

Abstract
Proinflammatory cytokines have been implicated in alcohol-induced neurodegeneration, but the role of the neuroimmune system in alcohol related behaviors has only recently come to the forefront. Herein, the effects of binge-like drinking on IL-1β mRNA and immunoreactivity within the amygdala were measured following the “drinking in the dark” (DID) paradigm, a model of binge-like ethanol drinking in C57BL/6J mice. Moreover, the role of IL-1 receptor signaling in the amygdala on ethanol consumption was assessed. Results indicated that a history of binge-like ethanol drinking promoted a significant increase of IL-1β mRNA expression within the amygdala, and immunohistochemistry analyses revealed that the basolateral amygdala (BLA), but not central amygdala (CeA), exhibited significantly increased IL-1β immunoreactivity. Fluoro-Jade® C labeling indicated that multiple cycles of the DID paradigm were not sufficient to elicit neuronal death. Bilateral infusions of IL-1 receptor antagonist (IL-1Ra) reduced ethanol consumption when infused into the BLA but not the CeA. These observations were specific to ethanol drinking as the IL-1Ra did not alter either sucrose drinking or open-field locomotor activity. The current findings highlight a specific role for IL-1 receptor signaling in modulating binge-like ethanol consumption and indicate that proinflammatory cytokines can be induced prior to dependence or any evidence of neuronal cell death. These findings provide a framework in which to understand how neuroimmune adaptations may alter ethanol consumption and therein contributing to alcohol abuse.
1. Introduction
Binge drinking is a prevalent problem within the US, and repeated bouts of high blood alcohol concentrations can fundamentally change normal neurobiological functions (CDC, 2013; Gunzerath et al., 2011). These neuroplastic changes are thought to be one of the underlying mechanisms that lead to alcohol use disorders (AUDs) (Der-Avakian and Markou, 2012). One such system altered by alcohol abuse that has recently emerged as an area of interest is the neuroimmune system (Crews et al., 2011; Hutchinson and Watkins, 2014). Studies of brains from people with a history of alcohol abuse show increased microglial activation and proinflammatory cytokines such as IL-1β (He and Crews, 2008; Zou and Crews, 2012). Pre-clinical studies concurring with this phenomenon have mainly focused on characterizing alcohol-induced neuroimmune responses in relation to cellular damage (Crews and Vetreno, 2014; Marshall et al., 2013) or the molecular mechanisms that underlie the initial immune response (Fernandez-Lizarbe et al., 2013). In particular, IL-1β has been shown to be upregulated in the CNS after various AUD models (Lippai et al., 2013; Qin et al., 2008). However, the contributions of neuroimmune responses to the modulation of ethanol consumption have been relatively ignored, especially in the transition between alcohol abuse and alcohol dependence. This study utilizes the “drinking-in-the-dark” paradigm (DID) because it is uniquely suited to study neuroadaptations that occur during the transitory window between excessive ethanol consumption prior to dependence in rodents (Rhodes et al., 2005; Thiele and Navarro, 2014).
Studies focused on the alcohol-related behavioral implications of the immune system have shown that modulation of the system alters ethanol consumption. In general, immune activating compounds increase ethanol consumption; whereas, anti-inflammatory agents reduce ethanol intake (Bell et al., 2013; Blednov et al., 2011). For example, peripheral administration of lipopolysaccharide, an inflammatory agent, resulted in prolonged increased voluntary ethanol consumption (Blednov et al., 2011), but the specific molecular mechanisms underlying these responses are still elusive as lipopolysaccharide can alter a bevy of cytokines and chemokines (Qin et al., 2007; Turrin et al., 2001). Moreover, it is unclear if reduced consumption is caused by peripheral or CNS actions of immune modulation. The present study specifically assessed the role of IL-1 receptor signaling by examining IL-1β, a pro-inflammatory cytokine, changes induced by binge-like ethanol drinking and then blocking IL-1 receptor signaling by intracranial site-specific infusions of IL-1 receptor antagonist (IL-1Ra).
Cytokines are more than immunomodulators and have been increasingly implicated in CNS actions including complex behaviors like addiction (Coller and Hutchinson, 2012). IL-1β has come to the forefront as a key player in other behavioral phenotypes underlying AUDs including depression and pain disorders (Egli et al., 2012; Markou et al., 1998). Its signaling has been implicated in a variety of neuroplastic events in other maladaptations including anxiety and emotional disorders (Jones et al., 2015; Wohleb et al., 2014). The specific contribution of IL-1β within the CNS to AUDs has not been determined, but modulation of the IL-1 system through peripheral IL-1Ra injections has been shown to decrease alcohol-induced sedation, motor impairment, as well as ethanol consumption (Lippai et al., 2013; Wu et al., 2011). Moreover, genetic polymorphisms of IL-1 have been found in alcohol-dependent patients compared with healthy controls (Liu et al., 2009), but deletion of the gene that encodes IL-1Ra reduced ethanol preference and consumption in transgenic mice (Blednov et al., 2012) . While these studies suggest a link between IL-1 signaling and ethanol consumption, the conflicting findings and whether these effects are from peripheral or CNS contributions of IL-1 receptor signaling are not known.
Although the neurocircuitry that modulates ethanol consumption is complex, the present study focuses on the role of IL-1 receptor signaling within the amygdala. Amygdalar IL-1 receptor signaling is associated with increased anxiety, which is believed to be a potential underlying factor in ethanol consumption and craving (Chiu et al., 2014; Fox et al., 2007). Furthermore, neuropeptides like corticotrophin releasing factor (CRF) and neuropeptide Y (NPY), both of which modulate stress and anxiety, can modulate IL-1β concentrations (Ferreira et al., 2010; Gilpin et al., 2015; Kageyama et al., 2010; Rivest and Rivier, 1994). Our laboratory and others have previously shown that manipulating both CRF and NPY signaling within the amygdala and extended amygdala modulates binge-like ethanol consumption in mice (Correia et al., 2015; Lowery-Gionta et al., 2012; Pleil et al., 2015). Together these findings indicate a potential interplay between IL-1 signaling and ethanol consumption within the amygdala.
Given the link between IL-1 receptor signaling and CRF activity, and the known role of CRF in modulating binge-like ethanol drinking, we hypothesized that IL-1 receptor activity in the amygdala also modulates binge-like drinking and that it is not solely associated with alcohol-induced neurodegeneration. The current experiments tests these hypotheses by determining: a) the effects of binge-like drinking on IL-1β immunoreactivity and mRNA expression, b) whether ethanol-induced neuroimmune activity occurs independent of neurodegeneration, and finally c) the effects of blocking of IL-1 receptor signaling on ethanol consumption.
2. Methods
2.1 Animals
Male C57BL /6J 6-8 weeks old mice (Jackson Laboratories; Bar Harbor, ME) were individually housed in a reversed 12:12 hour light:dark cycle vivarium maintained at 22°C. During experiments, animals had ad libitum access to Prolab® RMH 3000 (Purina LabDiet®; St. Louis, MO) or Teklad Diet® 2920X (Harlan Laboratories Inc.; Indianapolis, IN) and water unless otherwise indicated. All animals were given at least a week to acclimate to the environment before experimentation. The procedures used in this study were all approved by the University of North Carolina Institutional Animal Care and Use Committee and followed the Guidelines for the Care and Use of Laboratory Animals.
2.2 “Drinking in the Dark” Procedures
Binge drinking was modeled using a 4 day DID paradigm as previously described (Rhodes et al., 2005). Briefly, home cage water bottles were removed, and mice were allotted two hour access to a single bottle of 20% (v/v) ethanol solution or a control solution of either 3% (w/v) sucrose or water three hours into the dark cycle. On the fourth day, also called the test day, consumption (g/kg) was measured for four hours. Each DID cycle was separated by three days for mice undergoing multiple 4-day DID cycles.
Tail blood samples (≈60μL) were taken immediately following the DID procedure to determine blood ethanol concentration (BEC) except in the qRT-PCR experiment. For mRNA studies, trunk blood samples were taken in the group immediately euthanized following the DID cycle rather than tail vein samples. The groups euthanized 1 day or 7 days after DID did not have tail blood samples taken to avoid any restraint stress associated with tail blood collection, which has previously been shown to alter IL-1β mRNA expression (Minami et al., 1991). Blood samples collected were centrifuged to obtain serum samples and run in duplicate with the AM1 Alcohol Analyzer (Analox, London, UK) for BEC determination (mg/dL).
2.3 qRT-PCR
Mice used in PCR studies experienced one 4-day DID cycle receiving either ethanol or water. Ethanol and water animals were further divided by euthanasia timepoint (0, 1 or 7 days post DID; Ethanol n= 7-10/subgroup; H2O n= 6-10/subgroup). Following rapid decapitation, brains were extracted, flash frozen in isopentane, and stored at −80°C. Cylindrical punches of the amygdala (1mm x 1mm) were harvested and immediately submerged in RNALater Ice® (Ambion; Carlsbad, CA), according to manufacturer instructions. Amygdalar tissue punches were manually homogenized in Tri-Reagent (1ml/50-100mg tissue, Molecular Research Center, Inc.; Cincinnati, OH) with a glass dounce tissue grinder and briefly sonicated. RNA was separated using 1-bromo-3-chloropropane (Molecular Research, Inc.) and purified using isopropanol. AdvantageTM RT-for-PCR kit (Clontech; Mountain View, CA) was used to create cDNA samples via reverse transcription. Samples of cDNA were analyzed using the UNC Animal Clinical Chemistry and Gene Expression Laboratories similar to protocols previously reported (Jones et al., 2015; Kim et al., 2002). The nucleotide sequences of the PCR primers and fluorogenic probes were developed by UNC Animal Clinical Chemistry and Gene Expression Laboratories. The nucleotide sequences of the PCR primers and fluorogenic probes used for the IL-1β and β-actin genes are as follows. : IL-1β forward: 5’-AAT TTC TTG CAG CCG GAG CA-3’, reverse: 5’-AGG CGG AGG AAG TAT TCT TC-3’, probe: 5’-FCC AGC AAC CTC AGC CGG TTC TGA Q-3’; β-actin forward: 5’-CTG CCT GAC GGC CAG GTC-3’, reverse: 5’-CAA GAA GGA AGG CTG GAA AAG A-3’, probe: 5’-FCA CTA TTG GCA ACG AGC GGT TCC GQ-3’. . Measurements were normalized to water groups as previously described and are expressed as fold change (Kim et al., 2002) .
2.4 Tissue Processing
Mice used in all other experiments were overdosed with a 0.15mL intraperitoneal (ip) injection of an anesthetic cocktail of ketamine (66.67mg/mL) and xylazine (6.67mg/mL) in 0.9% saline. Animals were then transcardially perfused with 0.1M phosphate buffer saline (PBS; pH=7.4) and 4% paraformaldehyde in PBS (pH=7.4). Extracted brains were postfixed in paraformaldehyde for 24 hours and sectioned coronally at 40μm using a vibratome (Leica VT1000S; Wetzlar, Germany). Sections were collected in a 1:4 series for both immunohistochemistry (IHC) and Fluoro-Jade C (FJC) processing and stored in cryopreserve at −20°C.
2.5 Immunohistochemistry & Immunoreactivity Quantification
Animals used for IHC analysis underwent one or three, 4-day cycles of DID receiving either ethanol or sucrose or 3 cycles with water and were euthanized immediately at the end of their final cycle (Ethanol n= 9/subgroup; Sucrose n= 9-10/subgroup; H2O n= 9). Every fourth section collected was used for IL-1β immunoreactivity detection. Free floating sections were rinsed in 0.1M PBS before endogenous peroxidases were quenched with 0.6% H2O2. After serial PBS washes, nonspecific binding was blocked using rabbit serum (PBS/0.1% triton-X/3% rabbit serum; Vector Laboratories; Burlingame, CA). Sections were then incubated in goat IL-1β primary antibody (1:1000; R&D Systems; Minneapolis, MN) for 48 hours at 4°C. After serial washes in rabbit serum to remove the primary, incubation in biotinylated rabbit anti-goat secondary antibody, avidin-biotin-peroxidase complex (ABC elite kit, Vector Labs), and the chromagen, 3,3’-diaminobenzidine tetrahydrochloride (Polysciences; Warrington, PA) were used for detection of IL-1β. Sections were then mounted onto glass slides and coverslipped with SHUR/Mount™ (Triangle Biomedical Sciences; Durham, NC).
Images of slides were taken with a Zeiss Axio Zoom V16 macroscope (Jena, Germany) installed on an HP Z820 Workstation at 100x magnification. Slide images were coded to ensure that the experimenter was blind to treatment conditions during quantification. The basolateral amygdala (BLA) and the central nucleus of the amygdala (CeA) were separately traced on sections between Bregma −0.70mm and −2.06mm (Paxinos and Franklin, 2004). IL-1β immunoreactivity was measured using Zen Pro 2012. Immunopositive pixels were determined by optical density from experimenter determined threshold so that immunoreactivity is expressed as percent area (immunoreactive positive area/total area of region of interest).
2.6 Fluoro-Jade C & Quantification
FJC is a method of assessing neurodegeneration comparable to amino-cupric silver staining (Schmued et al., 2005). This staining was performed on animals following one or three, 4-day DID cycles and were euthanized immediately following the final DID cycle (Ethanol n= 8-9/subgroup; Sucrose n=6-7/subgroup; H2O n=10). FJC processing was performed in accordance with manufacture instructions (Millipore, Billerica, MA) similar to previously published methods (Liput et al., 2013; Qin and Crews, 2012b). Briefly, every fourth section was mounted on Superfrost-Plus slides (Fisher Scientific, Pittsburgh, PA). After drying overnight in a 37°C incubator, sections were rehydrated with the following steps: 5 min in 1% sodium hydroxide/ 80% ethanol; 2 min in 70% ethanol; 2 min ddH2O. After rehydration, slides were incubated in 0.06% potassium permanganate for 10 min while gently shaking and then rinsed in ddH2O for 2 min. Sections were then stained with 0.001% (w/v) FJC in 0.1% (v/v) acetic acid for 20 min. Finally residual FJC was removed by rinses (3 × 1 min; ddH2O). Slides were allowed to dry overnight and coverslipped using SHUR/Mount™. FJC+ cells within the amygdalar subregions (Bregma −0.70mm and −2.06mm) were quantified at 200x using an Olympus BX-51 microscope (Center Valley, PA) and expressed as cell/section.
2.7 Surgeries & IL-1Ra Intervention
Animals used to manipulate IL-1 receptor signaling underwent the 4-day DID procedure prior to surgery. For cannulae placement surgery, animals were anesthetized using a cocktail of xylazine (10mg/kg) and ketamine (100mg/kg) delivered ip (1.5mL/kg). Bilateral 26-G guide cannulae (Plastics One; Roanoke, VA) were aimed at the BLA (AP:−1.22, ML:±3.01, DV: −4.75) or the CeA (AP:−1.06, ML:±2.50, DV: −4.64) using an Angle IITM Stereotax (Leica Instruments, Buffalo Grove, IL) (Paxinos and Franklin, 2004). After a week of recovery, animals were subjected to the DID procedure again. On the test day, before alcohol access, 0.9μg of IL-1Ra (GenScript; Piscataway, NJ) dissolved in 0.9% saline (3mg/mL) or saline alone were infused at a rate of 0.15μL/min for two minutes using a Hamilton syringe (Reno, NV) attached to a Harvard Apparatus PHD 2000 infusion pump (Holliston, MS) similar to previous studies (Lowery-Gionta et al., 2012). Injectors were left in the guide cannulae for an additional 3 minutes to allow proper diffusion. All studies involving IL-1Ra used Latin-Square designs so that each mouse was dosed with both IL-1Ra and saline in a counterbalanced order for ethanol (BLA n=12; CeA n= 14) or sucrose (BLA n=13). At the conclusion of all behavioral analyses injection placements were histologically verified using an identical volume of Alcian blue dye (0.3μL/injection site) as the IL-1Ra experiments.
2.8 Open Field Tests
After a week of recovery, mice used in the sucrose test were then split into two groups receiving microinfusions in the BLA, as previously described, of either IL-1Ra (n=7) or saline (n=6) and subjected to open-field locomotor activity tests to assess the effects of IL-1Ra on general locomotor activity as well as its anxiolytic properties (Prut and Belzung, 2003). Following microinfusions, mice were placed into an open-field arena identical to previous descriptions (Cox et al., 2013; Fee et al., 2004). Briefly, animal movement was tracked using the VersaMax® software program (AccuScan Instruments, Inc., Columbus, OH) over a two hour period with fifteen minute bin outputs using VersaDat® Version 4.00 (AccuScan Instruments, Inc.). Total distance traveled was measured to assess the effect of IL-Ra on locomotor activity; whereas center distance and center time were recorded to determine the anxiolytic effects of IL-1Ra (Prut and Belzung, 2003).
2.9 Statistical Analysis
Prism Version 5.0 (GraphPad Software, Inc. La Jolla, Ca) was used to analyze and graph all data reported herein. Two-way analyses of variance (ANOVAs) were used to determine the effect of treatment and time for IL-1β mRNA expression and behaviors within the open-field test (OFT). However, one-way ANOVAs were used to assess data collected from both the BLA and the CeA for immunoreactivity of IL-1β and counts of FJC+ cells as well as to compare ethanol consumption in different cohorts of animals. Ethanol consumption, BECs, and sucrose consumption following IL-Ra manipulation were compared using a two-way (drug x order of drug presentation) repeated measures (RM) ANOVA. Bonferroni posthoc tests or t-tests were only conducted if a significant interaction or main effect of treatment was observed. All data are reported as the mean ± standard error of the mean and considered significant if p<0.05, two-tailed.
3. Results
3.1 Binge-Like Ethanol Consumption Increased IL-1β mRNA expression
Animals used in the qRT-PCR experiment had a grand mean ethanol consumption of 4.8±0.16g/kg. The BECs from the group immediately euthanized averaged 106.5±16.5mg/dL. A one-way ANOVA indicated no significant differences in consumption between animals used at different time-points ([F(2,21)=0.07, p=0.93]; Table 1). Ethanol exposure resulted in increased IL-1β mRNA in the amygdala by approximately 6-fold compared with the water drinking control group. A two-way ANOVA revealed a main effect of binge-like ethanol versus water consumption history [F(1,43)=19.33, p<0.001] but no effect of time of abstinence after DID [F(2,43)=0.00, p=0.99] or an interaction [F(2,43)=0.01, p=0.99]. Collapsing across time, a posthoc t-test indicated a significant increase of IL-1β mRNA in mice that consumed ethanol compared with water [t(48)=4.21, p<0.001] (Figure 1). Importantly, a two-way ANOVA showed no differences in the crossing point of the housekeeping gene, β-actin, between the water group and animals that received ethanol (Table 2).These indicate that a short history of binge-like ethanol drinking promotes increased IL-1β expression that persists up to a week following binge-like consumption.
Table 1.
ETHANOL INTAKE AND BLOOD ETHANOL LEVELS FROM THE mRNA, IMMUNOHISTOCHEMISTRY, AND FLUOROADE C EXPERIMENTS
| Experiment | Time | Consumption (g/kg/4hr) | BEC (mg/dL) |
|---|---|---|---|
| mRNA Expression | 0d after 1-DID cycle | 4.8±0.0.3 | 106.5±16.5 |
| 1d after 1-DID cycle | 4.7±0.2 | N/A | |
| 7d after 1-DID cycle | 4.9±0.3 | N/A | |
| Immunohistochemistry | 1-DID cycle | 4.2±0.4 | 54.10±13.3 |
| 3-DID Cycles | 4.7±0.4 | 91.33±16.3 | |
| Fluoro-Jade C | 1-DID cycle | 4.6±0.3 | 67.3±15.7 |
| 3-DID Cycles | 4.5±0.3 | 71.9±16.3 |
Figure 1.
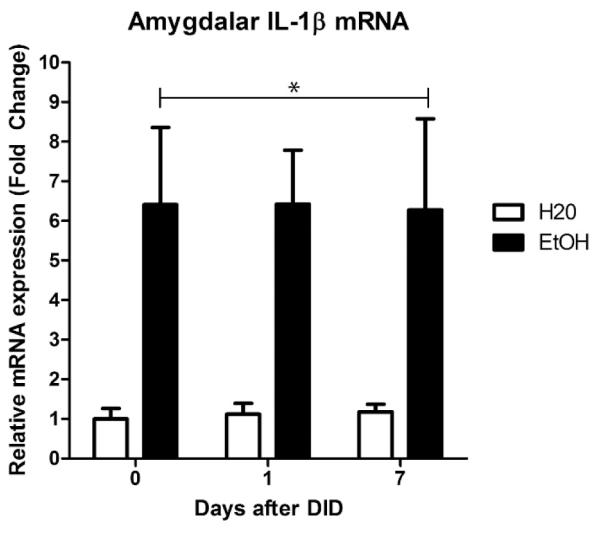
One cycle of DID induced increased mRNA expression of IL-1β when compared to the group that only received water. This effect persisted even a week into abstinence (*p<0.05 compared to water group). All data are presented as mean ± SEM.
Table 2.
CROSSING THRESHOLDS FOR qRT-PCR STUDY
| Group | β-Actin (Ct) | IL-1β (Ct) | |
|---|---|---|---|
| Water: | 0D after 1-DID cycle | 23.19 ± 0.3 | 37.35 ± 0.6 |
| 1d after 1-DID cycle | 22.19 ± 0.4 | 37.05 ± 0.3 | |
| 7d after 1-DID cycle | 23.58 ± 0.4 | 37.07 ± 0.5 | |
| Ethanol | 0D after 1-DID cycle | 22.62 ± 0.3 | 34.27 ± 0.4 |
| 1d after 1-DID cycle | 22.36 ± 0.2 | 34.77 ± 0.4 | |
| 7d after 1-DID cycle | 22.91 ± 0.4 | 35.49 ± 0.3 | |
3.2 Binge-Like Ethanol Consumption Increased IL-1β Immunoreactivity
Animals used in the IL-1β immunoreactivity experiment had a grand mean ethanol consumption of 4.5±0.35g/kg and BEC of 70.4±11.4mg/dL. T-tests indicated no significant differences in ethanol consumption [t(17) =0.02, p=0.98] or BECs [t(17) =1.57, p=0.14] between animals that had one or three 4-day DID cycles (Table 1). Photomicrographs indicated IL-1β positive cells were apparent in the amygdala of both ethanol and control animals as previously shown in rodents (Figure 2) (Ericsson et al., 1995; Yabuuchi et al., 1994). Binge-like ethanol drinking resulted in an increased IL-1β immunoreactivity compared with the water drinking control group in the BLA (Figure 2A). A one-way ANOVA revealed a main effect of group [F(2,23)=4.25, p=0.03]. Posthoc Bonferroni tests indicated a significant increase of IL-1β immunoreactivity in mice that experienced 3 cycles of DID compared with water drinking control mice (p<0.05). In the CeA, a one-way ANOVA did not reach statistical significance ([F(2,23)=0.95, p=0.40]; Figure 2C). These data indicate that a history of binge-like ethanol drinking promotes increased IL-1β immunoreactivity specifically in the BLA and not the CeA. Importantly, no difference was indicated by one-way ANOVAs between animals drinking water versus sucrose in the BLA [F(2,24)=0.05, p=0.95] or the CeA [F(2,24)=0.36, p=0.70] (Figure 2B,D), suggesting that increased IL-1β was specific to ethanol consumption.
Figure 2.

Ethanol induced increased IL-1β immunoreactivity within the BLA after 3 cycles compared with the water control (A); however, no statistically significant difference was seen within the CEA between water and ethanol (C) or between water and sucrose were seen in either the BLA (B) or CEA (D; *p<0.05 compared to water group). Representative photomicrographs of the basolateral amygdala of mice exposed to water (E), 1 ethanol DID cycle (F), and 3 ethanol DID cycles (G) show an upregulation of IL-1β in the BLA. Scale bar in panel G= 20μm; dashed lines delineate the external capsule surrounding the basolateral amygdala. All data are presented as mean ± SEM.
3.3 Binge-Like Ethanol Drinking Elicits Neuroimmune Responses Independent of Neurodegeneration
Neuroimmune responses are often elicited by neurodegeneration, so FJC was used to assess potential ethanol-induced damage stemming from binge-like ethanol intake. Animals used to assess neurodegeneration had a grand mean ethanol consumption of 4.5±0.28g/kg and BEC of 71.9±16.25mg/dL (Table 1). Very few FJC+ cells were observed in the amygdala of animals that received water (Figure 3E) or following 1 or 3, 4-day cycles of binge-like ethanol drinking as evident in the photomicrographs (Figure 3F,G); however, an LPS treated positive control was run in conjunction to ensure the FJC protocol was successful (Figure 3H). One-way ANOVAs indicated no significant effect of ethanol or sucrose drinking compare with water consumption in either the BLA [F(2,24)=0.22, p=0.81; F(2,20)=0.88, p=0.43] (Figure 3A,B) or the CeA [F(2,24)=0.79, p=0.46; F(2,20)=1.31, p=0.29] (Figure 3C, D).
Figure 3.

Neither ethanol (A,C) nor sucrose (B,D) induced any changes in FJC+ cells. Representative photomicrographs of the FJC stain for mice that received water (E) or underwent 1 (F) or 3 (G) DID cycles of or ethanol show very little FJC+ cells. An LPS treated positive control can be seen in panel H. Scale bar in panel H= 20μm; dashed lines delineate the external capsule surrounding the basolateral amygdala. All data are presented as mean ± SEM.
3.4 IL-1Ra Infusion in BLA Reduced Binge-Like Ethanol but not Sucrose Consumption
There was an 87% success rate in bilateral cannula placements. Six animals whose cannulas were not within the BLA or CeA were removed from analyses. With respect to the BLA, a two-way RM ANOVA (drug x order of drug presentation) on ethanol consumption indicated a main effect of drug treatment [F(1,11)=12.30, p=0.005] but no main effect of drug order [F(1,11)<0.01, p=0.97] or interaction between order and treatment [F(1,11)=4.64, p=0.06] (Figure 4A). Likewise, a two-way RM ANOVA (drug x order of drug presentation) of BECs indicated a main effect of drug treatment [F(1,11)=15.28, p=0.002] but no main effect of drug order [F(1,11)=0.42, p=0.53] or interaction between order and treatment [F(1,11)=0.51, p=0.49] (Figure 4B). However, a two-way RM ANOVA of binge-like ethanol consumption in the CeA experiment revealed no interaction [F(1,13)=1.32, p=0.27] or main effects of IL-1Ra [F(1,13)=0.63, p=0.44] or order of drug treatment [F(1,13)=0.07, p=0.80] (Figure 4C). No effect of IL-1Ra administration in the CeA [F(1,13)=1.03, p=0.33], order of treatment [F(1,13)=1.02, p=0.33], or interaction [F(1,13)=2.79, p=0.12] was determined using a two-way RM ANOVA examining BECs (Figure 4D). The effects of manipulating IL-1 receptor signaling via IL-1Ra in the BLA was specific to ethanol consumption as a two-way RM ANOVA comparing groups that consumed sucrose did not indicate any significant effect of the IL1Ra [F(1,12)=2.12, p=0.17], drug presentation order [F(1,12)=2.33, p=0.15], or any interaction [F(1,12)=0.18, p=0.68] (Figure 5A).
Figure 4.

Graphs represent the collapsed data of the Latin-Square design. IL-1Ra pretreatment in the BLA resulted in reduced ethanol consumption (A) during the DID session and resulted in lower BECs (B) compared with saline. No significant effect of IL-1Ra manipulation within the CEA was observed in either consummatory data (C) or in BECs (D). Approximate injection sites are shown in panel E. Grey circles represent cannulae placement within the BLA while black circles represent those in the CeA. All data are presented as mean ± SEM.
Figure 5.

Collapsing across drug presentation groups, IL-1Ra administration in the BLA did not result in significant differences in sucrose consumption (A).Additionally IL-1Ra treatment in the BLA did not significantly affect locomotor activity (B) or any measures of anxiety [Center Time (C); Center Distance (D)]. Approximate injection sites are represented by grey circles in panel E. All data are presented as mean ± SEM.
3.5 IL-1Ra Infusion in BLA had No Effects in Open-Field Test
Reduced ethanol consumption caused by IL-1Ra manipulation in the BLA is not likely due to IL-Ra induced changes in locomotor as a two-way repeated measures ANOVA only indicated a significant main effect of time [F(7,77)= 42.20, p<0.001] but not treatment [F(1,77)= 0.11, p=0.74] or any interaction [F(7,77)=1.47, p=0.19] on total distance traveled (Figure 5B). Moreover, no significant main effects or interactions were observed after T-tests of total time spent [t(11) =0.28, p=0.79] or distance traveled [t(11) =1.13, p=0.28] in the center of the open field arena suggesting that intra-BLA IL-1Ra does not induce an anxiolytic state which would also affect ethanol consumption (Figure 5C, 5D).
4. Discussion
Alcohol’s effects on the neuroimmune system has usually been studied in relation to alcohol-induced neurodegeneration, but the current study shows that alcohol-induced changes to the neuroimmune system have behavioral implications independent of neuronal loss. This work highlights a role of IL-1 receptor signaling in a site-specific manner within the CNS. The data herein provides evidence that: a) binge-like ethanol consumption increased IL-1β mRNA expression in the amygdala and immunoreactivity in the BLA, b) modulation of the neuroimmune system occurs independently of neuronal cell death, and c) inhibition of IL-1 receptor signaling within the BLA results in reduced binge-like ethanol consumption. Cumulatively these studies suggest that alcohol-induced increases of IL-1β in the BLA promote continued binge-like ethanol drinking which can be blunted with blockade of IL-1Ra in the BLA.
Manipulation of the immune system effects on ethanol consumption are not completely novel (Blednov et al., 2011; Blednov et al., 2012), but the present study not only identifies a direct role of a specific cytokine system, IL-1, but more importantly it shows IL-1 receptor signaling affects ethanol consumption via CNS actions rather than as a byproduct of peripheral actions. The importance of the IL-1 system in ethanol consumption has previously been highlighted by gene array studies and further validated with behavioral experiments in knockout mice (Blednov et al., 2012; Mulligan et al., 2006). However, global knockouts and peripheral manipulations lack site-specificity and cannot separate the developmental role of the cytokine and its specific actions on ethanol consumption. For example, IL1rn (the gene that encodes IL-1Ra) knockout mice consumed less ethanol compared to their wild-type controls (Blednov et al., 2012). This is contrary to the general consensus that anti-inflammatories reduce ethanol consumption and could potentially be due to compensatory developmental adaptations stemming from lifelong constitutive deletion of the gene. The present study, however, shows the relevance of site-directed pharmacology and concurs with the general consensus that an anti-inflammatory state reduces ethanol consumption. The critical role of IL-1 signaling on alcohol-induced behaviors has also been shown, as peripheral IL-1Ra but not minocycline reduce ethanol-induced sedation (Wu et al., 2011). However, the data presented herein provide direct evidence that neuroimmune signaling, specifically BLA IL-1 signaling, regulates ethanol consummatory behaviors. IL-1 receptor’s signaling effects on binge-like drinking were confined to the BLA as IL-1Ra had no effect in the adjoining CeA used as a site-directed control. The BLA also appeared to be particularly sensitive in terms of ethanol-induced changes in IL-1β immunoreactivity as no change was seen in the CeA. Importantly, the dose of IL-1Ra infused within the BLA had no independent effects on anxiety-like behavior or motor function according to data collected in the OFT. Moreover, the effect of IL-1Ra infusions appears to be specific to a function of ethanol as it had no bearing on sucrose consumption. IL-1 signaling within the BLA has also been shown to modulate heroin abuse indicating that the relationship between IL-1 signaling and the amygdala may not be unique to ethanol but may be involved in other drugs of abuse (Coller and Hutchinson, 2012; Szczytkowski and Lysle, 2008, 2010).
The present study shows an increase in a proinflammatory cytokine in a model of binge-like ethanol intake representing the early, more experimental stages of ethanol misuse in non-dependent subjects (Thiele and Navarro, 2014). Human alcoholics and preclinical models indicate an increased proinflammatory state (Crews et al., 2011; He and Crews, 2008), but these findings were after a lifetime of alcohol abuse or in models with repeated binge exposure (Osterndorff-Kahanek et al., 2015; Qin and Crews, 2012b). However, what is unique about the present studies is that changes in IL-1β immunoreactivity and mRNA expression occurred prior to dependence and in subjects that were self-administering ethanol. Moreover, the alcohol-induced cytokine increase occurred at BECs well below those reported in previous acute and chronic studies, with BECs around 300mg/dL (Hayes et al., 2013; Qin and Crews, 2012a). Others have shown decreases in IL-1β expression following a bolus ethanol intragastric challenge in rats after a period of abstinence agreeing with others who report that the timing of analysis can affect neuroimmune responses (Doremus-Fitzwater et al., 2014; Marshall et al., 2013). The findings herein, in respect to other studies on alcohol-induce alteration of IL-1β, indicate that changes in IL-1β occur during early ethanol consumption and at more moderate BECs than previously described. Importantly, our findings provide the first evidence that IL-1 signaling is recruited during excessive ethanol consumption and prior to dependence, and thus may play a critical role in modulating neuroplastic changes that occur during the transition to ethanol dependence.
Ethanol-induced upregulation of IL-1β mRNA expression observed within the amygdala persisted even in the absence of alcohol. These findings are in agreement with others showing ethanol elicited neuroimmune system dysfunction even a month after abstinence (Marshall et al., 2013). Persistent activation of glia and other neuroimmune responses can lead to exacerbated dysfunction in the presence of other immune challenges (Vetreno and Crews, 2014; Williamson et al., 2011). Moreover, the IL-1β mRNA response was initiated after just one cycle of DID, whereas the increase in protein expression was not significantly different until after three cycles. The strong trend of an increase after one week suggests that it does not necessarily take multiple binge exposures to increase the protein, but the possibility that there is a delayed response from just 1 cycle that causes the increase after 3 cycles cannot be completely ruled out with the current data set. Regardless of when the IL-1β response is initiated, these studies highlight the enduring nature of increases in IL-1β mRNA in a model that represents the early stages of alcohol abuse. This plastic change is particularly alarming and suggests the neuroimmune response could be an early indicator of future addiction liability and may eventually contribute to cellular damage with repetitive exposure.
The persisting nature of the alcohol-induced neuroimmune response and the fact that it is proinflammatory led to the theory that alcohol-induced neuroimmune responses occur because of neurodegeneration but also potentially exacerbate damage (Crews et al., 2011; Vetreno and Crews, 2014). However, following multiple cycles of DID, this study indicates that neurodegeneration does not occur. The lack of cell death indicates that ethanol can elicit a neuroimmune response independent of significant brain damage. It is important to denote that the mechanism by which Fluoro-jade staining works is unknown, therefore it is perceivable that early stages of cellular death may not be incorporated in these cell counts (Poirier et al., 2000). However, very little FJC+ cells were expected to be induced by three cycles of DID as the BECs produced in the DID paradigm likely do not reach a level that would cause neurodegeneration (Liput et al., 2013). This is only the second paper to demonstrate that the DID paradigm does not elicit cellular damage (Sprow et al., 2015), but the first to use a marker of neurodegeneration. The enduring change of IL-1β mRNA without any neurodegeneration in this model further implicates IL-1 in a signaling role and not just as an indicator of cellular stress.
The underlying signaling pathways responsible for altered neurotransmission remain elusive with the current set of data. However, it has been previously shown that IL-1Ra can exert both indirect (glial driven) and direct (neuronal driven) control of glutamatergic transmission (Chu et al., 2010; De Chiara et al., 2013). In particular, glutamatergic projections from the BLA, where IL-1 signaling had an effect on ethanol consumption, convey nociceptive information to the CeA (Veinante et al., 2013). Evidence suggests that CeA neuropeptides associated with nociception, such as CRF and NPY, modulate binge-like drinking (Lowery-Gionta et al., 2012; Sparrow et al., 2012; Veinante et al., 2013). Although these studies did not determine an effect of IL-1Ra administration in the CeA, recent evidence revealed that IL-Ra and IL-1β can regulate GABAergic transmission in response to ethanol in the CeA (Bajo et al., 2015a; Bajo et al., 2015b). Although Bajo and colleagues only examined the CeA, they suggest that the effects of IL-1 signaling in relation to alcohol-induced GABAergic tone would likely be seen in other regions as well (Bajo et al., 2015a). While the studies herein show that IL-1Ra administration within the BLA reduced binge-like drinking, it is perceivable that IL-1 receptor signaling in other regions include the CeA may have a different behavioral outcomes. Future studies are necessary to elucidate the underlying neurocircuitry and cell signaling cascades of IL-1 receptor signaling that contribute to binge-like drinking as well as other stages of AUD.
Alcoholism as with many drugs of abuse often develops after an experimental period. In alcohol misuse, the experimental period often includes binge drinking. The current findings provide evidence that binge-like consumption can alter the neuroimmune system prior to dependence or evidence of neuronal cell death. Furthermore, these results show that manipulating IL-1 signaling centrally modulates binge-like ethanol consumption. Together these findings with previous research indicate that the neuroimmune system remains a viable target for exploration in reducing binge-like drinking, but the impacts of other cytokine receptors, especially anti-inflammatory cytokines, in the amygdala as well as other regions of interests on ethanol consumption remain elusive.
Amygdalar IL-1β mRNA is increased for an extended period after binge-like drinking
Alcohol-induced IL-1β increased immunoreactivity is specifically in the BlA
Ethanol associated changes in IL-1β occurred independent of neurodegeneration
IL-1R antagonist administration in the BlA but not CeA reduced ethanol consumption
IL-1 receptor signaling in the amygdala has a role in binge-like drinking
Acknowledgments
The authors thank Timothy P. Gilliam and Rhiannon D. Thomas for their technical assistance.
Funding
This work was supported by National Institutes of Health grants AA022048, AA013573, AA015148, AA021611, AA011605, & GM000678.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Disclosure:
The authors declare no conflict of interest.
5. References
- Bajo M, Herman MA, Varodayan FP, Oleata CS, Madamba SG, Harris RA, Blednov YA, Roberto M. Role of the IL-1 receptor antagonist in ethanol-induced regulation of GABAergic transmission in the central amygdala. Brain Behav Immun. 2015a;45:189–197. doi: 10.1016/j.bbi.2014.11.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bajo M, Varodayan FP, Madamba SG, Robert AJ, Casal LM, Oleata CS, Siggins GR, Roberto M. IL-1 interacts with ethanol effects on GABAergic transmission in the mouse central amygdala. Frontiers in pharmacology. 2015b;6:49. doi: 10.3389/fphar.2015.00049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bell RL, Lopez MF, Cui C, Egli M, Johnson KW, Franklin KM, Becker HC. Ibudilast reduces alcohol drinking in multiple animal models of alcohol dependence. Addict Biol. 2013 doi: 10.1111/adb.12106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blednov YA, Benavidez JM, Geil C, Perra S, Morikawa H, Harris RA. Activation of inflammatory signaling by lipopolysaccharide produces a prolonged increase of voluntary alcohol intake in mice. Brain Behav Immun. 2011;25(Suppl 1):S92–S105. doi: 10.1016/j.bbi.2011.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blednov YA, Ponomarev I, Geil C, Bergeson S, Koob GF, Harris RA. Neuroimmune regulation of alcohol consumption: behavioral validation of genes obtained from genomic studies. Addict Biol. 2012;17:108–120. doi: 10.1111/j.1369-1600.2010.00284.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- CDC Vital signs: binge drinking prevalence, frequency, and intensity among adults - United States, 2010. MMWR Morb Mortal Wkly Rep. 2013;61:14–19. [PubMed] [Google Scholar]
- Chiu GS, Darmody PT, Walsh JP, Moon ML, Kwakwa KA, Bray JK, McCusker RH, Freund GG. Adenosine through the A2A adenosine receptor increases IL-1beta in the brain contributing to anxiety. Brain Behav Immun. 2014;41:218–231. doi: 10.1016/j.bbi.2014.05.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chu YX, Zhang Y, Zhang YQ, Zhao ZQ. Involvement of microglial P2X7 receptors and downstream signaling pathways in long-term potentiation of spinal nociceptive responses. Brain Behav Immun. 2010;24:1176–1189. doi: 10.1016/j.bbi.2010.06.001. [DOI] [PubMed] [Google Scholar]
- Coller JK, Hutchinson MR. Implications of central immune signaling caused by drugs of abuse: mechanisms, mediators and new therapeutic approaches for prediction and treatment of drug dependence. Pharmacol Ther. 2012;134:219–245. doi: 10.1016/j.pharmthera.2012.01.008. [DOI] [PubMed] [Google Scholar]
- Correia D, Martynhak BJ, Pereira M, Siba IP, Ribeiro AF, Camarini R, Boerngen-Lacerda R. Reduction of ethanol intake by corticotropin-releasing factor receptor-1 antagonist in "heavy-drinking" mice in a free-choice paradigm. Psychopharmacology (Berl) 2015 doi: 10.1007/s00213-015-3909-y. [DOI] [PubMed] [Google Scholar]
- Cox BR, Olney JJ, Lowery-Gionta EG, Sprow GM, Rinker JA, Navarro M, Kash TL, Thiele TE. Repeated cycles of binge-like ethanol (EtOH)-drinking in male C57BL/6J mice augments subsequent voluntary EtOH intake but not other dependence-like phenotypes. Alcohol Clin Exp Res. 2013;37:1688–1695. doi: 10.1111/acer.12145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crews FT, Vetreno RP. Neuroimmune basis of alcoholic brain damage. Int Rev Neurobiol. 2014;118:315–357. doi: 10.1016/B978-0-12-801284-0.00010-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crews FT, Zou J, Qin L. Induction of innate immune genes in brain create the neurobiology of addiction. Brain Behav Immun. 2011;25(Suppl 1):S4–S12. doi: 10.1016/j.bbi.2011.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Chiara V, Motta C, Rossi S, Studer V, Barbieri F, Lauro D, Bernardi G, Centonze D. Interleukin-1beta alters the sensitivity of cannabinoid CB1 receptors controlling glutamate transmission in the striatum. Neuroscience. 2013;250:232–239. doi: 10.1016/j.neuroscience.2013.06.069. [DOI] [PubMed] [Google Scholar]
- Der-Avakian A, Markou A. The neurobiology of anhedonia and other reward-related deficits. Trends Neurosci. 2012;35:68–77. doi: 10.1016/j.tins.2011.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doremus-Fitzwater TL, Buck HM, Bordner K, Richey L, Jones ME, Deak T. Intoxication- and withdrawal-dependent expression of central and peripheral cytokines following initial ethanol exposure. Alcohol Clin Exp Res. 2014;38:2186–2198. doi: 10.1111/acer.12481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Egli M, Koob GF, Edwards S. Alcohol dependence as a chronic pain disorder. Neurosci Biobehav Rev. 2012;36:2179–2192. doi: 10.1016/j.neubiorev.2012.07.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ericsson A, Liu C, Hart RP, Sawchenko PE. Type 1 interleukin-1 receptor in the rat brain: distribution, regulation, and relationship to sites of IL-1-induced cellular activation. J Comp Neurol. 1995;361:681–698. doi: 10.1002/cne.903610410. [DOI] [PubMed] [Google Scholar]
- Fee JR, Sparta DR, Knapp DJ, Breese GR, Picker MJ, Thiele TE. Predictors of high ethanol consumption in RIIbeta knock-out mice: assessment of anxiety and ethanol-induced sedation. Alcohol Clin Exp Res. 2004;28:1459–1468. doi: 10.1097/01.ALC.0000141809.53115.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernandez-Lizarbe S, Montesinos J, Guerri C. Ethanol induces TLR4/TLR2 association, triggering an inflammatory response in microglial cells. J Neurochem. 2013 doi: 10.1111/jnc.12276. [DOI] [PubMed] [Google Scholar]
- Ferreira R, Xapelli S, Santos T, Silva AP, Cristovao A, Cortes L, Malva JO. Neuropeptide Y modulation of interleukin-1{beta} (IL-1{beta})-induced nitric oxide production in microglia. J Biol Chem. 2010;285:41921–41934. doi: 10.1074/jbc.M110.164020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fox HC, Bergquist KL, Hong KI, Sinha R. Stress-induced and alcohol cue-induced craving in recently abstinent alcohol-dependent individuals. Alcohol Clin Exp Res. 2007;31:395–403. doi: 10.1111/j.1530-0277.2006.00320.x. [DOI] [PubMed] [Google Scholar]
- Gilpin NW, Herman MA, Roberto M. The central amygdala as an integrative hub for anxiety and alcohol use disorders. Biol Psychiatry. 2015;77:859–869. doi: 10.1016/j.biopsych.2014.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gunzerath L, Hewitt BG, Li TK, Warren KR. Alcohol research: past, present, and future. Ann N Y Acad Sci. 2011;1216:1–23. doi: 10.1111/j.1749-6632.2010.05832.x. [DOI] [PubMed] [Google Scholar]
- Hayes DM, Deeny MA, Shaner CA, Nixon K. Determining the threshold for alcohol-induced brain damage: new evidence with gliosis markers. Alcohol Clin Exp Res. 2013;37:425–434. doi: 10.1111/j.1530-0277.2012.01955.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- He J, Crews FT. Increased MCP-1 and microglia in various regions of the human alcoholic brain. Experimental neurology. 2008;210:349–358. doi: 10.1016/j.expneurol.2007.11.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hutchinson MR, Watkins LR. Why is neuroimmunopharmacology crucial for the future of addiction research? Neuropharmacology. 2014;76:218–227. doi: 10.1016/j.neuropharm.2013.05.039. Pt B. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones ME, Lebonville CL, Barrus D, Lysle DT. The role of brain interleukin-1 in stress-enhanced fear learning. Neuropsychopharmacology. 2015;40:1289–1296. doi: 10.1038/npp.2014.317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kageyama K, Kagaya S, Takayasu S, Hanada K, Iwasaki Y, Suda T. Cytokines induce NF-kappaB, Nurr1 and corticotropin-releasing factor gene transcription in hypothalamic 4B cells. Neuroimmunomodulation. 2010;17:305–313. doi: 10.1159/000292022. [DOI] [PubMed] [Google Scholar]
- Kim HS, Lee G, John SW, Maeda N, Smithies O. Molecular phenotyping for analyzing subtle genetic effects in mice: application to an angiotensinogen gene titration. Proc Natl Acad Sci U S A. 2002;99:4602–4607. doi: 10.1073/pnas.072083799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lippai D, Bala S, Petrasek J, Csak T, Levin I, Kurt-Jones EA, Szabo G. Alcohol-induced IL-1beta in the brain is mediated by NLRP3/ASC inflammasome activation that amplifies neuroinflammation. J Leukoc Biol. 2013;94:171–182. doi: 10.1189/jlb.1212659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liput DJ, Hammell DC, Stinchcomb AL, Nixon K. Transdermal delivery of cannabidiol attenuates binge alcohol-induced neurodegeneration in a rodent model of an alcohol use disorder. Pharmacology, biochemistry, and behavior. 2013;111:120–127. doi: 10.1016/j.pbb.2013.08.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu L, Hutchinson MR, White JM, Somogyi AA, Coller JK. Association of IL-1B genetic polymorphisms with an increased risk of opioid and alcohol dependence. Pharmacogenetics and genomics. 2009;19:869–876. doi: 10.1097/FPC.0b013e328331e68f. [DOI] [PubMed] [Google Scholar]
- Lowery-Gionta EG, Navarro M, Li C, Pleil KE, Rinker JA, Cox BR, Sprow GM, Kash TL, Thiele TE. Corticotropin releasing factor signaling in the central amygdala is recruited during binge-like ethanol consumption in C57BL/6J mice. J Neurosci. 2012;32:3405–3413. doi: 10.1523/JNEUROSCI.6256-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Markou A, Kosten TR, Koob GF. Neurobiological similarities in depression and drug dependence: a self-medication hypothesis. Neuropsychopharmacology. 1998;18:135–174. doi: 10.1016/S0893-133X(97)00113-9. [DOI] [PubMed] [Google Scholar]
- Marshall SA, McClain JA, Kelso ML, Hopkins DM, Pauly JR, Nixon K. Microglial activation is not equivalent to neuroinflammation in alcohol-induced neurodegeneration: The importance of microglia phenotype. Neurobiol Dis. 2013;54:239–251. doi: 10.1016/j.nbd.2012.12.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Minami M, Kuraishi Y, Yamaguchi T, Nakai S, Hirai Y, Satoh M. Immobilization stress induces interleukin-1 beta mRNA in the rat hypothalamus. Neurosci Lett. 1991;123:254–256. doi: 10.1016/0304-3940(91)90944-o. [DOI] [PubMed] [Google Scholar]
- Mulligan MK, Ponomarev I, Hitzemann RJ, Belknap JK, Tabakoff B, Harris RA, Crabbe JC, Blednov YA, Grahame NJ, Phillips TJ, Finn DA, Hoffman PL, Iyer VR, Koob GF, Bergeson SE. Toward understanding the genetics of alcohol drinking through transcriptome meta-analysis. Proc Natl Acad Sci U S A. 2006;103:6368–6373. doi: 10.1073/pnas.0510188103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Osterndorff-Kahanek EA, Becker HC, Lopez MF, Farris SP, Tiwari GR, Nunez YO, Harris RA, Mayfield RD. Chronic ethanol exposure produces time- and brain region-dependent changes in gene coexpression networks. PLoS One. 2015;10:e0121522. doi: 10.1371/journal.pone.0121522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paxinos G, Franklin KBJ. Elsevier Academic Press; Amsterdam ; Boston: 2004. The mouse brain in stereotaxic coordinates. [Google Scholar]
- Pleil KE, Rinker JA, Lowery-Gionta EG, Mazzone CM, McCall NM, Kendra AM, Olson DP, Lowell BB, Grant KA, Thiele TE, Kash TL. NPY signaling inhibits extended amygdala CRF neurons to suppress binge alcohol drinking. Nat Neurosci. 2015;18:545–552. doi: 10.1038/nn.3972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poirier JL, Capek R, De Koninck Y. Differential progression of Dark Neuron and Fluoro-Jade labelling in the rat hippocampus following pilocarpine-induced status epilepticus. Neuroscience. 2000;97:59–68. doi: 10.1016/s0306-4522(00)00026-9. [DOI] [PubMed] [Google Scholar]
- Prut L, Belzung C. The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: a review. Eur J Pharmacol. 2003;463:3–33. doi: 10.1016/s0014-2999(03)01272-x. [DOI] [PubMed] [Google Scholar]
- Qin L, Crews FT. Chronic ethanol increases systemic TLR3 agonist-induced neuroinflammation and neurodegeneration. J Neuroinflammation. 2012a;9:130. doi: 10.1186/1742-2094-9-130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin L, Crews FT. NADPH oxidase and reactive oxygen species contribute to alcohol-induced microglial activation and neurodegeneration. J Neuroinflammation. 2012b;9:5. doi: 10.1186/1742-2094-9-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin L, He J, Hanes RN, Pluzarev O, Hong JS, Crews FT. Increased systemic and brain cytokine production and neuroinflammation by endotoxin following ethanol treatment. J Neuroinflammation. 2008;5:10. doi: 10.1186/1742-2094-5-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin L, Wu X, Block ML, Liu Y, Breese GR, Hong JS, Knapp DJ, Crews FT. Systemic LPS causes chronic neuroinflammation and progressive neurodegeneration. Glia. 2007;55:453–462. doi: 10.1002/glia.20467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rhodes JS, Best K, Belknap JK, Finn DA, Crabbe JC. Evaluation of a simple model of ethanol drinking to intoxication in C57BL/6J mice. Physiol Behav. 2005;84:53–63. doi: 10.1016/j.physbeh.2004.10.007. [DOI] [PubMed] [Google Scholar]
- Rivest S, Rivier C. Stress and interleukin-1 beta-induced activation of c-fos, NGFI-B and CRF gene expression in the hypothalamic PVN: comparison between Sprague-Dawley, Fisher-344 and Lewis rats. Journal of neuroendocrinology. 1994;6:101–117. doi: 10.1111/j.1365-2826.1994.tb00559.x. [DOI] [PubMed] [Google Scholar]
- Schmued LC, Stowers CC, Scallet AC, Xu L. Fluoro-Jade C results in ultra high resolution and contrast labeling of degenerating neurons. Brain Res. 2005;1035:24–31. doi: 10.1016/j.brainres.2004.11.054. [DOI] [PubMed] [Google Scholar]
- Sparrow AM, Lowery-Gionta EG, Pleil KE, Li C, Sprow GM, Cox BR, Rinker JA, Jijon AM, Pena J, Navarro M, Kash TL, Thiele TE. Central neuropeptide Y modulates binge-like ethanol drinking in C57BL/6J mice via Y1 and Y2 receptors. Neuropsychopharmacology. 2012;37:1409–1421. doi: 10.1038/npp.2011.327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sprow GM, Rinker JA, Lowery-Gointa EG, Sparrow AM, Navarro M, Thiele TE. Lateral hypothalamic melanocortin receptor signaling modulates binge-like ethanol drinking in C57BL/6J mice. Addict Biol. 2015 doi: 10.1111/adb.12264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szczytkowski JL, Lysle DT. Conditioned effects of heroin on proinflammatory mediators require the basolateral amygdala. Eur J Neurosci. 2008;28:1867–1876. doi: 10.1111/j.1460-9568.2008.06472.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szczytkowski JL, Lysle DT. Dopamine D1 receptors within the basolateral amygdala mediate heroin-induced conditioned immunomodulation. J Neuroimmunol. 2010;226:38–47. doi: 10.1016/j.jneuroim.2010.05.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thiele TE, Navarro M. "Drinking in the dark" (DID) procedures: A model of binge-like ethanol drinking in non-dependent mice. Alcohol (Fayetteville, N.Y. 2014;48:235–241. doi: 10.1016/j.alcohol.2013.08.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turrin NP, Gayle D, Ilyin SE, Flynn MC, Langhans W, Schwartz GJ, Plata-Salaman CR. Pro-inflammatory and anti-inflammatory cytokine mRNA induction in the periphery and brain following intraperitoneal administration of bacterial lipopolysaccharide. Brain Res Bull. 2001;54:443–453. doi: 10.1016/s0361-9230(01)00445-2. [DOI] [PubMed] [Google Scholar]
- Veinante P, Yalcin I, Barrot M. The amygdala between sensation and affect: a role in pain. Journal of molecular psychiatry. 2013;1:9. doi: 10.1186/2049-9256-1-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vetreno RP, Crews FT. Current hypotheses on the mechanisms of alcoholism. Handbook of clinical neurology. 2014;125:477–497. doi: 10.1016/B978-0-444-62619-6.00027-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williamson LL, Sholar PW, Mistry RS, Smith SH, Bilbo SD. Microglia and memory: modulation by early-life infection. J Neurosci. 2011;31:15511–15521. doi: 10.1523/JNEUROSCI.3688-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wohleb ES, Patterson JM, Sharma V, Quan N, Godbout JP, Sheridan JF. Knockdown of interleukin-1 receptor type-1 on endothelial cells attenuated stress-induced neuroinflammation and prevented anxiety-like behavior. J Neurosci. 2014;34:2583–2591. doi: 10.1523/JNEUROSCI.3723-13.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu Y, Lousberg EL, Moldenhauer LM, Hayball JD, Robertson SA, Coller JK, Watkins LR, Somogyi AA, Hutchinson MR. Attenuation of microglial and IL-1 signaling protects mice from acute alcohol-induced sedation and/or motor impairment. Brain Behav Immun. 2011;25(Suppl 1):S155–164. doi: 10.1016/j.bbi.2011.01.012. [DOI] [PubMed] [Google Scholar]
- Yabuuchi K, Minami M, Katsumata S, Satoh M. Localization of type I interleukin-1 receptor mRNA in the rat brain. Brain Res Mol Brain Res. 1994;27:27–36. doi: 10.1016/0169-328x(94)90180-5. [DOI] [PubMed] [Google Scholar]
- Zou J, Crews FT. Inflammasome-IL-1beta Signaling Mediates Ethanol Inhibition of Hippocampal Neurogenesis. Frontiers in neuroscience. 2012;6:77. doi: 10.3389/fnins.2012.00077. [DOI] [PMC free article] [PubMed] [Google Scholar]