

Abstract
This study analyzes the impact of droughts, compared with average climatic conditions, on the supporting ecosystem service water provision in sub‐watersheds in managed alpine grasslands in two climatically different regions of the Alps, Lautaret (French Alps) and Stubai (Austrian Alps). Soil moisture was modelled in the range of 0–0.3 m. At both sites, current patterns showed that the mean seasonal soil moisture was (1) near field capacity for grasslands with low management intensity and (2) below field capacity for grasslands with higher land‐use intensity. Soil moisture was significantly reduced by drought at both sites, with lower reductions at the drier Lautaret site. At the sub‐watershed scale, soil moisture spatial heterogeneity was reduced by drought. Under drought conditions, the evapotranspiration to precipitation ratios at Stubai was slightly higher than those at Lautaret, indicating a dominant ‘water spending’ strategy of plant communities. Regarding catchment water balance, deep seepage was reduced by drought at Stubai more strongly than at Lautaret. Hence, the observed ‘water spending’ strategy at Stubai might have negative consequences for downstream water users. Assessing the water provision service for alpine grasslands provided evidence that, under drought conditions, evapotranspiration was influenced not only by abiotic factors but also by the water‐use strategy of established vegetation. These results highlight the importance of ‘water‐use’ strategies in existing plant communities as predictors of the impacts of drought on water provision services and related ecosystem services at both the field and catchment scale. © 2015 The Authors. Ecohydrology published by John Wiley & Sons, Ltd.
Keywords: climate change, deep percolation, evaporation, HILLFLOW model, spatial analyses, transpiration
Introduction
Increased frequencies and intensities of droughts are extreme events that are projected to occur in terrestrial ecosystems by the end of the 21st century at the latest (Reichstein et al., 2013; Bahn et al., 2014). Given these expectations, the related changes in ecosystem services (ESs) (e.g. biomass and forage quality, carbon sequestration and water provision) are being studied intensively (Jentsch et al., 2011; Walter et al., 2012; Lamarque et al., 2014), as well as their effects on society, economy and the Earth system (Cerda et al., 1998; Vetter & Bond, 2012; Martínez‐Garza et al., 2013; Reichstein et al., 2013). A crucial step for the successful implementation of the concept of ES (Lamarque et al., 2011a) in regional resource management and policy is to quantify and map the provisioning of ESs (Seppelt et al. 2011; Burkhard et al. 2012). Quantification is necessary to evaluate the trade‐offs between ESs when making decisions (Egoh et al., 2008; Seppelt et al., 2011; Crossman et al., 2012). The most promising techniques for quantification, reported in the study of Martínez‐Harms and Balvanera (2012), are the following: (1) extrapolation of primary data to the analysed area, producing spatially explicit results by combining quantitative and qualitative aspects; however, careful interpretation of these results is needed because of the potential lack of representation of the stochastic, scale‐dependent and nonlinear nature of ecological processes, thereby producing a uniformity of error. This may occur when an average ES value is attributed to a distinct cartographical unit (i.e. land use/cover type) and (2) a combination of plot measurements and empirical geostatistical models to spatially model ESs provision. Detailed reviews on the studies can be found in the study of Martínez‐Harms and Balvanera (2012) and Nemec and Raudsepp‐Hearne (2013).
Nevertheless, depending on the nature of the ecosystem functions and processes, the use of a process‐based (e.g. physical) environmental model is occasionally obligatory to correctly reproduce spatial configurations and patterns (Eigenbrod et al., 2010). This is particularly true for hydrological ESs that are strongly affected by the changes in ecosystems due to global change (Beniston et al., 2007; Bangash et al., 2013; Crossman et al., 2013; Liu et al., 2013), for which topographical and physical characteristics and the related three‐dimensional movement of the water must be taken into account. Hydrological ES in mountain areas is based on complex interactions between topography, soil characteristics, vegetation and climate, all of which influence the runoff production (Wigmosta et al., 1994). These characteristics require high‐quality models for quantification. However, complex models propagate uncertainty based on two causes: (1) model uncertainty, i.e. uncertainty regarding the description of processes in the models and uncertainties due to the parameter interactions in more complex models and (2) multiplication of parameters, which results in additional sources of uncertainty.
In particular, mountainous areas are important target regions for the quantification of hydrological ESs due to their vital role as the water towers of the world (Viviroli et al., 2003; Messerli et al., 2004). Although only a quarter of the world's population lives in mountainous areas (Meybeck et al., 2001), more than half of it relies on water coming from the mountains (Beniston, 2006). Additionally, mountain areas are known to be more affected by extraordinary rainfall events than other landscapes (Serrano‐Muela et al., 2013; Taguas et al., 2013; Wang et al., 2014). The future climate will involve, not only increased air temperatures but also more frequent and more intense extreme events (Seneviratne et al., 2012; Bahn et al., 2014; Köplin et al., 2014). Grasslands are primarily susceptible to drought events and less susceptible to other extremes (Reichstein et al., 2013). Drought affects water provision and consequently important ESs in managed grasslands such as forage production or forage quality (Zwicke et al., 2013, but refer to the study of Jentsch et al. (2011)). However, Benot et al. (2013a) found that the short‐term influence of management on plant diversity and biomass production was stronger than the influence of extreme summer weather for the upper valley of the Romanche River in the Central French Alps. Therefore, both climate change effects in the long term and distinct responses of grassland ecosystems to extreme events are expected because grassland ES is affected by both climate‐use and land‐use changes (Soussana & Luscher, 2007).
This study aimed to analyse the impacts of drought on the supporting water provision ES in managed mountain grasslands in two climatically different regions of the Alps. Stakeholder consultations in both regions revealed the importance of water provision for forage production, grazing and hydropower (Lamarque et al., 2011b). As indicators of ES water provision, stakeholders living in the French Alps stressed the importance of soil moisture affected by drier climatic conditions, while stakeholders from the study site in the Austrian Alps mentioned water quantity (i.e. hydropower) as being more important (Lamarque et al., 2011b). To address the future challenges for grassland management due to climate change, the impacts of seasonal droughts on soil moisture and water quantity based on highly probable precipitation scenarios (Beniston, 2006; Beniston, 2012; Strauss et al., 2013) were evaluated.
Our objectives were as follows:
to implement a physically based, well‐calibrated hydrological model for alpine grassland sites at two climatically different study sites;
to quantify the water provision service by modelling soil moisture and water quantity for the current land use/cover at both sites under current climate conditions;
to evaluate the impacts of seasonal drought on soil moisture and water quantity at each of these sites given the current land‐use conditions
Materials and Methods
Study areas
The experimental long‐term socio‐ecological research (LTSER) site Lautaret (F) (Lavorel et al., 2013) covers 12.92 km2 between 1650 m and 2500 m a.s.l. and is located on the south facing the slopes of the valley above the village of Villar d'Arène in the central French Alps (N45.04°, E6.34°; Figure 1). Lautaret is characterized by a subalpine climate with a strong continental influence due to a rain shadow effect deriving from dominant westerly winds. The mean annual air temperature is 5 °C at 1650 m a.s.l. and 3 °C at 2000 m a.s.l.; the mean annual precipitation is 956 mm. Most of the precipitation falls as snow during the winter, whereas ca 18% of the annual rainfall occurs during the summer. The current landscape is dominated by grassland ecosystems of the soil type Cambisol. These grasslands are still used by a small and active farming community centred around sheep and cattle rearing for lamb and steer production. At lower altitudes (e.g. 1650–2000 m a.s.l.), former arable fields have been abandoned and subsequently converted to terraced grasslands used for hay production or grazing. At midslope (e.g., 1800‐2200 m a.s.l.), ancient, never ploughed hay meadows are increasingly used for light summer grazing by sheep or cattle; a small fraction of these areas are no longer cut or grazed at all. The upper slopes (2200–2500 m a.s.l.) are grazed by transhumant flocks during the summer (Quetier et al., 2007).
Figure 1.
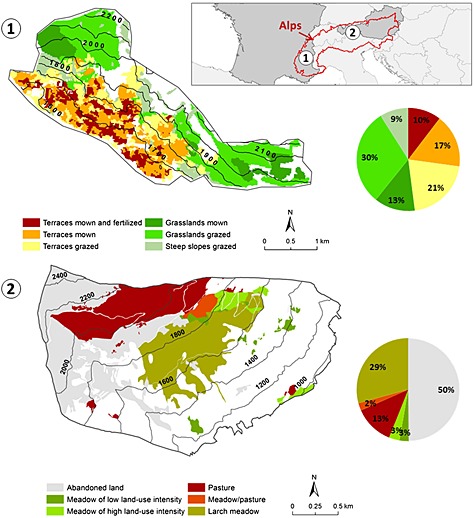
Location of the study areas in the Alps: site Lautaret in the Upper Romanche Valley in France (1); site Stubai in the Stubai Valley in Austria (2). The regions of the study areas depicted in white represent non‐grassland.
The experimental site Stubai (A) is located at the LTSER site Kaserstattalm (Tappeiner et al., 2013) in the Stubai Valley in the Austrian Alps and covers 4.93 km2 between 970 and 2200 m a.s.l. (N47.13°, E11.33°; Figure 1). The mean annual precipitation ranges from 852 to 1097 mm at 970 and 1900 m a.s.l., respectively. The mean annual air temperature is 6.5 °C at 970 m a.s.l. and 3.0 °C at 1900 m a.s.l. The present grasslands are found on the soil type Dystric Cambisol and differ in management intensity. A tendency towards reduced management and less intensive grazing since the 1950s has resulted in a successive change of abandoned areas to shrublands and young forest stands, shaping the landscape structure of the Stubai Valley mountain region.
Further details on grassland types as well as relevant soil and vegetation characteristics can be found in Table 1. Please note that to allow comparison with other studies at our experimental sites, numbering of grassland types does not follow chronological order.
Table 1.
Area of investigated grassland types for Lautaret (L) and Stubai (S) with soil hydrological properties at a depth of 0–0.3 m [field capacity (FC), gross pore volume (GPV), permanent wilting point (PWP)], evapotranspiration of grassland community May–October (ET) and vegetation type (data for Lautaret: Quetier et al. (2007), Obojes et al. (2014); data for Stubai: Obojes et al. (2014), unpublished measurements by the authors).
| Grassland type | Description | Area | FC | PWP | ET | Plant community |
|---|---|---|---|---|---|---|
| (ha) | (Vol%) | (Vol%) | (mm d−1) | |||
| L1 | Previously cultivated terraces (1550–1950 m), now manured and mown for hay | 73.1 | 37 | 25 | 2.9–4.1 | Mown and fertilized meadows dominated by Dactylis glomerata, and Trisetetum flavescens |
| L2 | Previously cultivated terraces (1550–1950 m), now mown for hay but not manured | 118.1 | 40 | 23 | 2.8–3.5 | Mown meadows dominated by B. erectus and Sesleria caeruela |
| L3 | Previously cultivated terraces (1550–1950 m), now unmown and grazed in spring and autumn | 146.3 | 38 | 22 | 2.9–3.6 | Pastures with low management intensity dominated by B. erectus and S. caeruela |
| L4 | Never cultivated unterraced grasslands (1700–2000 m), currently mown for hay | 90.0 | 37 | 22 | 3.0–3.6 | Mown meadows dominated by Festuca paniculata, Meum athamanticum, and Trifolium alpinum |
| L5 | Never cultivated unterraced grasslands (1700–2000 m), summer grazed | 213.2 | 38 | 21 | 3.4–3.9 | Pastures of low management intensity dominated by F. paniculata and Festuca nigrescens |
| L9 | Steep and rocky slopes, grazed | 63.0 | 37 | 20 | 2.9–3.6 | Sloping pastures with very low management intensity dominated by B erectus, S. caeruela and several xeric species |
| S2 | Abandoned land | 113.0 | 40 | 20 | 3.8–6.9 | Abandoned meadow dominated by Luzula sylvatica, Geranium sylvaticum and Potentilla aurea, increasing abundance of Vaccinium sp. |
| S3 | Pasture | 28.0 | 37 | 21 | 3.8–7.2 | Pasture of high management intensity dominated by A. vulgaris, L. hispidus, and R. acris |
| S4 | Meadow of low land‐use intensity (1100–1900 m), mown every 2 years, not manured | 6.5 | 37 | 20 | 4.1–7.2 | Mown meadows of low management intensity dominated by Leontodon hispidus, Agrostis capillaris, and Plantago lanceolata |
| S6 | Meadow of high land‐use intensity (1600–1900 m), 1–2 mowings per year, manured | 7.5 | 37 | 20 | 4.1–7.2 | Mown and fertilized meadows of high management intensity dominated by Alchemilla vulgaris, A. capillaris, and Ranunculus acris |
| S8 | Meadow/pasture | 5.3 | 42 | 21 | 4.1–7.0 | Mown meadow and pasture of low management intensity dominated by A. vulgaris, A. capillaris, and R. acris |
| S9 | Larch meadow | 66.3 | 40 | 20 | 3.6–6.7 | Mown meadow of low management intensity with Larix decidua dominated by L. hispidus, A. capillaris, and P. lanceolata |
Hydrological model HILLFLOW
Given that both investigated study areas are not closed catchments, hydrological modelling was performed with the 3D version of the hillslope model HILLFLOW developed by Axel Bronstert (University of Potsdam) and Erich Plate (University of Karlsruhe). HILLFLOW is described by its authors as a physically based, distributed hydrological model at the hillslope and small catchment scale, although there are some sub‐models containing empirical approaches (e.g. Mualem–van Genuchten model for unsaturated water flow). Detailed information can be found, e.g. in the study of Bronstert (1999), Bronstert and Bardossy (1999) and Bronstert and Plate (1997).
Hydrological modelling and model calibration were performed based on land‐use/cover (LUC) maps and a comprehensive database on (micro) climatic variables, botanical compositions, plant functional traits, soil characteristics and a range of ecosystem properties (e.g. evapotranspiration, biomass production, litter decomposition, nitrogen stocks and fluxes, etc.) of more than 60 permanent plots distributed across different LUC types and altitudes (Lavorel et al., 2013; Tappeiner et al., 2013). HILLFLOW 3D has a grid‐based discretization with quadratic‐shaped, constant‐sized grids (Bronstert, 1995). Based on the available resolution of digital elevation models and LUC‐maps, the grid size was 75 m for Lautaret and 50 m for Stubai. As input data for each cell, parameters regarding the existing soil type (e.g. soil texture, saturated water content, field capacity, residual soil water content, saturated hydraulic conductivity, soil depth and macropores), vegetation type (e.g. root depth, evapotranspiration and canopy interception), topography (e.g. slope and altitude) and climate (e.g. precipitation) were required for the existing grassland types (Table 1). Among these parameters, Mualem–van Genuchten parameters (α, n), saturated water content, saturated hydraulic conductivity, canopy interception and proportion of macropores were estimated from available soil moisture observations. Drought effects were modelled by reducing the long‐term average (e.g. normal) precipitation (daily temporal resolution, Table 2). At Lautaret, the simulation of drought conditions was based on reducing the long‐term average (normal) precipitation (1970–2000) of a first drought in the spring and a second one in summer (refer to the study of Lamarque et al. (2013)). The selected precipitation scheme for Lautaret reflects the possible scenarios according to Beniston (2006), given that Mediterranean‐influenced areas will be faced with more extensive droughts throughout the vegetation period (refer to the study of Benot et al. (2013a)). At Stubai, the daily precipitation sum of the long‐term average (normal) from the years 1990–2010 was reduced in relative numbers to match the total precipitation in the year 2003 (e.g. a very dry summer). According to the studies of Strauss et al. (2013), Beniston (2006), and Beniston (2012), more frequent summer droughts, as in the year 2003 (ZAMG, 2014), are the most likely scenario in the coming 30 to 50 years. Selected precipitation regimes (e.g. normal and dry) were assumed to be homogenously distributed throughout each site. The modelling period was from May to September, covering the growing seasons at both sites. Thirty days prior to the modelled period were included as a ‘model wind up’ period to establish realistic soil water conditions.
Table 2.
Monthly and seasonal (vegetation season) precipitation [mm] for the normal and dry scenario at Lautaret and the Stubai Valley.
| Precipitation [mm] | Lautaret | Lautaret | Stubai Valley | Stubai Valley |
|---|---|---|---|---|
| normal | dry | normal | dry | |
| May | 65.9 | 0.0 | 89.0 | 40.8 |
| June | 73.4 | 36.7 | 130.0 | 52.9 |
| July | 62.0 | 0.0 | 154.3 | 43.0 |
| August | 76.4 | 38.2 | 152.3 | 40.6 |
| September | 77.4 | 77.4 | 106.0 | 50.0 |
| Sum | 355.1 | 152.3 | 631.6 | 227.3 |
For model calibration at Lautaret, soil moisture data at a depth of 0.1 m (sensor type: Echo‐probe, Decagon Devices Inc., USA) was used. For Stubai, model calibration was performed using soil moisture data at depths of 0.05 m and 0.15 m (sensor type: ThetaProbe, Delta‐T Devices Ltd., UK). The calibration period was set from the beginning of May until the end of September, including a ‘model wind up’ period of 1 month, covering the moist conditions after the snow melt as well as droughts during the summer months. Thus, plausible parameter estimation over the entire realistic soil moisture range can be ensured (Gan et al., 1997). To calibrate the model, a least squares optimization was applied. In addition to standard regression parameters, model evaluation statistics for calibrated grassland types included Nash–Sutcliffe efficiency (NSE), percent BIAS, root mean square error (RMSE) and the RMSE observations standard deviation ratio (RSR). A detailed description of each statistical parameter is given by Moriasi et al. (2007). To evaluate the goodness of fit, the NSE of each calibration was classified according to the scheme from the study of Moriasi et al. (2007) (0.75 < NSE ≤ 1.00 very good; 0.65 < NSE < 0.75 good; 0.50 < NSE < 0.65 satisfactory; NSE ≤ 0.50 unsatisfactory).
For grasslands without available soil moisture data, soil parameters were taken from the calibrated grassland type, which was most likely representative, based on available soil data: L3(calibrated) = L9; S2(calibrated) = S9; S8(calibrated) = S3,S4.
Results
Parameterization and calibration
The results confirmed the applicability of the calibrated models, classifying five out of eight as very good (Table 3). The only exception was L5, which had a NSE coefficient of 0.48 and underestimated the soil moisture content. However, as indicated by a RMSE of 1.67, differences from the measured soil moisture content were low.
Table 3.
Model evaluation statistics for calibrated grassland types. Statistics are standard regression parameters [slope, offset and coefficient of determination (R2)], Nash–Sutcliffe efficiency (NSE), percent BIAS (PBIAS), root mean square error (RMSE) and RMSE observations standard deviation ratio (RSR). Model performance was classified according to Moriasi et al. (2007) (+++ very good, ++ good, + satisfactory, − unsatisfactory) using NSE and RSR.
| Model evaluation statistics | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Standard regression | |||||||||
| Grassland type | Slope | Offset | R2 | NSE | PBIAS | RMSE | RSR | ||
| L1 | 0.87 | 3.81 | 0.84 | 0.81 | +++ | 1.86 | 1.61 | 0.44 | +++ |
| L2 | 0.95 | 2.01 | 0.81 | 0.79 | +++ | −0.14 | 1.46 | 0.46 | +++ |
| L3 | 1.04 | −1.07 | 0.82 | 0.76 | +++ | −1.08 | 2.24 | 0.49 | +++ |
| L4 | 0.71 | 10.42 | 0.69 | 0.54 | + | 3.52 | 2.47 | 0.68 | + |
| L5 | 0.71 | 12.08 | 0.66 | 0.48 | − | 2.19 | 1.67 | 0.72 | − |
| S2 | 0.52 | 17.32 | 0.76 | 0.66 | ++ | −2.01 | 2.55 | 0.58 | ++ |
| S6 | 0.78 | 8.23 | 0.92 | 0.89 | +++ | −1.03 | 1.29 | 0.33 | +++ |
| S8 | 0.74 | 11.38 | 0.85 | 0.84 | +++ | −0.09 | 1.47 | 0.40 | +++ |
Parameter estimates revealed the differences in physical and hydrological soil parameters between Stubai (A) and Lautaret (F) (Figure 2). Regarding soil hydrological parameters, saturated conductivity (Ks) and residual soil water content (Ɵres) showed similar values. However, saturated soil water content (Ɵsat) was slightly higher at the Lautaret site. This fact was corroborated by the calibrated Mualem–van Genuchten parameters α and n having lower values at Lautaret indicating higher clay content (Hartge & Horn, 1999). The parameter TR described the soil water tension (e.g. suction head) at which potential transpiration is reduced by plants. This process is mainly regulated by the stomatal control of the plants and is therefore highly related to species composition. In more detail, the HILLFLOW model required two different values for TR varying with different rates of potential transpiration (TR): (1) TR high for high potential transpiration rates (>5 mm d−1) and (2) TR low for low potential transpiration rates (<1 mm d−1). For both TRhigh and TRlow, the best parameter estimates indicated a reduction in the potential transpiration rate at higher soil water tension levels at Lautaret compared with Stubai. Although saturated water contents as well as clay content (as a proxy for soil water retention capacity) were higher at Lautaret, parameterization of TR high and TR low indicated that plants were closing their stomata earlier to reduce transpiration rates and to save available water in the root zone.
Figure 2.
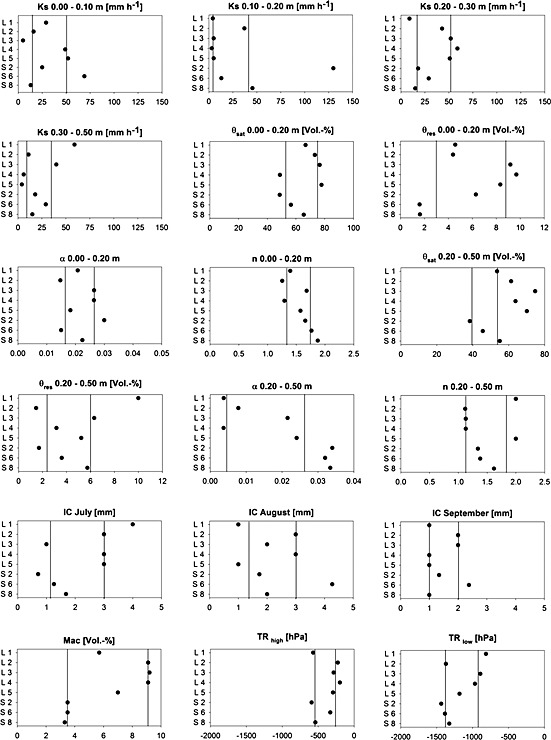
Parameter estimates from model calibration: Ks = saturated hydraulic conductivity; Ɵsat = saturated water content; Ɵres = residual soil water content; α, n = Mualem–van Genuchten parameters; IC = canopy interception (water); Mac = proportion of macropores; TR = transpiration reduction (at high/low potential rates). Black vertical lines mark the interquartile range of all grassland types.
Soil moisture
Soil moisture was modelled in the range of 0–0.3 m to reproduce the impact of drought on the main rooting depth of grassland communities (Tasser & Tappeiner, 2005). At Lautaret, grasslands with higher management intensities (L1 > L2 > L3 on terraces; L4 > L5 on unterraced grassland) were prone to lower soil moisture under both normal and dry conditions (Table 4). Unterraced grassland showed generally higher soil moisture values than terraced grassland but also exhibited the strongest reductions with drought. At Stubai, no distinct correlation between management intensity and soil moisture was observed. However, grasslands with the highest management intensities (S6, S8) were more strongly affected by drought. In contrast, pastures showed the lowest negative impact of drought (Table 4). The mean soil moisture at drought was generally lower, and the relative reduction of soil moisture by drought was generally higher at Stubai compared with Lautaret for land‐use types with similar management intensity (i.e. S6 vs L1; S4 vs L2 and L4; S3 vs L3 and L5; S8 vs L4; S9 vs L4; Table 4).
Table 4.
Modelled mean soil moisture (Vol.−%) and standard deviation for different grassland types for normal (n) and dry (d) years; difference (Δ n–d) in absolute values (Vol.−%) and relative numbers (%) at a depth of 0–0.3 m.
| Site‐ID | Description | Normal (n) year | Dry (d) year | Δ n–d (%) |
|---|---|---|---|---|
| L1 | Previously cultivated terraces (1550–1950 m), now manured and mown | 33.63 (±1.4) | 26.01 (±0.6) | −7.62 (−22.7%) |
| L2 | Previously cultivated terraces (1550–1950 m), now mown but not manured | 35.76 (±1.8) | 26.01 (±1) | −9.75 (−27.3%) |
| L3 | Previously cultivated terraces (1550–1950 m), now unmown and grazed in spring and autumn | 36.56 (±1.8) | 26.22 (±1.3) | −10.34 (−28.3%) |
| L4 | Never cultivated unterraced grasslands (1700–2000 m), currently mown | 39.49 (±1.3) | 28.28 (±1) | −11.21 (−28.4%) |
| L5 | Never cultivated unterraced grasslands (1700–2000 m), summer grazed | 43.21 (±2.0) | 27.48 (±1.9) | −15.73 (−36.4%) |
| L9 | Steep and rocky slopes, grazed | 37.75 (±2) | 26.91 (±1.9) | −10.84 (−28.7%) |
| S2 | Abandoned land | 33.03 (±0.6) | 20.27 (±1.3) | −12.76 (−38.6%) |
| S4 | Meadow of low land‐use intensity (mowing every 2 years) | 33.00 (±1.4) | 21.34 (±2.4) | −11.66 (−35.3%) |
| S6 | Meadow of high land‐use intensity (1–2 mowings per year) | 35.04 (±1.3) | 20.19 (±1.3) | −14.85 (−42.4%) |
| S3 | Pasture | 29.58 (±1.6) | 21.85 (±1.1) | −7.73 (−26.1%) |
| S8 | Meadow/pasture | 39.42 (±2.6) | 22.35 (±1.1) | −17.07 (−43.3%) |
| S9 | Larch meadow | 32.92 (±0.4) | 20.55 (±1.1) | −12.37 (−37.6%) |
The soil moisture patterns in Figure 3 are a graphic representation of the results presented in Table 4. Regarding the differences across the grassland types under current (i.e. normal) conditions, greenish areas mark the mean seasonal soil moisture close to field capacity (cf., Table 1, Table 4). At both sites, this category was attributed to grasslands with low management intensity (i.e. unterraced, no fertilisation, not heavily grazed, max. 1 cut per year). On the other hand, grasslands of higher land‐use intensity (i.e. terraced grassland at Lautaret and meadows of high land‐use intensities as well as larch meadows at Stubai) revealed the mean soil moisture below field capacity, indicated in yellow and orange. As the soil types present (Dystric Cambisol, Cambisol) did not vary significantly regarding soil hydrological properties (Table 1), soil moisture is reduced by varying the biotic responses (i.e. ET). Even under ‘normal’ conditions, soil moisture at the most productive grassland sites was below its optimum for plant growth. Pastures (S3) at Stubai showed the lowest mean soil moisture values caused by simultaneously high ET and low management intensity (i.e. low biomass removal) – a unique combination compared with other grassland types.
Figure 3.
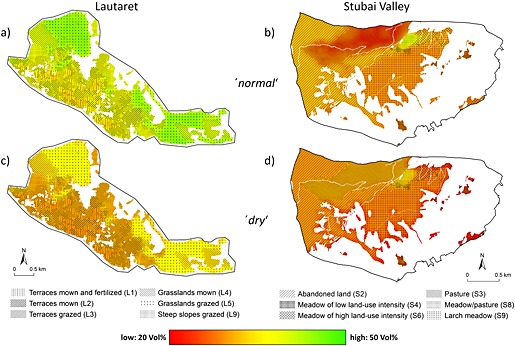
Spatial soil moisture distribution at a depth of 0–0.3 m under ‘normal’ (a, b) and ‘dry’(c, d) conditions at Lautaret and Stubai.
Under ‘dry’ conditions, both study sites showed clearly reduced soil moisture for each grassland type, but the effect was smaller at Lautaret. Current differences between grasslands were almost levelled out by drought effects at Stubai. At Lautaret, unterraced grassland types (L4, L5, L9) showed clearly higher soil moisture values compared with terraced grassland of higher land‐use intensities. Generally, spatial heterogeneity of soil moisture was reduced under ‘dry’ conditions.
Water quantity
The following results refer to the totals for the modelling period May to September, unless otherwise specified. In absolute numbers, under ‘normal’ conditions, ET made up 234 mm and 366 mm at Lautaret and Stubai, respectively. Under ‘dry’ conditions, ET was reduced to 122 mm and 202 mm at Lautaret and Stubai, respectively. Water quantity [i.e. deep seepage (DS)] was evaluated at the catchment scale and resulted from the HILLFLOW model output. Relating DS to precipitation (P) and ET to P showed the differences in the water balance between the drier site, Lautaret, and the more humid site, Stubai. The model estimated that under ‘normal’ conditions, 31% (110 mm) and 39% (246 mm) of precipitation drained as DS water (Figure 4) at Lautaret and Stubai, respectively. Under ‘dry’ conditions, DS was reduced to 19 mm and 26 mm at Lautaret and Stubai, respectively. While higher DS/P values at Stubai indicate more humid conditions than at Lautaret, higher ET/P values indicate drier conditions, which were only observed for Lautaret under ‘normal’ conditions. Under ‘dry’ conditions, ET/P ratios at Stubai were slightly higher, indicating a ‘water spending’ strategy.
Figure 4.
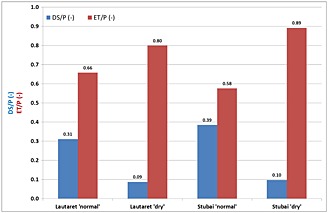
Relating deep seepage and evapotranspiration to precipitation for evaluating water balance components under ‘normal’ and ‘dry’ conditions at Lautaret and Stubai.
Discussion
Assessing the water provision service for alpine grasslands at two climatically different regions across the Alps provided evidence that, under drought conditions, ET is influenced not only by abiotic factors (i.e. soil and climate) but also by the water‐use strategy of the vegetation present. This influence was obvious at the drier site, Lautaret, where the mean soil moisture under drought conditions was higher than at the more humid site, Stubai. Given soil depths of at least 0.3 m (lowest on terraced grassland, Lautaret) (Robson et al., 2007; Leitinger et al., 2010), thereby not limiting the main rooting depth of the investigated plant communities (e.g. Tasser and Tappeiner (2005)), it must be noted that the depletion of soil moisture mainly depended on potential ET rates defined for every existing vegetation type. To overcome a possible bias by overestimating ET, our parameterization and calibration procedure covered wet and dry periods as well as all parameters that trigger ET reduction due to lower soil moisture and general climatic conditions (i.e. clear vs overcast skies). The amount of soil moisture depletion during the modelled drought periods was within the range covered by the period of model calibration, and the results are therefore considered highly reliable. The performance of the model was at least satisfactory for the seven out of eight investigated grassland types (Table 3), while only L5 was not calibrated satisfactorily. This finding was attributed to the high litter accumulation in this grassland type (Gross et al., 2007), which affected the soil moisture in two counteracting ways that are difficult to quantify. Litter layers may both reduce soil evaporation and increase interception of rainfall (Gross et al. 2008). Nevertheless, given the RMSE of 1.67 Vol% (RMSE observations standard deviation ratio of 0.72, Table 3), the calibration of L5 can still be considered suitable for modelling the soil moisture dynamics on a daily basis.
In general, modelled soil moisture was consistent with the observations from the study of Gross et al. (2008) in 12 plots at 0–15 cm depth for Lautaret in the ‘normal’ precipitation scheme, whereas the highest soil moisture throughout the growing season was observed on unterraced grasslands (L4, L5) and the driest soils were on mown terraces (L1, L2), which are shallower and rockier (Robson et al., 2007). At Stubai, Leitinger et al. (2010) found the mean soil moisture values at a depth of 0–0.2 m for the period from 23 May 2007 to 02 October 2007 (752 mm of precipitation) of 39.44°Vol% (±6.6) and 33.62°Vol% (±9.2) for abandoned land (S2) and pasture (S3), respectively. Given the lower soil depth of observation and higher precipitation (i.e. 752 mm compared with the long‐term average and model input of 631 mm between May and September), these results can be considered comparable to the modelled mean soil moisture from this study (cf., Table 4); this is particularly true with regard to the generally higher mean soil moisture at abandoned lands compared with pastures. Surprisingly, under ‘normal’ conditions, no clear differences in soil moisture patterns between the two sites were found. This result suggests that, while the mean soil moisture throughout the season did not differ significantly between the two sites, the stronger soil moisture dynamics reported by Obojes et al. (2014) at the drier Lautaret site promoted the establishment of plant communities adapted to generally drier conditions, as shown by lower Ellenberg moisture (F) values.
Drought impact on soil moisture
Under ‘dry’ conditions, water shortage leads to lower stomatal conductance and reduced photosynthesis, as well as reduced biomass production (Chaves et al., 2002). Regarding the overall effects of drought at each site, Lautaret revealed higher soil moisture values than Stubai even though Lautaret is characterized by a drier climate where drought would be expected to have a stronger effect on soil moisture. In other words, the depletion of soil moisture under dry conditions was lower at Lautaret than at Stubai, which indicates different plant feedbacks driven by different climatic conditions (i.e. either precipitation amount or frequency). In general, drought leads to a more uniform soil moisture distribution in both catchments (Figure 3), which reflects a decoupling from the influence of vegetation given by soil moisture conditions close to the permanent wilting point (PWP).
The impact of drought at Lautaret was decreased soil moisture, ranging from −22.7% to −36.4% (Table 4), which is in line with the findings from the study of Benot et al. (2013b) who observed a decrease in the mean soil moisture of between −28% and −32% in a subalpine grassland site at Lautaret during a simulated extreme drought in July and August (−80% precipitation). The remaining soil moisture was above the PWP (cf., Table 1 and Table 4), indicating that plant communities were following a ‘water saving’ strategy to avoid structural or physiological damage. This is corroborated by the findings from the study of Benot et al. (2013a) at Lautaret, where community structure of a subalpine grassland was not affected after 2 years of consecutive summer drought due to well‐adapted plant communities. On the contrary, at Stubai, soil moisture decreased between −26.3% and −43.3%, reaching the PWP for most grassland types (cf., Table 1 and Table 4). Schmitt et al. (2010) and Brilli et al. (2011) report that, for differently managed grassland ecosystems at Stubai, low soil moisture did not lead to strong reductions in the net ecosystem CO2 and H2O exchange. Experiments conducted by Brilli et al. (2011) confirmed that at least the dominant grassland plant species were not sensitive to long drought periods until very low soil moisture was reached (<0.10 m3 m‐3).
Plant communities at Stubai more efficiently deplete soil moisture by acting as ‘water spending’ plants (Levitt, 1980; Moreno‐Gutierrez et al., 2012), which is reflected by higher estimates for the parameters TR low and higher TR high (Figure 2) at Lautaret compared with Stubai. This means that, at Lautaret, plants reduce ET earlier than at Stubai by closing their stomata (Lavorel, unpublished observation). With respect to drought periods, it can be concluded that drought would affect the soil moisture of grassland ecosystems at Stubai more strongly than at Lautaret unless physiological adaptations or shifts in plant communities are taking place. In regard to that, it remains debatable whether plants can initially adapt to drought physiologically by changing their water‐use strategy from ‘water spending’ to ‘water saving’ until such time as a shift to a more adapted plant community takes place (Jentsch et al., 2011; Reyer et al., 2013).
Drought impact on water quantity
In the catchment water balance, DS is more strongly reduced by drought at Stubai than at Lautaret in both relative and absolute numbers (Figure 4). This study therefore confirms that the observed ‘water spending’ vegetation strategy at Stubai may have negative consequences for downstream water users (Brilli et al., 2011), i.e. reduced water quantity. At Lautaret, Obojes et al. (2014) found slightly higher precipitation rates of 2.75 mm d−1 compared with 2.3 mm d−1 in our study, as well as a DS values of 0.77 mm d−1 (±0.22) and ET rates of 2.07 mm d−1 (±0.24) under ‘normal’ conditions. Given the modelling period of 153 days, the findings from our study correspond to approximately 0.7 mm d−1 and 1.5 mm d−1 of DS and ET, respectively. At Stubai, a DS value of 1.6 mm d−1 and ET value of 2.4 mm d−1 at P = 4.1 mm d−1 were found. Obojes et al. (2014) report a DS of 3.04 mm d−1 (±0.39) and ET of 2.15 mm d−1 (±0.32), which is again realistic given the higher P rates of 5.17 mm d−1 (±0.21) leading to more DS, in contrast with the higher ET in a non‐water limited grassland ecosystem (Wieser et al., 2008). ET values under ‘normal’ conditions at Stubai also correspond to the results from the study Wieser et al. (2008) who compared the experimental data from 16 grassland sites in the Austrian Alps between 580 and 2550 m a.s.l. and estimated the mean ET rates of 2.2 mm d−1. They further stated that during 2001–2006, annual ET/P values ranged from 0.53 (2002) to 0.91 (2006), while the fraction of precipitation evaporating to the atmosphere increased with decreasing precipitation. Wieser et al. (2008) state, in this context, that even during droughts in which up to 90% of P evaporates, these systems are not faced with water stress. For the investigated grassland sites, they also specified that the limitations of ET by closing stomata due to low soil moisture played a minor role, which is again in line with our findings concerning ‘dry’ conditions at Stubai.
Della Chiesa et al. (2014) evaluated the changes in grassland hydrological cycling along an elevation gradient in the Alps. Their results indicated the water stress conditions for vegetation in each year at lower altitude (1000 m a.s.l.) with a mean annual temperature of 8.1–9.5 °C and annual precipitation of 580 to 636 mm. At medium altitudes (1500 m a.s.l., mean temperature of 5.3–7.1 °C, precipitation of 632–686 mm), only a warmer and drier year caused drought, whereas no water stress was observed at highest altitude (2000 m a.s.l., mean temperature of 2.5–3.5 °C, precipitation of 620–706 mm). While the annual precipitation in the study of Della Chiesa et al. (2014) was lower than for both sites in this study, temperatures and according ET were comparable. Given the general climatic conditions at Lautaret and Stubai, the results from Della Chiesa et al. (2014) support the findings from our modelling study; that is, under ‘normal’ conditions, generally no drought effect due to low soil moisture occurred. They also modelled the effects of a precipitation change of 30% on ET and showed a slight reduction in ET at middle elevations and no change at high elevation. This corroborates our findings that well‐adapted plant communities can easily cope with the reductions in precipitation without strongly affecting ET.
The ET/P ratios of 0.66 at Lautaret and 0.58 at Stubai under ‘normal’ conditions were within the range of values reported by Wieser et al. (2008), collected for 16 grassland sites. They reported decreasing ET/P values from 0.7 to 0.1 with increasing P and higher ET/P values in the drier inner Alpine region compared with the more humid parts of the Alps. Nevertheless, the modelled soil moisture gave no hint that water limitation took place at both sites. This fact is in line with findings from the study of Everson (2001) who observed ET/P values of 0.44–0.56 in wetter and 0.64–0.69 in drier years in montane grasslands of South Africa that they classified as being not limited by soil moisture. In comparison, Gu et al. (2008) found ET/P values of 0.6 at a meadow on the Qinghai–Tibetan Plateau that was then classified as water‐limited. The modelled ET/P values in this study under ‘dry’ conditions revealed higher ET/P values for the Stubai site (0.89), indicating stronger water limitation than at Lautaret (0.80). At Stubai, at least the dominant plant species act as ‘water spending’ plants, maintaining maximum transpiration rates despite strong reductions in soil moisture. Again, we would like to note that Brilli et al. (2011) showed in laboratory experiments that plant species at Stubai did not regulate stomatal conductance until soil moisture reached the PWP. Our study therefore confirms that the impacts of drought on water provision services can only be evaluated by taking water‐use strategies of the dominant plant species into account. This fact will gain additional importance in a future climate where increasing air temperatures foster higher evapotranspiration rates and the main factor for runoff formation changes throughout ecosystems (Liu et al., 2014; Olang et al., 2014).
Ecosystem service ‘water provision’
Based on the study of Lamarque et al. (2011b) on stakeholder perceptions of grassland ESs, eight ESs were considered to be of high importance for local farmers at the two sites, Lautaret and Stubai: (1) soil stability, (2) soil fertility, (3) water provision, (4) water quality, (5) forage quality, (6) forage quantity, (7) aesthetic value, and (8) carbon storage. In the following, we discuss the impacts of changes in water provision (i.e. soil moisture and water quantity) on the other relevant ESs with a special focus on the water‐related services.
In the short term, soil fertility will decrease, and water quality will increase as drought conditions increase soil mineral nitrogen and nitrogen mineralization and reduce leaching losses (Bloor & Bardgett, 2012). The most obvious impacts are implications for plant growth and tissue quality, leading to changes in forage quality and forage quantity. Forage quality would be slightly reduced because of lower tissue water and nitrogen content (Quetier et al., 2007; Benot et al., 2013a). Forage quantity would be lower because of the significantly lower evapotranspiration of plants (De Boeck et al., 2011; Schirpke et al., 2013a).
Longer‐term effects are associated with species turnover towards more parsimonious water management strategies. Drought will increase the root/shoot ratio (Chaves et al., 2002) and consequently soil stability by increasing rooting density (Tasser et al., 2003). Additionally, drought leads to an increasing abundance of shrubs or tussock grasses with a ‘water saving’ strategy, which significantly reduces snow gliding and related erosion risk because of the growth form (Tasser et al., 2003; Leitinger et al., 2008). Forage quality would be further reduced because of a decline of forbs and increase in shrubs or grasses (Harte et al., 2006). Management decisions due to recurrent droughts and accompanied disadvantages for agronomy would lead to a decrease in the aesthetic value due to abandonment (Schirpke et al., 2013b). Carbon storage might be reduced because of reduced vegetation productivity. However, lagged and legacy effects as well as other mechanisms of drought are complex and intertwined and act synergistically or antagonistically (Reichstein et al., 2013). Additionally, our evaluation of impacts of drought on ESs is based on existing farming systems. However, studies already report an adaptation to climate change rather than a stop of farm activity (Bindi and Olesen, 2011; Lamarque et al., 2013; Varadan and Kumar, 2014), and promising techniques to design adaptation strategies at farming level have developed (Schaap et al., 2013, among others). Hence, the impact of drought on a specific ES or even bundled ES cannot be ascertained with certainty nor is it generalizable for managed alpine grasslands. However, by taking the results of this study in two climatically different regions of the Alps into account, the impacts of drought on numerous ES can be determined as long as preconditions concerning the ‘water‐use’ strategy of existing plant communities are taken into account.
Conclusions
Our modelling analysis of the impact of droughts on the supporting ES water provision in sub‐watersheds dominated by managed alpine grasslands in two climatically different regions of the Alps revealed common responses of decreasing soil moisture and DS. However, these reductions were lower in the generally drier French region because of a stronger ‘water saving’ strategy of plant communities. The expected impacts of drought on water provision services can only be evaluated by considering water‐use strategies of the dominant plant species. Thus, whether more frequent droughts will lead to a shift to better adapted plant communities at the more humid Austrian region remains debatable and depends on whether and to which extent plants can initially adapt to drought physiologically by changing their water‐use strategy.
Acknowledgements
We wish to thank Axel Bronstert for allocating the HILLFLOW model, Gerhard Niederwieser and Sabine Kreidl from the IT‐Centre (University of Innsbruck) for their continuing technical support and Erich Tasser (EURAC) for providing data. This research was funded by the ERA‐Net BiodivERsA project ‘VITAL’, with the national funders Austrian Science Fund (FWF: I 242‐B17) and ANR within the 2008 BiodivERsA call for research proposals. RR was partly funded by the research funding price of the Hypo Bank Tirol (2013). This research was conducted on the long‐term research site ‘Zone Atelier Alpes’, a member of the ILTER‐Europe and LTSER networks, and the LTER site ‘Stubai Valley’, a member of the Austrian LTSER Platform ‘Tyrolean Alps’. UIBK is part of the Interdisciplinary Research Center ‘Ecology of the Alpine Region’ within the major research area ‘Alpine Space – Man and Environment’ at the University of Innsbruck. Modelling was performed within the ‘High‐Performance Computing (HPC)’ Research Center part of the ‘Scientific Computing’ major research area at the University of Innsbruck.
Leitinger, G. , Ruggenthaler, R. , Hammerle, A. , Lavorel, S. , Schirpke, U. , Clement, J.‐C. , Lamarque, P. , Obojes, N. , and Tappeiner, U. (2015) Impact of droughts on water provision in managed alpine grasslands in two climatically different regions of the Alps. Ecohydrol., 8: 1600–1613. doi: 10.1002/eco.1607.
References
- Bahn M, Reichstein M, Dukes JS, Smith MD, McDowell NG. 2014. Climate‐biosphere interactions in a more extreme world. The New Phytologist 202: 356–359. DOI: 10.1111/nph.12662. [DOI] [PubMed] [Google Scholar]
- Bangash RF, Passuello A, Sanchez‐Canales M, Terrado M, Lopez A, Elorza FJ, Ziv G, Acuna V, Schuhmacher M. 2013. Ecosystem services in Mediterranean river basin: climate change impact on water provisioning and erosion control. Science of the Total Environment 458: 246–255. DOI: 10.1016/j.scitotenv.2013.04.025. [DOI] [PubMed] [Google Scholar]
- Beniston M. 2006. Mountain weather and climate: a general overview and a focus on climatic change in the Alps. Hydrobiologia 562: 3–16. DOI: 10.1007/s10750-005-1802-0. [Google Scholar]
- Beniston M. 2012. Impacts of climatic change on water and associated economic activities in the Swiss Alps. Journal of Hydrology 412: 291‐296 DOI: 10.1016/j.jhydrol.2010.06.046. [Google Scholar]
- Beniston M, Stephenson DB, Christensen OB, Ferro CAT, Frei C, Goyette S, Halsnaes K, Holt T, Jylha K, Koffi B, Palutikof J, Scholl R, Semmler T, Woth K. 2007. Future extreme events in European climate: an exploration of regional climate model projections. Climatic Change 81: 71–95. DOI: 10.1007/s10584-006-9226-z. [Google Scholar]
- Benot ML, Saccone P, Pautrat E, Vicente R, Colace M‐P, Grigulis K, Clément J‐C, Lavorel S. 2013a. Stronger short‐term effects of mowing than extreme summer weather on a subalpine grassland. Ecosystems 1‐15 DOI: 10.1007/s10021-013-9734-4. [Google Scholar]
- Benot ML, Saccone P, Vicente R, Pautrat E, Morvan‐Bertrand A, Decau ML, Grigulis K, Prud'homme MP, Lavorel S. 2013b. Subalpine grasslands. Plant Ecology Divers 6: 393–404. DOI: 10.1080/17550874.2013.784818. [Google Scholar]
- Bindi M, Olesen JE. 2011. The responses of agriculture in Europe to climate change. Regional Environmental Change 11: S151–S158. DOI: 10.1007/s10113-010-0173-x. [Google Scholar]
- Bloor JMG, Bardgett RD. 2012. Stability of above‐ground and below‐ground processes to extreme drought in model grassland ecosystems: interactions with plant species diversity and soil nitrogen availability. Perspectives in Plant Ecology 14: 193–204. DOI: 10.1016/j.ppees.2011.12.001. [Google Scholar]
- Brilli F, Hortnagl L, Hammerle A, Haslwanter A, Hansel A, Loreto F, Wohlfahrt G. 2011. Leaf and ecosystem response to soil water availability in mountain grasslands. Agricultural and Forest Meteorology 151: 1731–1740. DOI: 10.1016/j.agrformet.2011.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bronstert A. 1995. User manual for the HILLFLOW‐3D catchment modelling system, Version 1.1 (English Edition). CRC‐Working Document 95/4, Cooperative Research Centre for Catchment Hydrology and CSIRO Division of Water Resources: Canberra. [Google Scholar]
- Bronstert A. 1999. Capabilities and limitations of detailed hillslope hydrological modelling. Hydrological Processes 13: 21–48. DOI: 10.1002/(Sici)1099-1085(199901)13:1<21::Aid-Hyp702>3.3.Co;2-W. [Google Scholar]
- Bronstert A, Bardossy A. 1999. The role of spatial variability of soil moisture for modelling surface runoff generation at the small catchment scale. Hydrology Earth System Science 3: 505–516. [Google Scholar]
- Bronstert A, Plate EJ. 1997. Modelling of runoff generation and soil moisture dynamics for hillslopes and micro‐catchments. Journal of Hydrology 198: 177–195. [Google Scholar]
- Burkhard B, Kroll F, Nedkov S, Muller F. 2012. Mapping ecosystem service supply, demand and budgets. Ecological Indicators 21: 17–29. DOI: 10.1016/j.ecolind.2011.06.019. [Google Scholar]
- Cerda A, Schnabel S, Ceballos A, Gomez‐Amelia D. 1998. Soil hydrological response under simulated rainfall in the Dehesa land system (extremadura, SW Spain) under drought conditions. Earth Surface Proceedings of the Land 23: 195–209. DOI: 10.1002/(Sici)1096-9837(199803)23:3<195::Aid-Esp830>3.0.Co;2-I. [Google Scholar]
- Chaves MM, Pereira JS, Maroco J, Rodrigues ML, Ricardo CPP, Osorio ML, Carvalho I, Faria T, Pinheiro C. 2002. How plants cope with water stress in the field. Photosynthesis and growth. Annals Botany‐London 89: 907–916. DOI: 10.1093/Aob/Mcf105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crossman J, Futter MN, Oni SK, Whitehead PG, Jin L, Butterfield D, Baulch HM, Dillon PJ. 2013. Impacts of climate change on hydrology and water quality: future proofing management strategies in the Lake Simcoe watershed, Canada. Journal of Great Lakes Research 39: 19–32. DOI: 10.1016/j.jglr.2012.11.003. [Google Scholar]
- Crossman ND, Burkhard B, Nedkov S. 2012. Quantifying and mapping ecosystem services. International Journal of Biodiversity Science, Ecosystem Services & Management 8: 1–4. DOI: 10.1080/21513732.2012.695229. [Google Scholar]
- De Boeck HJ, Dreesen FE, Janssens IA, Nijs I. 2011. Whole‐system responses of experimental plant communities to climate extremes imposed in different seasons. New Phytologist 189: 806–817. DOI: 10.1111/j.1469-8137.2010.03515.x. [DOI] [PubMed] [Google Scholar]
- Della Chiesa S, Bertoldi G, Niedrist G, Obojes N, Endrizzi S, Albertson JD, Wohlfahrt G, Hörtnagl L, Tappeiner U. 2014. Modelling changes in grassland hydrological cycling along an elevational gradient in the Alps. Ecohydrology 7: 1453–1473. DOI: 10.1002/eco.1471. [Google Scholar]
- Egoh B, Reyers B, Rouget M, Richardson DM, Le Maitre DC, van Jaarsveld AS. 2008. Mapping ecosystem services for planning and management. Agriculture, Ecosystems & Environment 127: 135‐140. DOI: 10.1016/j.agee.2008.03.013. [Google Scholar]
- Eigenbrod F, Armsworth PR, Anderson BJ, Heinemeyer A, Gillings S, Roy DB, Thomas CD, Gaston KJ. 2010. The impact of proxy‐based methods on mapping the distribution of ecosystem services. Journal of Applied Ecology 47: 377‐385. DOI: 10.1111/j.1365-2664.2010.01777.x. [Google Scholar]
- Everson CS. 2001. The water balance of a first order catchment in the montane grasslands of South Africa. Journal of Hydrology 241: 110‐123. DOI: 10.1016/S0022-1694(00)00376-0. [Google Scholar]
- Gan TY, Dlamini EM, Biftu GF. 1997. Effects of model complexity and structure, data quality, and objective functions on hydrologic modeling. Journal of Hydrology 192: 81–103. [Google Scholar]
- Gross N, Robson TM, Lavorel S, Albert C, Le Bagousse‐Pinguet Y, Guillemin R. 2008. Plant response traits mediate the effects of subalpine grasslands on soil moisture. The New Phytologist 180: 652–662. DOI: 10.1111/j.1469-8137.2008.02577.x. [DOI] [PubMed] [Google Scholar]
- Gross N, Suding KN, Lavorel S. 2007. Leaf dry matter content and lateral spread predict response to land use change for six subalpine grassland species. Journal of Vegetation Science 18: 289–300. DOI: 10.1111/j.1654-1103.2007.tb02540.x. [Google Scholar]
- Gu S, Tang YH, Cui XY, Du M, Zhao L, Li Y, Xu SX, Zhou H, Kato T, Qi PT, Zhao X. 2008. Characterizing evapotranspiration over a meadow ecosystem on the Qinghai‐Tibetan Plateau. Journal of Geophysical Research‐Atmospheres 113 DOI: 10.1029/2007jd009173. [Google Scholar]
- Harte J, Saleska S, Shih T. 2006. Shifts in plant dominance control carbon‐cycle responses to experimental warming and widespread drought. Environmental Research Letters 1 DOI: 10.1088/1748-9326/1/1/014001. [Google Scholar]
- Hartge KH, Horn R. 1999. Einführung in die Bodenphysik, Schweizerbart'Sche Verlagsbuchhandlung: Stuttgart. [Google Scholar]
- Jentsch A, Kreyling J, Elmer M, Gellesch E, Glaser B, Grant K, Hein R, Lara M, Mirzae H, Nadler SE, Nagy L, Otieno D, Pritsch K, Rascher U, Schadler M, Schloter M, Singh BK, Stadler J, Walter J, Wellstein C, Wollecke J, Beierkuhnlein C. 2011. Climate extremes initiate ecosystem‐regulating functions while maintaining productivity. Journal of Ecology 99: 689‐702. DOI: 10.1111/j.1365-2745.2011.01817.x. [Google Scholar]
- Köplin N, Rößler O, Schädler B, Weingartner R. 2014. Robust estimates of climate‐induced hydrological change in a temperate mountainous region. Climatic Change 122: 171–184. DOI: 10.1007/s10584-013-1015-x. [Google Scholar]
- Lamarque P, Lavorel S, Mouchet M, Quétier F. 2014. Plant trait‐based models identify direct and indirect effects of climate change on bundles of grassland ecosystem services. Proceedings of the National Academy Sciences USA, in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lamarque P, Artaux A, Barnaud C, Dobremez L, Nettier B, Lavorel S. 2013. Taking into account farmers' decision making to map fine‐scale land management adaptation to climate and socio‐economic scenarios. Landscape Urban Plan 119: 147‐157. DOI: 10.1016/j.landurbplan.2013.07.012. [Google Scholar]
- Lamarque P, Quetier F, Lavorel S. 2011a. The diversity of the ecosystem services concept and its implications for their assessment and management. Cr Biologies 334: 441‐449. DOI: 10.1016/j.crvi.2010.11.007. [DOI] [PubMed] [Google Scholar]
- Lamarque P, Tappeiner U, Turner C, Steinbacher M, Bardgett RD, Szukics U, Schermer M, Lavorel S. 2011b. Stakeholder perceptions of grassland ecosystem services in relation to knowledge on soil fertility and biodiversity. Regional Environmental Change 11: 791–804. DOI: 10.1007/s10113-011-0214-0. [Google Scholar]
- Lavorel S, Spiegelberger T, Mauz I, Bigot S, Granjou C, Dobremez L, Nettier B, Thuiller W, Brun J‐J, Cozic P. 2013. Fostering research into coupled long‐term dynamics of climate, land use, ecosystems and ecosystem services in the central French Alps In Singh SJ, Haberl H, Chertow M, Mirtl M, Schmid M. (eds) Long Term Socio‐Ecological Research. Springer: Netherlands; 485–501. [Google Scholar]
- Leitinger G, Holler P, Tasser E, Walde J, Tappeiner U. 2008. Development and validation of a spatial snow‐glide model. Ecological Modelling 211: 363–374. DOI: 10.1016/j.ecolmodel.2007.09.015. [Google Scholar]
- Leitinger G, Tasser E, Newesely C, Obojes N, Tappeiner U. 2010. Seasonal dynamics of surface runoff in mountain grassland ecosystems differing in land use. Journal of Hydrology 385: 95–104. DOI: 10.1016/j.jhydrol.2010.02.006. [Google Scholar]
- Levitt J. 1980. Responses of Plants to Environmental Stresses, Academic Press Inc.: New York. [Google Scholar]
- Liu S, Crossman ND, Nolan M, Ghirmay H. 2013. Bringing ecosystem services into integrated water resources management. Journal of Environmental Management 129: 92‐102. DOI: 10.1016/j.jenvman.2013.06.047. [DOI] [PubMed] [Google Scholar]
- Liu Z, Yao Z, Huang H, Wu S, Liu G. 2014. Land use and climate changes and their impacts on runoff in the Yarlung Zangbo river basin, China. Land Degradation and Development 25: 203–215. DOI: 10.1002/ldr.1159. [Google Scholar]
- Martínez‐Garza C, Tobon W, Campo J, Howe HF. 2013. Drought mortality of tree seedlings in an eroded tropical pasture. Land Degradation and Development 24: 287–295. DOI: 10.1002/ldr.1127. [Google Scholar]
- Martínez‐Harms MJ, Balvanera P. 2012. Methods for mapping ecosystem service supply: a review. International Journal of Biodiversity Science, Ecosystem Services & Management 8: 17–25. DOI: 10.1080/21513732.2012.663792. [Google Scholar]
- Messerli B, Viviroli D, Weingartner R. 2004. Mountains of the world: vulnerable water towers for the 21(st) century. Ambio Special Report 13: 29–34. [PubMed] [Google Scholar]
- Meybeck M, Green P, Vorosmarty C. 2001. A new typology for mountains and other relief classes: an application to global continental water resources and population distribution. Mountain Research and Development 21: 34‐45. DOI: 10.1659/0276-4741(2001)021[0034:Antfma]2.0.Co;2. [Google Scholar]
- Moreno‐Gutierrez C, Dawson TE, Nicolas E, Querejeta JI. 2012. Isotopes reveal contrasting water use strategies among coexisting plant species in a Mediterranean ecosystem. New Phytologist 196: 489–496. DOI: 10.1111/j.1469-8137.2012.04276.x. [DOI] [PubMed] [Google Scholar]
- Moriasi DN, Arnold JG, Van Liew MW, Bingner RL, Harmel RD, Veith TL. 2007. Model evaluation guidelines for systematic quantification of accuracy in watershed simulations. Transactions of the Asabe 50: 885–900. [Google Scholar]
- Nemec KT, Raudsepp‐Hearne C. 2013. The use of geographic information systems to map and assess ecosystem services. Biodiversity and Conservation 22: 1–15. DOI: 10.1007/s10531-012-0406-z. [Google Scholar]
- Obojes N, Bahn M, Tasser E, Walde J, Inauen N, Hiltbrunner E, Saccone P, Lochet J, Clément JC, Lavorel S, Tappeiner U, Körner Ch. 2014. Vegetation effects on the water balance of mountain grasslands depend on climatic conditions. Ecohydrology. DOI: 10.1002/eco.1524. [Google Scholar]
- Olang LO, Kundu PM, Ouma G, Fürst J. 2014. Impacts of land cover change scenarios on storm runoff generation: a basis for management of the Nyando Basin, Kenya. Land Degradation and Development 25: 267–277. DOI: 10.1002/ldr.2140. [Google Scholar]
- Quetier F, Thebault A, Lavorel S. 2007. Plant traits in a state and transition framework as markers of ecosystem response to land‐use change. Ecological Monographs 77: 33–52. DOI: 10.1890/06-0054. [Google Scholar]
- Reichstein M, Bahn M, Ciais P, Frank D, Mahecha MD, Seneviratne SI, Zscheischler J, Beer C, Buchmann N, Frank DC, Papale D, Rammig A, Smith P, Thonicke K, van der Velde M, Vicca S, Walz A, Wattenbach M. 2013. Climate extremes and the carbon cycle. Nature 500: 287‐295. DOI: 10.1038/Nature12350. [DOI] [PubMed] [Google Scholar]
- Reyer CPO, Leuzinger S, Rammig A, Wolf A, Bartholomeus RP, Bonfante A, de Lorenzi F, Dury M, Gloning P, Abou Jaoude R, Klein T, Kuster TM, Martins M, Niedrist G, Riccardi M, Wohlfahrt G, de Angelis P, de Dato G, Francois L, Menzel A, Pereira M. 2013. A plant's perspective of extremes: terrestrial plant responses to changing climatic variability. Global Change Biology 19: 75‐89. DOI: 10.1111/Gcb.12023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robson TM, Lavorel S, Clement JC, Le Roux X. 2007. Neglect of mowing and manuring leads to slower nitrogen cycling in subalpine grasslands. Soil Biology and Biochemistry 39: 930‐941. DOI: 10.1016/j.soilbio.2006.11.004. [Google Scholar]
- Serrano‐Muela MP, Nadal‐Romero E, Lana‐Renault N, González‐Hidalgo JC, López‐Moreno JI, Beguería S, Sanjuan Y, García‐Ruiz JM. 2013. An exceptional rainfall event in the central western Pyrenees: spatial patterns in discharge and impact. Land Degradation and Development. DOI: 10.1002/ldr.2221. [Google Scholar]
- Schaap BF, Reidsma P, Verhagen J, Wolf J, van Ittersum MK. 2013. Participatory design of farm level adaptation to climate risks in an arable region in The Netherlands. European Journal of Agronomy 48, 30–42. DOI: 10.1016/j.eja.2013.02.004. [Google Scholar]
- Schirpke U, Leitinger G, Tasser E, Schermer M, Steinbacher M, Tappeiner U. 2013a. Multiple ecosystem services of a changing Alpine landscape: past, present and future. International Journal of Biodiversity Science, Ecosystem Services & Management 9: 123–135. DOI: 10.1080/21513732.2012.751936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schirpke U, Tasser E, Tappeiner U. 2013b. Predicting scenic beauty of mountain regions. Landscape Urban Plan 111: 1‐12. DOI: 10.1016/j.landurbplan.2012.11.010. [Google Scholar]
- Schmitt M, Bahn M, Wohlfahrt G, Tappeiner U, Cernusca A. 2010. Land use affects the net ecosystem CO2 exchange and its components in mountain grasslands. Biogeosciences 7: 2297‐2309. DOI: 10.5194/bg-7-2297-2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seneviratne SI, Nicholls N, Easterling D, Goodess CM, Kanae S, Kossin J, Luo Y, Marengo J, McInnes K, Rahimi M, Reichstein M, Sorteberg A, Vera C, Zhang X, Rusticucci M, Semenov V, Alexander LV, Allen S, Benito G, Cavazos T, Clague J, Conway D, Della‐Marta PM, Gerber M, Gong S, Goswami BN, Hemer M, Huggel C, van den Hurk B, Kharin VV, Kitoh A, Tank AMGK, Li G, Mason S, McGuire W, van Oldenborgh GJ, Orlowsky B, Smith S, Thiaw W, Velegrakis A, Yiou P, Zhang T, Zhou T, Zwiers FW, Field CB, Barros V, Stocker TF, Dahe Q. 2012. Changes in Climate Extremes and their Impacts on the Natural Physical Environment In: Field CB, Barros V, Stocker TF, Dahe Q, Dokken DJ, Ebi KL, Mastrandrea MD, Mach KJ, Plattner G‐K, Allen SK, Melinda T, Midgley PM. (eds). Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation. Cambridge University Press, Cambridge, UK, and New York, USA; pp 109–230. [Google Scholar]
- Seppelt R, Dormann CF, Eppink FV, Lautenbach S, Schmidt S. 2011. A quantitative review of ecosystem service studies: approaches, shortcomings and the road ahead. Journal of Applied Ecology 48: 630‐636. DOI: 10.1111/j.1365-2664.2010.01952.x. [Google Scholar]
- Soussana JF, Luscher A. 2007. Temperate grasslands and global atmospheric change: a review. Grass and Forage Science 62: 127–134. DOI: 10.1111/j.1365-2494.2007.00577.x. [Google Scholar]
- Strauss F, Formayer H, Schmid E. 2013. High resolution climate data for Austria in the period 2008‐2040 from a statistical climate change model. International Journal of Climatology 33: 430‐443. DOI: 10.1002/Joc.3434. [Google Scholar]
- Taguas EV, Carpintero E, Ayuso JL. 2013. Assessing land degradation risk through the long‐term analysis of erosivity: a case study in southern Spain. Land Degradation and Development 24: 179–187. DOI: 10.1002/Ldr.1119. [Google Scholar]
- Tappeiner U, Borsdorf A, Bahn M. 2013. Long‐term socio‐ecological research in mountain regions: perspectives from the Tyrolean Alps In: Singh SJ, Haberl H, Chertow M, Mirtl M, Schmid M. (eds). Long Term Socio‐Ecological Research. Springer; Netherlands; pp 505–525. [Google Scholar]
- Tasser E, Mader M, Tappeiner U. 2003. Effects of land use in alpine grasslands on the probability of landslides. Basic and Applied Ecology 4: 271‐280. DOI: 10.1078/1439-1791-00153. [Google Scholar]
- Tasser E, Tappeiner U. 2005. New model to predict rooting in diverse plant community compositions. Ecological Modelling 185, 195–211. DOI: 10.1016/j.ecolmodel.2004.11.024. [Google Scholar]
- Varadan RJ, Kumar P. 2014. Indigenous knowledge about climate change: validating the perceptions of dryland farmers in Tamil Nadu. Indian Journal of Traditional Knowledge 13: 390–397. [Google Scholar]
- Vetter S, Bond WJ. 2012. Changing predictors of spatial and temporal variability in stocking rates in a severely degraded communal rangeland. Land Degradation and Development 23: 190‐199. DOI: 10.1002/Ldr.1076. [Google Scholar]
- Viviroli D, Weingartner R, Messerli B. 2003. Assessing the hydrological significance of the world's mountains. Mountain Research and Development 23: 32‐40. DOI: 10.1659/0276-4741(2003)023[0032:Athsot]2.0.Co;2. [Google Scholar]
- Walter J, Grant K, Beierkuhnlein C, Kreyling J, Weber M, Jentsch A. 2012. Increased rainfall variability reduces biomass and forage quality of temperate grassland largely independent of mowing frequency. Agriculture, Ecosystems & Environment 148: 1‐10. DOI: 10.1016/j.agee.2011.11.015. [Google Scholar]
- Wang N, Jiao JY, Lei D, Chen Y, Wang DL. 2014. Effect of rainfall erosion: seedling damage and establishment problems. Land Degradation and Development 25: 565–572. DOI: 10.1002/ldr.2183. [Google Scholar]
- Wieser G, Hammerle A, Wohlfahrt G. 2008. The water balance of grassland ecosystems in the Austrian Alps. Arctic Antarctic and Alpine Research 40: 439‐445. DOI: 10.1657/1523-0430(07-039)[Wieser]2.0.Co;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wigmosta MS, Vail LW, Lettenmaier DP. 1994. A distributed hydrology‐vegetation model for complex terrain. Water Resources Research 30: 1665‐1679. DOI: 10.1029/94wr00436. [Google Scholar]
- ZAMG . 2014. Current Climate Monitoring. Central Institute for Meteorology and Geogynamics, Vienna, Austria. http://www.zamg.ac.at/cms/en/climate/climate‐overview/climate_monitor/jahr/wien_hohe_warte/?jahr=2013 (09.06.2014).
- Zwicke M, Alessio GA, Thiery L, Falcimagne R, Baumont R, Rossignol N, Soussana JF, Picon‐Cochard C. 2013. Lasting effects of climate disturbance on perennial grassland above‐ground biomass production under two cutting frequencies. Global Change Biology 19: 3435–3448. DOI: 10.1111/Gcb.12317. [DOI] [PubMed] [Google Scholar]