

Abstract
Background
The root of Panax quinquefolium L., famous as American ginseng all over the world, is one of the most widely-used medicinal or edible materials. Ginsenosides are recognized as the main bioactive chemical components responsible for various functions of American ginseng. In this study, tissue-specific chemicals of P. quinquefolium were analyzed by laser microdissection and ultra-high performance liquid chromatography- quadrupole/time-of-flight-mass spectrometry (UHPLC-Q/TOF–MS) to elucidate the distribution pattern of ginsenosides in tissues. The contents of ginsenosides in various tissues were also compared.
Results
A total of 34 peaks were identified or temporarily identified in the chromatograms of tissue extractions. The cork, primary xylem or cortex contained higher contents of ginsenosides than phloem, secondary xylem and cambium. Thus, it would be reasonable to deduce that the ratio of total areas of cork, primary xylem and the cortex to the area of the whole transection could help to judge the quality of American ginseng by microscopic characteristics.
Conclusion
This study sheds new light on the role of microscopic research in quality evaluation, and provides useful information for probing the biochemical pathways of ginsenosides.
Graphical abstract.
.
Keywords: Ginsenosides, Panax quinquefolium L., Tissue-specific, Laser microdissection, UHPLC-Q/TOF–MS
Background
Microscopic authentication refers to examine the structure, cell and internal features of herbal medicines using a microscope and its derivatives. It has been recorded in many Pharmacopoeias as an authentication method, such as Chinese Pharmacopoeia, United States Pharmacopeia, European Pharmacopoeia, British Pharmacopoeia, Japanese Pharmacopoeia, and Korean Pharmacopoeia. Distinctly, microscopic authentication has been commonly used in the authentication of herbal medicines. As we know, the secondary metabolites of herbal medicine contribute to its effects. Nevertheless, the normal microscopic identification cannot provide the useful information of secondary metabolites in different herbal materials directly. Thus, microscopic method can identify the source species but not evaluate the quality of herbal medicines.
By using techniques of anatomy and histochemistry, some studies have demonstrated that there is a close relationship between microscopic characteristics and active components of herbal medicines. For example, the histochemical techniques and phytochemical methods have been applied in the distribution and accumulation of active components in Sinomenium acutum, Aloe vera var. chinensis, Gynostemma pentaphyllum, Dioscorea zingiberensis and Macrocarpium officinacle [1–5]. However, these studies used routine chemical reactions and thus the distribution of the detailed active components could not be identified. Moreover, those agents usually have poor specificity, which leads to the increase of false positive results. Also, it is noteworthy that these investigations lacked objective data and had not been validated by other methods yet. Recently, the combination of fluorescence microscopy, laser microdissection (LMD), and ultra-high performance liquid chromatography-quadrupole/time-of-flight-mass spectrometry (UHPLC-Q/TOF–MS) has been successfully applied to explore the distribution pattern of secondary metabolites among different tissues from several Chinese medicinal materials (CMMs) [6–11]. This method can obtain the exact quantitative and qualitative data to profile the chemicals in tissues and cells of medicinal materials.
American ginseng, the root of Panax quinquefolium L., is one of the most recognized herbal medicines all over the world. Also, American ginseng has become popular in oriental countries as dietary health supplements or additives to foods and beverages [12]. In the herbal markets, various specifications or grades of American ginseng can be found, including main root, rootlet and fibrous root. Production area also affects the grade or price of the commercial medicine. As we know, American ginseng contains the major bioactive triterpene saponins named ginsenosides, such as ginsenosides Rg1, 20(S)-Rg2, Re, 20(S)-Rh1, Rb1, Rb2 and Rd, which possess a wide range of pharmacological effects, including cardiovascular, anti-diabetic, anti-inflammatory and anti-tumor properties [13–16].
To evaluate the quality of American ginseng, a number of analytical methods to determine the total ginsenoside content or the target compounds have been developed [17–19]. However, few of them focus on the distribution rules of ginsenosides among tissues or detect the relationship of the quality and the microscopic characteristics. Until now, ginsenosides in the rhizome and root of P. ginseng Meyer has already been located: the cork contained more kinds of ginsenosides than did the cortex, phloem, xylem and resin canals [8]. But whether this rule applies to P. quinquefolium or not still waits to be found out. Analyzing the distribution of ginsenosides in different anatomical structures will establish the relationship between microscopic features and active components. Then the microscopic features used for the quality evaluation and classification of different specifications or grades of American ginseng can be validated or clarified.
In this study, fluorescence microscopy, LMD and UHPLC-Q/TOF–MS were used to analyze and compare the spatial chemical profiles of various tissues from P. quinquefolium to correlate the relationship between microscopic features and active components for the quality evaluation of American ginseng, shedding new light on the role of microscopic research in quality evaluation.
Results and discussion
Microscopic examination and dissection by LMD
In this study, four fresh P. quinquefolium samples (Pq1–4) and nine dried commercial samples were collected for analysis (see Table 1; Fig. 1). As shown under the normal light and fluorescence mode (see Fig. 2), the transverse section of American ginseng was comprised of cork, cortex, phloem, cambium and xylem. The cork was consisted of several rows of densely-arranged flat cells. Red fluorescence was emitted from the cork while blue color was shown in other tissues. Cortex was narrow. Cracks could be seen in phloem. Resin ducts with orange red fluorescence were scattered in the cortex and phloem. Cambium was arranged in a ring, showing strong florescence. Xylem was broad, usually differentiated into primary xylem with strong florescence and secondary xylem with common florescence. Since our study on localization of ginsenosides in the rhizome and root of P. ginseng illustrated that the resin ducts contained few ginsenosides, the resin ducts of P. quinquefolium samples were not examined here. The cork, cortex, phloem, secondary xylem and primary xylem were dissected from the main roots of Pq1–4 and Pq5–13. For the branch roots of Pq1–4, the xylem was hardly seen differentiation, and was thus examined as a whole. Compared with other samples, the cambium in the cross sections of Pq6 and Pq8 was obvious with relative more layers of cells, hence, the cambium of Pq6 and Pq8 were also investigated. Therefore, various tissues possessed different features and could be recognized under fluorescence mode. According to previous reports [6–8], the size of about 2,500,000 and 1,000,000 μm2 of each separated tissues of fresh and dried materials were dissected by LMD respectively which could detect the chemicals containing in tissues.
Table 1.
Information of commercial samples of Panax quinquefolium materials
| Sample no. | Commercial name | Specification | Harvest time | Harvest place |
|---|---|---|---|---|
| Pq1 | American ginseng | – | September 12th, 2014 | Cultivation in Mulin County, Mudanjiang City, Heilongjiang Province, China |
| Pq2 | American ginseng | – | September 12th, 2014 | Cultivation in Mulin County, Mudanjiang City, Heilongjiang Province, China |
| Pq3 | American ginseng | – | September 12th, 2014 | Cultivation in Mulin County, Mudanjiang City, Heilongjiang Province, China |
| Pq4 | American ginseng | – | September 12th, 2014 | Cultivation in Mulin County, Mudanjiang City, Heilongjiang Province, China |
| Pq5 | Wild-mountain pao-shen no. 1 | HK$ 66,137.57/1000 g | – | Wildlife in America |
| Pq6 | Wild-mountain small pao-shen no. 3.5 | HK$ 34,391.53/1000 g | – | Wildlife in America |
| Pq7 | Wild-mountain small and rouond pao-shen | HK$ 25,873.02/1000 g | – | Wildlife in America |
| Pq8 | Wild-mountain pao-mian no. 3.5 | HK$ 76,190.48/1000 g | – | Wildlife in America |
| Pq9 | Wild-mountain pao-mian no. 4 | HK$ 52,645.5/1000 g | – | Wildlife in America |
| Pq10 | Wild-mountain small and rouond pao-mian | HK$ 44,973.54/1000 g | – | Wildlife in America |
| Pq11 | Cultivated big-branch Pao-shen | HK$ 1534.39/1000 g | – | Cultivation in Canada |
| Pq12 | Cultivated middle-branch Pao-shen | HK$ 1428.57/1000 g | – | Cultivation in Canada |
| Pq13 | Cultivated shen no. 4 | HK$ 1111.11/1000 g | – | Cultivation in Canada |
Fig. 1.
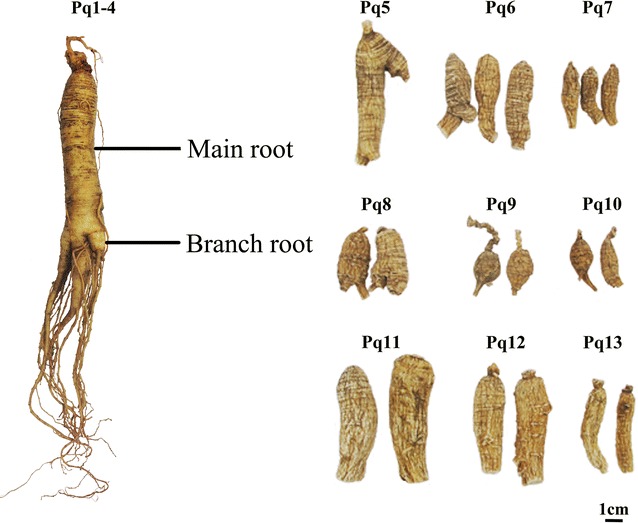
Morphological features of Panax quinquefolium materials
Fig. 2.
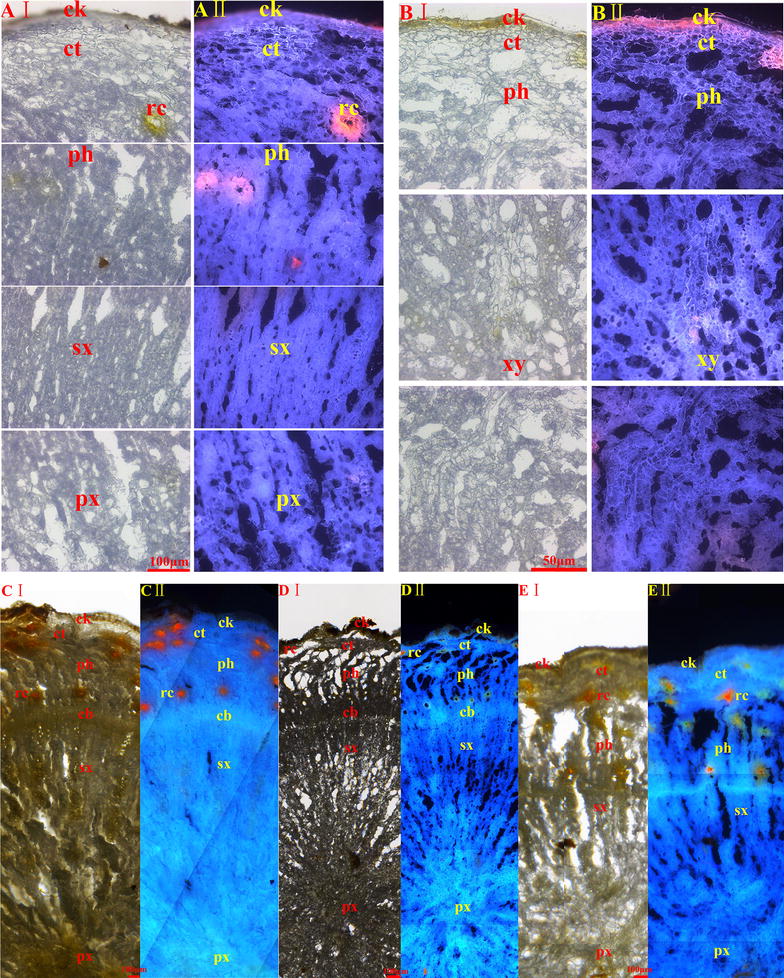
Microscopic characteristics of P. quinquefolium. I Under normal light microscope, II under fluorescence mode with dichromatic mirror. a, b represented the main root and branch root of Pq1; c–e represented Pq6, Pq8 and Pq10 respectively. ck cork, ct cortex, ph phloem, rc resin canals, cb cambium, xy xylem, sx secondary xylem, px primary xylem, pt pith
Tissue-specific chemical profiles
By UHPLC-Q/TOF–MS technique, tissue-specific chemical profiles of each sample were obtained as total ion chromatograms (see Figs. 3, 4). A total of 34 peaks were detected in all the tissue extractions. By comparing retention times, accurate mass weights, and mass ions with the reference compounds, six peaks (Peaks 3, 4, 14, 15, 23, 29) were unambiguously identified as ginsenosides Rg1, Re, 20(S)-Rg2, 20(S)-Rb1, Rb2 and Rd. By matching those data with the components reported in the literature, 25 compounds were tentatively authenticated [12, 20–24]. The identification result is shown in Table 2.
Fig. 3.
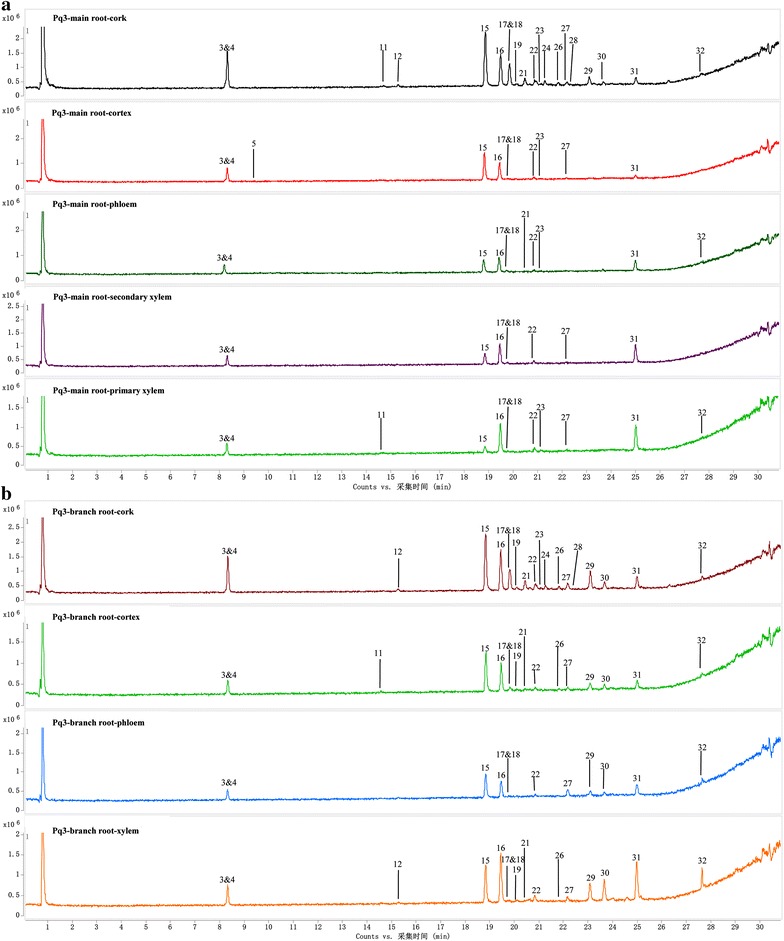
The total ions current (TIC) chromatograms of microdissected tissues from main root (a) and branch root (b) of P. quinquefolium samples. The peak numbers referred to Table 2
Fig. 4.
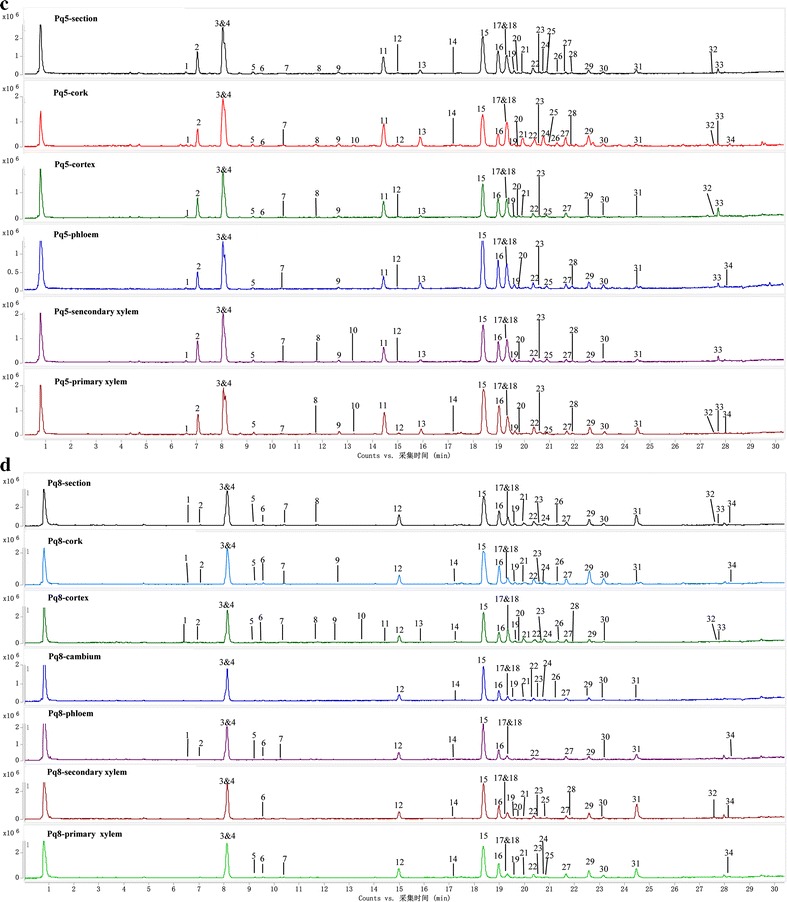
The total ions current (TIC) chromatograms of microdissected tissues from P. quinquefolium samples of Pq5 (c) and Pq8 (d). The peak numbers referred to Table 2
Table 2.
Compounds identified from tissue extractions of Panax quinquefolium samples
| Peak no. | Identity | tR (min) | Molecular formular | [M−H]+ | [M−H+HCOOH]+ (mass accuracy, ppm) | Fragments of [M−H]+ (m/z) | ||
|---|---|---|---|---|---|---|---|---|
| Mean measured mass (Da) | Theoretical exact mass (Da) | Mass accuracy (ppm) | ||||||
| 1 | 20-Glc-G-Rf | 6.58 | C48H82O19 | 961.5522 | 961.5378 | 14.98 | 1007.5578 | 799.5047 [M−H−Glc]− |
| 2 | Notoginsenoside R1 | 7.04 | C47H80O18 | 931.5186 | 931.5278 | −9.88 | 977.5465 | 799.5026 [M−H−Xyl]−
637.4285[M−H−Glc−Xyl]−; |
| 3 | G-Rga1 | 8.04 | C42H72O14 | 799.4975 | 799.4849 | 15.76 | 845.5026 | 637.4360 [M−H−Glc] −
475.3785 [M−H−2Glc]− |
| 4 | G-Rea | 8.12 | C48H82O18 | 945.5548 | 945.5428 | 12.69 | 991.5630 | 799.4935 [M−H−Rha]−
783.5029 [M−H−Glc]− 637.4407 [M−H−Rha−Glc] – |
| 5 | Malonyl-G-Rg1 | 9.25 | C45H74O17 | 885.5082 | 885.4853 | 25.86 | – | 841.3240 [M−H−CO2]− |
| 6 | Malonyl-G-Re isomer | 9.56 | C51H84O21 | 1031.5547 | 1031.5432 | 11.15 | – | 987.5678[M−H−CO2]− |
| 7 | Malonyl-G-Re | 10.32 | C51H84O21 | 1031.5549 | 1031.5432 | 11.34 | – | 987.5644[M−H−CO2]− |
| 8 | Floralquinquenoside B | 11.73 | C42H72O15 | 815.4884 | 815.4793 | 11.16 | – | 637.4381[M−H−Rha−CH3OH]− |
| 9 | Floralquinquenoside D | 12.65 | C42H72O15 | 815.4882 | 815.4793 | 10.91 | 861.5002 | 653.4360 [M−H−Glc]− |
| 10 | Unknown | 13.26 | – | 961.5559 | – | – | 1007.5580 | – |
| 11 | Notoginsenoside Rw2 | 14.43 | C41H70O14 | 785.4780 | 785.4687 | 11.84 | 831.4871 | 653.4361 [M−H−Xyl]−
491.3674 [M−H−Xyl−Glc]− |
| 12 | Pseudoginsenoside F11 | 14.99 | C42H72O14 | 799.4831 | 799.4844 | −1.63 | 845.5015 | 653.4385 [M−H−Rha]− |
| 13 | Notoginsenoside R2 | 15.89 | C41H70O13 | 769.4573 | 769.4738 | −21.44 | 815.4730 | 637.4392 [M−H−Xyl]−
475.3839 [M−H− Xyl−Glc]− |
| 14 | 20 (S)-G-Rga2 | 17.23 | C42H72O13 | 783.5029 | 783.4900 | 16.46 | 829.5054 | 637.4394 [M−H−Rha]−
475.3734 [M−H−Rha−Glc]− |
| 15 | G-Rba1 | 18.38 | C54H92O23 | 1107.6097 | 1107.5957 | 12.64 | – | 945.5552[M−H−Glc]−
783.5012 [M−H−2Glc] − |
| 16 | Malonyl-G-Rb1 | 18.99 | C57H94O26 | 1193.6113 | 1193.5961 | 12.73 | – | 1149.6201[M−H−CO2]− |
| 17 | G-Ro | 19.33 | C48H76O19 | 955.5077 | 955.4908 | 17.69 | – | 793.2586[M−H−Glc]− |
| 18 | G-Rc | 19.34 | C53H90O22 | 1077.5730 | 1077.5871 | −13.08 | – | 945.5660 [M−H−Araf]−
783.4980 [M−H−Araf −Glc]− |
| 19 | Malonyl-G-Rb1 isomer I | 19.63 | C57H94O26 | 1193.6142 | 1193.5961 | 15.16 | – | 1149.6185[M−H−CO2]− |
| 20 | Unknown | 19.80 | – | 1087.5461 | – | – | – | |
| 21 | Malonyl-G-Ra2 | 19.97 | C56H92O25 | 1163.5993 | 1163.5855 | 11.86 | – | 1119.6041[M−H−CO2]− |
| 22 | Malonyl-G-Rb1 isomer II | 20.38 | C57H94O26 | 1193.6101 | 1193.5961 | 11.73 | – | 1149.6192[M−H−CO2]− |
| 23 | G-Rba2 | 20.47 | C53H90O22 | 1077.5683 | 1077.5851 | −15.59 | 1123.6337 | 945.5674 [M−H−Arap]− |
| 24 | G-Rb3 | 20.79 | C53H90O22 | 1077.5977 | 1077.5851 | 11.69 | 1123.6637 | 945.5587 [M−H−Xyl]−
915.5474 [M−H−Glc]− |
| 25 | Unknown | 20.91 | – | 1119.6015 | – | – | – | 925.4844 |
| 26 | Ma- Rb2/Rb3 isomer | 21.34 | C56H92O25 | 1163.5992 | 1163.5849 | 12.29 | – | 1119.6007[M−H−CO2]− |
| 27 | O-acetyl-G-Rb1 | 21.68 | C56H94O24 | 1149.6198 | 1149.6062 | 11.83 | 1195.6270 | 1107.6067 [M−H−Acetyl]−
945.5466 [M−H−Acetyl−Glc]− |
| 28 | Zingibroside R1 | 21.92 | C42H65O14 | 793.4479 | 793.4374 | 13.23 | – | 631.3332[M−H−Glc]− |
| 29 | G-Rda | 22.59 | C48H82O18 | 945.5548 | 945.5428 | 12.69 | 991.5613 | 783.4985 [M−H−Glc]−
621.4432 [M−H−2Glc]− |
| 30 | Malonyl-G-Rd | 23.18 | C51H84O21 | 1031.5614 | 1031.5432 | 17.64 | – | 987.5682[M−H−CO2]− |
| 31 | G-Rd isomer | 24.49 | – | 945.5543 | 945.5428 | 12.16 | 991.5069 | 783.4985 [M−H−Glc]−
621.4432 [M−H−2Glc]− |
| 32 | 20 (S)-G-Rg3 | 27.55 | C42H72O13 | 783.4978 | 783.4900 | 9.96 | 829.5057 | 621.4375 [M−H−Glc]−
459.4088 [M−H−2Glc]− |
| 33 | Chikusetsusaponin IVa | 27.69 | C42H66O14 | 793.4367 | 793.4380 | −1.64 | – | – |
| 34 | 20 (R)-G-Rg3 | 28.14 | C42H72O13 | 783.4982 | 783.4900 | 10.47 | 829.5065 | 621.4375 [M−H−Glc]−
459.3964 [M−H−2Glc]− |
G ginsenoside, Glc β-d-glucopyranosyl, Rha α-l-rhamnopyranosyl, Xyl β-d-xylopyranosyl, Araf α-l-arabinofuranosyl, Arap α-l-arabinopyranosyl
aIdentified with chemical marker
As seen from Figs. 3, 4, the distribution differences of gensenosides in various tissues from American ginseng were not as distinct as Asian ginseng [8]. The cork extractions usually had the most peaks (20–34 peaks). The cortex and primary xylem took the second place, namely 11–31 peaks and 12–30 peaks respectively. The secondary xylem (9–28 peaks), phloem (11–27 peaks) and cambium (24 peaks for Pq6, 18 peaks for Pq8) possessed the least peaks. For example, the cork, cortex, phloem, secondary xylem and primary xylem of Pq1 showed 34, 29, 29, 28 and 30 peaks separately. The tissues above of Pq7 had 32, 19, 14, 19 and 21 peaks respectively. Thus, the cork, primary xylem and cortex possessed the most kinds of saponin compounds.
For most samples, the areas of Peaks 21–30 in the cork were larger than those in other tissues. Peaks 21–30 represented compounds with medium or low polarity, which might be concerned with the protection function of the cork. In the xylem, especially the primary xylem, the areas of Peaks 17–31 were larger than those in cortex, phloem and cambium, which might be relevant with the lignification, suberification and the channel function of xylem cells.
Quantification of ginsenosides in various tissues
Ginsenosides Rg1, Re, Rh1, 20(S)-Rg2, 20(S)-Rb1, Rb2 and Rd in various tissues of different samples were determined by UHPLC-Q/TOF–MS. The results are given in Table 3 and Fig. 5. For most samples (Pq1–5, Pq7–10), the cork contained the most ginsenosides compared with other tissues, with the content ranging from 1094.58 to 269944.16 ng/105 μm2. Sometimes, the primary xylem possessed the highest level of ginsenosides (Pq6, Pq11–13), or possessed the second highest level (main root of Pq1, Pq5, Pq7–10), whereas sometimes low ginsenoside level was found in the primary xylem (main root of Pq2–4). The amounts of ginsenosides fluctuated in the cortex. It seemed that if the contents of ginsenosides were low in primary xylem, the contents would be high in cortex (main root of Pq2–4); and if the contents of ginsenosides were high in primary xylem, the cortex would have a medium (main root of Pq1, Pq5, Pq7, Pq8, Pq10) or low (Pq6, Pq9, Pq11–13) level of ginsenosides. The phloem, secondary xylem and cambium usually had fewer ginsenosides than other tissues. For the branch roots of Pq1-4, the cork, xylem and cortex occupied higher contents of ginsenosides than phloem did. Thus, the distribution pattern of ginsenosides in American ginseng was quite distinct from Asian ginseng. Distinctly, the cork, primary xylem or cortex had more ginsenosides than phloem, secondary xylem and cambium in American ginseng. Based on all the above, it was reasonable to deduce that the ratio of total areas of cork, primary xylem and the cortex to the area of whole transection could help to evaluate the quality of American ginsengs.
Table 3.
Contents of ginsenosides in the tissues from Panax quinquefolium samples
| Sample no. | Tissue | Amount in unit area (ng/105μm2) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Rga1 | Re | Rh1 | Rg2 | Rb1 | Rb2 | Rd | Sum | ||
| Pq1 main root | Cork | 67.31 | 34.58 | 0.40 | 8.83 | 13,247.66 | 25.51 | 13.18 | 13,397.47 |
| Cortex | 18.77 | 9.50 | –b | 1.83 | 5576.43 | 1.02 | 1.05 | 5608.60 | |
| Phloem | 11.53 | 7.46 | 0.25 | 1.87 | 4734.50 | 1.28 | 1.53 | 4758.42 | |
| Secondary xylem | 9.38 | 10.74 | – | 2.11 | 3176.85 | 2.20 | 1.69 | 3202.97 | |
| Primary xylem | 31.12 | 12.11 | 0.30 | 2.07 | 8104.59 | 2.15 | 1.49 | 8153.83 | |
| Pq1 branch root | Cork | 74.68 | 50.67 | – | 1.16 | 16,897.70 | 31.86 | 7.00 | 17,063.07 |
| Cortex | 10.84 | 9.88 | – | 2.81 | 7608.60 | 1.63 | 3.24 | 7637.00 | |
| Phloem | 5.50 | 5.46 | 0.35 | 2.17 | 5073.24 | 0.56 | 2.72 | 5090.00 | |
| Xylem | 8.10 | 9.36 | – | 3.97 | 7239.49 | 1.68 | 12.29 | 7274.89 | |
| Pq2 main root | Cork | 130.53 | 69.38 | 0.27 | 3.45 | 16,012.69 | 42.06 | 27.09 | 16,285.47 |
| Cortex | 36.33 | 19.63 | – | 0.75 | 5244.41 | 1.16 | 0.95 | 5303.23 | |
| Phloem | 16.39 | 8.66 | – | 2.57 | 3840.21 | 1.17 | 0.72 | 3869.72 | |
| Secondary xylem | 23.34 | 28.46 | – | 7.57 | 2344.29 | 2.51 | 1.50 | 2407.67 | |
| Primary xylem | 27.65 | 29.74 | – | 4.72 | 2688.51 | 3.21 | 0.93 | 2754.76 | |
| Pq2 branch root | Cork | 62.44 | 46.66 | 0.30 | 13.93 | 17,558.77 | 52.02 | 29.52 | 17,763.64 |
| Cortex | 11.64 | 8.78 | 0.36 | 2.84 | 3371.72 | 1.44 | 2.04 | 3398.82 | |
| Phloem | 11.57 | 8.17 | 0.37 | 4.20 | 3159.24 | 1.82 | 3.55 | 3188.92 | |
| Xylem | 23.09 | 18.41 | 0.34 | 9.31 | 5805.28 | 1.48 | 17.17 | 5875.08 | |
| Pq3 main root | Cork | 41.66 | 18.92 | 0.39 | 4.15 | 269,855 | 16.80 | 7.24 | 269,944.16 |
| Cortex | 18.67 | 7.19 | 0.59 | 1.61 | 145,606.6 | 3.61 | 1.15 | 145,639.42 | |
| Phloem | 11.31 | 6.40 | 0.51 | 0.98 | 67,598.38 | – | 1.21 | 67,618.79 | |
| Secondary xylem | 11.69 | 6.42 | 0.31 | 0.84 | 50,655.09 | – | – | 50,674.35 | |
| Primary xylem | 10.03 | 5.35 | 0.33 | 0.87 | 19,113.26 | 0.46 | 0.30 | 19,130.60 | |
| Pq3 branch root | Cork | 23.16 | 20.68 | 0.32 | 4.98 | 252,865.9 | 12.32 | 17.92 | 252,945.28 |
| Cortex | 6.69 | 6.20 | 0.39 | 2.06 | 114,430.5 | 3.50 | 4.44 | 114,453.78 | |
| Phloem | 5.17 | 4.43 | 0.32 | 1.56 | 85,663.43 | 0.95 | 3.07 | 85,678.93 | |
| Xylem | 6.18 | 5.95 | 0.27 | 0.30 | 134,882.3 | 0.70 | 9.10 | 134,904.80 | |
| Pq4 main root | Cork | 48.13 | 23.62 | 0.32 | 0.56 | 11,972.59 | 20.01 | 8.35 | 12,073.58 |
| Cortex | 11.50 | 5.10 | 0.31 | 0.84 | 4151.39 | 1.61 | 1.43 | 4172.18 | |
| Phloem | 11.85 | 5.45 | 0.33 | 0.52 | 1685.59 | – | 1.28 | 1705.02 | |
| Secondary xylem | 9.70 | 5.35 | 0.48 | 0.69 | 2659.33 | 1.51 | 1.45 | 2678.51 | |
| Primary xylem | 6.37 | 3.43 | 0.43 | 0.54 | 1766.77 | 0.74 | 0.47 | 1778.75 | |
| Pq4 branch root | Cork | 19.34 | 20.25 | 0.30 | 3.61 | 20,298.81 | 19.70 | 23.67 | 20,385.68 |
| Cortex | 7.09 | 7.63 | 0.40 | 1.48 | 12,388.83 | 5.48 | 7.41 | 12,418.32 | |
| Phloem | 2.94 | 4.66 | 0.32 | 1.07 | 5156.83 | 1.73 | 9.49 | 5177.04 | |
| Xylem | 7.50 | 8.57 | 0.35 | 2.25 | 15,479.39 | 2.95 | 9.38 | 15,510.39 | |
| Pq5 | Cork | 1723.58 | 838.53 | 10.24 | 11.41 | 869.15 | 167.96 | 229.08 | 3849.94 |
| Cortex | 920.69 | 365.92 | 4.64 | 3.35 | 764.67 | 11.79 | 20.06 | 2091.13 | |
| Phloem | 527.99 | 390.62 | 2.07 | 1.50 | 885.82 | 6.30 | 70.71 | 1884.99 | |
| Secondary xylem | 798.60 | 434.04 | 0.95 | 2.28 | 821.14 | 6.09 | 26.04 | 2089.14 | |
| Primary xylem | 1028.47 | 924.56 | 1.19 | 5.86 | 1365.07 | 32.65 | 144.22 | 3502.02 | |
| Pq6 | Cork | 670.07 | 34.99 | 6.03 | 1.00 | 582.31 | 149.25 | 155.58 | 1599.24 |
| Cortex | 320.81 | 13.99 | 3.79 | 0.84 | 364.01 | 41.22 | 47.94 | 792.61 | |
| Phloem | 417.83 | 18.60 | 2.80 | 0.94 | 432.25 | 5.95 | 25.73 | 904.10 | |
| Cambium | 605.12 | 26.43 | 7.10 | 1.03 | 600.16 | 6.08 | 40.85 | 1286.77 | |
| Secondary xylem | 906.45 | 35.99 | 4.26 | 0.85 | 814.07 | 7.77 | 59.66 | 1829.04 | |
| Primary xylem | 1501.30 | 74.73 | 5.11 | 1.32 | 1115.92 | 23.22 | 179.22 | 2900.82 | |
| Pq7 | Cork | 166.40 | 327.34 | 1.71 | 3.93 | 401.22 | 66.79 | 127.19 | 1094.58 |
| Cortex | 174.18 | 207.77 | 1.30 | 2.98 | 163.75 | 16.12 | 24.81 | 590.91 | |
| Phloem | 119.12 | 131.49 | 0.65 | 1.40 | 191.36 | 4.16 | 21.26 | 469.43 | |
| Secondary xylem | 158.80 | 110.53 | 0.60 | 0.88 | 157.42 | 4.33 | 3.80 | 436.35 | |
| Primary xylem | 187.65 | 173.03 | 0.71 | 0.92 | 333.41 | 11.19 | 30.31 | 737.22 | |
| Pq8 | Cork | 149.28 | 1827.33 | 0.67 | 12.70 | 1347.97 | 41.12 | 429.50 | 3808.57 |
| Cortex | 180.35 | 714.05 | 0.74 | 19.35 | 1173.10 | 80.68 | 82.68 | 2250.96 | |
| Phloem | 141.83 | 732.05 | 0.56 | 6.91 | 1002.23 | 9.38 | 49.03 | 1941.99 | |
| Cambium | 144.34 | 723.85 | 0.62 | 9.33 | 1154.96 | 5.40 | 61.17 | 2099.69 | |
| Secondary xylem | 144.52 | 987.34 | 0.80 | 9.24 | 1478.13 | 11.02 | 163.51 | 2794.55 | |
| Primary xylem | 145.17 | 1302.97 | 0.95 | 11.91 | 1365.79 | 12.07 | 218.33 | 3057.19 | |
| Pq9 | Cork | 907.61 | 14.06 | 2.08 | 0.88 | 799.16 | 195.43 | 170.42 | 2089.63 |
| Cortex | 160.10 | 1.99 | – | – | 179.45 | 7.47 | 4.61 | 353.61 | |
| Phloem | 74.54 | 1.52 | – | 0.95 | 60.01 | 4.97 | 2.16 | 144.15 | |
| Secondary xylem | 392.43 | 2.41 | – | – | 430.97 | 3.61 | 22.49 | 851.91 | |
| Primary xylem | 676.25 | 2.78 | 0.84 | – | 1019.24 | 5.45 | 69.53 | 1774.09 | |
| Pq10 | Cork | 668.61 | 712.57 | 0.84 | 6.36 | 986.29 | 19.83 | 67.40 | 2461.89 |
| Cortex | 139.10 | 669.75 | 0.54 | 14.92 | 635.65 | 78.61 | 39.19 | 1577.77 | |
| Phloem | 123.70 | 611.79 | 0.61 | 6.61 | 434.33 | 3.12 | 14.90 | 1195.07 | |
| Secondary xylem | 146.61 | 697.48 | 0.66 | 5.26 | 538.81 | 16.77 | 19.34 | 1424.93 | |
| Primary xylem | 147.68 | 743.10 | 0.62 | 7.22 | 714.65 | 2.12 | 20.17 | 1635.56 | |
| Pq11 | Cork | 62.97 | 537.33 | 0.88 | 5.07 | 511.33 | 65.37 | 188.27 | 1371.23 |
| Cortex | 24.68 | 320.33 | – | 3.50 | 503.92 | 2.51 | 36.75 | 891.69 | |
| Phloem | 21.42 | 344.88 | – | 3.88 | 670.87 | 2.04 | 83.75 | 1126.83 | |
| Secondary xylem | 8.58 | 340.35 | – | 3.60 | 564.94 | 2.98 | 159.17 | 1079.62 | |
| Primary xylem | 22.13 | 619.29 | 0.56 | 7.31 | 916.71 | 3.81 | 364.94 | 1934.75 | |
| Pq12 | Cork | 115.07 | 518.18 | 1.85 | 6.69 | 634.75 | 104.45 | 319.73 | 1700.73 |
| Cortex | 67.05 | 342.52 | 0.55 | 3.34 | 560.80 | 9.01 | 43.48 | 1026.75 | |
| Phloem | 69.79 | 375.80 | – | 3.56 | 871.87 | 3.74 | 48.98 | 1373.75 | |
| Secondary xylem | 45.97 | 610.82 | 0.61 | 4.39 | 1117.76 | 6.46 | 211.26 | 1997.28 | |
| Primary xylem | 147.28 | 871.49 | 0.47 | 5.95 | 1021.44 | 9.45 | 132.19 | 2188.28 | |
| Pq13 | Cork | 82.82 | 766.07 | 0.92 | 22.68 | 568.79 | 88.17 | 125.20 | 1654.66 |
| Cortex | 33.11 | 428.44 | – | 6.46 | 453.02 | 7.45 | 37.26 | 965.74 | |
| Phloem | 34.04 | 547.62 | – | 5.32 | 526.29 | 3.31 | 41.08 | 1157.65 | |
| Secondary xylem | 41.02 | 453.95 | 0.52 | 8.36 | 772.36 | 5.45 | 104.84 | 1386.50 | |
| Primary xylem | 37.32 | 893.53 | – | 14.27 | 922.80 | 3.81 | 166.35 | 2038.07 | |
aGinsenoside
bUnder detection limit
Fig. 5.
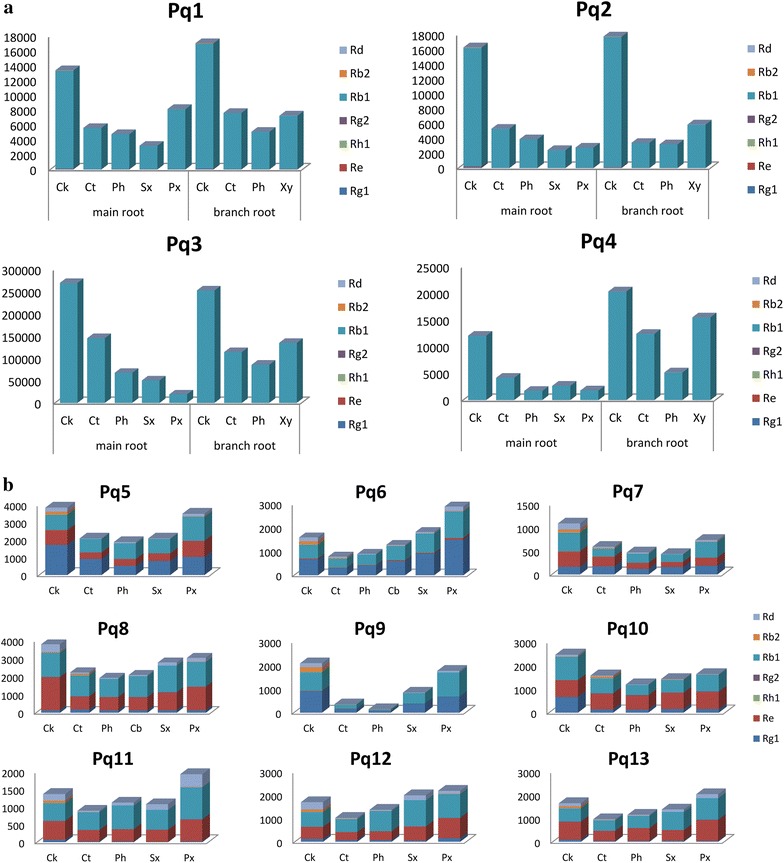
Contents of ginsenosides in different tissues of Pq1-4 (a) and Pq5-13 (b). Ck cork, Ct cortex, Ph phloem, Cb cambium, Sx secondary xylem, Px primary xylem
It was reported that the outer part of the P. quinquefolium root contained more ginsenosides than the center part [25]. However, another paper found that the peak areas of ginsenosides in the center part were larger than those of the outer part [26]. The outer part includes the cork and cortex, while the center part represented the primary xylem for most samples or xylem for branch roots. Our research illustrated that the both situations existed simultaneously in American ginseng.
Although P. quinquefolium and P. ginseng were closely related species which contained many common saponin constituents, their distribution patterns of ginsenosides were quite different. The most obvious difference was that the ginsenosides were not only concentrated in the cork and cortex, but also inclined to be accumulated in the primary xylem in American ginseng. This was identical with the morphological and microscopical characteristics of Asian and American ginseng. In detail, American ginseng was harder than Asian ginseng, and was more difficult to be broken. At the same time, under the fluorescence microscope, it was found that xylem of American ginseng usually differentiated into primary and secondary xylem, while the differentiation was scarely seen in the xylem of Asian ginseng. That is to say that the developed primary xylem was absent in Asian ginseng. The different microscopic structures between American ginseng and Asian ginseng may explain their distinct distribution patterns of ginsenosides in various tissues..
Such similar phenomenon was also found in Bupleuri Radix material. Bupleurum chinense DC. and B. scorzoneri folium Willd. were both original plants of Bupleuri Radix in China. Meanwhile, B. falcatum L. was recorded by Japanese Pharmacopoeia as the original plant of Bupleuri Radix. Recent research found that although saikosaponins were mostly distributed in the cork and cortex in the three species, the cork of B. scorzoneri folium and B. falcatum contained more saikosaponin a, c, d than the cortex, while the opposite situation was found in B. chinense [7]. Thus, the phenomenon that related plants had different distribution patterns of the same secondary metabolites was not an accident. The exact mechanism causing the phenomenon deserved to be further explored.
Conclusion
In conclusion, LMD, fluorescence microscopy, and UHPLC-Q/TOF–MS were applied to profile and determine tissue-specific chemicals of P. quinquefolium in this study. As a result, the cork, primary xylem or cortex had more ginsenosides than phloem, secondary xylem and cambium in American ginseng. Thus, the ratio of total areas of cork, primary xylem and the cortex to the area of the whole transection showed a potential to be used as a reference to judge the quality of American ginsengs.
Experimental
Plant material
As seen from Table 1 and Fig. 1, four fresh P. quinquefolium samples (Pq1–4) were collected from Mulin County, Mudanjiang City, Heilongjiang Province, China. Nine dried samples (Pq5–13) of various commercial types were purchased from Hong Kong herbal markets. All of them were identified by Dr. Zhitao Liang from the School of Chinese Medicine, Hong Kong Baptist University. The voucher specimens were deposited in the Bank of China (Hong Kong) Chinese Medicines Centre of Hong Kong Baptist University. Collected samples were stored at −20 °C before use.
Chemicals and reagents
Chemical standards of ginsenosides Rg1, 20(S)-Rg2, Re, 20(S)-Rh1, Rb1, Rb2 and Rd were purchased from Shanghai Tauto Biotech Company (Shanghai, China). Acetonitrile and methanol of HPLC grade were from E. Merck (Darmstadt, Germany), and formic acid of HPLC grade was from Tedia (Fairfield, USA). Water was prepared by a Milli-Q system (Millipore, Bedford, MA, USA).
Laser microdissection and sample solution preparations
The dried materials were firstly softened by infiltrating with water-soaked-non-cellulose paper before frozen section. The softened and fresh roots were cut into small sections, embedded in cryomatrixTM (Thermo Shandon Limited, U.K.), and then placed on a cutting platform in the cryobar of a cryostat (Thermo Shandon As620 Cryotome, U.K.) at −20 °C. Serial slices of 40 μm in thickness were cut at −10 °C. Each sectioned tissue slice was mounted directly to a non-fluorescent PET microscope steel frame slide (76 mm × 26 mm, 1.4 μm thick, Leica Microsystems, Germany). The slide was observed with a Leica LMD 7000 microscope system (Leica, Benshein, Germany) in fluorescence mode with a dichromatic mirror. Microdissection was conducted by a DPSS laser beam at 349 nm wavelength, aperture of 12, speed of 10, power of 50–60 μJ and pulse frequency of 2895 Hz under a Leica LMD-BGR fluorescence filter system at 10x magnification. Tissue parts within an area of approximately 1 × 106 μm2 were determined as the investigated size and dissected separately under fluorescence inspection mode. The microdissected tissues fell into caps of 500 μL microcentrifuge tubes (Leica, Germany) by gravity.
The separated tissue part in each cap was transferred to the bottom of the tube through centrifugation (Centrifuge 5415R, Eppendorf, Hamburg, Germany) at 12,000 rpm for 5 min. 100 μL methanol was added into each microcentrifuge tube. The tube was sonicated for 30 min (CREST 1875HTAG ultrasonic processor, USA). The microcentrifuge tube was centrifuged again for 10 min at 12,000 rpm, and 4 °C. 90 μL of the supernatant was transferred to a glass insert with plastic bottom spring (400 μL, Grace, USA) in a 1.5 mL brown HPLC vial (Grace, USA) and stored at 4 °C for analysis.
Qualitative and quantitative analysis
UHPLC-QTOF–MS analysis was performed on an Agilent 6540 ultra-high definition accurate mass quadrupole time-of-flight spectrometer with UHPLC (UHPLC-QTOF–MS, Agilent Technologies, USA). A UPLC C18 analytical column (2.1 mm × 100 mm, I.D. 1.7 μm, ACQUITY UPLC® BEH, Waters, USA) was used for separation, coupled with a C18 pre-column (2.1 mm × 5 mm, I.D. 1.7 μm, VanGuardTM BEH, Waters, USA) at room temperature of 20 °C. The mobile phase was a mixture of water (A) and acetonitrile (B), both containing 0.1 % formic acid, with an optimized linear gradient elution as follows: 0–3 min, 10–20 % B; 3–25 min, 20–38 % B; 25–30 min, 38–85 % B; 30–30.1 min, 85–100 % B; 30.1–32 min, 100 % B; 32–32.1 min 100–10 % B with 4 min of balance. The injection volume was 3 μL for tissue sample. The flow rate was set at 0.35 mL/min. The mass spectra were acquired in negative mode by scanning from 100 to 1700 in mass to charge ratio (m/z). The MS analysis was performed under the following operation parameters: dry gas temperature 300 °C, dry gas (N2) flow rate 8 L/min, nebulizer pressure 45 psi, Vcap 3000, nozzle voltage 500 V, and fragmentor voltage 180 V. The energies for collision-induced dissociation (CID) were set at 30 and 45 eV respectively for the fragmentation information.
Data analysis was performed with Agilent MassHunter Workstation software-Qualitative Analysis and Q-TOF Quantitative Analysis (version B.04.00, Build 4.0.479.5, Service Pack 3, Agilent Technologies, Inc. 2011). By searching databases including PubMed of the US National Library Medicine and the National Institutes of Health, Scifinder Scholar of American Chemical Society and Chinese National Knowledge Infrastructure (CNKI) of Tsinghua University, all chemicals reported in the literatures as derived from Panax species were summarized in a Microsoft Office Excel table to establish a database, which includes the name, molecular formula, and molecular weight of each chemical. The “Search Database” in the “Identify Compounds” in Agilent MassHunter Workstation software-Qualitative Analysis was used to identify the chromatographic peaks.
To semi-quantitatively determine the spatial distributions of the individual metabolites in different tissue regions, the contents of chemical markers including ginsenosides Rg1, 20(S)-Rg2, Re, 20(S)-Rh1, Rb1, Rb2 and Rd in various microdissected tissues were relatively determined using the above UHPLC-QTOF–MS method. Linearity was examined within selected concentration range with different levels and applied to calculate the amounts of these analytes in tissue extracts.
Authors’ contributions
ZL initiated and all authors designed the study. YC and YZ carried out the experimental study. YC drafted the manuscript. LX collected the herbal samples. All authors contributed to the data analysis and to finalizing the manuscript. All authors read and approved the final version.
Acknowledgements
This work is sponsored by the Faculty Research Grant of Hong Kong Baptist University (FRG2/12-13/030) and Innovation and Technology Fund (ITS/185/13FX). We are grateful to Mr. Alan Ho from the School of Chinese Medicine, Hong Kong Baptist University for his technical support.
Competing interests
The authors declare that they have no competing interests.
Abbreviations
- LMD
laser microdissection
- UHPLC-Q/TOF-MS
ultra-high performance liquid chromatography-quadrupole/time-of-flight- mass spectrometry
Contributor Information
Yujie Chen, Email: chenyujie.1986@163.com.
Liang Xu, Email: 861364054@qq.com.
Yuancen Zhao, Email: yuan.cen@icloud.com.
Zhongzhen Zhao, Email: zzzhao@hkbu.edu.hk.
Hubiao Chen, Email: hbchen@hkbu.edu.hk.
Tao Yi, Email: yitao@hkbu.edu.hk.
Minjian Qin, Phone: +86 25 86185130, Email: minjianqin@163.com.
Zhitao Liang, Phone: +852 1534118066, Email: lzt23@hkbu.edu.hk.
References
- 1.Cai X, Zhang AX, Wu H, Hu ZH. Histochemistry of sinomenine in the stem of Sinomenium acutum and Sinomenium acutum var. cinereum. Acta BotBoreali-Occidentalia Sin. 1999;19:104–107. [Google Scholar]
- 2.Shen ZG, Chauser-volfson E, Gutterman Y, Hu ZH. Anatomy, histochemistry and phytochemistry of leaves in Aloe vera var. chinensis. Acta Bot Sin. 2001;43:780–787. [Google Scholar]
- 3.Lin R, Cao YF, Hu ZH. Anatomical structure of vegetative organs and histochemical localization of ginsenosides in Gynostemma pentaphyllum. Acta Bot Boreali-Occidentalia Sin. 2002;22:796–800. [Google Scholar]
- 4.Cao YF, Lin R, Hu ZH. Studies on the developmental anatomy of Rhizome of Dioscorea zingiberensis and its histochemistry. J Wuhan Bot Res. 2003;21:288–294. [Google Scholar]
- 5.Qiao Q, Xiao YP, Wang ZZ. Anatomical structure and histochemical localization of the drupe of Macrocarpium officinacle. Acta Bot Yunnanica. 2004;26:651–655. [Google Scholar]
- 6.Liang ZT, Sham TT, Yang GY, Yi L, Chen HB, Zhao ZZ. Profiling of secondary metabolites in tissues from Rheum palmatum L. using laser microdissection and liquid chromatography mass spectrometry. Anal Bioanal Chem. 2013;405:4199–4212. doi: 10.1007/s00216-013-6819-z. [DOI] [PubMed] [Google Scholar]
- 7.Liang ZT, Oh KY, Wang YQ, Yi T, Chen HB, Zhao ZZ. Cell type-specific qualitative and quantitative analysis of saikosaponins in three Bupleurum species using laser microdissection and liquid chromatography–quadrupole/time of flight-mass spectrometry. J Pharm Biomed Anal. 2014;97:157–165. doi: 10.1016/j.jpba.2014.04.033. [DOI] [PubMed] [Google Scholar]
- 8.Liang ZT, Chen YJ, Xu L, Qin MJ, Yi T, Chen HB, Zhao ZZ. Localization of ginsenosides in the rhizome and root of Panaxginseng by laser microdissection and liquid chromatography–quadrupole/time of flight-mass spectrometry. J Pharm Biomed Anal. 2015;105c:121–133. doi: 10.1016/j.jpba.2014.12.005. [DOI] [PubMed] [Google Scholar]
- 9.Yi L, Liang ZT, Peng Y, Yao X, Chen HB, Zhao ZZ. Tissue-specific metabolite profiling of alkaloids in Sinomenii Caulis using laser microdissection and liquid chromatography-quadrupole/time of flight-mass spectrometry. J Chromatogr A. 2012;1248:93–103. doi: 10.1016/j.chroma.2012.05.058. [DOI] [PubMed] [Google Scholar]
- 10.Chen YJ, Liang ZT, Zhu Y, Xie GY, Tian M, Zhao ZZ, Qin MJ. Tissue-specific metabolites profiling and quantitative analyses of flavonoids in the rhizome of Belamcanda chinensis by combining laser-microdissection with UHPLC-Q/TOF-MS and UHPLC-QqQ-MS. Talanta. 2014;130:585–597. doi: 10.1016/j.talanta.2014.07.004. [DOI] [PubMed] [Google Scholar]
- 11.Jaiswal Y, Liang ZT, Ho A, Wong LL, Yong P, Chen HB, Zhao ZZ. Distribution of toxic alkaloids in tissues from three herbal medicine Aconitum species using laser micro-dissection, UHPLC–QTOF MS and LC–MS/MS techniques. Phytochemistry. 2014;107:155–174. doi: 10.1016/j.phytochem.2014.07.026. [DOI] [PubMed] [Google Scholar]
- 12.Sun BS, Xu MY, Li Z, Wang YB, Sang CK. UPLC-Q-TOF-MS/MS analysis for steaming times-dependent profiling of steamed Panax quinquefolius and its ginsenosides transformations induced by repetitious steaming. J Ginseng Res. 2012;36:277–290. doi: 10.5142/jgr.2012.36.3.277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Attele AS, Wu JA, Yuan CS. Ginseng pharmacology: multiple constituents and multiple actions. Biochem Pharmacol. 1999;58:1685–1693. doi: 10.1016/S0006-2952(99)00212-9. [DOI] [PubMed] [Google Scholar]
- 14.Dou DQ, Hou WB, Chen YJ. Studies on the characteristic constituents of Chinese ginseng and American ginseng. Planta Med. 1998;64:585–586. doi: 10.1055/s-2006-957526. [DOI] [Google Scholar]
- 15.Qi LW, Wang CZ, Yuan CS. Ginsenosides from American ginseng: chemical and pharmacological diversity. Phytochemistry. 2011;72:689–699. doi: 10.1016/j.phytochem.2011.02.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Yuan CS, Wang CZ, Wicks SM, Qi LW. Chemical and pharmacological studies of saponins with a focus on American ginseng. J Ginseng Res. 2010;34:160–167. doi: 10.5142/jgr.2010.34.3.160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Corbit RM, Ferreira JFS, Ebbs SD, Murphy LL. Simplified extraction of ginsenosides from American ginseng (Panax quinquefolius L.) for high-performance liquid chromatography-ultraviolet analysis. J Agric Food Chem. 2005;53:9867–9873. doi: 10.1021/jf051504p. [DOI] [PubMed] [Google Scholar]
- 18.Wang A, Wang CZ, Wu JA, Osinski J. Determination of major ginsenosides in Panax quinquefolius (American ginseng) using high-performance liquid chromatography. Phytochem Anal. 2005;16:272–277. doi: 10.1002/pca.838. [DOI] [PubMed] [Google Scholar]
- 19.Qu CL, Bai YP, Jin XQ, Wang YT, Zhang K, You JY, Zhang HQ. Study on ginsenosides in different parts and ages of Panax quinquefolius L. Food Chem. 2009;115:340–346. doi: 10.1016/j.foodchem.2008.11.079. [DOI] [Google Scholar]
- 20.Zheng CN, Hao HP, Wang X, Wang XL, Wang GJ, Sang GW, Liang Y, Xie L, Xia CH, Yao XL. Diagnostic fragment-ion-based extension strategy for rapid screening and identification of serial components of homologous families contained in traditional Chinese medicine prescription using high-resolution LC-ESI-IT-TOF/MS: Shengmai injection as an example. J Mass Spectrom. 2009;44:230–244. doi: 10.1002/jms.1502. [DOI] [PubMed] [Google Scholar]
- 21.Qi LW, Wang CZ, Yuan CS. Isolation and analysis of ginseng: advances and challenges. Nat Prod Rep. 2011;28:467–495. doi: 10.1039/c0np00057d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Li SL, Shen H, Zhu LY, Xu J, Jia XB, Zhang HM, Lin G, Cai H, Cai BC, Chen SL, Xu HX. Ultra-high-performance liquid chromatography-quadrupole/time of flight mass spectrometry based chemical profiling approach to rapidly reveal chemical transformation of sulfur-fumigated medicinal herbs, a case study on white ginseng. J Chromatogr A. 2012;1231:31–45. doi: 10.1016/j.chroma.2012.01.083. [DOI] [PubMed] [Google Scholar]
- 23.Zhang HM, Li SL, Zhang H, Wang Y, Zhao ZL, Chen SL, Xu HX. Holistic quality evaluation of commercial white and red ginseng using a UPLC-QTOF-MS/MS-based metabolomics approach. J Pharm Biomed Anal. 2012;62:258–273. doi: 10.1016/j.jpba.2012.01.010. [DOI] [PubMed] [Google Scholar]
- 24.Du XW, Wills RBH, Stuart DL. Changes in neutral and malonyl ginsenosides in American ginseng (Panax quinquefolium) during drying, storage and ethanolic extraction. Food Chem. 2004;86:155–159. doi: 10.1016/j.foodchem.2003.11.003. [DOI] [Google Scholar]
- 25.Liu WC, Zhang MP, Li CS, Sun CY, Jiang SC, Li C, Wang Y. Determination of ginsenosides for different parts in Panax quinquefolium L. by HPLC. Chin J Ginseng Res. 2012;22:20–23. [Google Scholar]
- 26.Qu YX, Wang ZZ. A study on saponins for different parts in Panax quinquefolium L. Chin J Prog in Modern Biomed. 2006;6:32–35. [Google Scholar]