

Abstract
Key to understanding the implications of climate and land use change on biodiversity and natural resources is to incorporate the physiographic platform on which changes in ecological systems unfold. Here, we advance a detailed classification and high-resolution map of physiography, built by combining landforms and lithology (soil parent material) at multiple spatial scales. We used only relatively static abiotic variables (i.e., excluded climatic and biotic factors) to prevent confounding current ecological patterns and processes with enduring landscape features, and to make the physiographic classification more interpretable for climate adaptation planning. We generated novel spatial databases for 15 landform and 269 physiographic types across the conterminous United States of America. We examined their potential use by natural resource managers by placing them within a contemporary climate change adaptation framework, and found our physiographic databases could play key roles in four of seven general adaptation strategies. We also calculated correlations with common empirical measures of biodiversity to examine the degree to which the physiographic setting explains various aspects of current biodiversity patterns. Additionally, we evaluated the relationship between landform diversity and measures of climate change to explore how changes may unfold across a geophysical template. We found landform types are particularly sensitive to spatial scale, and so we recommend using high-resolution datasets when possible, as well as generating metrics using multiple neighborhood sizes to both minimize and characterize potential unknown biases. We illustrate how our work can inform current strategies for climate change adaptation. The analytical framework and classification of landforms and parent material are easily extendable to other geographies and may be used to promote climate change adaptation in other settings.
Introduction
The science underpinning climate adaptation planning is nascent and complex, but recent guidance emphasizes the need to characterize environmental heterogeneity that underlies a variety of strategies to conserve ecological patterns and processes in the face of climate change (e.g., [1,2,3,4]). One of the increasingly used strategies is to characterize environmental heterogeneity by identifying “arenas of biological activity,” “land facets” or physiographic settings (e.g., [5,6,7]). Of the general adaptation strategies identified by climate-smart conservation [3], protecting different components of environmental heterogeneity forms the basis for at least four of them. One key strategy is to protect key ecosystem features ([4,8,9,10,11]), because areas with high environmental heterogeneity support high genetic and species diversity ([3,5,12,13,14,15,16), including rare plant species of conservation concern [7]. A second strategy is to support evolutionary potential by representing a variety of environmental settings, which facilitates adaptation of populations and reassembly of communities [17], and species’ responses to climate change [18,19]. A third strategy is to protect refugia—places that are less likely to be influenced by climate change because site-level climate conditions are decoupled from changes in regional climate [20]. A fourth strategy is to ensure connectivity among different habitat types and physiographic settings, and many of the landscape features such as riparian zones that are important for connectivity are strongly aligned with certain landforms (e.g., valley bottoms).
Here we describe the development of a novel classification and maps of landforms and physiography to be used as foundational data for ecological modeling of species and biodiversity to inform climate adaptation planning activities. We recognize that physiographic settings can be considered over a variety of spatial and temporal scales, and that some landforms (especially locally) can change due to extreme events, such as flash floods or earthquakes. This has led scientists to view geomorphology as non-linear and dynamic [21]. Yet, here we generalize these dynamics to emphasize geophysical settings that occur over broad spatial extents (>100 km2) and change, on average, at relatively slow time scales (centuries to millennia). In comparison to the dramatic climatic and biotic responses that are forecast to occur due to climate change [22], we view these settings as relatively static. Our characterization of landforms confers a practical advantage for climate adaptation because it isolates those features that will likely remain on the landscape and allow managers to examine and focus on the relatively high uncertainty in dynamic variables, such as climate model projections of temperature or precipitation.
Seminal work by ecologists has long recognized that landforms [23] structure ecological patterns and processes (e.g., [24–27]). More recently, scientists have brought this concept into conservation planning by identifying land facets [6,28], abiotic units [29–32], geodiversity [33,34], geomorphological units [35], and the geophysical stage [7,36]. These physical features have the potential to inform climate adaptation strategies by helping characterize and interpret how species’ distributions are locally influenced by climate and other broad scale dynamic factors. Importantly, not all species are expected to interact with the physiographic setting in the same way. Species’ ecological niches (i.e., the environmental conditions and resources required by species for survival) are shaped in part by biophysical variables (e.g., temperature), which are in turn shaped by physical variables that define the physiographic setting [37, 38]. Physical variables are broadly measurable at the topographic and even micro scales at which many species interact with the environment. Hence, they are important for understanding how species’ ecological niches translate geographically into fine-scale distributions within the physiographic setting. Such resolution issues pertaining to scale have an immense effect on identifying climate change refugia and opportunities for maintaining connectivity [18]. In addition, the influence of physiographic setting on the fine-scale distributions of plant species further determines habitat composition and structure for many animal species. Combined, the fine-scale patterning and partitioning of species distributions within a physiographic setting affect overall richness and the likelihood of stochastic extirpation through the area-heterogeneity tradeoff [39].
Our work to characterize environmental heterogeneity was guided by three criteria. First, we excluded factors that have been used to define physiographic units [7,28,29,40] that change relatively rapidly, such as climate and biotic interactions. Instead, we used only physical factors that remain relatively stable over centuries to millennia (solar insolation, hillslope position, lithology), recognizing that these physical factors are ultimately shaped by tectonic processes, climate, and biota over longer time scales [41]. The advantages of characterizing landforms and physiographic diversity solely on relatively static, physical variables are that: (a) the ecological interpretation is straightforward and parsimonious because solar insolation, hillslope position, and lithology underlie dynamic processes; (b) it explicitly avoids confounding classification thresholds with contemporary patterns defined on the basis of climatological and biotic distributions, which may change dramatically and in unprecedented ways over a management planning time horizon (10–100 years; [42]); and (c) it provides a basis to evaluate the proportion of current biological and environmental diversity that is explained by stable factors that will not change over a planning time horizon, thus offering a baseline for interpreting the magnitudes and uncertainties of future change.
A second guiding criterion was that estimates of physiographic diversity are sensitive to spatial resolution and extent [43], so we derived landforms using a multi-scale analysis [44]. The multi-scale approach informs a wider range of ecological questions by identifying the scale(s) at which particular physiographic settings are defined and experienced by individual species. Fine-grained features can be locally and regionally important [45] and so the resolution of the data should be as detailed as practically possible to assist managers in making decisions at a local scale, typically identified to be at roughly 10–90 m. Landforms generated from high-resolution datasets are especially important as a way to better “downscale” typical climate-based outputs for ecological applications, which tend to be too coarse to be operational at the local level.
A final guiding criterion was the need to make these data relevant to natural resource managers by generating a parsimonious, easy to explain and interpretable model [46]. Moreover, because conservation in the era of rapid climate and land use change forces us to plan at broader extents [47], we wanted to provide comprehensive data that are consistent across broadly defined regions, such as the U.S. Department of Interior Landscape Conservation Cooperatives (LCCs; [48]).
Our goal in this paper is to describe an ecologically relevant classification and map of landforms and physiographic classes that are suitable for climate adaptation planning. We examined the sensitivity of our landform classes to changes in spatial resolution by comparing datasets generated at 10, 30, 90, 270, and 810 m resolution. We illustrate the utility of our datasets by calculating the amount of explanatory power they have against other common indicators of biodiversity patterns and spatial climatic gradients. We also summarized the patterns of landforms and physiography across the conterminous USA, including by LCCs and conduct a “gap analysis” [49] of level of land protection for landform types. Because there is large uncertainty associated with future climate conditions and even more uncertainty around ecological responses, which likely will include novel communities, we believe providing information about what is likely not to change offers a strong foundation for managers to build robust climate adaptation plans.
Materials and Methods
We generated three related maps: (1) landforms; (2) physiography (landforms combined with lithology); and (3) physiographic diversity.
Landforms
We developed a comprehensive classification of landforms based on hillslope position and dominant physical processes. Following work articulating hillslope and soil formation (aka topographic sequences; [50,51]), we distinguished four hillslope positions that form a natural sequence of topographic units along the catena: ridges/peaks (summits), upper slopes (shoulders), lower slopes (foot slopes), and valley bottoms (toe slopes). Next, we differentiated the position within each of these hillslopes as a function of solar orientation to reflect how ecological processes (especially soil moisture and evapotranspiration) are strongly influenced by the intensity of insolation and/or shading [24]. Finally, we identified features at the extremes of hillslope gradients, including areas that are very flat (i.e. areas <2°) or very steep (i.e. “cliffs” >50°).
To quantify hillslope position, we calculated a multi-scale topographic position index (TPI) that measures relative topographic relief [52,53]:
where E 0 is the elevation in meters at a given location (or cell) from the National Elevation Dataset (originally at 10 m resolution, for the contiguous USA; excluding Alaska, Hawaii, and territories) [54], and E n is the mean elevation of all cells within a neighborhood specified by radius r. Highly positive values are associated with peaks and ridges, while highly negative values are associated with valley bottoms and sinks. Locations with values approaching zero typically occur on uniform hillslopes or on flat lands (e.g., plateaus). We standardized TPI following [55] to better resolve fine-scale features:
where E s is the standard deviation of elevation for all cells within a neighborhood specified by radius r. Previously, TPI has been calculated for broad regions using only a single neighborhood, such as a radius of 200 [6], 500 [56], or 564 m globally [57]. So that our analysis would be less sensitive to a particular scale and following [52], we calculated TPIs at multiple scales, selecting neighborhood sizes that would differentiate relatively small geomorphological features such as local hills or ridges from large mountain peaks and divides, and large, broad valleys across the USA. That is, we calculated TPI using radii (r) of 270, 810, and 2430 m using a standard progression [58] in multiples of 3 from the base 10 meter native resolution of the DEM that scales roughly in order of magnitude of area and nest with other data used in this analysis (e.g., derived from Landsat imagery at 30 m). Initially we explored additional larger radii (7.2 and 21.8 km), but found they did not discriminate fine-grain (<1 km) features.
We calculated mTPIs as:
The break between the upper slope and mid-slope is found where mTPIs = 0. Hills were differentiated as upper slopes that had between 30–300 m relief (measured as the elevation gain from its base to highest point; [59]). Ridges/peaks were at least 300 m, and mountains or divides (ridges or peaks that have regional significance) also had at least 300 m relief, but for TPIs where r = 2430 m.
Valley bottoms were distinguished from lower slopes where mTPIs < -0.75, adjusted from -1.0 to reflect the asymmetrical distribution of elevation values (due to gravity). Narrow and relatively deep valleys (canyons) were found where mTPIs < -1.2 and the absolute relief exceeded 5 meters [60]. We also used slope to distinguish two special classes: cliffs were identified as those locations with greater than 50° slope, and flats were identified as areas that have slopes less than 2° when calculated ~810 m resolution to filters out small “speckles” and stripes that can occur from artifacts in digital elevation models. We confirmed these thresholds by visually comparing cliff and flat landforms to 30 m datasets on hillshade and delineated geomorphological types from a high resolution soils database (http://tinyurl.com/ssurgo), at 3 random locations in nearly 60 ecoregions across the USA.
We used an estimate of incident radiation and heat load [61] that combines slope, aspect, and latitude to predict the ecological effects of potential direct radiation. In contrast to other methods that have relied on aspect [7,36] or slope and solar insolation [52,62], heat load has a strong empirical relationship to evapotranspiration that controls vegetation distributions [56,61] and can be readily calculated at broad extents and high resolution (30–90 m).
We modified the original heat-load index [61] in two ways to construct a continuous heat-insolation load index (CHILI). First, we modified the formula to explicitly incorporate latitude as a variable so that heat load is calculated on a continuous (latitudinal) basis, assuming insolation at the equinox [52]. Second, the original heat-load index is based on a 45° “folding” to mimic patterns of evapotranspiration and measured heat load that increase in the afternoon. We modified the folding to 22.5°, based on more recent empirical studies that found that thermal south ranged from ~10° to 30° west of south over the growing season [63], and mean maximum daily temperatures were reached at ~20°-45° west of south [64]. We did not include assumptions that might influence the amount of radiation due to atmospheric conditions (e.g., due to clouds) or other specific assumptions about growing seasons.
We classified the heat load index to distinguish “warm”, “neutral”, and “cool” portions of a landscape. We established these classes from the CHILI values (warm > 0.767, cool <0.448, neutral 0.448–0.767) by finding the thresholds that resulted in equal areas of each class on a simulated landscape. We did this using an idealized mountain formed by a Gaussian surface, where we controlled the height (i.e., population) to width (i.e., bandwidth) of a kernel density function to minimize areas with either >50° (“cliff”) or <2° (flat). The warm/neutral/cool patterns generated from the thresholds aligned well with independent estimates for a range of physiographic provinces across the USA.
Our final comprehensive classification of landforms consists of 15 classes (Table 1). We implemented the landform algorithm (S1 Fig) using the Google Earth Engine platform [65], at five resolutions (or grains: 10, 30, 90, 270, and 810 m) using a 10 m USGS DEM [54].
Table 1. A hierarchical classification of ecologically relevant landforms.
| Hillslope position | ID | Class name | TPI | Slope (°) | CHILI |
|---|---|---|---|---|---|
| Summit | 11 | Peak/ridge warm | (0.0 < mTPIs < 1.0) and (30 < (E o -E n) < 300) | Warm | |
| Summit | 12 | Peak/ridge | (0.0 < mTPIs < 1.0) and (30 < (E o -E n) < 300) | Neutral | |
| Summit | 13 | Peak/ridge cool | (0.0 < mTPIs < 1.0) and (30 < (E o -E n) < 300) | Cool | |
| Summit | 14 | Mountain/divide* | (0.0 < mTPIs < 1.0) and ((E o -E n) ≥ 300) | ||
| Summit | 15 | Cliff | >50 | ||
| Upper slope | 21 | Upper slope warm | (0.0 < mTPIs < 1.0) and ((E o -E n) ≤ 30) | Warm | |
| Upper slope | 22 | Upper slope neutral | (0.0 < mTPIs < 1.0) and ((E o -E n) ≤ 30) | Neutral | |
| Upper slope | 23 | Upper slope cool | (0.0 < mTPIs < 1.0) and ((E o -E n) ≤ 30) | Cool | |
| Upper slope | 24 | Upper slope flat | (0.0 < mTPIs < 1.0) and ((E o -E n) ≤ 30) | <2 | |
| Lower slope | 31 | Lower slope warm | (-0.75 < mTPIs < 0.0) and ((E o -E n) > -5) | Warm | |
| Lower slope | 32 | Lower slope neutral | (-0.75 < mTPIs < 0.0) and ((E o -E n) > -5) | Neutral | |
| Lower slope | 33 | Lower slope cool | (-0.75 < mTPIs < 0.0) and ((E o -E n) > -5) | Cool | |
| Lower slope | 34 | Lower slope flat | (-0.75 < mTPIs < 0.0) and ((E o -E n) > -5) | <2 | |
| Valley bottom | 41 | Valley | (mTPIs < -0.75) | ||
| Valley bottom | 42 | Valley (narrow) | (mTPIs < -1.2) and ((E o -E n) ≤ -5) |
Classes were based on dominant hillslope position and defined using the topographic position index (TPI), slope, and continuous heat load index (CHILI).
*Difference in elevation calculated at r = 2430, others are calculated at r = 810. ID is the unique identifier used to label each class in the landform dataset.
Lithology
We used the best-available national-level dataset on lithology [66], which specifies 18 ecologically relevant parent material classes defined by textural and chemical similarities and mapped as polygons consistently across the USA (1:1,000,000). Rather than soil class information, we used geologic parent material because of the strong linkage between these characteristics and ecological response [7]. That is, in addition to differentiating crude soil texture categories, lithology incorporates differences in chemical properties found to be important to vegetation. We converted the polygons to a 270 m resolution raster to differentiate linear, narrow (~540 m in width) polygons. We then overlayed the lithology classes with the land form classes, generating the physiography dataset at 270 m resolution. We explored the higher spatial resolution SSURGO (http://tinyurl.com/ssurgo) and STATSGO (http://www.tinyurl.com/statsgo) soils databases, but these had numerous inconsistencies in classifications, were incomplete in roughly 20% of the USA (with higher proportions of missing classes in terrain with higher relief), and had attributes on parent material available for only roughly 10% of the USA.
Multi-scale diversity
We estimated environmental heterogeneity by calculating the diversity of physiographic units using Shannon’s equitability (E H; 0 to 1), which is calculated by normalizing the Shannon-Weaver diversity index (H):
where p i is the proportion of observations (cells) of type i in a given neighborhood and S is the number of physiographic types. We used a multi-scale approach to calculate the diversity of types using moving-window neighborhoods based on the average area of hydrologic unit codes 4, 6, 8, 10 12, 14, and 16 (i.e. 115.8, 89.9, 35.5, 13.1, 5.6, 2.8, and 1.2 km radius). We used the area of hydrologic units as the basis for the multi-scale moving window analysis because they are hierarchically organized and represent ecologically relevant scales [67]. Note that the resolution (or grain) of the diversity dataset is 270 m based on the physiography data layer described above. We conducted these spatial analyses using ArcGIS v10 (Esri 2014, Redlands, CA).
Relationship with existing indicators
To examine some of the linkages between the physiographic setting and climate adaptation strategies, in accordance with our starting criteria, we asked a series of questions that could be answered using a combination of our landform and physiographic diversity datasets, as well as others commonly used to understand patterns of biodiversity and climatic variation. Our first question related strongly to the strategy to protect key ecosystem features: What proportion of variation in biodiversity is explained by variation in physiographic setting? To assess the relationship with patterns of biodiversity, we examined physiographic diversity in relation to vertebrate species richness [68] and ecological system richness [69]. Range maps for USA mammals (91 species) and amphibians (n = 48) and bird breeding ranges (n = 200) were compiled at 10 km resolution. We calculated richness as the multi-scale average of the number of species using the same watershed-based scales described earlier. The multi-scale average of the number of ecological systems (579 vegetation types) was calculated from 30 m resolution data [67].
Because the environmental heterogeneity that influences key ecosystem features is influenced by a combination of climatic and physiographic factors, we posed a second related question: What proportion of variation in current meso-scale climate (840 m resolution) was explained by variation in physiographic setting? We answered this question in two ways. First, to assess the relationship between our physiographic datasets and meso-climatic variation, we compared landform diversity against historical temperature variability. Landform diversity was calculated as the average of the landform E H (at 840 m resolution) across moving windows for the multi-scale watersheds. Temperature variability was calculated as the multi-scale average of the standard deviation in historical (1950–2000) annual mean temperature from WorldClim [70], also at 840 m resolution. Second, we compared spatial variation between mTPI and historical estimates of meso-climate from WorldClim, where slope (rise/run) was calculated using the average maximum technique [22]. We calculated slopes on the raw mTPI values (before classifying into slope position categories; m/km) and the denominator of climate velocity (°C/km).
In answering the above questions, we used Pearson’s product-moment correlation coefficients [71] to assess how much variation in the indicator response variables was explained by the landform and physiographic predictor variables. Our interest was to simply quantify the proportion of current environmental heterogeneity that is explained by stable factors that change relatively little over a planning time horizon.
Results
We first analyzed how our landform classes were sensitive to resolution, and found that general trends in class proportions were consistent throughout the USA over a range of scales (resolution), but strong differences for classes that occupied small portions of the USA (Table 2) were evident as well. We found a more than 2-fold difference for: hill/ridge (cool) and cliff (30:90 m ratio); upper slope (cool) and lower slope (cool) (30:270 m ratio); and upper slope (warm), upper slope, and lower slope (warm). Interestingly, we found that “cool” landforms that are likely to be least affected by warming climates [19] were particularly sensitive to resolution and are under-represented at coarser-resolutions. The valley bottom (narrow) class showed directional changes, first slightly decreasing in representation as a function of coarser resolution, but at 810 m exhibited a 25% increase in representation (compared to 30 m).
Table 2. Landform classes in the conterminous USA at different resolutions.
| Percent in each class at resolution (m) | Ratio | ||||||
|---|---|---|---|---|---|---|---|
| Name | 30 | 90 | 270 | 810 | 30:90 | 30:270 | 30:810 |
| Hill/ridge (warm) | 0.31% | 0.31% | 0.38% | 0.48% | 1.00 | -1.24 | -1.57 |
| Hill/ridge | 0.69% | 0.73% | 0.66% | 0.64% | 0.94 | 1.03 | 1.07 |
| Hill/ridge (cool) | 0.05% | 0.01% | 0.00% | 0.00% | 4.03 | 51.51 | 6318.84 |
| Peak/divide | 0.05% | 0.05% | 0.05% | 0.07% | -1.01 | -1.02 | -1.51 |
| Cliff | 0.07% | 0.02% | 0.01% | 0.00% | 3.18 | 11.68 | n/a |
| Upper slope (warm) | 12.40% | 11.39% | 9.34% | 5.04% | 1.09 | 1.33 | 2.46 |
| Upper slope | 14.32% | 13.05% | 11.30% | 7.05% | 1.10 | 1.27 | 2.03 |
| Upper slope (cool) | 0.69% | 0.39% | 0.13% | 0.01% | 1.77 | 5.48 | 96.50 |
| Upper slope (flat) | 18.19% | 19.61% | 23.41% | 33.50% | -1.08 | -1.29 | -1.84 |
| Lower slope (warm) | 12.23% | 11.04% | 9.30% | 5.80% | 1.11 | 1.31 | 2.11 |
| Lower slope | 10.99% | 10.29% | 9.96% | 6.84% | 1.07 | 1.10 | 1.61 |
| Lower slope (cool) | 0.41% | 0.23% | 0.09% | 0.00% | 1.80 | 4.70 | 83.54 |
| Lower slope (flat) | 20.16% | 23.45% | 25.80% | 28.75% | -1.16 | -1.28 | -1.43 |
| Valley bottom | 8.29% | 8.30% | 8.48% | 10.41% | 1.00 | -1.02 | -1.25 |
| Valley bott. (narrow) | 1.14% | 1.13% | 1.09% | 1.43% | 1.01 | 1.05 | -1.25 |
This table shows the proportion of the conterminous USA by landform class, as a function of resolution.
Because we found that landforms were clearly sensitive to spatial resolution, we provide our results below based on the 30 m resolution dataset. Some general patterns are evident by examining the variation in the proportion of each class within each LCC across the USA (Fig 1A; Table 3), but landforms also exhibit complex local patterns that affect climate heterogeneity (Fig 1B). By hillslope position, we found that 1.16% of the conterminous USA is considered to be peaks/divides and hills/ridges, 45.61% on upper slopes, 43.79% on lower slopes, and 9.44% in valley bottoms. “Neutral” heat load dominates most hillslope position classes. As a proportion of the USA, hills/ridges had 0.69% in neutral heat settings, while 0.31% was warm and only 0.05% is cool. Upper slopes were dominated as well by “neutral” heat load situations (with 14% neutral and 18% flat, with 12% “warm” and 0.7% “cool”). Lower slopes also were dominated by neutral heat loads with 11% neutral and 20% flat, with 12% warm and 0.4% cool. Finally, deep and narrow valleys cover 1.14%, while broad valley bottoms cover 8.29% of the USA.
Fig 1. Landforms of the conterminous USA.

(A) A landform map of the USA, with Landscape Conservation Cooperatives used by the Department of Interior to guide climate change adaptation. Labels a-h refer to inset examples and legend for class types. (B) Examples of landform classes, zoomed in to illustrate different patterns: (a) the Pacific Northwest around Mount St. Helens (1:175,000); (b) along the Missouri River at the boundary of Montana and North Dakota (1:200,000); (c) near Milton, Pennsylvania (1:500,000); (d) in the Sky Islands of southern Arizona (1:500,000); (e) Estes Park, Colorado (1:175,000); (f) near Smithfield, North Carolina (1:400,000); (g) along the Ogeechee River near Statesboro, Georgia (1:300,000); and (h) south Texas tablelands (1:200,000).
Table 3. Ecoregional distribution of landforms.
| Area | Summit | Upper slope | Lower slope | Valley bottom | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LCC | (km2) | Peak/ridge warm | Peak/ridge neutral | Peak/ridge cool | Mtn/divide | Cliff | Warm | Neutral | Cool | Flat | Warm | Neutral | Cool | Flat | Valley | Narrow |
| North Pacific | 179,408 | 0.369 | 2.885 | 0.100 | 0.278 | 0.027 | 10.02 | 27.98 | 2.79 | 2.80 | 11.20 | 24.61 | 1.61 | 5.77 | 8.33 | 1.23 |
| California | 210,173 | 1.046 | 1.384 | 0.007 | 0.125 | 0.039 | 18.21 | 13.32 | 0.34 | 9.21 | 21.17 | 10.00 | 0.22 | 16.81 | 7.38 | 0.74 |
| Great Northern | 660,379 | 0.124 | 1.791 | 0.098 | 0.245 | 0.087 | 6.48 | 30.04 | 2.92 | 4.89 | 7.43 | 27.12 | 1.85 | 8.83 | 7.18 | 0.92 |
| Great Basin | 562,700 | 0.319 | 1.133 | 0.009 | 0.070 | 0.010 | 8.49 | 17.12 | 0.35 | 9.90 | 13.34 | 17.68 | 0.15 | 26.63 | 4.43 | 0.36 |
| Gulf Coast Prairie | 382,730 | 0.141 | 0.015 | 0.000 | 0.000 | 0.000 | 12.10 | 0.11 | 0.00 | 34.28 | 9.28 | 0.04 | 0.00 | 34.07 | 9.20 | 0.76 |
| Southern Rockies | 516,755 | 0.645 | 1.136 | 0.016 | 0.103 | 0.148 | 21.34 | 14.85 | 0.47 | 7.29 | 22.17 | 11.75 | 0.23 | 12.51 | 6.48 | 0.86 |
| Plains and Prairie Potholes | 783,896 | 0.008 | 0.265 | 0.002 | 0.001 | 0.000 | 0.44 | 20.34 | 0.02 | 25.21 | 0.57 | 15.93 | 0.01 | 27.38 | 9.00 | 0.82 |
| Great Plains | 782,004 | 0.041 | 0.054 | 0.000 | 0.000 | 0.000 | 7.44 | 6.12 | 0.00 | 34.42 | 6.44 | 3.87 | 0.00 | 32.90 | 7.77 | 0.93 |
| Upper Midwest & Great Lakes | 540,064 | 0.011 | 0.226 | 0.000 | 0.001 | 0.000 | 0.88 | 15.70 | 0.00 | 28.55 | 0.90 | 10.22 | 0.00 | 32.85 | 9.44 | 1.20 |
| Eastern T. Prairie & Big Rivers | 535,630 | 0.010 | 0.026 | 0.000 | 0.000 | 0.000 | 6.30 | 11.15 | 0.00 | 31.46 | 5.41 | 6.06 | 0.00 | 28.30 | 9.84 | 1.45 |
| Gulf Coast Prairie | 382,730 | 0.141 | 0.015 | 0.000 | 0.000 | 0.000 | 12.10 | 0.11 | 0.00 | 34.28 | 9.28 | 0.04 | 0.00 | 34.07 | 9.20 | 0.76 |
| Gulf Coastal Plains and Ozarks | 728,639 | 0.234 | 0.150 | 0.000 | 0.000 | 0.000 | 20.66 | 2.12 | 0.00 | 22.17 | 14.30 | 1.07 | 0.00 | 26.98 | 10.71 | 1.61 |
| Appalachian | 592,906 | 1.007 | 2.061 | 0.001 | 0.007 | 0.000 | 22.80 | 17.66 | 0.06 | 4.36 | 20.20 | 11.45 | 0.02 | 6.81 | 11.08 | 2.50 |
| North Atlantic | 289,740 | 0.146 | 1.127 | 0.006 | 0.009 | 0.000 | 8.01 | 23.51 | 0.10 | 13.02 | 6.96 | 20.53 | 0.02 | 16.52 | 8.80 | 1.24 |
| South Atlantic | 357,847 | 0.060 | 0.041 | 0.000 | 0.000 | 0.000 | 18.96 | 0.45 | 0.00 | 29.80 | 13.38 | 0.20 | 0.00 | 24.91 | 10.08 | 2.12 |
| Peninsular Florida | 93,383 | 0.002 | 0.000 | 0.000 | 0.001 | 0.000 | 0.73 | 0.00 | 0.00 | 43.81 | 0.36 | 0.00 | 0.00 | 44.48 | 9.59 | 1.04 |
This table shows the percentage of each landform class in the conterminous USA by Landscape Conservation Cooperative ecoregions (LCC).
We overlaid the 15 landform types with 18 classes of soil parent material, resulting in 269 unique combinations of physiographic types (270 possible; Fig 2; Table 4). The multi-scale physiographic E H averaged 0.382 (SD = 0.068), with high mean values located in the North Pacific (0.671), Great Northern (0.574), and Southern Rockies (0.526), and low mean values located in Peninsular Florida (0.120), South Atlantic (0.314), and the Gulf Coast Prairie (0.321), and the Great Plains (0.347). Strong patterns were evident and coincide with major mountain ranges of the Cascades, Bitterroot/Absaroka, Sierra Nevada, Southern Rockies, and the Appalachians. High diversity also occured in areas with major river systems, throughout the Midwest in particular.
Fig 2. Physiographic diversity in the conterminous USA.

Multi-scale physiographic diversity, calculated using the Shannon-Weaver index. Labels are mean diversity by Landscape Conservation Cooperatives, with standard deviations in parentheses.
Table 4. Parent soil material types by landform class in the conterminous USA.
| Summit | Peak/ridge | Upper slope | Lower slope | Valley bottom | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parent material | Mtn/divide | Cool | Neutral | Warm | Cliff | Flat | Cool | Neutral | Warm | Flat | Cool | Neutral | Warm | Valley | Narrow | % |
| Water areas | 0 | 116 | 2 | 2 | 0 | 2,756 | 2,446 | 39 | 6,663 | 4,738 | 4,652 | 70 | 44,408 | 5,819 | 300 | 2.84% |
| Carbonate | 3,041 | 3,821 | 69 | 427 | 436 | 107,376 | 39,875 | 2,397 | 56,633 | 87,812 | 26,630 | 1,313 | 53,079 | 38,899 | 6,949 | 5.38% |
| Non-carbonate | 6,470 | 17,207 | 413 | 1,103 | 689 | 301,360 | 310,013 | 10,669 | 266,030 | 266,910 | 250,832 | 5,696 | 301,094 | 158,182 | 21,688 | 24.12% |
| Alkaline intrusive | 30 | 86 | 0 | 6 | 1 | 1,285 | 3,635 | 23 | 865 | 1,280 | 2,753 | 14 | 1,093 | 645 | 140 | 0.15% |
| Silicic residual | 7,051 | 16,749 | 460 | 1,981 | 531 | 175,353 | 194,703 | 15,058 | 28,853 | 175,545 | 147,859 | 8,997 | 26,370 | 60,104 | 10,510 | 10.94% |
| Extrusive volcanic | 212 | 552 | 12 | 32 | 10 | 7,400 | 13,656 | 289 | 17,639 | 7,274 | 11,073 | 153 | 22,492 | 3,433 | 289 | 1.06% |
| Colluvial sediment | 5,098 | 9,865 | 5 | 38 | 4 | 130,028 | 102,240 | 283 | 56,663 | 112,038 | 62,194 | 109 | 54,174 | 63,951 | 13,855 | 7.68% |
| Glacial till—clay | 3 | 29 | 0 | 0 | 0 | 411 | 5,053 | 0 | 42,074 | 437 | 3,339 | 0 | 38,469 | 8,079 | 784 | 1.24% |
| Glacial till loamy | 251 | 2,090 | 6 | 7 | 1 | 26,116 | 133,142 | 171 | 293,759 | 23,075 | 84,497 | 91 | 229,810 | 79,783 | 10,866 | 11.12% |
| Glacial till coarse | 356 | 3,183 | 32 | 39 | 8 | 9,742 | 56,763 | 656 | 6,289 | 12,025 | 53,989 | 459 | 13,285 | 14,451 | 1,087 | 2.17% |
| Glacial lake sediment fine textured | 28 | 457 | 11 | 7 | 4 | 2,402 | 31,878 | 542 | 79,935 | 3,476 | 29,812 | 553 | 107,186 | 29,783 | 3,500 | 3.65% |
| Glacial outwash coarse textured | 20 | 184 | 2 | 1 | 1 | 1,550 | 12,353 | 40 | 40,864 | 1,713 | 10,909 | 57 | 56,492 | 13,173 | 1,130 | 1.76% |
| Hydric, peat, muck | 0 | 2 | 0 | 0 | 0 | 52 | 321 | 0 | 19,683 | 27 | 241 | 0 | 24,215 | 4,903 | 215 | 0.63% |
| Eolian sediment (coarse) | 161 | 266 | 1 | 2 | 10 | 12,200 | 19,100 | 13 | 114,581 | 11,286 | 12,020 | 4 | 113,351 | 21,676 | 2,146 | 3.86% |
| Eolian sediment (fine) | 24 | 209 | 1 | 0 | 1 | 14,876 | 40,851 | 63 | 74,135 | 11,921 | 25,201 | 28 | 56,507 | 20,675 | 3,112 | 3.11% |
| Saline lake sediment | 74 | 288 | 2 | 10 | 2 | 3,010 | 4,744 | 62 | 17,498 | 7,576 | 9,444 | 28 | 65,828 | 5,329 | 357 | 1.44% |
| Coastal fine-textured sediment | 1,136 | 1,689 | 12 | 17 | 18 | 92,631 | 48,071 | 338 | 377,499 | 134,719 | 67,369 | 410 | 598,242 | 113,265 | 10,804 | 18.08% |
| Coastal coarse-textured sediment | 1 | 6 | 0 | 0 | 0 | 369 | 63 | 0 | 31,266 | 201 | 59 | 0 | 24,049 | 5,420 | 733 | 0.79% |
| % | 0.31% | 0.73% | 0.01% | 0.05% | 0.02% | 11.39% | 13.05% | 0.39% | 19.61% | 11.04% | 10.29% | 0.23% | 23.45% | 8.30% | 1.13% | 100.00% |
This table shows the area (km2) in the conterminous USA for each type of parent soil material type by landform class.
We also examined landform classes in relation to protected areas (aka a “gap analysis” [48]; Fig 3). Not surprisingly, we found that the highest levels of protection tended to occur on peaks/divides and hills/ridges, while the lowest level of protection occurred at lower slope and valley bottoms locations. Only 5 of 15 landform types were represented at the Aichi Biodiversity Target level (17%): peak/divide, cliff, hill/ridge (cool), upper slope (cool), and lower slope (cool).
Fig 3. Level of land protection by landform class.
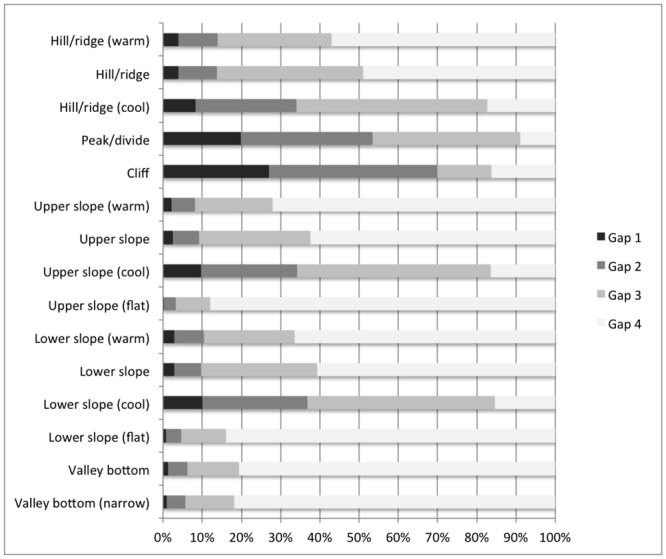
Results from a “gap” analysis of landforms showing the level of land protection for each land form class. Level of protection follows the gap status classes [48]: Status 1—permanent protection from conversion and natural disturbances; Status 2—permanent protection from conversion but may have some modification of natural disturbances; Status 3—protection from conversion for most areas but extractive uses may be allowed; and Status 4—no known formal legal mandates or restrictions to prevent conversion of natural habitat types.
We found varying levels of explanatory power among our variables and other indicators of biodiversity commonly used in climate adaptation planning. Overall, the Pearson’s product-moment correlation between physiographic diversity and vertebrate species richness was 0.449, while the correlation with ecological systems was 0.285. The strongest correlation we found was between slopes of topographic position and the denominator of climate velocity (0.792). The correlation between landform diversity (excluding parent material) and temperature variability (SD) was 0.483.
Discussion
The conterminous USA exhibits a wide range of landforms and parent material, resulting in considerable physiographic diversity. Our maps of landform classes and physiographic diversity are novel contributions that characterize ecologically relevant aspects of abiotic environmental heterogeneity. These databases provide a consistent and comprehensive platform for climate adaptation planning at local to regional and national scales.
Climate adaptation planning will likely involve pursuing multiple strategies, and below we briefly explore how landforms and physiographic diversity could inform such efforts—especially protecting key ecosystem features, ensuring connectivity, supporting evolutionary potential, and protecting refugia. Note that our intent is to provide a few illustrative examples of how our datasets can be applied within an adaptation framework, not to argue that these are the only datasets or uses. Indeed, we expect future research will further explore the use of physiographic variables for climate adaptation planning, especially to evaluate indicators and available datasets, and to compare and evaluate data gaps.
We found that our measures of physiographic diversity had moderate levels of explanatory power related to current patterns of biodiversity (0.449 for species richness), suggesting that our physiographic datasets meaningfully capture key ecosystem features that structure biodiversity and can aid coarse-filter approaches [5] to conservation planning. Our datasets may also be extended to support “fine-filter” conservation planning and vulnerability assessments focused on a particular ecosystem feature as a conservation target (e.g., an individual species or important habitat). Fine-filter approaches might instead select particular spatial extents relevant to the conservation target or particular landform or physiographic classes with which a given species has high affinity. These may be used to better understand the fine scale distribution of particular species, as well as the topographic and micro environmental influences of climate and other important environmental variables influencing a species niche. Similarly, our physiographic dataset can be leveraged to help ensure connectivity for fine-filter conservation targets by identifying potential movement pathways among physiographic classes or to identify locations of key landform types that facilitate ecological flows, such as slopes and valley bottoms [72]. Likewise, it can be used to identify locations for protecting refugia, which might be located on cool slopes or valley bottoms where cold-air pooling has potential to occur [19].
We found that the spatial rate of change in mTPI (m/km) was highly correlated with the spatial rate of change in temperature (°C/km) (0.792), suggesting that our physiographic diversity dataset is predictive of areas with strong temperature gradients. Importantly, we expect the relationship to be scale dependent [73], but also note that our mTPI was calculated over a range of biologically relevant scales (270 to 2430 m) that encompassed meso-climate (840 m). The presence and location of strong climatic and edaphic gradients identified by our physiographic diversity dataset would provide information useful for strategies aiming to support evolutionary potential, given the importance of these gradients to evolutionary processes such as diversification, speciation, and extinction [74–76].
An important application of landforms is to “place” coarser-scale climate change information to allow resource managers to understand how a particular species or ecological process might respond to climate change at a finer scale. There is a strong theoretical basis for understanding the relationship of landform types to climate [19]. Although geologic parent material has a less clear role, it can indirectly influence climate through soil characteristics (e.g., productivity and water-holding capacity) and—over shorter time scales—vegetation [77].
Not surprisingly, we found a strong scaling effect on the amount and especially pattern of landform classes as a function of resolution (Table 1. The interaction between physiography and climate is highly scale dependent [18]–and it is important to note that most climate information is still relatively coarse (~1 km is considered highly “downscaled” climate data). Our findings that landforms that are most likely to be related to climate refugia (“cool” and valley bottom) are especially sensitive to scaling suggests that work on identifying these areas should consider representation and patterning effects of scale. Recent work on micro-scale climatology has found that environmental heterogeneity varies in ecologically significant ways at horizontal scales as small as 1 m2, and vertical scales even smaller at 1 cm2 [78,79]. Currently these resolutions are too small to be mapped in a national-level database, but may be feasible for localized studies. More generally, opportunities exist to develop stronger linkages between geomorphology and vegetation by building in explicit ecological mechanisms and processes, including soils and other climate measures that may reflect physiological tolerances (e.g., maximum daily temperature), but also insolation (slope and aspect) and resulting water-balance [78]. Because the lithology dataset is a relatively coarse scale (~1:1,000,000; converted to 270 m resolution), we recommend limiting interpretation of physiographic patterns to features at least 1 km2 in size..
Many agencies and organizations are engaging in large landscape conservation and general guidance has begun to emerge on how to conduct and coordinate climate adaptation planning [2,11,30,80–82]. Although analyses of climate impacts on conservation targets are a key component of most climate adaption planning frameworks, there is little guidance on how to address broad, landscape-level impacts [18,80]. Rather, in contrast to species or habitat-level conservation, the physiographic approach provides insight into the enduring features that will continue to structure biodiversity in the face of climate change. We recognize that different regional conservation groups, such as the LCCs, will likely need to adopt different strategies for adapting to climate change, to allow for regional differences in climate change impacts, but they may consider our landform and physiographic diversity datasets as a foundation for pursuing climate adaptation strategies on where and how to invest in climate adaptation.
Supporting Information
Landforms were defined using basic topographic measures derived directly from the USGS 10 meter Digital Elevation Model, as well as latitude. Physiographic classes were generated by overlaying the landforms and lithology (parent material) converted from polygons (1:1,000,000 scale) to 270 m raster grid. The program used to generate these datasets is available from the authors upon request.
(PNG)
{kind=link}
Acknowledgments
We thank F. Davis and J. Evans for early discussions, and N. Fisichelli, P. Jantz, B. McRae and G. Schuurman for comments and suggestions that greatly improved earlier versions of the manuscript. We also appreciate the helpful comments and suggestions from two peer reviewers.
Data Availability
Datasets are available at: https://www.sciencebase.gov/catalog/item/564b4bb0e4b0ebfbef0d31d2.
Funding Statement
This work was supported by the NASA Applied Sciences program (http://www.nasa.gov/applied-sciences/) through award number 10-BIOCLIM10-0034. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. Conservation Science Partners provided support in the form of salaries for authors (DMT, DH-A, CMA), but did not have any additional role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript. The specific roles of these authors are articulated in the ‘author contributions’ section.
References
- 1. Glick P, Stein BA, Edelson NA. Scanning the Conservation Horizon: A Guide to Climate Change Vulnerability Assessment. Washington, D.C: National Wildlife Federation; 2011. [Google Scholar]
- 2. National Fish, Wildlife and Plants Climate Adaptation Partnership (NFWPCAP). National Fish, Wildlife and Plants Climate Adaptation Strategy Report from the Association of Fish and Wildlife agencies, Council on Environmental Quality, Great Lakes Indian Fish and Wildlife Commission, National Oceanic and Atmospheric Administration, and U.S. Fish and Wildlife Service; Washington, DC; 2011. [Google Scholar]
- 3. Stein B, Glick P, Edelson N, Staudt A. Climate-smart conservation: putting adaptation principles into practice. Washington, DC: National Wildlife Federation; 2014. [Google Scholar]
- 4. Schmitz OJ, Lawler JJ, Beier P, Groves C, Knight G, Boyce DA, et al. Conserving biodiversity: Practical guidance about climate change adaptation approaches in support of land-use planning. Nat Areas J. 2015; 35(1): 190–203. [Google Scholar]
- 5. Hunter ML, Jacobson GL, Webb T. Paleoecology and the coarse-filter approach to maintaining biological diversity. Conserv Biol. 1988; 2: 375–385. [Google Scholar]
- 6. Beier P, Brost B. Use of land facets to plan for climate change: conserving the arenas, not the actors. Conserv Biol. 2010; 24: 701–710. 10.1111/j.1523-1739.2009.01422.x [DOI] [PubMed] [Google Scholar]
- 7. Anderson MG, Ferree CE. Conserving the stage: climate change and the geophysical underpinnings of species diversity. PLOS ONE. 2010; e11554 10.1371/journal.pone.0011554 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Heller NE, Zavaleta ES. Biodiversity management in the face of climate change: A review of 22 years of recommendations. Biol Conserv. 2009; 142: 14–32. [Google Scholar]
- 9. Game ET, Lipsett-Moore G, Saxon E, Peterson N, Sheppard S. Incorporating climate change adaptation into national conservation assessments. Glob Chang Biol. 2011; 10.1111/j.1365-2486.2011.02457.x [DOI] [Google Scholar]
- 10. Groves CR, Game ET, Anderson MG, Cross M, Engquist C, Ferdana Z, et al. Incorporating climate change into systematic conservation planning. Biodivers Conserv. 2012; 21:1651–1671. [Google Scholar]
- 11. Stein BA, Staudt A, Cross MS, Dubois NS, Enquist C, Griffis R, et al. Preparing for and managing change: climate adaptation for biodiversity and ecosystems. Front Ecol Environ. 2013; 11(9): 502–510. [Google Scholar]
- 12. Huston MA. Biological diversity The coexistence of species on changing landscapes. Cambridge: Cambridge University Press; 1994. [Google Scholar]
- 13. Rosenzweig ML. Species diversity in space and time. Cambridge: Cambridge University Press; 1995. [Google Scholar]
- 14. Kruckeberg AR. Geology and plant life: The effects of landforms and rock types on plants. Seattle, Washington: University of Washington Press; 2004. [Google Scholar]
- 15. Vellend M, Geber MA. Connections between species diversity and genetic diversity. Ecol Lett. 2005; 8: 767–781. [Google Scholar]
- 16. Potts AJ, Hedderson TA, Cowling RM. Testing large-scale conservation corridors designed for patterns and processes: comparative phylogeography of three tree species. Divers Distrib. 2013; 19: 1418–1428. [Google Scholar]
- 17. Moritz C. Strategies to protect biological diversity and the evolutionary processes that sustain it. Syst Biol. 2002; 51: 238–254. [DOI] [PubMed] [Google Scholar]
- 18. Ackerly DD, Loarie SR, Cornwell WK, Weiss SB, Hamilton H, Branciforte R, et al. The geography of climate change: implications for conservation biogeography. Divers Distrib. 2010; 16: 476–487. [Google Scholar]
- 19. Dobrowski SZ. A climatic basis for microrefugia: the influence of terrain on climate. Glob Chang Biol. 2010; [Google Scholar]
- 20. Keppel G, van Niel KP, Wardell-Johnson GW, Yates CJ, Byrne M, Mucina L, et al. Refugia: identifying and understanding safe havens for biodiversity under climate change. Glob Ecol Biogeog. 2012; 21: 393–404. [Google Scholar]
- 21. Phillips JD. Evolutionary geomorphology: thresholds and nonlinearity in landform response to environmental change. Hydr Earth Sys Sci Disc. 2006; 3(2): 365–394. [Google Scholar]
- 22. Loarie SR, Duffy PB, Hamilton H, Asner GP, Field CB, Ackerly DD. The velocity of climate change. Nature. 2009; 462(7276): 1052–1055. 10.1038/nature08649 [DOI] [PubMed] [Google Scholar]
- 23. Hammond EH. Small-scale continental landform maps. Ann Assoc Amer Geogr. 1954; 54: 33–42. [Google Scholar]
- 24. Whittaker RH. Vegetation of the Great Smoky Mountains. Ecol Monogr. 1956; 26(1): 1–80. [Google Scholar]
- 25. Peet RK. Forest vegetation of the Colorado Front Range. Vegetatio. 1981; 45: 3–75. [Google Scholar]
- 26. Davis FW, Goetz S. Modeling vegetation pattern using digital terrain data. Landsc Ecol. 1990; 4(1): 69–80. [Google Scholar]
- 27. Iverson LR, Dale ME, Scott CT, Prasad A. A GIS-derived integrated moisture index to predict forest composition and productivity of Ohio forests (USA). Landsc Ecol. 1997; 12: 331–348. [Google Scholar]
- 28. Wessels KJ, Freitag S, van Jaarsveld AS. The use of land facets as biodiversity surrogates during reserve selection at a local scale. Biol Conserv. 1999; 89: 21–38. [Google Scholar]
- 29. Pressey RL, Cabeza M, Watts ME, Cowling RM, Wilson KA. Conservation planning in a changing world. TREE. 2007; 22: 583–592. [DOI] [PubMed] [Google Scholar]
- 30. Klausmeyer KR, Shaw MR, MacKenzie JB, Cameron DR. Landscape-scale indicators of biodiversity’s vulnerability to climate change. Ecosph. 2011; 2(8):88. [Google Scholar]
- 31. Schloss CA, Lawler JJ, Larson ER, Papendick HL, Case MJ, Evans DM, et al. Systematic conservation planning in the face of climate change on the Columbia Plateau. PLOS ONE. 2011; 10.1371/journal.pone.0028788 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Sayre R, Comer P, Warner H, Cress J. A new map of standardized terrestrial ecosystems of the conterminous United States. Washington, DC: USGS; Professional Paper 1768. 2009. [Google Scholar]
- 33. Gray M. Geodiversity. West Sussex, England: John Wiley & Sons; 2004. [Google Scholar]
- 34. Hjort J, Gordon JE, Gray M, Hunter ML Jr. Why geodiversity matters in valuing nature’s stage. Cons Biol. 2015; 29(3): 630–639. [DOI] [PubMed] [Google Scholar]
- 35. Klingseisen B, Metternicht G, Paulus G. Geomorphometric landscape analysis using a semi-automated GIS-approach. Environ Model Softw. 2008; 23(1):109–121. [Google Scholar]
- 36. Anderson M, Clark M, Sheldon AO. Estimating climate resilience for conservation across geophysical settings. Cons Biol. 2014; 28(4): 959–970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Peterson AT, Soberon J, Pearson RG, Anderson RP, Martinez-Meyer E, et al. Ecological niches and geographic distributions. Princeton, NJ: Princeton University Press; 2011. [Google Scholar]
- 38. Lawler JJ, Ackerly DD, Albano CM, Anderson MG, Dobrowski SZ, et al. The theory behind, and the challenges of, conserving nature’s stage in a time of rapid change. Conserv Biol. 2015; 29(3): 618–629. 10.1111/cobi.12505 [DOI] [PubMed] [Google Scholar]
- 39. Kadmon R, Allouche O. Integrating the effects of area, isolation, and habitat heterogeneity on species diversity: a unification of island biogeography and niche theory. Am Nat. 2007; 170(3): 443–54. [DOI] [PubMed] [Google Scholar]
- 40. Pressey RL, Hager TC, Ryan KM, Schwarz J, Wall S, Ferrier S, et al. Using abiotic data for conservation assessments over extensive regions: quantitative methods applied across New South Wales, Australia. Biol Conserv. 2000; 96: 55–82. [Google Scholar]
- 41. Geiger R, Aron RH, Todhunter P. The Climate Near the Ground. 6th Edition Lanham, MD, USA: Rowman and Littlefield Publishers; 2003. [Google Scholar]
- 42. Williams JW, Jackson ST. Novel climates, no-analog communities, and ecological surprises. Fron Ecol Environ. 2007; 5(9): 475–482. [Google Scholar]
- 43. Albano C. Identification of geophysically diverse locations that may facilitate species’ persistence and adaptation to climate change in the southwestern United States. Landsc Ecol. 2015; 10.1007/s10980-015-0167-7 [DOI] [Google Scholar]
- 44. Dufour A, Gadallah F, Wagner HH, Guisan A, Buttler A. Plant species richness and environmental heterogeneity in a mountain landscape: effects of variability and spatial configuration. Ecograph. 2006; 29: 573–584. [Google Scholar]
- 45. Lawson CR, Bennie J, Hodgson JA, Thomas CD, Wilson RJ. Topographic microclimates drive microhabitat associations at the range margin of a butterfly. Ecograph. 2014; 37: 732–740. [Google Scholar]
- 46. Theobald DM, Hobbs NT, Bearly T, Zack J, Shenk T, Riebsame WE. Incorporating biological information into local land-use decision making: Designing a system for conservation planning. Landsc Ecol. 2000; 15(1): 35–45. [Google Scholar]
- 47. Clement JP, Belin A, Bean MJ, Boling TA, Lyons JR. A strategy for improving the mitigation policies and practices of the Department of the Interior A report to the Secretary of the Interior from the Energy and Climate Change Task Force, Washington, D.C., 2014. [Google Scholar]
- 48.Secretary of the Interior. Addressing the impacts of climate change on America’s water, land, and other natural and cultural resources. Secretarial Order 3289; 2009.
- 49. Jennings MD. Gap analysis: concepts, methods, and recent results. Landsc Ecol. 2000; 15(1): 5–20. [Google Scholar]
- 50.Ruhe RV, Walker PH. Hillslope models and soil formation II: open systems. In: Proceedings of the 9th Congress of the International Soil Science Society, Adelaide, Australia, 1968; 551–560.
- 51. Dikau R. Geomorphic landform modeling based on hierarchy theory In: Brassel K, Kishimoto H. Proceedings of the 4th International Symposium on Spatial Data Handling. Zürich, Switzerland: Department of Geography, University of Zürich; 1990. pp. 230–239 [Google Scholar]
- 52. Guisan A, Weiss SB, Weiss AD. GLM versus CCA spatial modeling of plant species distribution. Plant Ecol. 1999; 143: 107–122. [Google Scholar]
- 53. Dickson BG, Beier P. Quantifying the influence of topographic position on cougar (Puma concolor) movement in southern California, USA. J Zool. 2007; 271: 270–277. [Google Scholar]
- 54. Gesch D, Oimoen M, Greenlee S, Nelson C, Steuck M, et al. The National Elevation Dataset. Photogr Eng Rem Sens. 2002; 68(1): 5–11. [Google Scholar]
- 55. De Reu J, Bourgeois J, Bats M, Zwertvaegher A, Gelorini V, De Smedt P. Application of the topographic position index to heterogeneous landscapes. Geomorphol. 2013; 186: 39–49. [Google Scholar]
- 56. Ashcroft MB, Gollan JR. Fine-resolution (25 m) topoclimatic grids of near-surface (5 cm) extreme temperatures and humidities across various habitats in a large (200 x 300 km) and diverse region. Int J Climatol. 2012; 32: 2134–2148. [Google Scholar]
- 57. Sayre R, Dangermond J, Frye C, Vaughan R, Aniello P, et al. A New Map of Global Ecological Land Units—An Ecophysiographic Stratification Approach. Washington, DC: Association of American Geographers; 2014. [Google Scholar]
- 58. Theobald DM. Estimating changes in natural landscapes from 1992 to 2030 for the conterminous United States. Lands Ecol. 2010; 25(7): 999–1011. [Google Scholar]
- 59.US Department of Agriculture, Natural Resources Conservation Service (NRCS). National soil survey handbook, part 629 –Glossary of Landform and Geologic Terms. Available: http://tinyurl.com/nsshsoils. Accessed 9 September 2015.
- 60. Bates RL, Jackson JA. Glossary of Geology. 2nd ed Alexandria, Virginia: American Geological Institute; 1995. [Google Scholar]
- 61. McCune B, Keon D. Equations for potential annual direct incident radiation and heat load. J Vegetat Sci. 2002; 13(4): 603–606. [Google Scholar]
- 62. Pierce KB Jr, Lookingbill T, Urban D. A simple method for estimating potential relative radiation (PRR) for landscape-scale vegetation analysis. Landsc Ecol. 2005; 20:137–147. [Google Scholar]
- 63. Holden ZA, Jolly WM. Modeling topographic influences on fuel moisture and fire danger in complex terrain to improve wildland fire management decision support. For Ecol Managem. 2011; 262: 2133–2141. [Google Scholar]
- 64. Deng Y, Goodchild MF, Chen X. Using NDVI to define thermal south in several mountainous landscapes of California. Comp Geosc. 2009. 35: 327–336. [Google Scholar]
- 65. Hansen MC, Potapov PV, Moore R, Hancer M, Turubanova SA, Tyukvina A, et al. High-resolution global maps of 21st-century forest cover change. Science. 2013; 10.1126/science.1244693 [DOI] [PubMed] [Google Scholar]
- 66. Soller DR, Reheis MC, Garrity CP, Van Sistine DR. Map database for surficial materials in the conterminous United States. US Geological Survey Data Series 425. 2009. [Google Scholar]
- 67. Theobald DM. A general model to quantify ecological integrity for landscape assessments and US application. Landsc Ecol: 2013; 10.1007/s10980-013-9941-6 [DOI] [Google Scholar]
- 68. Jenkins CN, Pimm SL, Joppa LN. Global patterns of terrestrial vertebrate diversity and conservation. PNAS. 2013; 10.1073/pnas.1302251110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.US Geological Survey, Gap Analysis Program (USGS-GAP) (2010) National GAP Land Cover, Version 1. Available: http://gapanalysis.usgs.gov. Accessed 29 July 2011.
- 70. Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A. Very high resolution interpolated climate surfaces for global land areas. Intern J Climatol. 2005; 25: 1965–1978. [Google Scholar]
- 71.R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing. 2014. http://www.R-project.org
- 72. Brierley G, Fryirs K, Jain V. Landscape connectivity: the geographic basis of geomorphic applications. Area. 2006; 38(2): 165–174. [Google Scholar]
- 73. Dobroski SZ, Abatzoglou J, Swanson AK, Greenberg JA, Mynsberge AR, et al. The climate velocity of the contiguous United States during the 20th century. Glob Chnge Biol. 2013; 19(1): 241–251. [DOI] [PubMed] [Google Scholar]
- 74. Cowling R, Pressey R. Rapid plant diversification: planning for an evolutionary future. PNAS. 2001; 98: 5452–5457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Rouget M, Cowling RM, Lombard AT, Knight AT, Kerley GH. Designing large-scale conservation corridors for pattern and process. Conserv Biol. 2006; 20: 549–561. [DOI] [PubMed] [Google Scholar]
- 76. Davis EB, Koo MS, Conroy C, Patton JL, Moritz C. The California Hotspots Project: identifying regions of rapid diversification of mammals. Molecul Ecol. 2008; 17: 120–38. [DOI] [PubMed] [Google Scholar]
- 77. Seneviratne SI, Corti T, Davin EL, Hirschi M, Jaeger EB, et al. Investigating soil moisture-climate interactions in a changing climate: a review. Earth-Science Rev. 2010; 99: 125–161. [Google Scholar]
- 78. Flint LE, Flint AL. Downscaling future climate scenarios to fine scales for hydrologic and ecological modeling and analysis. Ecol Process. 2011; 1:1. [Google Scholar]
- 79. Potter KA, Arthur-Woods H, Pincebourde S. Microclimatic challenges in global change biology. Glob Chang Biol. 2013; 19: 2932–2939. 10.1111/gcb.12257 [DOI] [PubMed] [Google Scholar]
- 80. Mawdsley JR, O’Malley R, Ojima DS. A review of climate-change adaptation strategies for wildlife management and biodiversity conservation. Conserv Biol. 2009; 23(5): 1080–1089. 10.1111/j.1523-1739.2009.01264.x [DOI] [PubMed] [Google Scholar]
- 81. Dawson TP, Jackson ST, House JI, Prentice IC, Mace GM. Beyond predictions: biodiversity conservation in a changing climate. Science. 2011; 332(6025): 53–58. 10.1126/science.1200303 [DOI] [PubMed] [Google Scholar]
- 82. US Forest Service (USFS). Forest Service global change research strategy, 2009–2019 implementation plan. FS-9. 2010. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Landforms were defined using basic topographic measures derived directly from the USGS 10 meter Digital Elevation Model, as well as latitude. Physiographic classes were generated by overlaying the landforms and lithology (parent material) converted from polygons (1:1,000,000 scale) to 270 m raster grid. The program used to generate these datasets is available from the authors upon request.
(PNG)
Data Availability Statement
Datasets are available at: https://www.sciencebase.gov/catalog/item/564b4bb0e4b0ebfbef0d31d2.