

Abstract
Background
Increasing numbers of genetic variants are being recognized among piroplasms, but the precise taxonomical status, the tick vector and the geographical range of several species or genotypes are still unknown. Bovine piroplasmosis was reported to re-emerge in north-east Hungary. Because Theileria-infection was newly diagnosed in one cattle herd in the same region of the country, the aim of this study was to molecularly identify the relevant agent, to find its local vector tick species, and to examine the range of Babesia/Theileria spp. of ruminants in Haemaphysalis sp. ticks collected previously in Hungary.
Findings
Blood samples were drawn on two occasions from 90 dairy cattle in northern Hungary, and ticks were collected on their pastures. In addition, questing ticks (315 Haemaphysalis inermis, 259 H. concinna and 22 H. punctata), which originated mainly in the same region of the country from 2007, were included in the study. DNA was extracted from these samples, followed by molecular analysis for piroplasms.
In the cattle Theileria orientalis was identified, with 100 % sequence homology to isolates from Japan, China, South-Africa and Australia. Based on GenBank data this genotype has not been previously reported in Europe. The prevalence of infection in the herd remained almost constant in the main tick season, suggesting exposure in previous years.
Retrospective analysis of ticks revealed the presence of Babesia crassa in H. inermis, for the first time in Europe and in this tick species. On the other hand, H. concinna carried five different piroplasms, including B. motasi that was also newly detected in Central-Eastern Europe and in this tick species; whereas H. punctata harboured Theileria sp. OT3, hitherto known to occur in the Mediterranean region.
Conclusions
Results of this study broaden the range of piroplasms that are infective for ruminants in Central-Eastern Europe. Although bovine babesiosis and theileriosis was known to occur in Hungary, molecular evidence is provided here for the first time on the presence of Babesia and/or Theileria spp. of sheep, goats and cervids in Hungary.
Keywords: Babesia, Theileria, Tick, Cattle, Small ruminants, Cervids
Findings
Background
Piroplasms (Apicomplexa: Piroplasmida) are unicellular, tick-borne parasites that infect blood cells of vertebrates. In this category both the genus Babesia and the genus Theileria appear to be geographically widespread [1], affecting domesticated and game animal species, even humans, according to their lower or higher pathogenicity.
Piroplasms represent an unfolding field of research in parasitology, because Babesia and Theileria spp. are not as host-specific as previously thought [2] and the taxonomy of several species has been revised (e.g. [3]). Apart from this, a steadily increasing number of new species (e.g. Babesia vulpes: [4]) and genotypes [5] are being molecularly characterized, the latter frequently with unknown pathogenicity.
In this context, it is also evident that babesioses and/or theilerioses may show a changing epidemiological situation, including regionally disappearing and emerging species. In particular, it was reported that B. divergens is becoming extinct in north-east Hungary [6], and B. major and T. buffeli were newly identified in the same region [7]. Recently, during routine haematological examination, further cattle herds were recognized to harbour piroplasms in the same region of the country. Therefore the present study was initiated in order to identify any relevant agents and to uncover the range of Babesia and Theileria species in the local tick population. Because B. major, members of the T. orientalis complex and several other piroplasms of ruminants are transmitted by Haemaphysalis spp. in a worldwide context [8–10], the molecular investigation of potentially present Babesia and Theileria species focused on this hard tick genus.
Methods
Individual, EDTA-anticoagulated blood samples were collected by coccygeal venipuncture in a dairy cattle herd in which piroplasms had been detected microscopically during routine haematological examination of asymptomatic cattle. The herd consisted of 90 cows and their calves that are kept extensively (grazing pastures from April until November) in northern Hungary (Nógrád county, Pásztó, geographical coordinates: 47° 55′ 26.9″ N, 19° 42′ 21.4″ E). All cows were blood-sampled prior to the grazing period (April, 2015) and after two months of grazing (June, 2015). In addition, 73 ticks were collected from the pastures with the dragging-flagging method in the main tick season (April, 2015).
The DNA was extracted using QIAamp DNA Mini kit (Qiagen, Hilden, Germany) from all blood samples and ticks as described [7, 11]. The presence of piroplasms was investigated by a conventional PCR that amplifies an approx. 500 bp long part of the 18S rDNA gene of Babesia/Theileria spp. with the primers BJ1 (forward: 5′-GTC TTG TAA TTG GAA TGA TGG-3′) and BN2 (reverse: 5′-TAG TTT ATG GTT AGG ACT ACG-3′) [7]. This was followed by sequencing 20 PCR positive samples. Phylogenetic analysis was carried out according to the neighbor-joining method as reported [12]. The sequence was submitted to GenBank (KT725847). Exact confidence intervals (CI) for the prevalence rates were calculated at the level of 95 %.
To trace potential vectors of the T. orientalis complex and other piroplasms infecting ruminants in the region, a larger number of ticks (596 Haemaphysalis specimens, including 315 H. inermis, 259 H. concinna and 22 H. punctata) were also screened. These ticks had been collected from the vegetation by the dragging-flagging method from March to July in 2007, on 20 locations (pastures, forests, meadows) mainly in northern Hungary [13]. DNA extraction from pools of these ticks (Table 1) was done by the MagNA Pure LC total nucleic acid isolation kit (Roche Diagnostics, Rotkreuz, Switzerland), and prior to testing for piroplasms the amplifiable DNA contents of each tick pool were evaluated as reported [14]. These DNA samples were stored at −80 °C until molecular analysis (PCR and sequencing of 24 PCR positive samples) as described above. Sequences were submitted to GenBank (KT725848-54: Table 1).
Table 1.
Results of molecular analyses of 596 ticks collected in 2007 in Hungary
| Haemaphysalis species | Tick stage or sex | Number of ticks per pool | PCR positive/all pools | Results of sequencing (length, % identity, sample number) | Location in Hungarya | Reference sequence | Accession number of sequence in this study (name of isolate) |
|---|---|---|---|---|---|---|---|
| H. inermis | male | 5 | 7/24 | Babesia crassa (410 bp, 98.6 %, 4×) | B, P, S | KF791205 | KT725848 (Bcr-Hu1) |
| female | 5 | 12/40 | Babesia crassa (410 bp, 98.6 %, 6×) | B, P, S | KF791205 | KT725848 (Bcr-Hu1) | |
| H. concinna | male | 3 | 10/29 | Babesia sp. Kh-Hc222 (409 bp, 99.8 %, 1×) | P | KJ486568 | KT725849 (BKh-Hu1) |
| Babesia sp. Irk-Hc133 (409 bp, 100 %, 2×) | B, P, A, C | KJ486563 | KT725852 (BIrk-Hu1) | ||||
| Theileria capreoli (439 bp, 99.8 %, 2×) | B, P | AY726008 | KT725850 (Tc-Hu1) | ||||
| Theileria sp. ZS TO4b (440 bp, 100 %, 1×) | P | DQ520836 | KT725851 (TZSTO4-Hu1) | ||||
| female | 3 | 5/16 | Babesia motasi (403 bp, 99.3 %, 1×) | B | AY260179 | KT725853 (Bmo-Hu1) | |
| Babesia sp. Irk-Hc133 (409 bp, 100 %, 1×) | B, P, A, C | KJ486563 | KT725852 (BIrk-Hu1) | ||||
| Theileria sp. ZS TO4 (440 bp, 100 %, 1×) | P | DQ520836 | KT725851 (TZSTO4-Hu1) | ||||
| nymph | 10 | 7/7 | Babesia sp. Irk-Hc133 (409 bp, 100 %, 1×) | B, P, A, C | KJ486563 | KT725852 (BIrk-Hu1) | |
| Theileria capreoli (439 bp, 99.8 %, 1×) | B, P | AY726008 | KT725850 (Tc-Hu1) | ||||
| Theileria sp. ZS TO4 (440 bp, 100 %, 1×) | P | DQ520836 | KT725851 (TZSTO4-Hu1) | ||||
| larva | 52 | 0/1 | - | - | - | ||
| H. punctata | male | 5 | 1/5 | Theileria sp. OT3 (160 bp, 100 %, 1×) | P | KF470868 | KT725854 (TOT3-Hu1) |
| female | 3 | 1/3 | Theileria sp. OT3 (160 bp, 100 %, 1×) | V | KF470868 | KT725854 (TOT3-Hu1) | |
| nymph | 1 | 0/1 | - | - | - |
a Abbreviations: B Börzsöny Mountains, P Pilis Mountains, S Szekszárd Hills, V Visegrád Mountains, A Ásotthalom, C Cserhát Mountains
bNote that Theileria sp. ZS TO4 and T. capreoli are 99.5 % identical, but for comparability with relevant literature data and because of their separate clustering on the phylogentic tree (Fig. 1) both names are maintained in the table and in the text
Ethical approval
The animals were sampled as part of the regular veterinary care.
Results and discussion
Investigation of the current epidemiological situation in a cattle herd
Before grazing 52 of 90 (57.7 %, CI: 46.9–68.1 %), then (after two months) during grazing 51 of 90 (56.6 %, CI: 45.8–67.1 %) cows were PCR positive. Three cows became PCR negative, and two became PCR positive between the two samplings. All the remaining 49 animals had parasitaemias (detectable by PCR) which persisted during the evaluated period. Similarly high rates of infections were reported from other continents (e.g. [15]) and can be explained by long-term persistence of theileriae in the blood stream [16].
In all 20 blood samples sequencing revealed the same Theileria orientalis/buffeli genotype (length: 432 bp), with 100 % identity to T. orientalis isolates from Japan (in cattle: AB668373), China (in buffalo: HM538223), South-Africa (in buffalo: GU733374) and Australia (in cattle: AB520953). Based on GenBank data this genotype has not been previously reported in Europe. The sequence was submitted to GenBank (KT725847). This genotype differed with five nucleotides from the T. buffeli isolate reported recently from cattle in Hungary (KJ756505: [7]) and these two clustered separately with a high bootstrap support on the phylogenetic tree (Fig. 1).
Fig. 1.
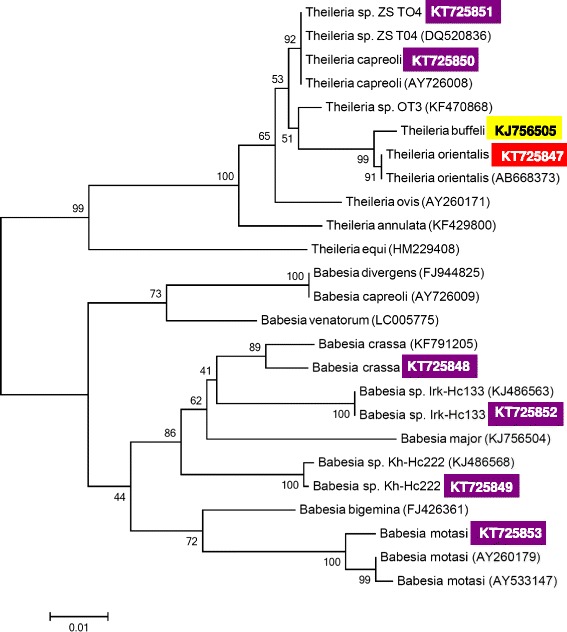
Phylogenetic comparison of partial 18S rDNA sequences of piroplasms identified in Hungary (inverse colour), with related sequences from the GenBank. Inverse purple colour indicates sequences amplified from ticks in the present study; yellow and red colour designate formerly and recently identified Theileria isolates from cattle, respectively. The names of piroplasm isolates obtained from ticks in the present study and corresponding lengths of sequences used for the phylogenetic tree are included in Table 1. The sequence of Theileria sp. OT3 obtained in this study was considerably shorter than those of other piroplasms, therefore it was excluded from the phylogenetic analysis. Branch lengths correlate to the number of substitutions inferred according to the scale shown. The isolation source and country/continent for reference sequences are the following: AY726008 (roe deer, Europe), KF470868 (sheep, China), DQ520836 (red deer, Poland), AY260171 (sheep, Sudan), KF429800 (cattle, Iran), AB668373 (cattle, Japan), HM229408 (horse, China), KJ486568 (Haemaphysalis concinna, Russia), KJ756504 (cattle, Hungary), KF791205 (H. parva, Turkey), KJ486563 (H. concinna, Russia), FJ944825 (cattle, France), AY726009 (roe deer, France), LC005775 (Ixodes persulcatus, Mongolia), FJ426361 (cattle, Brazil), AY260179 (sheep, Netherlands), AY533147 (sheep, Spain)
Ticks collected from the pastures (including all three Haemaphysalis spp.) were PCR negative (sample number per species not shown). Thus the local vector of T. orientalis could not be identified. Taken together, because in the relevant herd the main tick season in the spring apparently had minor influence on the prevalence of T. orientalis infection, it might have originated in previous years.
Molecular analysis of piroplasms in ticks collected in 2007
Seven piroplasm species or genotypes were shown to be present in these ticks (Table 1), all newly detected in Hungary. H. concinna carried five different piroplasms, as contrasted to one found in H. inermis, and another in H. punctata. The phylogenetic positions of these Babesia and Theileria spp. and relevant genotypes from GenBank are shown on Fig. 1.
H. inermis is the most abundant tick species of its genus in Hungary, and is regarded as an emerging species [13]. In ten pools of H. inermis (Table 1) a sequence was identified which had the highest (98.6 %) homology to (6 nucleotide difference from) two Turkish isolates of B. crassa: one from sheep (AY260177: [17]) and one from H. parva ticks (KF791205: [18]). The B. crassa genotype from H. inermis in the present study and the one previously reported from H. parva clustered together (Fig. 1). Babesia crassa has low pathogenicity in small ruminants [19] and its vector is/are most likely Haemaphysalis sp./spp., because (apart from H. parva as mentioned above) it was isolated from H. concinna in China (JX542614) and from H. sulcata in Turkey (KF034782) [20]. Until now this species or closely related genotypes were known to occur only in the Middle-East [17].
Another lowly pathogenic species, B. motasi was present in H. concinna. This piroplasm occurs in Northern, Western and Southern Europe, where it is transmitted by H. punctata [21]. To the best of our knowledge, B. motasi was not reported previously from H. concinna. The first Hungarian B. motasi genotype has 99.1–99.3 % sequence similarity to (i.e. three to four nucleotide difference from) isolates found in the Netherlands and in Spain, which differed from each other with two nucleotides, as reflected by the position of these three genotypes on the phylogenetic tree (Fig. 1).
Further piroplasm genotypes demonstrated from H. concinna or H. punctata in the present study (Table 1) are known to occur in countries close to Hungary (Theileria sp. ZS TO4 in red deer, H. concinna in Austria: [22]; T. capreoli in red deer in Poland: [23]), or in Southern Europe (Theileria sp. OT3 in sheep in Italy: [24]). Surprisingly, two Babesia genotypes found here in H. concinna were formerly reported in Far Eastern Russia (KJ486568 from Khabarovsk) and in East Siberia (KJ486563 from Irkutsk region), in both cases from H. concinna ticks [5]. In this way identical (or nearly identical) Babesia sequences were found in H. concinna ticks from different parts of Eurasia (Hungary, East Siberia, Far East), i.e. the relevant genotypes appear to be geographically very widespread. This may be related to the broad geographical range of H. concinna in Eurasia [25], and connectedness of its eastern and western habitats via longitudinal migration of birds [26]. Birds are known to be preferred hosts of this tick species [11, 25].
Taken together, in the present study several new genotypes of Babesia and Theileria spp. were identified. The high genetic variability of the 18S rDNA gene among piroplasms infecting ruminants is well known [5, 27]. Accordingly, Babesia and Theileria spp. detected in Haemaphysalis sp. ticks of the present study were found to exhibit pronounced heterogeneity in the amplified part of their 18S rDNA gene.
It was also shown here that different Babesia/Theileria genotypes or species are associated with different Haemaphysalis spp. in Central-Eastern Europe (i.e. none of the genovariants were shared between the ticks species analysed here). This is in line with the relatively narrow spectrum of competent vector tick species in the case of piroplasms [28].
Piroplasms with close phylogenetic relationships were reported to have similar host-associations as well [5]. In the present study H. inermis and H. punctata harboured only one Babesia and one Theileria sp. of sheep, respectively. In contrast to this, H. concinna carried piroplasms of wild cervids, as wells as of domestic small ruminants. The preferred hosts of H. concinna are red deer, roe deer, but it can also infest sheep and goats in Central-Eastern Europe [29]. Therefore it is likely that domestic and wild ruminants in the region are exposed to and infected with the detected piroplasms.
The high diversity of piroplasms of ruminants in ticks of the present study support previous literature data, because the number of Babesia and Theileria genotypes were shown to be highest in Haemaphysalis spp. as contrasted to other tick genera [5]. Interestingly, none of the nearly 600 Haemaphysalis sp. ticks, collected in 2007 and analysed here, harboured the three piroplasms (two genotypes of T. orientalis/buffeli, and B. major) of cattle newly detected in 2013–2015 in Hungary [7]. This may support that the latter piroplasms might have been introduced recently into the country. On the other hand, seven further Babesia/Theileria species or genotypes were identified in this study that were hitherto unknown to occur in the region.
Conclusions
This is the first simultaneous molecular investigation of piroplasms in representatives of all three Haemaphysalis spp., which occur in Central Europe. This report provides the first molecular evidence of piroplasms of small ruminants and cervids in Hungary, of B. motasi in Central-Eastern Europe, and of any B. crassa-like strain in Europe. The finding of B. crassa and B. motasi is also new in the tick species H. inermis and H. concinna, respectively. These data encourage scientists to broaden the scope of this study to other European countries.
Acknowledgements
Financial support was provided by OTKA 115854 and by the 9877–3/2015/FEKUT grant of the Hungarian Ministry of Human Resources. Laboratory work was partly performed with logistical support from the Center for Clinical Studies at the Vetsuisse Faculty of the University of Zurich.
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
SH initiated and organized the study, did most of the DNA extractions, wrote the manuscript. NT performed the PCRs. JK performed phylogenetic analysis. ZG and AM provided conditions of sampling for the study. SI collected blood samples. BF participated in DNA extractions. RF supervised parasitological work. RHL provided conditions for DNA extraction and supervised the study. All authors read and approved the manuscript.
Contributor Information
Sándor Hornok, Phone: 36-1-478-4187, Email: hornok.sandor@aotk.szie.hu.
Nóra Takács, Email: takacs.nora@aotk.szie.hu.
Jenő Kontschán, Email: jkontschan@gmail.com.
Zsolt György, Email: drgyorgyzs@gmail.com.
Attila Micsutka, Email: alpolgarmester@citromail.hu.
Serena Iceton, Email: serenaiceton@yahoo.co.uk.
Barbara Flaisz, Email: flaisz.barbara@aotk.szie.hu.
Róbert Farkas, Email: farkas.robert@aotk.szie.hu.
Regina Hofmann-Lehmann, Email: regina@hofmann-lehmann.com.
References
- 1.Homer MJ, Aguilar-Delfin I, Telford SR, 3rd, Krause PJ, Persing DH. Babesiosis. Clin Microbiol Rev. 2000;13:451–69. doi: 10.1128/CMR.13.3.451-469.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Criado A, Martinez J, Buling A, Barba JC, Merino S, Jefferies R, et al. New data on epizootiology and genetics of piroplasms based on sequences of small ribosomal subunit and cytochrome b genes. Vet Parasitol. 2006;142:238–47. doi: 10.1016/j.vetpar.2006.07.004. [DOI] [PubMed] [Google Scholar]
- 3.Mehlhorn H, Schein E. Redescription of Babesia equi Laveran, as 1901 as Theileria equi Mehlhorn, Schein 1998. Parasitol Res. 1998;1998(84):467–75. doi: 10.1007/s004360050431. [DOI] [PubMed] [Google Scholar]
- 4.Baneth G, Florin-Christensen M, Cardoso L, Schnittger L. Reclassification of Theileria annae as Babesia vulpes sp nov. Parasit Vector. 2015;8:207. doi: 10.1186/s13071-015-0830-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Rar VA, Epikhina TI, Suntsova OV, Kozlova IV, Lisak OV, Pukhovskaya NM, et al. Genetic variability of Babesia parasites in Haemaphysalis spp. and Ixodes persulcatus ticks in the Baikal region and Far East of Russia. Infect Genet Evol. 2014;28:270–5. doi: 10.1016/j.meegid.2014.10.010. [DOI] [PubMed] [Google Scholar]
- 6.Hornok S, Edelhofer R, Szotáczky I, Hajtós I. Babesia divergens becoming extinct in cattle of Northeast Hungary: new data on the past and present situation. Acta Vet Hung. 2006;54:493–501. doi: 10.1556/AVet.54.2006.4.7. [DOI] [PubMed] [Google Scholar]
- 7.Hornok S, Mester A, Takács N, Fernández de Mera IG, de la Fuente J, Farkas R. Re-emergence of bovine piroplasmosis in Hungary: has the etiological role of Babesia divergens been taken over by B. major and Theileria buffeli? Parasit Vectors. 2014;7:434. doi: 10.1186/1756-3305-7-434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Alani AJ, Herbert IV. The morphometrics of Babesia motasi (Wales) and its transmission by Haemaphysalis punctata (Canestrini and Fanzago 1877) to sheep. Vet Parasitol. 1988;30:87–95. doi: 10.1016/0304-4017(88)90155-0. [DOI] [PubMed] [Google Scholar]
- 9.Fujisaki K. A review of the taxonomy of Theileria sergenti/buffeli/orientalis group parasites in cattle. J Protozool Res. 1991;2:87–96. [Google Scholar]
- 10.Yin H, Lu W, Luo J, Zhang Q, Lu W, Dou H. Experiments on the transmission of Babesia major and Babesia bigemina by Haemaphysalis punctata. Vet Parasitol. 1996;67:89–98. doi: 10.1016/S0304-4017(96)01022-9. [DOI] [PubMed] [Google Scholar]
- 11.Hornok S, Kováts D, Csörgő T, Meli ML, Gönczi E, Hadnagy Z, et al. Birds as potential reservoirs of tick-borne pathogens: first evidence of bacteraemia with Rickettsia helvetica. Parasit Vectors. 2014;7:128. doi: 10.1186/1756-3305-7-128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hornok S, Estók P, Kováts D, Flaisz B, Takács N, Szőke K, et al. Screening of bat faeces for arthropod-borne apicomplexan protozoa: Babesia canis and Besnoitia besnoiti-like sequences from Chiroptera. Parasit Vectors. 2015;8:441. doi: 10.1186/s13071-015-1052-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hornok S, Farkas R. Influence of biotope on the distribution and peak activity of questing ixodid ticks in Hungary. Med Vet Entomol. 2009;23:41–6. doi: 10.1111/j.1365-2915.2008.00768.x. [DOI] [PubMed] [Google Scholar]
- 14.Boretti FS, Perreten A, Meli ML, Cattori V, Willi B, Wengi N, et al. Molecular Investigations of Rickettsia helvetica infection in dogs, foxes, humans, and Ixodes ticks. Appl Environ Microbiol. 2009;75:3230–7. doi: 10.1128/AEM.00220-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Eamens GJ, Bailey G, Gonsalves JR, Jenkins C. Distribution and temporal prevalence of Theileria orientalis major piroplasm surface protein types in eastern Australian cattle herds. Aust Vet J. 2013;91:332–40. doi: 10.1111/avj.12078. [DOI] [PubMed] [Google Scholar]
- 16.Mans BJ, Pienaar R, Latif AA. A review of Theileria diagnostics and epidemiology. Int J Parasitol Parasites Wildl. 2015;4:104–18. doi: 10.1016/j.ijppaw.2014.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Schnittger L, Yin H, Gubbels MJ, Beyer D, Niemann S, Jongejan F, et al. Phylogeny of sheep and goat Theileria and Babesia parasites. Parasitol Res. 2003;91:398–406. doi: 10.1007/s00436-003-0979-2. [DOI] [PubMed] [Google Scholar]
- 18.Orkun Ö, Karaer Z, Çakmak A, Nalbantoğlu S. Identification of tick-borne pathogens in ticks feeding on humans in Turkey. PLoS Negl Trop Dis. 2014;8 doi: 10.1371/journal.pntd.0003067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hashemi-Fesharki R, Uilenberg G. Babesia crassa n.sp. (Sporozoa, Babesiidae) of domestic sheep in Iran. Vet Q. 1981;3:1–8. doi: 10.1080/01652176.1981.9693787. [DOI] [PubMed] [Google Scholar]
- 20.Aktas M. A survey of ixodid tick species and molecular identification of tick-borne pathogens. Vet Parasitol. 2014;200:276–83. doi: 10.1016/j.vetpar.2013.12.008. [DOI] [PubMed] [Google Scholar]
- 21.Uilenberg G, Rombach MC, Perié NM, Zwart D. Blood parasites of sheep in the Netherlands. II. Babesia motasi (Sporozoa, Babesiidae) Vet Q. 1980;2:3–14. doi: 10.1080/01652176.1980.9693752. [DOI] [PubMed] [Google Scholar]
- 22.Fuehrer HP, Biro N, Harl J, Worliczek HL, Beiglböck C, Farkas R, et al. Molecular detection of Theileria sp. ZS TO4 in red deer (Cervus elaphus) and questing Haemaphysalis concinna ticks in Eastern Austria. Vet Parasitol. 2013;197:653–7. doi: 10.1016/j.vetpar.2013.07.005. [DOI] [PubMed] [Google Scholar]
- 23.Sawczuk M, Maciejewska A, Skotarczak B. Identification and molecular detection of Theileria sp. infecting red deer (Cervus elaphus) in north-western Poland. Eur J Wildl Res. 2008;54:225–30. doi: 10.1007/s10344-007-0133-z. [DOI] [Google Scholar]
- 24.Giangaspero A, Marangi M, Papini R, Paoletti B, Wijnveld M, Jongejan F. Theileria sp. OT3 and other tick-borne pathogens in sheep and ticks in Italy: molecular characterization and phylogeny. Ticks Tick Borne Dis. 2015;6:75–83. doi: 10.1016/j.ttbdis.2014.09.007. [DOI] [PubMed] [Google Scholar]
- 25.Nosek J. The ecology, bionomics and behaviour of Haemaphysalis (Haemaphysalis) concinna tick. Z Parasitenkd. 1971;36:233–41. doi: 10.1007/BF00348561. [DOI] [PubMed] [Google Scholar]
- 26.Boere G, Dodman T. The flyway approach to the conservation and wise use of waterbirds and wetlands: a training kit. Module 1: Understanding the flyway approach to conservation, 2010. Available from: http://www.eaaflyway.net/documents/resources/wow2-Module1.pdf. Accessed: 18/10/2015.
- 27.Chae JS, Lee JM, Kwon OD, Holman PJ, Waghela SD, Wagner GG. Nucleotide sequence heterogeneity in the small subunit ribosomal RNA gene variable (V4) region among and within geographic isolates of Theileria from cattle, elk and white-tailed deer. Vet Parasitol. 1998;75:41–52. doi: 10.1016/S0304-4017(97)00183-0. [DOI] [PubMed] [Google Scholar]
- 28.Jongejan F, Uilenberg G. The global importance of ticks. Parasitology. 2004;129(Suppl):S3–14. doi: 10.1017/s0031182004005967. [DOI] [PubMed] [Google Scholar]
- 29.Hornok S, Horváth G, Jongejan F, Farkas R. Ixodid ticks on ruminants, with on-host initiated moulting (apolysis) of Ixodes, Haemaphysalis and Dermacentor larvae. Vet Parasitol. 2012;187:350–3. doi: 10.1016/j.vetpar.2011.12.012. [DOI] [PubMed] [Google Scholar]