

Abstract
Background
In Staphylococcus aureus sub-populations of slow-growing cells forming small colony variants (SCVs) are associated with persistent and recurrent infections that are difficult to eradicate with antibiotic therapies. In SCVs that are resistant towards aminoglycosides, mutations have been identified in genes encoding components of the respiratory chain. Given the high frequencies of SCVs isolated clinically it is vital to understand the conditions that promote or select for SCVs.
Results
In this study we have examined how exposure to sub-inhibitory concentrations of antibiotics with different mechanism of action influence the formation of SCVs that are resistant to otherwise lethal concentrations of the aminoglycoside, gentamicin. We found that exposure of S. aureus to fluoroquinolones and mitomycin C increased the frequency of gentamicin resistant SCVs, while other antibiotic classes failed to do so. The higher proportion of SCVs in cultures exposed to fluoroquinolones and mitomycin C compared to un-exposed cultures correlate with an increased mutation rate monitored by rifampicin resistance and followed induction of the SOS DNA damage response.
Conclusion
Our observations suggest that environmental stimuli, including antimicrobials that reduce replication fidelity, increase the formation of SCVs through activation of the SOS response and thereby potentially promote persistent infections that are difficult to treat.
Keywords: Staphylococcus aureus, Small colony variants, Mutation, SOS response
Background
The formation of a small colony phenotype compared to wild type colony morphology has been observed in clinical specimens across many bacterial genera comprising Gram-positives and -negatives [1]. Clinically, the emergence of slow-growing Staphylococcus aureus mutants, known as small colony variants (SCVs), is associated with sub-acute infections displaying enhanced intracellular residency in host cells promoting persistent and recurrent infections and an increased risk of antibiotic treatment failure [1]. The SCV phenotype can be caused by disruptions of metabolic pathways, conferred either by mutations in the gene thyA of the pathway that converts uracil to thymidine [2] or by mutations in the menadione and hemin production pathways resulting in a dysfunctional electron transport chain [3, 4]. Depending on the pathways inactivated, antibiotic resistance to different classes of antimicrobial agents may occur [5]. Mutations in thyA confer trimethoprim-sulfamethoxazole resistance [2, 6], while mutations in genes of the biosynthesis pathways encoding menadione or hemin confer resistance to aminoglycosides [3, 4] and certain antimicrobial peptides [7, 8]. The resistance mechanism to aminoglycosides in SCVs defective in the electron transport chain has been linked to a decrease in membrane potential compared to wild type cells, causing a reduction in aminoglycosides uptake [1, 9]. SCVs with a defective electron transport chain exhibit distinct phenotypic characteristics affecting pathogenesis, i.e. reduced pigmentation, decreased hemolysis activity and most often display auxotrophy for menadione and hemin [1]. In a wild type S. aureus population a sub-population exists as SCVs [10], however environmental stimuli may increase the frequency of SCV formation. Increased frequency of S. aureus gentamicin resistant SCVs has been detected during co-cultures of S. aureus and Pseudomonas aeruginosa [11], in the intracellular milieu of endothelial cells [12] or in strains with reduced replication fidelity [13]. Short-term enhanced mutation rates of bacterial cells can occur through activation of the SOS response [14] and therefore activation of the SOS response could potentially increase the frequency of SCVs. The SOS regulon is generally repressed by LexA, but upon DNA damage it undergoes autocleavage in response to RecA binding to single-stranded DNA [15]. Inactivation of the LexA repressor leads to the expression of the SOS regulon. This regulon includes genes involved in DNA repair, recombination, as well as error-prone polymerases [16]. The error-prone polymerase V, encoded by the gene umuC, is highly up-regulated upon SOS response activation in S. aureus [17, 18]. In addition to traditional DNA damaging agents like UV and mitomycin C, other agents also induce the SOS response such as antibiotics of the flouroquinolone class, i.e. ciprofloxacin and moxifloxacin [17, 18]. Therefore we have investigated if antimicrobial compounds that interfere with DNA-, RNA-, protein- and cell wall synthesis affect the proportion of SCVs in the population.
Results and discussion
To address the effect of the SOS response and exposure to antimicrobials on the formation of gentamicin resistant SCVs in S. aureus we employed two strains, the wild type strain 8325-4 as well as an isogenic mutant expressing a non-cleavable LexA variant (lexA S130A) that is unable to induce the SOS response [18]. When measuring the minimum inhibitory concentration (MIC) of the strains, increased susceptibility was evident for the lexA(S130A) mutant towards chloramphenicol, as well as the DNA targeting antimicrobials; ciprofloxacin, moxifloxacin and mitomycin C, while equal MIC between the two strains was observed for gentamicin, rifampicin and vancomycin (Table 1).
Table 1.
MICs (μg/ml) of the antimicrobials used in this study for 8325-4 and its lexA derivative
| Antibiotic | 8325-4 | lexA(S130A) |
|---|---|---|
| Gentamicin | 1 | 1 |
| Ciprofloxacin | 1/2 | 1/4 |
| Moxifloxacin | 1/16 | 1/32 |
| Vancomycin | 1 | 1 |
| Chloramphenicol | 4 | 2 |
| Rifampicin | 1/512 | 1/512 |
| Mitomycin C | 1/8 | 1/32 |
Subsequently we measured the effect of sub-inhibitory concentrations (0.5 MIC) of antibiotics on the formation of gentamicin resistant SCVs using the wild type S. aureus strain 8235-4 and the isogenic lexA(S130A) mutant. The frequency of gentamicin resistant SCVs in an un-exposed culture of 8325-4 was on average 5.1 × 10−7, whereas 24 h exposure to mitomycin C, ciprofloxacin and moxifloxacin increased the frequencies of SCVs 11-, 15- and 8-fold, respectively, compared to the SCV frequency of the un-exposed 8325-4 (Fig. 1). The remaining three antimicrobials assayed, chloramphenicol, vancomycin and rifampicin, did not affect SCV formation. For the mutant strain expressing the non-cleavable lexA(S130A) exposure to none of the antimicrobials influenced SCV formation (Fig. 1). These results suggest that antimicrobials targeting DNA enhance SCV formation in an SOS dependent manner.
Fig. 1.
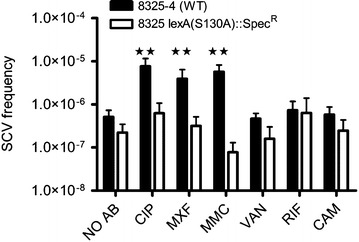
Effect of sub-inhibitory concentrations of antimicrobials on SCV frequency. The frequency of gentamicin resistant SCVs in WT 8325-4 (black) population and in the isogenic mutant lexA(S130A) (white) grown in absence of antibiotics (NO AB) or in the presence of sub-inhibitory concentrations of CIP (Ciprofloxacin), MXF (Moxifloxacin), MMC (Mitomycin C), VAN (Vancomycin), RIF (Rifampicin) and CAM (Chloramphenicol). Concentrations of antimicrobials are 0.5 MIC of the respective strains. The bars represent the mean from 8 independent selections of each condition and error bars represent 95 % confidence intervals
The increased frequency of SCVs observed in response to compounds causing DNA damage led us to investigate if the SOS regulon was activated. Activation of the SOS regulon was monitored with a recA::lacZ fusion in 8325-4. Expression of the recA gene was induced when exposed to mitomycin C, ciprofloxacin and moxifloxacin, while none of the remaining three agents increased the expression of recA (Fig. 2). The SOS regulon controls the expression of the error-prone polymerase Pol V, which confers increased mutation frequency in S. aureus [18]. Reduced replication fidelity could account for the increased number of SCV observed for the agents activating the SOS regulon. Mutation frequency has previously been monitored by rifampicin resistance, which arises by point mutations in rpoB [19]. The frequency of rifampicin resistant mutants in un-exposed cultures of the two strains was similar and approximately 3 × 10−8. In contrast exposure of 8325-4 to mitomycin C, ciprofloxacin and moxifloxacin increased the frequency of rifampicin resistant mutants 10-, 8- and sixfold respectively, whereas none of the remaining antibiotics for 8325-4 or any of the antibiotics for the lexA(S130A) mutant strain affected the frequency of rifampicin resistance (Fig. 3). Several of the antibiotics (ciprofloxacin, chloramphenicol and rifampicin) at the tested concentration also significantly reduced growth rates in exponential phase, yet no correlation between growth rate and increased frequency of SCVs was observed (Fig. 4).
Fig. 2.
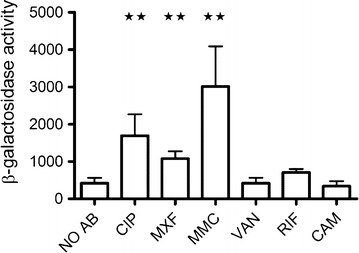
Sub-inhibitory concentrations of DNA-damaging agents activate the SOS-response. Activation of the SOS-response was monitored as β-galactosidase activity from a recA::lacZ fusion in strain 8325-4, when grown in absence (NO AB) or in the presence of sub-inhibitory concentrations of CIP (Ciprofloxacin), MXF (Moxifloxacin), MMC (Mitomycin C), VAN (Vancomycin), RIF (Rifampicin) and CAM (Chloramphenicol). The bars represent the mean from 3 independent experiments of each condition and error bars represent 95 % confidence intervals
Fig. 3.
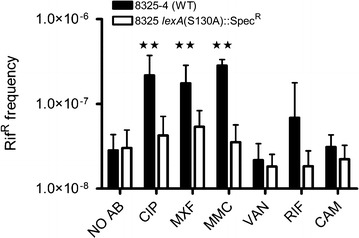
Effect of sub-inhibitory concentrations of antimicrobials on the frequency of rifampicin resistant mutants. The frequency of rifampicin resistant mutants in WT 8325-4 (black) population and in the isogenic mutant lexA(S130A) (white) grown in absence of antibiotics (NO AB) or in the presence of sub-inhibitory concentrations of CIP (Ciprofloxacin), MXF (Moxifloxacin), MMC (Mitomycin C), VAN (Vancomycin), RIF (Rifampicin) and CAM (Chloramphenicol). Concentrations of antimicrobials are 0.5 MIC of the respective strains. The bars represent the mean from 8 independent selections of each condition and error bars represent 95 % confidence intervals
Fig. 4.
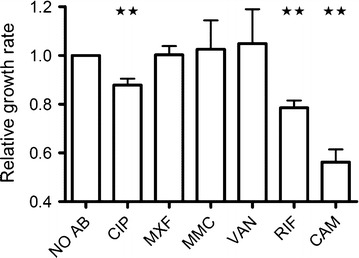
Sub-inhibitory concentrations of antimicrobials on the growth rate of strain 8325-4. The growth rate of strain 8325-4 grown in absence (NO AB) is set to 1 and the relative growth rates are presented of 8325-4 when grown in the presence of sub-inhibitory concentrations of CIP (Ciprofloxacin), MXF (Moxifloxacin), MMC (Mitomycin C), VAN (Vancomycin), RIF (Rifampicin) and CAM (Chloramphenicol). The bars represent the mean from 3 independent experiments of each condition and error bars represent 95 % confidence intervals
Strains displaying higher basal mutation rates have been detected in several clinical SCV isolates with auxotrophies for hemin [13] and thymidine [20]. Identification of hypermutators among clinical SCV isolates suggests that high mutations rates may contribute to the emergence and establishment of SCV infections [13, 20]. Not only exposure to sub-inhibitory concentrations of antibiotics that trigger activation of the SOS response and a temporary increase in mutation rate may facilitate the emergence of SCVs, but also conditions following contact with the immune system may activate the SOS response [21]. As part of the host defense mechanism against infections, neutrophils generate reactive oxygen species, such as hydrogen peroxide and superoxide [22] that can damage DNA and trigger expression of the SOS regulon [21]. It has recently been shown that exposure to sub-inhibitory concentrations of hydrogen peroxide indeed increases the frequency of gentamicin resistant SCVs via activation of the SOS response [23]. Therefore chemical inhibition that prevents activation of the SOS response could potentially be used as adjuvants to reduce the emergence and establishment of SCVs during infection, a strategy that has also been proposed to decelerate the evolution of antibiotic resistance towards various antimicrobial classes [24, 25].
Conclusion
In conclusion, our data suggests that environmental stimuli that elevate the mutation rate of S. aureus, i.e. through activation of the SOS response, stimulate the emergence of SCVs that are associated with persistent S. aureus infections. Caution should be taken when treating S. aureus empirically with broad-spectrum antibiotics such as fluoroquinolones when lacking resistance data, since sub-inhibitory exposure could increase the proportion of SCV and thereby limit further treatment options.
Methods
Bacterial strains, growth conditions and MIC
The strains used in this study include the S. aureus strains 8325-4, 8325 lexA(S130A)::SpecR [18] and HI2682, a 8325-4 strain containing a recA::lacZ fusion [26]. Strains were grown at 37 °C in tryptic soy broth (TSB) or on tryptic soy agar (TSA) plates with or without gentamicin (Sigma), ciprofloxacin (Sigma), moxifloxacin (Sigma), chloramphenicol (Sigma), vancomycin (Sigma), rifampicin (Sigma) and mitomycin C (Sigma). The minimum inhibitory concentrations (MIC) were determined by broth micro-dilution assay according to CLSI guidelines, except that cation-adjusted Mueller–Hinton broth was substituted with TSB.
Quantification of small colony variants and rifampicin resistant mutants
Over night (ON) cultures of 8325-4 and 8325 lexA(S130A)::SpecR were diluted 1000-fold in 1 ml TSB in a 10 ml falcon tube, supplemented with or without 0.5 MIC of the antimicrobials assayed. The cultures were grown for 24 h at 37 °C with shaking (200 rpm). Total colony forming units (CFU) were determined on TSA plates, the number of gentamicin resistant small colony variants were estimated on TSA supplemented with gentamicin (4 μg/ml) and rifampicin resistant mutant CFU were determined on TSA supplemented with rifampicin (5 μg/ml). Plates were incubated at 37 °C. Total CFU and rifampicin resistant mutants were counted following 24 h growth and SCVs were counted after 48 h growth. SCVs were defined as colonies resistant to gentamicin (>4 μg/ml) that displayed slow growth and significantly smaller colony morphology (approximately tenfold) compared to WT strain 8325-4. The frequency of SCV was determined as the number of SCV divided by the total CFU count. The same procedure was used in the calculation of rifampicin mutation frequency. The frequencies reported are a mean of 8 independent experiments.
β-galactosidase assay with recA::lacZ fusion
β-galactosidase activity was measured from strain HI2682, a 8325-4 strain with recA::lacZ fusion. Cells were grown in 10 ml TSB in 100 ml Erlenmeyer flasks at 37 °C with shaking with and without the presence of sub-inhibitory concentrations (0.5 MIC) of antimicrobials to an OD600 of 0.5. The assay was performed as described by Sambrook and Russell [27]. The reported β-galactosidase activities are a mean of three independent experiments.
Fitness measurement
The fitness of cells exposed to sub-inhibitory concentrations of antimicrobials was estimated by growth rate (µ) comparisons in exponential phase. Growth rates were determined for cells inoculated at 106 cells/ml and grown with or without antibiotics at a concentration of 0.5 MIC at 37 °C in a Bioscreen C (Oy Growth Curves Ab Ltd) with OD600 measurements at 5 min intervals. The relative fitness was calculated as: µ(mutant)/µ(WT).
Statistics
Significant difference was calculated by 1-way ANOVA, using log-transformed datasets for SCV and rifampicin resistance frequency, with a post hoc analysis of Dunnett Multiple Comparison Tests (*p < 0.05, **p < 0.01). Statistics were performed in GraphPad Prism 4 (GraphPad Software, Inc).
Authors’ contributions
MV conceived of the study, participated in its design, performed the experiments and participated in analysis of the data. WP and HI participated in the study design and analysis of the data. All authors participated in drafting the manuscript. All authors read and approved the final manuscript.
Acknowledgements
The Danish Research Council for Independent Research, Technology and Production nr. 12-127417.
Competing interests
The authors declare that they have no competing interests.
Contributor Information
Martin Vestergaard, Email: mave@sund.ku.dk.
Wilhelm Paulander, Email: wilhelm.paulander@gmail.com.
Hanne Ingmer, Email: hi@sund.ku.dk.
References
- 1.Proctor RA, von Eiff C, Kahl BC, Becker K, McNamara P, Herrmann M, Peters G. Small colony variants: a pathogenic form of bacteria that facilitates persistent and recurrent infections. Nat Rev Microbiol. 2006;4:295–305. doi: 10.1038/nrmicro1384. [DOI] [PubMed] [Google Scholar]
- 2.Chatterjee I, Kriegeskorte A, Fischer A, Deiwick S, Theimann N, Procter RA, Peters G, Herrmann M, Kahl BC. In vivo mutations of thymidylate synthase (Encoded by thyA) are responsible for thymidine dependency in clinical small-colony variants of Staphylococcus aureus. J Bacteriol. 2008;190:834–842. doi: 10.1128/JB.00912-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lannergård J, von Eiff C, Sander G, Cordes T, Seggewiβ J, Peters G, Procter RA, Becker K, Hughes D. Identification of the genetic basis for clinical menadione-auxotrophic small-colony variant isolates of Staphylococcus aureus. Antimicrob Agents Chemother. 2008;52:4017–4022. doi: 10.1128/AAC.00668-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Dean MA, Olsen RJ, Long SW, Rosato AE, Musser JM. Identification of point mutations in clinical Staphylococcus aureus strains that produce small-colony variants auxotrophic for menadione. Infect Immun. 2014;82:1600–1605. doi: 10.1128/IAI.01487-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kahl BC. Small colony variants (SCVs) of Staphylococcus aureus—a bacterial survival strategy. Infect Genet Evol. 2014;21:515–522. doi: 10.1016/j.meegid.2013.05.016. [DOI] [PubMed] [Google Scholar]
- 6.Besier S, Ludwig A, Ohlsen K, Brade V, Wichelhaus TA. Molecular analysis of the thymidine-auxotrophic small colony variant phenotype of Staphylococcus aureus. Int J Med Microbiol. 2007;297:217–225. doi: 10.1016/j.ijmm.2007.02.003. [DOI] [PubMed] [Google Scholar]
- 7.Samuelsen Ø, Haukland HH, Kahl BC, von Eiff C, Proctor RA, Ulvatne H, Sandvik K, Vorland LH. Staphylococcus aureus small colony variants are resistant to the antimicrobial peptide lactoferricin B. J Antimicrob Chemother. 2005;56:1126–1129. doi: 10.1093/jac/dki385. [DOI] [PubMed] [Google Scholar]
- 8.Pränting M, Andersson DI. Mechanisms and physiological effects of protamine resistance in Salmonella enterica serovar Typhimurium LT2. J Antimicrob Chemother. 2010;65:876–887. doi: 10.1093/jac/dkq059. [DOI] [PubMed] [Google Scholar]
- 9.Taber HW, Mueller JP, Miller PF, Arrow AS. Bacterial uptake of aminoglycoside antibiotics. Microbiol Rev. 1987;51:439–457. doi: 10.1128/mr.51.4.439-457.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Edwards AM. Phenotype switching Is a natural consequence of Staphylococcus aureus replication. J Bacteriol. 2012;194:5404–5412. doi: 10.1128/JB.00948-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hoffman LR, Déziel E, D’Argenio DA, Lépine F, Emerson J, McNamara S, Gibson RL, Ramsey BW, Miller SI. Selection for Staphylococcus aureus small-colony variants due to growth in the presence of Pseudomonas aeruginosa. PNAS. 2006;103:19890–19895. doi: 10.1073/pnas.0606756104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Vesga O, Groeschel MC, Otten MF, Brar DW, Vann JM, Proctor RA. Staphylococcus aureus small colony variants are induced by endothelial cell intracellular milieu. J Infect Dis. 1996;173:739–742. doi: 10.1093/infdis/173.3.739. [DOI] [PubMed] [Google Scholar]
- 13.Schaaff F, Bierbaum G, Baumert N, Bartmann P, Sahl HG. Mutations are involved in emergence of aminoglycoside-induced small colony variants of Staphylococcus aureus. Int J Med Microbiol. 2003;293:427–435. doi: 10.1078/1438-4221-00282. [DOI] [PubMed] [Google Scholar]
- 14.Sutton MD, Smith BT, Godoy VG, Walker GC. The SOS response: recent insights into umuDC-dependent mutagenesis and DNA damage tolerance. Ann Rev Genet. 2000;34:479–497. doi: 10.1146/annurev.genet.34.1.479. [DOI] [PubMed] [Google Scholar]
- 15.Butala M, Žgur-Bertok D, Busby SJW. The bacterial LexA transcriptional repressor. Cell Mol Life Sci. 2009;66:82–93. doi: 10.1007/s00018-008-8378-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Erill I, Campoy S, Barbé J. Aeons of distress: an evolutionary perspective on the bacterial SOS response. FEMS Microbiol Rev. 2007;31:637–656. doi: 10.1111/j.1574-6976.2007.00082.x. [DOI] [PubMed] [Google Scholar]
- 17.Mesak LR, Miao V, Davies J. Effects of subinhibitory concentrations of antibiotics on SOS and DNA repair gene expression in Staphylococcus aureus. Antimicrob Agents Chemother. 2008;52:3394–3397. doi: 10.1128/AAC.01599-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Cirz RT, Jones MB, Gingles NA, Minogue TD, Jarrahi B, Peterson SN, Romesberg FE. Complete and SOS-mediated response of Staphylococcus aureus to the antibiotic ciprofloxacin. J Bacteriol. 2007;189:531–539. doi: 10.1128/JB.01464-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Taddei F, Matic I, Radman M. cAMP-dependent SOS induction and mutagenesis in resting bacterial populations. PNAS. 1995;92:11736–11740. doi: 10.1073/pnas.92.25.11736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Besier S, Zander J, Kahl BC, Kraiczy P, Brade V, Wichelhaus TA. The thymidine-dependent small-colony-variant phenotype is associated with hypermutability and antibiotic resistance in clinical Staphylococcus aureus. Antimicrob Agents Chemother. 2008;52:2183–2189. doi: 10.1128/AAC.01395-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Imlay JA, Linn S. Mutagenesis and stress responses induced in Escherichia coli by hydrogen peroxide. J Bacteriol. 1987;169:2967–2976. doi: 10.1128/jb.169.7.2967-2976.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.DeLeo FR, Diep BA, Otto M. Host defense and pathogenesis in Staphylococcus aureus infections. Infect Dis Clin North Am. 2009;23:17–34. doi: 10.1016/j.idc.2008.10.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Painter KL, Strange E, Parkhill J, Bamford KB, Armstrong-James D, Edwards AM. Staphylococcus aureus adapts to oxidative stress by producing H2O2-resistant small colony variants via the SOS response. Infect Immun. 2015;83:1830–1844. doi: 10.1128/IAI.03016-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Cirz RT, Chin JK, Andes DR, de Crécy-Lagard V, Craig WA, Romesberg FE. Inhibition of mutation and combating the evolution of antibiotic resistance. PLoS Biol. 2005;3:e176. doi: 10.1371/journal.pbio.0030176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wigle TJ, Sexton JZ, Gromova AV, Hadimani MB, Hughes MA, Smith GR, Yeh Li-An, Singleton SF. Inhibitors of RecA activity discovered by high-throughput screening: cell-permeable small molecules attenuate the SOS response in Escherichia coli. J Biomol Screen. 2009;14:1092–1101. doi: 10.1177/1087057109342126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gottschalk S, Ifrah D, Lerche S, Gottlieb CT, Cohn MT, Hiasa H, Hansen PR, Gram L, Ingmer H, Thomsen LE. The antimicrobial lysine-peptoid hybrid LP5 inhibits DNA replication and induces the SOS response in Staphylococcus aureus. BMC Microbiol. 2013;13:192. doi: 10.1186/1471-2180-13-192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Sambrook J, Russel DWI. Molecular cloning: a laboratory manual. 3. Cold Spring Harbor: Cold Spring Harbor Laboratory Press; 2001. [Google Scholar]