

Abstract
Heartland virus (HRTV), a phlebovirus first isolated from two Missouri farmers in 2009, has been proposed to be transmitted to humans by the bite of infected Amblyomma americanum ticks. It is closely related to severe fever with thrombocytopenia syndrome virus (SFTSV) from China, another previously unrecognized phlebovirus that has subsequently been associated with hundreds of cases of severe disease in humans. To expand diagnostic capacity to detect HRTV infections, 20 hybridoma clones secreting anti-HRTV murine monoclonal antibodies (MAbs) were developed using splenocytes from HRTV-inoculated AG129 alpha/beta and gamma interferon receptor–deficient mice. Nine of these MAbs were characterized herein for inclusion in future HRTV diagnostic assay development. All of the MAbs developed were found to be non-neutralizing and reactive to linear epitopes on HRTV nucleocapsid protein. MAb 2AF11 was found to be cross-reactive with SFTSV.
Heartland virus (HRTV) is a newly identified virus member of the Phlebovirus genus in the family Bunyaviridae. It was first isolated from two hospitalized Missouri farmers in separate locations within the state in 2009.1 HRTV has been proposed to be transmitted to humans by the bite of infected Amblyomma americanum ticks.2 It causes severe disease characterized by fever, leukopenia, and thrombocytopenia.1 HRTV is closely related to severe fever with thrombocytopenia virus (SFTSV), a phlebovirus causing severe disease in China and neighboring countries.3 The average case fatality rate of SFTSV infection is between 6% and 17% with the most severe manifestations occurring in elderly individuals.4 HRTV has a single-stranded RNA genome of negative or ambisense polarity encoded on three segments. The large (L) segment encodes the RNA-dependent-RNA polymerase, the medium (M) segment encodes a precursor of the glycoproteins Gn and Gc, and the small (S) segment encodes the nucleocapsid (N) protein and a nonstructural (Ns) protein.5
Serological assays for the detection of HRTV are currently limited to the detection of neutralizing antibodies by plaque reduction neutralization test (PRNT). The limitation in the range of assays developed is due to the absence of suitable anti-HRTV monoclonal antibodies (MAbs). To develop diagnostic assays able to detect both recent and prior infections and to assess the disease burden of HRTV infection in the United States, anti-HRTV murine MAbs were developed and characterized. Interferon receptor–deficient AG129 mice approximately 3 weeks of age were inoculated intraperitoneally (IP) with 100 plaque-forming units (PFU) of HRTV strain MO-4. After 30 days, mice were bled and boosted with another 100 PFU of HRTV MO-4 IP. Splenocytes were harvested 4 days after the last inoculation for fusions with the mouse myeloma cell line P3X63Ag8.653 using the ClonaCell-HY Hybridoma cloning kit (StemCell Technologies, Vancouver, British Columbia, Canada).
This is the first report of the use of activated B cells from AG129 mice for the development of hybridomas. B-cell hybridoma clones are generally made by isolating activated B cells from the spleen of an immunized BALB/c mouse or mouse with a compatible major histocompatibility complex (MHC) haplotype (H2d) and fusing with a myeloma cell with the same haplotype. In this case, we used AG129 mice as B cell donors that have a different MHC haplotype (H2b) from the P3X63Ag8.653 myeloma cells used for fusions. Although the use of BALB/c mice may be appropriate for most infectious agents, some viruses may fail to initiate replication in immunocompetent mice and thus fail to mount a robust immune response. Using AG129 mice for hybridoma development may offer an alternative approach for developing MAbs for viruses incapable of replication in immunocompetent mice.
Sera from two HRTV infected mice taken on days 30 and 34 postinoculation (dpi) were assayed by enzyme-linked immunosorbent assay (ELISA) using purified HRTV at a dilution of 0.06 μg/well coated overnight at 4°C to 96-well plates in 50 mM sodium carbonate/50 mM sodium bicarbonate buffer, pH 9.6. Plates were washed five times in phosphate-buffered saline (PBS)/0.05% Tween before nonspecific binding sites were blocked with StartingBlock (ThermoFisher Scientific, Grand Island, NY). Sera diluted in PBS were incubated on the plates for 1 hour at 37°C. Plates were washed again before goat anti-mouse IgG conjugated to horseradish peroxidase diluted 1:5000 in PBS was incubated on the plates. After plates were washed a final time, reactions were developed using TMB K-blue substrate (KPL, Gaithersburg, MD) and stopped with the addition of 1 N H2SO4 before being read at 450 nm. On 30 dpi mice 1 and 2 had ELISA endpoint titers of < 2.7 log10 and ≥ 4.6 log10, respectively, indicating that mouse 1 did not develop an infection after the initial inoculation. On 34 dpi, those ELISA titers increased to 2.7 log10 and 5.6 log10, respectively, while PRNT80 titers on Vero cells were 1.9 log10 and 2.5 log10, respectively (Table 1).
Table 1.
HRTV-specific antibody responses in AG129 mice after primary and secondary immunizations with HRTV
| Mouse no. | Geometric mean reciprocal ELISA titer (log10) | Geometric mean reciprocal PRNT80 (log10) | |
|---|---|---|---|
| 30 DPI | 34 DPI | 34 DPI | |
| 1 | < 2.7 | 2.7 | 1.9 |
| 2 | ≥ 4.6 | 5.6 | 2.5 |
DPI = days postinfection; ELISA = enzyme-linked immunosorbent assay; HRTV = Heartland virus; PRNT = plaque reduction neutralization test.
To determine the viral protein specificity of the antibody response, purified HRTV (5 μg/well) was run on a 4–12% Bis-Tris polyacrylamide gel (ThermoFisher Scientific) under reducing conditions. Proteins were blotted electrophoretically from the gels onto nitrocellulose membranes and washed for 15 minutes in PBS/0.1% Tween wash buffer. Nonspecific binding sites were blocked with 10% goat serum in PBS for 1 hour while rocking. Sera diluted 1:200 were incubated with the membrane for 1 hour with gentle rocking. Membranes were washed again before goat anti-mouse conjugated to alkaline phosphatase (1:200; Jackson ImmunoResearch, West Grove, PA) was incubated on the membrane for 1 hour with gentle rocking. Membranes were washed and BCIP/NBT phosphatase substrate (KPL) was added until a color change appeared. Even though neutralizing antibody titers were detected for both mice, only mouse 2 produced IgG anti-glycoprotein (Gn and Gc) and anti-N antibody to HRTV detectable by western blot on 34 dpi (Figure 1 ).
Figure 1.
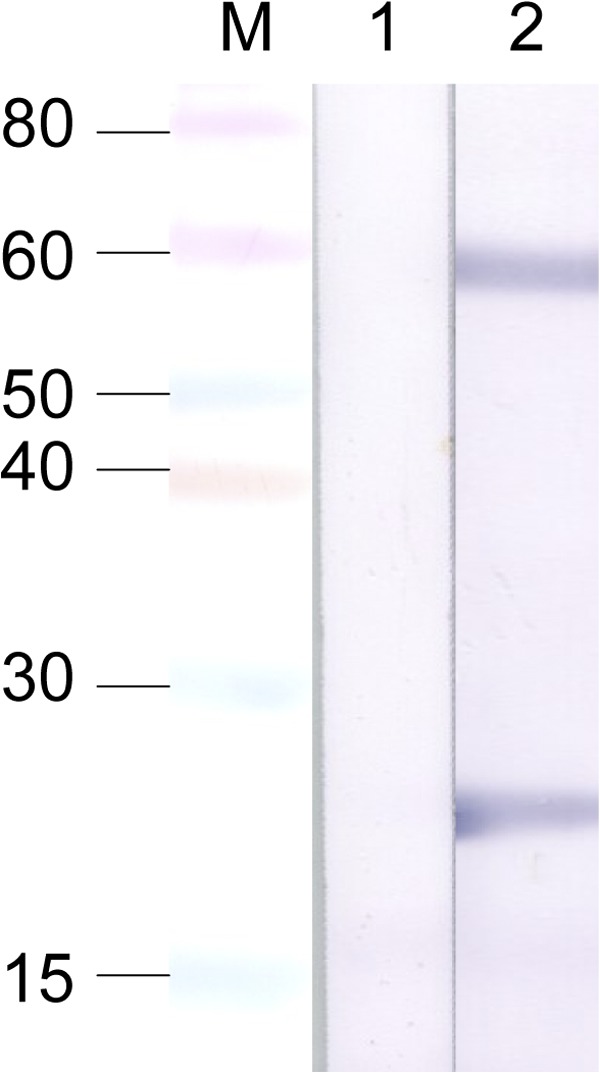
Reactivity of pooled mouse sera from 34 days postinoculation. M = Novex pre-stained low molecular mass marker (sizes are shown in kDa); lane 1 = mouse 1; lane 2 = mouse 2.
After the heterologous fusions, hybridomas were grown in high-glucose Dulbecco minimal essential medium supplemented with l-glutamine, 10% low IgG fetal bovine serum, 1 mM sodium pyruvate, 2 mM l-glutamine, 0.15% sodium bicarbonate, 100 U/mL penicillin G sodium, 100 μg/mL streptomycin sulfate, and 0.1 mM nonessential amino acids. MAbs were purified from highly concentrated culture media using protein-A sepharose (GE Healthcare, Pittsburgh, PA) according to the manufacturer's instructions. Purified MAbs were dialyzed against PBS and the concentration was measured by Bradford protein assay (Bio-Rad, Hercules, CA).
Twenty hybridoma clones secreting anti-HRTV MAbs were isolated from fusions. Of these, 19 were from fusions using splenocytes from mouse 2, while only one hybridoma clone was isolated from fusions using splenocytes from mouse 1. Of the hybridoma clones, 12 were of varying immunoglobulin (Ig) G isotypes, while the other eight were IgM (Table 2). Nine of the IgG MAbs were titrated by ELISA as previously described with starting concentrations of purified MAbs at 104 ng/mL (Table 2). An optical density of 2-fold the background was considered a positive result, and endpoints were expressed in ng/mL. The MAbs with the highest reactivities by ELISA were 2BB5, 2AG8, 2BB8, and 2AG9 with endpoint concentrations of 0.156, 0.381, 0.610, and 1.526 ng/mL, respectively. The MAbs with moderate reactivities by ELISA were 2BB7, 2BA2, and 2BB10 with endpoint concentrations of 24.41, 39.06, and 937.5 ng/mL. Two MAbs (2AF11 and 2BB6) had the lowest ELISA reactivities (104 ng/mL endpoints) (Table 2).
Table 2.
Serological and biological characteristics of anti-HRTV MAbs
| MAb | Isotype | ELISA endpoint* | Reciprocal PRNT80† | Immunoblot‡ | Cross-reactivity with SFTSV§ | |
|---|---|---|---|---|---|---|
| −ME | +ME | |||||
| 2BB5 | IgG3 | 0.156 | > 104 | N | N | NEG |
| 2AG8 | IgG2a | 0.381 | > 104 | N | N | NEG |
| 2BB8 | IgG2a | 0.610 | > 104 | N | N | NEG |
| 2AG9 | IgG2a | 1.526 | > 104 | N | N | NEG |
| 2BB7 | IgG2b | 24.41 | > 104 | N | N | NEG |
| 2BA2 | IgG1 | 39.06 | > 104 | N | N | NEG |
| 2BB10 | IgG1 | 937.5 | > 104 | N | N | NEG |
| 2AF11 | IgG2a | 104 | > 104 | N | N | 103 |
| 2BB6 | IgG2b | 104 | > 104 | N | N | NEG |
ELISA = enzyme-linked immunosorbent assay; HRTV = Heartland virus; MAb = monoclonal antibody; ME = β-mercaptoethanol; N = nucleocapsid; NEG = negative reactivity with SFTSV-infected cells; PRNT = plaque reduction neutralization test; SFTSV = severe fever with thrombocytopenia syndrome virus.
ELISA endpoint titers expressed as ng/mL.
PRNT endpoint titers expressed as ng/mL.
Protein specificities.
Cross-reactivity with SFTSV antigen determined by immunofluorescence assay (IFA) on SFTSV-infected Vero cells. IFA endpoint titers expressed as ng/mL.
To assess whether these MAbs could neutralize virus in vitro, purified MAbs were tested in a PRNT with HRTV. About 100 PFU of HRTV MO-4 were incubated with equal amounts of serial 2-fold dilutions of purified antibody starting at 104 ng/mL for 1 hour at 37°C. Six-well plates of Vero cells were then inoculated with the virus–antibody mixtures and incubated at 37°C with 5% CO2 for 1 hour after which cells were overlaid with 3 mL of medium containing 1% SeaKem LE agarose (Lonza, Rockland, ME) in nutrient medium (0.165% lactalbumin hydrolysate, 0.033% yeast extract, Earle's balanced salt solution, and 2% FBS). After incubation at 37°C for 7 days a second overlay containing an additional 80 μg/mL of neutral red vital stain (ThermoFisher Scientific) was added. Plaques were counted on day 9 and 10. None of the MAbs were found to neutralize virus in vitro when tested by PRNT using an 80% reduction in plaque formation as significant (PRNT80) (Table 2).
Viral glycoprotein specificity for these MAbs was determined by immunoblot as described above using purified HRTV with or without the reducing agent, β-mercaptoethanol. All anti-HRTV MAbs reacted specifically with the N protein from purified virus, which is approximately 20 kDa in mass. These MAbs were reactive under reducing and nonreducing conditions suggesting that all nine MAbs recognize linear epitopes on the N protein (Table 2).
To determine the cross-reactivity of anti-HRTV MAbs with SFTSV, Vero cells were inoculated with SFTSV, harvested 5 days after infection, and fixed to 12-well glass slides in 70% acetone in PBS. MAbs were added in doubling dilutions (20 μL/well) to the wells of the slide and incubated at 37°C for 30 minutes. Slides were washed three times in PBS before goat anti-mouse conjugated to fluorescein isothiocyanate (FITC) (ThermoFisher Scientific) diluted 1:200 in PBS was added (20 μL/well) and incubated at 37°C for 30 minutes. Only MAb 2AF11 was found to be cross-reactive with SFTSV with an endpoint titer of 103 ng/mL (Table 2).
This study reports the first generation of a panel of murine MAbs directed to HRTV as well as highlights the immunodominance of the nucleoprotein in HRTV infections. The majority of anti-SFTSV human and murine MAbs developed have been nucleocapsid-specific and non-neutralizing.6,7 The exception was a hMAb recognizing a linear epitope in the ectodomain of Gn found to be capable of neutralizing SFTSV in vitro by blocking virus cell–mediated attachment.7 The N protein has also been shown to be the immunodominant protein in other phleboviral infections including Rift Valley fever virus (RFTV) and Toscana (TOSV) viruses.8,9 It is highly conserved in bunyaviruses and is the most abundantly expressed viral protein in virus-infected cells.9–11 Even though the N protein does not produce neutralizing antibody, MAbs directed against this antigen can be protective in vivo against RVFV and Hantaan virus infections.12–15 Although neutralizing antibodies in bunyaviral infections are directed to the Gn or Gc glycoproteins, these antibodies may be harder to isolate since they usually recognize conformationally dependent epitopes requiring higher binding avidities.9,16,17
Serologic diagnostic assays that do not rely on the detection of neutralizing antibody and biosafety level 3 containment are needed for the detection of HRTV-specific IgM and IgG. Recombinant N protein has been used in diagnostic applications for phleboviruses including RVFV and SFTSV.18–20 An assay for the detection of total anti-HRTV antibody reactivity in serum samples may be more beneficial than neutralization assays that require special containment. The MAbs described herein are currently being evaluated for their inclusion in microsphere immunoassays and ELISAs for the detection of HRTV-specific IgM and IgG in human infections.
ACKNOWLEDGMENTS
We thank Olga Kosoy, Jason Velez, and Mary Crabtree for technical assistance.
Footnotes
Authors' addresses: Amanda E. Calvert, Arboviral Diseases Branch, U.S. Centers for Disease Control, Fort Collins, CO, E-mail: zpz0@cdc.gov. Aaron C. Brault, Division of Vector-Borne Diseases, Centers for Disease Control and Prevention, Fort Collins, CO, E-mail: acbrault1@mac.com.
References
- 1.McMullan LK, Folk SM, Kelly AJ, MacNeil A, Goldsmith CS, Metcalfe MG, Batten BC, Albarino CG, Zaki SR, Rollin PE, Nicholson WL, Nichol ST. A new phlebovirus associated with severe febrile illness in Missouri. N Engl J Med. 2012;367:834–841. doi: 10.1056/NEJMoa1203378. [DOI] [PubMed] [Google Scholar]
- 2.Savage HM, Godsey MS, Jr, Lambert A, Panella NA, Burkhalter KL, Harmon JR, Lash RR, Ashley DC, Nicholson WL. First detection of heartland virus (Bunyaviridae: Phlebovirus) from field collected arthropods. Am J Trop Med Hyg. 2013;89:445–452. doi: 10.4269/ajtmh.13-0209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Yu XJ, Liang MF, Zhang SY, Liu Y, Li JD, Sun YL, Zhang L, Zhang QF, Popov VL, Li C, Qu J, Li Q, Zhang YP, Hai R, Wu W, Wang Q, Zhan FX, Wang XJ, Kan B, Wang SW, Wan KL, Jing HQ, Lu JX, Yin WW, Zhou H, Guan XH, Liu JF, Bi ZQ, Liu GH, Ren J, Wang H, Zhao Z, Song JD, He JR, Wan T, Zhang JS, Fu XP, Sun LN, Dong XP, Feng ZJ, Yang WZ, Hong T, Zhang Y, Walker DH, Wang Y, Li DX. Fever with thrombocytopenia associated with a novel bunyavirus in China. N Engl J Med. 2011;364:1523–1532. doi: 10.1056/NEJMoa1010095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Xu B, Liu L, Huang X, Ma H, Zhang Y, Du Y, Wang P, Tang X, Wang H, Kang K, Zhang S, Zhao G, Wu W, Yang Y, Chen H, Mu F, Chen W. Metagenomic analysis of fever, thrombocytopenia and leukopenia syndrome (FTLS) in Henan Province, China: discovery of a new bunyavirus. PLoS Pathog. 2011;7:e1002369. doi: 10.1371/journal.ppat.1002369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Walter CT, Barr JN. Recent advances in the molecular and cellular biology of bunyaviruses. J Gen Virol. 2011;92:2467–2484. doi: 10.1099/vir.0.035105-0. [DOI] [PubMed] [Google Scholar]
- 6.Yu L, Zhang L, Sun L, Lu J, Wu W, Li C, Zhang Q, Zhang F, Jin C, Wang X, Bi Z, Li D, Liang M. Critical epitopes in the nucleocapsid protein of SFTS virus recognized by a panel of SFTS patients derived human monoclonal antibodies. PLoS One. 2012;7:e38291. doi: 10.1371/journal.pone.0038291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Guo X, Zhang L, Zhang W, Chi Y, Zeng X, Li X, Qi X, Jin Q, Zhang X, Huang M, Wang H, Chen Y, Bao C, Hu J, Liang S, Bao L, Wu T, Zhou M, Jiao Y. Human antibody neutralizes severe fever with thrombocytopenia syndrome virus, an emerging hemorrhagic fever virus. Clin Vaccine Immunol. 2013;20:1426–1432. doi: 10.1128/CVI.00222-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Martin-Folgar R, Lorenzo G, Boshra H, Iglesias J, Mateos F, Borrego B, Brun A. Development and characterization of monoclonal antibodies against Rift Valley fever virus nucleocapsid protein generated by DNA immunization. MAbs. 2010;2:275–284. doi: 10.4161/mabs.2.3.11676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Magurano F, Nicoletti L. Humoral response in Toscana virus acute neurologic disease investigated by viral-protein-specific immunoassays. Clin Diagn Lab Immunol. 1999;6:55–60. doi: 10.1128/cdli.6.1.55-60.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Schwarz TF, Gilch S, Pauli C, Jager G. Immunoblot detection of antibodies to Toscana virus. J Med Virol. 1996;49:83–86. doi: 10.1002/(SICI)1096-9071(199606)49:2<83::AID-JMV2>3.0.CO;2-F. [DOI] [PubMed] [Google Scholar]
- 11.Swanepoel R, Struthers JK, Erasmus MJ, Shepherd SP, McGillivray GM, Erasmus BJ, Barnard BJ. Comparison of techniques for demonstrating antibodies to Rift Valley fever virus. J Hyg (Lond) 1986;97:317–329. doi: 10.1017/s0022172400065414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Boshra H, Lorenzo G, Rodriguez F, Brun A. A DNA vaccine encoding ubiquitinated Rift Valley fever virus nucleoprotein provides consistent immunity and protects IFNAR(−/−) mice upon lethal virus challenge. Vaccine. 2011;29:4469–4475. doi: 10.1016/j.vaccine.2011.04.043. [DOI] [PubMed] [Google Scholar]
- 13.Jansen van Vuren P, Tiemessen CT, Paweska JT. Anti-nucleocapsid protein immune responses counteract pathogenic effects of Rift Valley fever virus infection in mice. PLoS One. 2011;6:e25027. doi: 10.1371/journal.pone.0025027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Nakamura T, Yanagihara R, Gibbs CJ, Jr, Amyx HL, Gajdusek DC. Differential susceptibility and resistance of immunocompetent and immunodeficient mice to fatal Hantaan virus infection. Arch Virol. 1985;86:109–120. doi: 10.1007/BF01314117. [DOI] [PubMed] [Google Scholar]
- 15.Yoshimatsu K, Yoo YC, Yoshida R, Ishihara C, Azuma I, Arikawa J. Protective immunity of Hantaan virus nucleocapsid and envelope protein studied using baculovirus-expressed proteins. Arch Virol. 1993;130:365–376. doi: 10.1007/BF01309667. [DOI] [PubMed] [Google Scholar]
- 16.Besselaar TG, Blackburn NK. Topological mapping of antigenic sites on the Rift Valley fever virus envelope glycoproteins using monoclonal antibodies. Arch Virol. 1991;121:111–124. doi: 10.1007/BF01316748. [DOI] [PubMed] [Google Scholar]
- 17.Lundkvist A, Horling J, Niklasson B. The humoral response to Puumala virus infection (nephropathia epidemica) investigated by viral protein specific immunoassays. Arch Virol. 1993;130:121–130. doi: 10.1007/BF01319001. [DOI] [PubMed] [Google Scholar]
- 18.Paweska JT, van Vuren PJ, Kemp A, Buss P, Bengis RG, Gakuya F, Breiman RF, Njenga MK, Swanepoel R. Recombinant nucleocapsid-based ELISA for detection of IgG antibody to Rift Valley fever virus in African buffalo. Vet Microbiol. 2008;127:21–28. doi: 10.1016/j.vetmic.2007.07.031. [DOI] [PubMed] [Google Scholar]
- 19.Paweska JT, Mortimer E, Leman PA, Swanepoel R. An inhibition enzyme-linked immunosorbent assay for the detection of antibody to Rift Valley fever virus in humans, domestic and wild ruminants. J Virol Methods. 2005;127:10–18. doi: 10.1016/j.jviromet.2005.02.008. [DOI] [PubMed] [Google Scholar]
- 20.Jiao Y, Zeng X, Guo X, Qi X, Zhang X, Shi Z, Zhou M, Bao C, Zhang W, Xu Y, Wang H. Preparation and evaluation of recombinant severe fever with thrombocytopenia syndrome virus nucleocapsid protein for detection of total antibodies in human and animal sera by double-antigen sandwich enzyme-linked immunosorbent assay. J Clin Microbiol. 2012;50:372–377. doi: 10.1128/JCM.01319-11. [DOI] [PMC free article] [PubMed] [Google Scholar]