(97)00131-5. [DOI] [PubMed] [Google Scholar]
- Davies PA. Allosteric modulation of the 5-HT3 receptor. Curr Opin Pharmacol. 2011;11:75–80. doi: 10.1016/j.coph.2011.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeFazio RA, Dvoryanchikov G, Maruyama Y, Kim JW, Pereira E, Roper SD, Chaudhari N. Separate populations of receptor cells and presynaptic cells in mouse taste buds. J Neurosci. 2006;26:3971–3980. doi: 10.1523/JNEUROSCI.0515-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dvoryanchikov G, Tomchik SM, Chaudhari N. Biogenic amine synthesis and uptake in rodent taste buds. J Comp Neurol. 2007;505:302–313. doi: 10.1002/cne.21494. [DOI] [PubMed] [Google Scholar]
- Finger TE, Danilova V, Barrows J, Bartel DL, Vigers AJ, Stone L, Hellekant G, Kinnamon SC. ATP signaling is crucial for communication from taste buds to gustatory nerves. Science. 2005;310:1495–1499. doi: 10.1126/science.1118435. [DOI] [PubMed] [Google Scholar]
- Foley JO, DuBois FS. An experimental study of the facial nerve. J Comp Neurol. 1943;79:79–105. doi: 10.1002/cne.900790107. [DOI] [Google Scholar]
- Fujimoto S, Ueda H, Kagawa H. Immunocytochemistry on the localization of 5-hydroxytryptamine in monkey and rabbit taste buds. Acta Anat (Basel) 1987;128:80–83. doi: 10.1159/000146320. [DOI] [PubMed] [Google Scholar]
- Galligan JJ. Pharmacology of synaptic transmission in the enteric nervous system. Curr Opin Pharmacol. 2002;2:623–629. doi: 10.1016/S1471-4892(02)00212-6. [DOI] [PubMed] [Google Scholar]
- Galligan JJ, LePard KJ, Schneider DA, Zhou X. Multiple mechanisms of fast excitatory synaptic transmission in the enteric nervous system. J Auton Nerv Syst. 2000;81:97–103. doi: 10.1016/S0165-1838(00)00130-2. [DOI] [PubMed] [Google Scholar]
- Hassaine G, Deluz C, Grasso L, Wyss R, Tol MB, Hovius R, Graff A, Stahlberg H, Tomizaki T, Desmyter A, Moreau C, Li XD, Poitevin F, Vogel H, Nury H. X-ray structure of the mouse serotonin 5-HT3 receptor. Nature. 2014;512:276–281. doi: 10.1038/nature13552. [DOI] [PubMed] [Google Scholar]
- Heath TP, Melichar JK, Nutt DJ, Donaldson LF. Human taste thresholds are modulated by serotonin and noradrenaline. J Neurosci. 2006;26:12664–12671. doi: 10.1523/JNEUROSCI.3459-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herness MS, Chen Y. Serotonergic agonists inhibit calcium-activated potassium and voltage-dependent sodium currents in rat taste receptor cells. J Membr Biol. 2000;173:127–138. doi: 10.1007/s002320001014. [DOI] [PubMed] [Google Scholar]
- Horio N, Yoshida R, Yasumatsu K, Yanagawa Y, Ishimaru Y, Matsunami H, Ninomiya Y. Sour taste responses in mice lacking PKD channels. PLoS One. 2011;6:e20007. doi: 10.1371/journal.pone.0020007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoyer D, Schoeffter P. 5-HT receptors: subtypes and second messengers. J Recept Signal Transduct. 1991;11:197–214. doi: 10.3109/10799899109066399. [DOI] [PubMed] [Google Scholar]
- Huang AL, Chen X, Hoon MA, Chandrashekar J, Guo W, Tränkner D, Ryba NJ, Zuker CS. The cells and logic for mammalian sour taste detection. Nature. 2006;442:934–938. doi: 10.1038/nature05084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang YA, Maruyama Y, Stimac R, Roper SD. Presynaptic (Type III) cells in mouse taste buds sense sour (acid) taste. J Physiol. 2008;586:2903–2912. doi: 10.1113/jphysiol.2008.151233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang YA, Dando R, Roper SD. Autocrine and paracrine roles for ATP and serotonin in mouse taste buds. J Neurosci. 2009;29:13909–13918. doi: 10.1523/JNEUROSCI.2351-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang YA, Pereira E, Roper SD. Acid stimulation (sour taste) elicits GABA and serotonin release from mouse taste cells. PLoS One. 2011;6:e25471. doi: 10.1371/journal.pone.0025471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang YJ, Maruyama Y, Lu KS, Pereira E, Plonsky I, Baur JE, Wu D, Roper SD. Mouse taste buds use serotonin as a neurotransmitter. J Neurosci. 2005;25:843–847. doi: 10.1523/JNEUROSCI.4446-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang YJ, Maruyama Y, Dvoryanchikov G, Pereira E, Chaudhari N, Roper SD. The role of pannexin 1 hemichannels in ATP release and cell-cell communication in mouse taste buds. Proc Natl Acad Sci U S A. 2007;104:6436–6441. doi: 10.1073/pnas.0611280104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ishida Y, Ugawa S, Ueda T, Yamada T, Shibata Y, Hondoh A, Inoue K, Yu Y, Shimada S. P2X2- and P2X3-positive fibers in fungiform papillae originate from the chorda tympani but not the trigeminal nerve in rats and mice. J Comp Neurol. 2009;514:131–144. doi: 10.1002/cne.22000. [DOI] [PubMed] [Google Scholar]
- Iwatsuki K, Ichikawa R, Hiasa M, Moriyama Y, Torii K, Uneyama H. Identification of the vesicular nucleotide transporter (VNUT) in taste cells. Biochem Biophys Res Commun. 2009;388:1–5. doi: 10.1016/j.bbrc.2009.07.069. [DOI] [PubMed] [Google Scholar]
- Jaber L, Zhao FL, Kolli T, Herness S. A physiologic role for serotonergic transmission in adult rat taste buds. PLoS One. 2014;9:e112152. doi: 10.1371/journal.pone.0112152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jackson MB, Yakel JL. The 5-HT3 receptor channel. Annu Rev Physiol. 1995;57:447–468. doi: 10.1146/annurev.ph.57.030195.002311. [DOI] [PubMed] [Google Scholar]
- Jenkins A, Franks NP, Lieb WR. Actions of general anaesthetics on 5-HT3 receptors in N1E-115 neuroblastoma cells. Br J Pharmacol. 1996;117:1507–1515. doi: 10.1111/j.1476-5381.1996.tb15314.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaya N, Shen T, Lu SG, Zhao FL, Herness S. A paracrine signaling role for serotonin in rat taste buds: expression and localization of serotonin receptor subtypes. Am J Physiol Regul Integr Comp Physiol. 2004;286:R649–R658. doi: 10.1152/ajpregu.00572.2003. [DOI] [PubMed] [Google Scholar]
- King MS, Bradley RM. Biophysical properties and responses to glutamate receptor agonists of identified subpopulations of rat geniculate ganglion neurons. Brain Res. 2000;866:237–246. doi: 10.1016/S0006-8993(00)02292-7. [DOI] [PubMed] [Google Scholar]
- Kinnamon JC, Taylor BJ, Delay RJ, Roper SD. Ultrastructure of mouse vallate taste buds. I. Taste cells and their associated synapses. J Comp Neurol. 1985;235:48–60. doi: 10.1002/cne.902350105. [DOI] [PubMed] [Google Scholar]
- Mathes CM, Spector AC. Systemic modulation of serotonergic synapses via reuptake blockade or 5HT1A receptor antagonism does not alter perithreshold taste sensitivity in rats. Chem Senses. 2014;39:583–593. doi: 10.1093/chemse/bju028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meredith TL, Corcoran A, Roper SD. Leptin's effect on taste bud calcium responses and transmitter secretion. Chem Senses. 2015;40:217–222. doi: 10.1093/chemse/bju066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murata Y, Yasuo T, Yoshida R, Obata K, Yanagawa Y, Margolskee RF, Ninomiya Y. Action potential-enhanced ATP release from taste cells through hemichannels. J Neurophysiol. 2010;104:896–901. doi: 10.1152/jn.00414.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelson G, Chandrashekar J, Hoon MA, Feng L, Zhao G, Ryba NJ, Zuker CS. An amino-acid taste receptor. Nature. 2002;416:199–202. doi: 10.1038/nature726. [DOI] [PubMed] [Google Scholar]
- Nelson W, Halberg F. An evaluation of time-dependent changes in susceptibility of mice to pentobarbital injection. Neuropharmacology. 1973;12:509–524. doi: 10.1016/0028-3908(73)90002-6. [DOI] [PubMed] [Google Scholar]
- Ninomiya Y, Tonosaki K, Funakoshi M. Gustatory neural response in the mouse. Brain Res. 1982;244:370–373. doi: 10.1016/0006-8993(82)90100-7. [DOI] [PubMed] [Google Scholar]
- Ninomiya Y, Higashi T, Katsukawa H, Mizukoshi T, Funakoshi M. Qualitative discrimination of gustatory stimuli in three different strains of mice. Brain Res. 1984a;322:83–92. doi: 10.1016/0006-8993(84)91183-1. [DOI] [PubMed] [Google Scholar]
- Ninomiya Y, Mizukoshi T, Higashi T, Katsukawa H, Funakoshi M. Gustatory neural responses in three different strains of mice. Brain Res. 1984b;302:305–314. doi: 10.1016/0006-8993(84)90244-0. [DOI] [PubMed] [Google Scholar]
- Ohkuri T, Horio N, Stratford JM, Finger TE, Ninomiya Y. Residual chemoresponsiveness to acids in the superior laryngeal nerve in “taste-blind” (P2X2/P2X3 double-KO) mice. Chem Senses. 2012;37:523–532. doi: 10.1093/chemse/bjs004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oka Y, Butnaru M, von Buchholtz L, Ryba NJ, Zuker CS. High salt recruits aversive taste pathways. Nature. 2013;494:472–475. doi: 10.1038/nature11905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Romanov RA, Rogachevskaja OA, Bystrova MF, Jiang P, Margolskee RF, Kolesnikov SS. Afferent neurotransmission mediated by hemichannels in mammalian taste cells. EMBO J. 2007;26:657–667. doi: 10.1038/sj.emboj.7601526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roper SD. Taste buds as peripheral chemosensory processors. Semin Cell Dev Biol. 2013;24:71–79. doi: 10.1016/j.semcdb.2012.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rüsch D, Braun HA, Wulf H, Schuster A, Raines DE. Inhibition of human 5-HT3A and 5-HT3AB receptors by etomidate, propofol and pentobarbital. Eur J Pharmacol. 2007;573:60–64. doi: 10.1016/j.ejphar.2007.07.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shin YK, Martin B, Golden E, Dotson CD, Maudsley S, Kim W, Jang HJ, Mattson MP, Drucker DJ, Egan JM, Munger SD. Modulation of taste sensitivity by GLP-1 signaling. J Neurochem. 2008;106:455–463. doi: 10.1111/j.1471-4159.2008.05397.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takai S, Yasumatsu K, Inoue M, Iwata S, Yoshida R, Shigemura N, Yanagawa Y, Drucker DJ, Margolskee RF, Ninomiya Y. Glucagon-like peptide-1 is specifically involved in sweet taste transmission. FASEB J. 2015;29:2268–2280. doi: 10.1096/fj.14-265355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomchik SM, Berg S, Kim JW, Chaudhari N, Roper SD. Breadth of tuning and taste coding in mammalian taste buds. J Neurosci. 2007;27:10840–10848. doi: 10.1523/JNEUROSCI.1863-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Buskirk C. The seventh nerve complex. J Comp Neurol. 1945;82:303–333. doi: 10.1002/cne.900820306. [DOI] [Google Scholar]
- Vandenbeuch A, Anderson CB, Parnes J, Enjyoji K, Robson SC, Finger TE, Kinnamon SC. Role of the ectonucleotidase NTPDase2 in taste bud function. Proc Natl Acad Sci U S A. 2013;110:14789–14794. doi: 10.1073/pnas.1309468110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vandenbeuch A, Larson ED, Anderson CB, Smith SA, Ford AP, Finger TE, Kinnamon SC. Postsynaptic P2×3-containing receptors in gustatory nerve fibres mediate responses to all taste qualities in mice. J Physiol. 2015;593:1113–1125. doi: 10.1113/jphysiol.2014.281014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Z-Y, Keith IM, Olson EB, Jr, Vidruk EH, Bisgard GE. Expression of 5-HT3 receptors in primary sensory neurons of the petrosal ganglion of adult rats. Auton Neurosci. 2002;95:121–124. doi: 10.1016/S1566-0702(01)00384-8. [DOI] [PubMed] [Google Scholar]
- Wu A, Dvoryanchikov G, Pereira E, Chaudhari N, Roper SD. Breadth of tuning in taste afferent neurons varies with stimulus strength. Nat Commun. 2015;6:8171. doi: 10.1038/ncomms9171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yee CL, Yang R, Böttger B, Finger TE, Kinnamon JC. “Type III” cells of rat taste buds: Immunohistochemical and ultrastructural studies of neuron-specific enolase, protein gene product 9.5, and serotonin. J Comp Neurol. 2001;440:97–108. doi: 10.1002/cne.1372. [DOI] [PubMed] [Google Scholar]
- Zeitz KP, Guy N, Malmberg AB, Dirajlal S, Martin WJ, Sun L, Bonhaus DW, Stucky CL, Julius D, Basbaum AI. The 5-HT3 subtype of serotonin receptor contributes to nociceptive processing via a novel subset of myelinated and unmyelinated nociceptors. J Neurosci. 2002;22:1010–1019. doi: 10.1523/JNEUROSCI.22-03-01010.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]