

Abstract
The roundworm C. elegans has been successfully used for more than 50 y as a genetically tractable invertebrate model in diverse biological fields such as neurobiology, development and interactions. C. elegans feeds on bacteria and can be naturally infected by a wide range of microorganisms, including viruses, bacteria and fungi. Most of these pathogens infect C. elegans through its gut, but some have developed ways to infect the epidermis. In this review, we will mainly focus on epidermal innate immunity, in particular the signaling pathways and effectors activated upon wounding and fungal infection that serve to protect the host. We will discuss the parallels that exist between epidermal innate immune responses in nematodes and mammals.
Keywords: collagen, cuticle, damage, Epidermal barrier, epidermis, GPCR, hemidesmosome, pathogen, Pathogen-barrier interactions, signaling, wounding
Introduction
A large body of knowledge has been accumulated for the tiny worm Caenorhabditis elegans since it was chosen as a model organism by Brenner 50 y ago.1 C. elegans, first described by Maupas at the start of the last century,2 is a free-living nematode, 1 mm long, found in rotting fruit. In the lab it can easily be grown on agar petri dishes or in liquid culture. Being a self-fertilizing hermaphrodite, it can be expanded as a clonal population, but males can also be maintained, allowing genetic approaches to be undertaken. With a short life cycle of 3 days, and 300 progeny per adult, large homogenous populations can be generated easily. Its transparency facilitates live imaging, very useful for cell biological studies. C. elegans has a simple anatomy with less than 1000 cells organized in a small number of tissues and internal organs.3
In C. elegans, as in mammals, the epidermis acts as a barrier to protect the organism from environmental damage and pathogens, prevents leakage of internal molecules and blocks the entrance of foreign compounds. Unlike the multilayered stratified epithelium in mammals, the adult C. elegans epidermis (also termed “hypodermis” for historical reasons) is mainly composed of one cell layer, including a large syncytial cell, called hypodermal cell 7 (hyp7), which surrounds the worm and covers most of the body length, and a line of specialized lateral cells on each side of the worm, called the seam cells (Fig. 1).4
Figure 1.
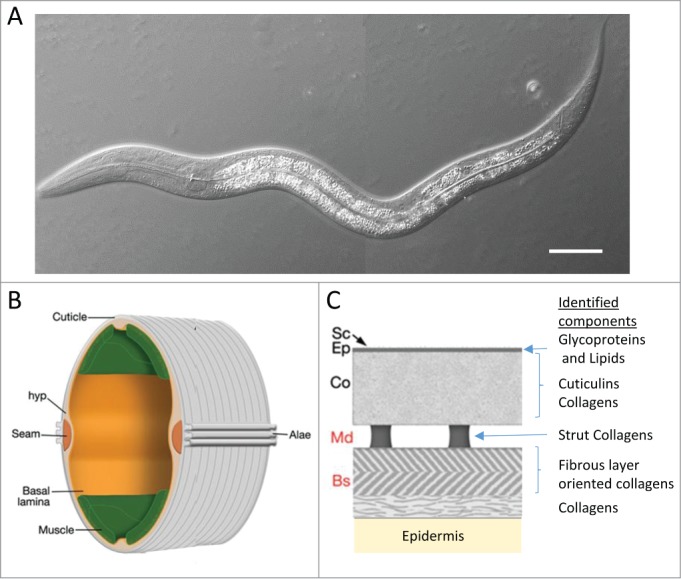
C. elegans anatomy. (A) Image of an early larval stage in DIC, scale bar is 20 μm. Schematic of an adult C. elegans body (B) and cuticle (C). The adult cuticle is approximately 0.5 μm in thickness and subdivided in (Bs) basal zone, (Md) medial zone, (Co) cortical zone, (Ep) epicuticle and (Sc) Surface coat “should be” subdivided in basal zone (Bs), medial zone (Md), cortical zone (Co), epicuticle (Ep) and surface coat (Sc). Collagen is present in all the major layers, except for Ep and Sc which are rich in lipids and glycoproteins, respectively. The cuticle is synthesized by the epidermis, formed by the hypodermis (hyp), a simple epidermal syncitium and the lateral seam cells (adapted from Wormatlas, http://www.wormatlas.org/hermaphrodite/cuticle/mainframe.htm).
As in most invertebrates, C. elegans possesses an exoskeleton. In nematodes, this is a tough, but flexible external cuticle secreted by the epidermis. This collagenous extracellular matrix (ECM) maintains the integrity of the worm, defines the body shape of the animal and is required for locomotion, through its attachment to the underlying muscles. The cuticle is subdivided into basal, medial and cortical zones, overlaid by the epicuticle and the most external layer, the surface coat. While the cuticle is mainly composed of collagen and insoluble proteins, called cuticlins, the epicuticle and the surface coat are rich in lipids and structural glycoproteins such as mucins, respectively. Secreted non-structural proteins also likely make up an important part of the cuticle, such as enzymes involved in post-secretion modification and cross-linking of matrix proteins or structural proteins associated with the surface coat.5,6 In mammals, the most external layer of the skin, the stratum corneum (SC) is the final product of keratinocyte differentiation, resulting from denucleation and crosslinking of intracellular proteins. It is mainly composed of keratin, cholesterol, free fatty acids and ceramides.7 Although differing in composition, the C. elegans cuticle can be considered analogous to the SC as they both function as a permeability barrier.8
Cuticle Collagen: A Key Component in Barrier Integrity
Collagen is the main structural protein of the extracellular matrix in animals, and the most abundant protein in mammals. It is an essential component of the skin and plays a key role in organogenesis and body morphology. In C. elegans, 2 major collagen families are present, the cuticle and the basement membrane collagens. While only 3 genes encodes type IV and type XVIII basement membrane collagens, the cuticle collagens are encoded by more than 170 genes. They are most similar to the fibril-associated collagens with interrupted triple helices (FACIT) found in vertebrates.5 They are regulated differentially, most of them following the 4 molting cycles during larval development and are involved in the formation of distinct sub-cuticle structures (e.g. annular furrows and alae) and layers.9 Collagen biogenesis involves a complex maturation process, partially shared between nematodes and vertebrates, during which pro-collagen undergoes several co-translational and post-translational modifications. Collagen maturation ends with the secretion of collagen triple helices that are covalently cross-linked via tyrosine residues. This determines the characteristic stiffness and integrity of the worm cuticle.5 During this step, a peroxidase (MLT-7) cooperates with an NADPH dual oxidase (BLI-3), to promote reactive oxygen species (ROS)-mediated catalysis of di- and tri- tyrosine covalent bonds.10,11 Recently, a tetraspanin protein (TSP-15) was found to form a complex with BLI-3/DUOX and MLT-7/peroxidase and to be required for ROS generation.12 In the absence of any one of these 3 proteins, no di- and tri- tyrosine bonds are made and the cuticle is fragile and blistered. In vertebrates, the formation of covalent cross-links of proteins such as involucrin and loricrin in the SC is also essential for the function of mammalian skin as an external barrier. It occurs via the action of transglutaminases that leads to the oxidation of amino group of lysine or glutamate residues into aldehyde derivatives. Collagen helices are cross-linked by lysyl oxidase in vertebrates between lysine and hydroxyl lysine residues. Lysyl oxidase-mediated cross-linkages are not found in the C. elegans cuticle, they only contribute to cross-linking of type IV collagen in the basement membrane.13,14
In human skin, collagens are mainly found in the dermis. Many mutations in different collagen genes have been associated with various diseases.15 For example, patients with epidermolysis bullosa, carry mutations in COL7A1 or COL17A1 that lead to blisters in the skin and mucosal membranes, and mutations in COL3A1 or COL5A1 have been found in patients with Ehlers–Danlos syndrome, with various symptoms including a fragile skin.16,17 Similarly, in nematodes, mutations in genes encoding collagens, like DPY-9, ROL-6, BLI-1, or processing enzymes, including the prolyl 4-hydroxylase DPY-18 or the disulfide isomerase PDI-2, result in diverse body morphology defects described as ROLler (Rol: helical twisting of the animal's body), DumPY (Dpy: shortening in the body length), or BLIster (Bli: blistering of cuticle material away from the surface of the animal).5
The nematode cuticle is made of successive layers of different collagens. Interestingly, some mutants in specific collagen genes, such as dpy-2, dpy-7 or dpy-10, which are presumed to be the most external collagens of the cuticle, show high osmotic resistance, due to a higher internal level of glycerol. These same collagens are also required for proper circumferential furrow formation.18 Wheeler and Thomas proposed that these collagens on the furrows could act as a stretch sensor, monitoring the turgor pressure of the cuticle. OSM-7 and OSM-11, which are nematode-specific proteins structurally related to the Notch ligand Delta, possibly associated with the cuticle, could transduce the signal, leading to the increase in glycerol.18 The same mutants also show a constitutive activation of innate immunity (see below).19,20 As previous studies found these mutants to be involved in the suppression of diverse hypomorphic mutations, probably through the induction of chaperone function,21-24 these results suggest that some collagens are part of a general stress sensing mechanism.
Epidermal Wounding: Cell Autonomous Innate and Healing Response
Just as mutation of genes required for the proper development of the worm's cuticle can trigger an innate immune response, so too does directly wounding the worm. This can be done either by hand, with a fine glass needle, when the worm is pricked at the tail, or in a more controlled manner with a laser that enables precise wounding of the epidermis. In both cases, at the wound site there is an immediate deposition of as-yet uncharacterized autofluorescent material that perdures for many hours. Later, the secretion of basal cuticle components leads to the repair of the damaged cuticle, leaving a scar at the wound site. Moreover, injury provokes a rapid induction of antimicrobial peptide (AMP) gene expression in the epidermis. For example a rise in the expression of nlp-29 is detectable within one hour after injury and is sustained for several hours (Fig. 2A).20 This induction of innate effectors upon wounding is not due to microbe invasion but is a preventive response of the tissues, as previously shown in other species.25 Indeed, mutants that are defective for the induction of defense gene expression upon wounding succumb to opportunistic infections.26
Figure 2.
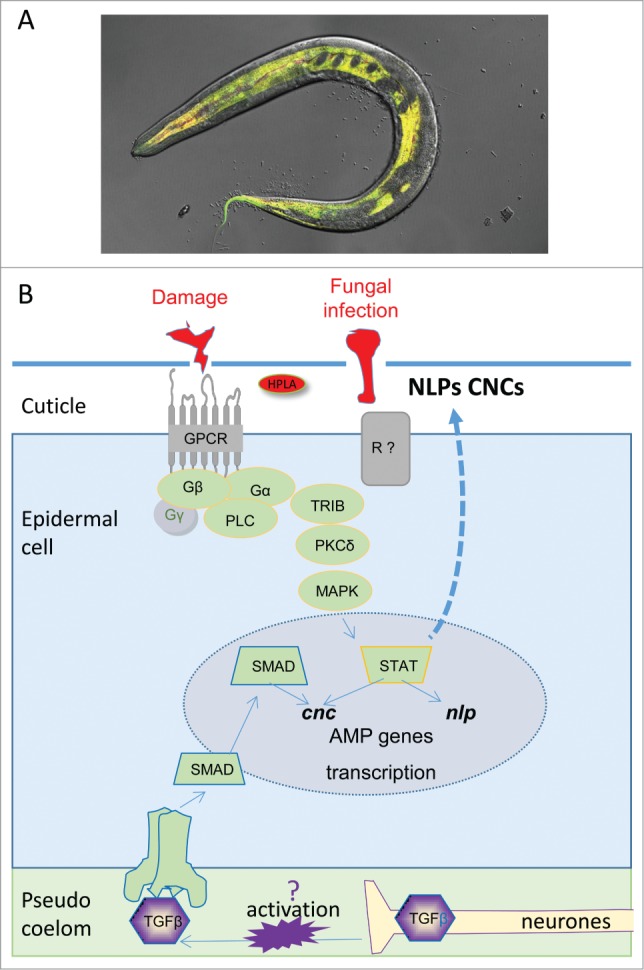
(A) Images of an adult worm infected with D. coniospora inducing an AMP reporter gene linked to GFP in his epidermis. (B) Schematic representation of the innate immunity pathways activated in C. elegans epidermis upon D. coniospora infection and wounding. AMP, antimicrobial peptide.
In order to identify the first event which initiates the epidermal response to damage, Xu and Chisholm focused their attention on calcium signaling, as it was already known to be involved in embryonic wound healing and the response to membrane injury in single cells.27,28 Upon injury to the epidermal syncytium, an immediate Ca2+ wave, released from internal stores, can be detected at the wound site and provokes local formation of a dense actin ring which, in turn, will trigger complete wound closure in 24 hrs. This epidermal Ca2+ response involves the epidermal transient receptor potential channel, metastatin family (TRPM) GTL-2, the Gαq EGL-30 and its effector PLCβ EGL-8.29 Subsequently it was shown that specific Ca2+ uptake into the mitochondria close to the wound site leads to local mitochondrial ROS (mtROS) production and that mtROS release inhibits the Rho GTPase RHO-1, promoting actin polymerization and filopodial-like protrusion in a Cdc-42 and Arp2/3 dependent manner.29,30 This wound closure process is different from the most common Rho GTPase-dependent purse-string mechanism that occurs, for instance, after wounding in the Drosophila embryo.31
While the pathways leading either to wound healing or AMP production (see below) are genetically and mechanistically distinct,29 the worm homolog of the tumor suppressor death-associated protein kinase, DAPK-1, functions as a negative regulator of both barrier repair and AMP expression. Loss of function of dapk-1 leads to spontaneous cuticle hypertrophy with associated appearance of autofluorescent aggregates and to the constitutive upregulation of epidermal AMPs.26 Thus as for other immune responses, the epidermal response to damage has to be tightly regulated to prevent the deleterious consequences of chronic activation.
Pathogen Infection
While some pathogens will only passively enter the host through a wound, others have developed active ways to infect the worm through the cuticle. The nematophagous fungus Lecanicillium psalliotae is able to degrade the worm cuticle by secreting a serine protease, thus facilitating mycelia penetration and complete nematode degradation.32 Other fungi, such as Arthrobotrys oligospora, Drechslerella stenobrocha and Dactylellina ellipsospora, are able to form traps to capture, kill and digest worms as a food source.33 One of the most studied pathogens able to infect the worms via its cuticle is Drechmeria coniospora. This endoparasitic fungi is frequently found associated with nematodes in nature.34,35 Its conidia (non-motile spores) can bind to the worm's cuticle through specialized adhesive knobs and upon germination spread hyphae throughout the worm's body, causing death after as little as 48 hours.36 Some bacteria also adhere to the cuticle, like Leucobacter spp Verde37 or Microbacterium nematophilum that colonizes the rectal cuticle and provokes swelling in the adjacent epithelia.38,39
Fungi and bacteria take advantage of the carbohydrate-rich surface coat and are believed to use lectins to adhere to the nematode cuticle. Recent studies in C. elegans have identified mutants with defects in their surface coat carbohydrates and an increased adhesion of D. coniospora. For instance, the bus-2 mutant carries a mutation in the enzyme galactosyltransferase and is enriched in core-2 fucosyl O-glycans at the expense of core-1 glycans, suggesting that there is an increase in fucose exposure in this mutant.40,41 Moreover a fucose-specific lectin binds the vulva of wild-type worms, a preferential site for D. coniospora adhesion. Taken together, these results suggest that fucosyl glycans are a target for the binding of D. coniospora spores.42
It is interesting to note that if mutants like bus-2 have an increased adhesion to D. coniospora, they conversely have a reduced adhesion to the bacterial pathogen M. nematophilum, hence their name Bus for bacterial unswollen.43 The same opposite effect has been revealed with 2 different strains of Leucobacter spp Verde. Thus, resistance to some natural pathogens can be associated with increased susceptibility to others, revealing the opposing selective forces that must shape the evolution of C. elegans.37
The surface coat composition changes during development through the different larval stages and it was also shown to change upon environmental signals,44,45 which could be presages, for instance, of the presence of pathogens. Thus, it is reasonable to think that this switch could enable evasion of pathogens or, in case of the parasitic nematodes, escape of the host immune response.6 This would complement the purely mechanical shedding of pathogens that can accompany molting.46
AMP Induction in C. elegans Epidermis
If pathogens have developed ways to attach to the cuticle, it is now clear that the worm is able to mount an innate immune response after infection. Using cDNA and genomic DNA microarrays or more recently RNAseq, transcriptomic profiling has identified many genes induced upon infection with Monacrosporium haptotylum47 or/and D. coniospora.19,48-50 Among this latter class, some were predicted to encode AMPs, including 2 related families: a subset of neuropeptide-like proteins (NLPs) and the caenacins (CNCs).48,51 Members of these 2 families are present in 2 distinct genomic clusters: the nlp-29 (including nlp-27, nlp-28, nlp-29, nlp-30, nlp-31 and nlp-34) and the cnc-2 (including cnc-1, cnc-2, cnc-4, cnc-5, and cnc-11) clusters, which contribute to resistance to D. coniospora infection.19,52,53 While these 2 classes of genes are upregulated by infection and act in the epidermis, they are regulated by different signaling pathways (see below, Fig. 2B). The main immune players were mostly identified through genetic screens, through the selection of mutants, termed Nipi, which fail to express antimicrobial peptide genes after D. coniospora infection.20
A Cell Autonomous Response via the p38 MAPK Pathway
After D. coniospora infection or wounding, the expression of nlp genes is induced by the activation of a G-protein-coupled-receptor (GPCR) DCAR-1 in an epidermis-specific and cell-autonomous way (Fig. 2B). DCAR-1 is expressed in the apical part of the epidermal syncytium (hyp7) and is activated by HPLA (4-hydroxyphenyllactic acid). The level of this ligand is increased after infection, and also in mutants with defects in the cuticle like dpy-10. Thus, it seems that HPLA acts as a DAMP (damage-associated molecular pattern). Even though dcar-1 orthologs are only found in nematodes, these findings point to an ancient role for GPCR in innate immunity.54 Interestingly HPLA, a common tyrosine metabolite, is detected across all species and its elevated concentration was detected in patients with phenylketonuria (PKU), tyrosinemia or with Zellweger's syndrome.55-57
DCAR-1 acts upstream of the Gα protein GPA-12 and of the Gβ protein RACK-1 (orthologous to the mammalian Gα G12 and similar to the human Gβ-2-like 1, respectively), which in turn, act genetically upstream of the serine, threonine Protein Kinase C (PKC) TPA-1, whose activation is diacylglycerol (DAG) dependent. DAG is a second messenger produced from phosphatidylinositol biphosphate (PIP2) by the catalytic action of the phospholipase C (PLC) EGL-8 and PLC-3 (a PLCγ homolog).54,58
In mammals and insect, a family of adaptor proteins containing a domain named TIR, for Toll Interleukin Receptor, appears to be crucial for Toll Like receptor (TLR) signaling.59 C. elegans has only 2 genes encoding proteins with a TIR domain, the TLR tol-1 and the SARM adaptor tir-1. Upon fungal infection, AMP induction is tol-1 independent but tir-1 dependent.48 tir-1 acts upstream of the nsy-1/sek-1/pmk-1 p38-MAPK pathway to induce epidermal AMP expression20 and it is required for full resistance to infection.48 Both the STAT family protein STA-2 and the GATA transcription factor ELT-3 are required for the transcription of nlps19,60 (Fig. 2B).
Parallel to this pathway, a conserved Tribbles-like kinase, NIPI-3, has been found to act upstream of TPA-1 and the p38/MAPK cassette specifically upon D. coniospora infection.20,58 The tribbles family of pseudokinases are key controllers of signal transduction via their interactions with diverse kinases, ubiquitin ligases and transcription factors. Early studies in the fly also associated Tribbles-like kinase with the control of cell cycle during development.61 It will be interesting to test if NIPI-3/TriB has also a general function in cell cycle regulation in C. elegans.
A Paracrine role of Non-Canonical DBL-1/TGFβ Pathway
cnc gene expression is induced both after D. coniospora infection and injury, but is controlled in a complex way depending on the stimulus (Fig. 2B). It was shown that the induction of the cnc-2 gene cluster is p38-MAPK dependent after wounding, while it is p38-MAPK independent upon infection. After infection, cnc genes are upregulated by a non-canonical DBL-1/TGFβ pathway.53
The canonical DBL-1/TGFβ pathway was previously shown to be involved in regulation of body size and male tail development in C. elegans. DBL-1 binding to the heterodimeric DAF-4/SMA-6 receptor activates the intracellular Smad signal-transducer homolog SMA-2/SMA-3/SMA-4 complex which, in turn, will control gene expression through recruitment of the zinc-finger transcription factor, SMA-9, in the nucleus.62 If the entire SMA-2/SMA-3/SMA-4 complex is needed also for a response to P. aeruginosa,63 instead, only SMA-3 is required for a response to D. coniospora. The identity of the transcription factor involved in this process is still unclear (Fig. 2B).53 Strikingly, the DBL-1 ligand is mainly expressed in the nervous system, so it might act in a paracrine way to induce cnc gene expression in the epidermis,53 in addition to controlling defense gene expression in the intestine.64 It's still unknown, however, how the signal is transmitted from the neurons to the epidermis. It's possible that DBL-1-secreting neurons are directly in contact with the pseudocoelom, suggesting that DAF-4/SMA-6 receptor could be activated from the basal side of the epidermis.46
Role of Solute Carrier Family (SLC) and Pseudokinases in Immunity
C. elegans genetic screens highlighted also the presence of unexpected players in these innate immune pathways, such as the sodium-neurotransmitter symporter SNF-12, a member of the solute carrier family (SLC6). Interestingly, SNF-12 controls the constitutive expression of cnc genes and nlp gene induction after infection and wounding. SNF-12 physically interacts with the transcription factor STA-2 and they have been found to colocalise in dyn-1-dependent-endocytic vesicles through which they regulate part of the innate immune response (Fig. 2B).60 SNF-12/SLC6 is predicted to be a transporter of a bioactive amine, but its endogenous substrate has yet to be identified.
Studies in vertebrates revealed that another class of SLC protein, transporting oligopeptides, might have a crucial function for the antibacterial or antiviral activity of macrophages, mature dendritic cells (mDCs)65 and plasmacytoid dendritic cells (pDCs).66 In mouse mDCs and macrophages, Slc15a mediates the internalization in late endosomes/lysosome of fragments of peptidoglycan coming from bacteria cell wall, and stimulates NF-kB and other immune effector pathways.65 Moreover, it was shown that the peptide/histidine transporter Slc15a4 is present both in mouse pDCs66 and B cells lysosomes where, by maintaining the appropriate pH, it can promote optimal TLR7 and TLR9 activation.67 Interestingly, Slc15a4 is associated with systemic lupus erythematosus (SLE).68
Another player genetically interacting with STA-2 and SNF-12 in C. elegans is the pseudokinase NIPI-4 (Fig. 2B).69 Interestingly, even though pseudokinases lack the ability to phosphorylate substrates, they are still able to regulate cellular processes, forming heterodimers with other protein kinases and controlling their activity.70 For instance, in humans, the pseudokinase STe20 Related ADaptor (STRAD)α, with its closed conformation typical of active protein kinases, can bind the tumor suppressor protein kinase LKB-1, induce a conformational change in LKB-1, which, in turn, enables cell proliferation, and regulates cell polarity as well as energy levels.70,71 NIPI-4 is expected to have a similar regulatory role. Although the identity of its target kinase is unknown, the p38 MAPK PMK-1 is a prime candidate.
Is Mechanosensation Involved in Innate Immunity Induction?
It is still not clear how p38 MAPK-dependent STA-2 activation and trafficking are regulated in C. elegans epidermis. A very recent study suggests that in the case of severe epidermal injury, upregulation of nlp-29 and cnc-2 could occur in a p38 independent, but STA-2 dependent way.72 STA-2 was found in the nucleus but also associated with hemidesmosomes (CeHDs),60,72 structures responsible for the attachment of the epidermal cells to the cuticle.73,74 Zhang et al. showed that STA-2 physically interacts with one apical component of CeHDs, MUP-4 a transmembrane multidomain protein. They hypothesized that the interaction of STA-2 with CeHDs inhibits its activity and that severe damage or CeHDs disruption would free STA-2 to induce AMP production.72 They further propose that this mechanism is conserved in humans where β-defensin transcription correlates with disruption of hemidesmosomes protein complexes (HPC) and inactivation of STA-3 or STA-5B attenuates AMPs upregulation after HPC disassembly in human epidermal keratinocyte (HEKa) cells.72 The suggestion that hemidesmosomes will act as a damage-sensor raises very interesting questions, like how do the HDs perceive the damage? And what are the signals that will activate HDs? Some hints have come from studies of epidermal development.
In C. elegans, it was known that muscle contraction is required in the embryo during morphogenesis for epidermal elongation. From a recent study, it was shown that upon muscle contraction p21-activated kinase (PAK-1), a component of CeHDs, is activated and together with the adaptor GIT-1 (G-protein-coupled receptor kinase interactor) and its partner PIX-1 (PAK-interacting exchange factor) can phosphorylate epidermal intermediate filaments and promote CeHD biogenesis. In mutants defective for muscle contraction, GIT-1 progressively leaves HDs. Zhang et al. could show that applying external mechanical pressure on these mutants considerably retarded the diffusion of GIT-1. Thus, they demonstrated that CeHDs are mechanosensitive and direct targets of physical forces.75 In the future it would be of interest to address whether injury or spore adhesion could induce a mechanical signal that could be sensed by apical CeHDs.
Interestingly, there is evidence suggesting a role for collagens in HDs integrity. In humans, mature type-1 HDs contain the bullous pemphigoid (BP) antigens BP180/collagen XVII as well as integrin α6β4 and plectin. Mutations in the corresponding genes in patients with epidermolysis bullosa alter hemidesmosomal structure.76 Also during carcinogenesis, extracellular cleavage of collagen XVII by matrix-metalloprotease MMP-9 disrupts HDs.77 In C. elegans, worms with mutations in collagen genes, such as dpy-9 and rol-6, present constitutive upregulation of AMPs.19 As the extracellular portion of MUP-4, the apical CeHD component, contains a von Willebrand factor A domain for collagen binding,72 it is possible that MUP-4 acts as a regulator of the epidermal immune response conceivably sensing, after injury or spore adhesion, collagen disruption.
Conclusions
C. elegans lacks both an adaptive immune system and specialized immune cells. This obviously limits its relevance for the understanding of human immunity. On the other hand, its relative simplicity and the extensive range of available experimental methods facilitate its use in biology. The major molecular mechanisms involved in the epidermal innate immune response in C. elegans have been identified. As in humans, they involve the upregulation of AMP expression and wound healing, governed by a combination of highly conserved and nematode-specific pathways. A common theme that has emerged is the importance of damage recognition as a trigger for the induction of defense genes. Future studies should reveal the mechanisms leading to the production and/or release of DAMPs and the identity of the elusive “fungus-receptor” putatively responsible for triggering pathogen-specific innate immune defenses. Together with a better understanding of how cross-tissue communication contributes to the regulation of host immunity, this will lead to a more comprehensive insight into how an organism can re-establish homeostasis after the integrity of epithelial barriers has been compromised by injury or infection.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank Jonathan Ewbank for critical reading.
Funding
Work in our lab is supported by institutional funding from INSERM, CNRS and AMU, and program grants from the Research National Agency ANR-12-BSV3–0001–01(FUNEL) and ANR Investissements d'Avenir ANR-11-LABX-0054 (Labex INFORM) ANR-10-INSB-04–01 (France-BioImaging) and ANR-11-IDEX-0001–02 (A*MIDEX)).
References
- 1.Brenner S. The genetics of Caenorhabditis elegans. Genetics 1974; 77:71-94; PMID:4366476 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Maupas E. Modes et formes de reproduction des nematodes. Archives de Zoologie Experimentale et Generale 1900(8):463-624 [Google Scholar]
- 3.Corsi AK, Wightman B, Chalfie M. A Transparent window into biology: A primer on Caenorhabditis elegans. WormBook 2015:1-31; PMID:26087236; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Altun ZF, Hall DH. Epithelial system, hypodermis. WormAtlas 2009 [Google Scholar]
- 5.Page AP, Johnstone IL. The cuticle. WormBook: The C. elegans Research Community ed, 2007:1-15; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Davies KG, Curtis RH. Cuticle surface coat of plant-parasitic nematodes. Annu Rev Phytopathol 2011; 49:135-56; PMID:21568702 [DOI] [PubMed] [Google Scholar]
- 7.Madison KC. Barrier function of the skin: “la raison d'etre” of the epidermis. J Invest Dermatol 2003; 121:231-41; PMID:12880413 [DOI] [PubMed] [Google Scholar]
- 8.Xu S, Hsiao TI, Chisholm AD. The wounded worm: Using C. elegans to understand the molecular basis of skin wound healing. Worm 2012; 1:134-8; PMID:24058838 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Johnstone IL. Cuticle collagen genes. Expression in Caenorhabditis elegans. Trends Genet 2000; 16:21-7; PMID:10637627 [DOI] [PubMed] [Google Scholar]
- 10.Thein MC, Winter AD, Stepek G, McCormack G, Stapleton G, Johnstone IL, Page AP. Combined extracellular matrix cross-linking activity of the peroxidase MLT-7 and the dual oxidase BLI-3 is critical for post-embryonic viability in Caenorhabditis elegans. J Biol Chem 2009; 284:17549-63; PMID:19406744 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Edens WA, Sharling L, Cheng G, Shapira R, Kinkade JM, Lee T, Edens HA, Tang X, Sullards C, Flaherty DB, et al.. Tyrosine cross-linking of extracellular matrix is catalyzed by Duox, a multidomain oxidase/peroxidase with homology to the phagocyte oxidase subunit gp91phox. J Cell Biol 2001; 154:879-91; PMID:11514595 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Moribe H, Konakawa R, Koga D, Ushiki T, Nakamura K, Mekada E. Tetraspanin is required for generation of reactive oxygen species by the dual oxidase system in Caenorhabditis elegans. PLoS Genet 2012; 8:e1002957; PMID:23028364 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Norman KR, Moerman DG. The let-268 locus of Caenorhabditis elegans encodes a procollagen lysyl hydroxylase that is essential for type IV collagen secretion. Dev Biol 2000; 227:690-705; PMID:11071784 [DOI] [PubMed] [Google Scholar]
- 14.Cox GN, Staprans S, Edgar RS. The cuticle of Caenorhabditis elegans. II. Stage-specific changes in ultrastructure and protein composition during postembryonic development. Dev Biol 1981; 86:456-70; PMID:6269931 [DOI] [PubMed] [Google Scholar]
- 15.Myllyharju J, Kivirikko KI. Collagens, modifying enzymes and their mutations in humans, flies and worms. Trends Genet 2004; 20:33-43; PMID:14698617 [DOI] [PubMed] [Google Scholar]
- 16.Has C, Bruckner-Tuderman L. The Genetics of Skin Fragility. Annu Rev Genom Hum G 2014; 15:245-68; http://dx.doi.org/ 10.1146/annurev-genom-090413-025540 [DOI] [PubMed] [Google Scholar]
- 17.Frank M, Albuisson J, Ranque B, Golmard L, Mazzella JM, Bal-Theoleyre L, Fauret AL, Mirault T, Denarie N, Mousseaux E, et al.. The type of variants at the COL3A1 gene associates with the phenotype and severity of vascular Ehlers-Danlos syndrome. Eur J Hum Genet 2015; PMID:25758994 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Wheeler JM, Thomas JH. Identification of a novel gene family involved in osmotic stress response in Caenorhabditis elegans. Genetics 2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Pujol N, Zugasti O, Wong D, Couillault C, Kurz CL, Schulenburg H, Ewbank JJ. Anti-fungal innate immunity in C. elegans is enhanced by evolutionary diversification of antimicrobial peptides. PLoS Pathog 2008; 4:e1000105; PMID:18636113; http://dx.doi.org/ 10.1371/journal.ppat.1000105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Pujol N, Cypowyj S, Ziegler K, Millet A, Astrain A, Goncharov A, Jin Y, Chisholm AD, Ewbank JJ. Distinct innate immune responses to infection and wounding in the C. elegans epidermis. Curr Biol 2008; 18:481-9; PMID:18394898; http://dx.doi.org/ 10.1016/j.cub.2008.02.079 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Goh PY, Bogaert T. Positioning and maintenance of embryonic body wall muscle attachments in C. elegans requires the mup-1 gene. Development 1991; 111:667-81; PMID:1879334 [DOI] [PubMed] [Google Scholar]
- 22.Kramer JM, Johnson JJ. Analysis of mutations in the sqt-1 and rol-6 collagen genes of Caenorhabditis elegans. Genetics 1993; 135:1035-45; PMID:8307321 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Levy AD, Yang J, Kramer JM. Molecular and genetic analyses of the Caenorhabditis elegans dpy-2 and dpy-10 collagen genes: a variety of molecular alterations affect organismal morphology. Mol Biol Cell 1993; 4:803-17; PMID:8241567; http://dx.doi.org/ 10.1091/mbc.4.8.803 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Nishiwaki K, Miwa J. Mutations in genes encoding extracellular matrix proteins suppress the emb-5 gastrulation defect in Caenorhabditis elegans. Mol Gen Genet 1998; 259:2-12; PMID:9738874; http://dx.doi.org/ 10.1007/s004380050782 [DOI] [PubMed] [Google Scholar]
- 25.Sorensen OE, Thapa DR, Roupe KM, Valore EV, Sjobring U, Roberts AA, Schmidtchen A, Ganz T. Injury-induced innate immune response in human skin mediated by transactivation of the epidermal growth factor receptor. J Clin Invest 2006; 116:1878-85; PMID:16778986; http://dx.doi.org/ 10.1172/JCI28422 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Tong A, Lynn G, Ngo V, Wong D, Moseley SL, Ewbank JJ, Goncharov A, Wu YC, Pujol N, Chisholm AD. Negative regulation of Caenorhabditis elegans epidermal damage responses by death-associated protein kinase. Proc Natl Acad Sci U S A 2009; 106:1457-61; PMID:19164535; http://dx.doi.org/ 10.1073/pnas.0809339106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.McNeil PL, Steinhardt RA. Plasma membrane disruption: repair, prevention, adaptation. Annu Rev Cell Dev Biol 2003; 19:697-731; PMID:14570587; http://dx.doi.org/ 10.1146/annurev.cellbio.19.111301.140101 [DOI] [PubMed] [Google Scholar]
- 28.Stanisstreet M. Calcium and wound healing in Xenopus early embryos. J Embryol Exp Morphol 1982; 67:195-205; PMID:6806425 [PubMed] [Google Scholar]
- 29.Xu S, Chisholm AD. A Galpha(q)-Ca(2+) signaling pathway promotes actin-mediated epidermal wound closure in C. elegans. Curr Biol 2011; 21:1960-7; PMID:22100061; http://dx.doi.org/ 10.1016/j.cub.2011.10.050 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Xu S, Chisholm AD. C. elegans epidermal wounding induces a mitochondrial ROS burst that promotes wound repair. Dev Cell 2014; 31:48-60; PMID:25313960; http://dx.doi.org/ 10.1016/j.devcel.2014.08.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wood W, Jacinto A, Grose R, Woolner S, Gale J, Wilson C, Martin P. Wound healing recapitulates morphogenesis in Drosophila embryos. Nat Cell Biol 2002; 4:907-12; PMID:12402048; http://dx.doi.org/ 10.1038/ncb875 [DOI] [PubMed] [Google Scholar]
- 32.Yang J, Huang X, Tian B, Wang M, Niu Q, Zhang K. Isolation and characterization of a serine protease from the nematophagous fungus, Lecanicillium psalliotae, displaying nematicidal activity. Biotechnol Lett 2005; 27:1123-8; PMID:16132863; http://dx.doi.org/ 10.1007/s10529-005-8461-0 [DOI] [PubMed] [Google Scholar]
- 33.Xie H, Aminuzzaman FM, Xu L, Lai Y, Li F, Liu X. Trap induction and trapping in eight nematode-trapping fungi (Orbiliaceae) as affected by juvenile stage of Caenorhabditis elegans. Mycopathologia 2010; 169:467-73; PMID:20146004; http://dx.doi.org/ 10.1007/s11046-010-9279-4 [DOI] [PubMed] [Google Scholar]
- 34.Felix MA, Duveau F. Population dynamics and habitat sharing of natural populations of Caenorhabditis elegans and C. briggsae. BMC Biol 2012; 10:59; PMID:22731941; http://dx.doi.org/ 10.1186/1741-7007-10-59 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Jansson HB. Adhesion of conidia of Drechmeria coniospora to Caenorhabditis elegans wild type and mutants. J Nematol 1994; 26:430-5; PMID:19279912 [PMC free article] [PubMed] [Google Scholar]
- 36.Jansson HB, von Hofsten A, von Mecklenburg C. Life cycle of the endoparasitic nematophagous fungus Meria coniospora: a light and electron microscopic study. Antonie Van Leeuwenhoek 1984; 50:321-7; PMID:6543109; http://dx.doi.org/ 10.1007/BF00394645 [DOI] [PubMed] [Google Scholar]
- 37.Hodgkin J, Felix MA, Clark LC, Stroud D, Gravato-Nobre MJ. Two Leucobacter strains exert complementary virulence on Caenorhabditis including death by worm-star formation. Curr Biol 2013; 23:2157-61; PMID:24206844; http://dx.doi.org/ 10.1016/j.cub.2013.08.060 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Nicholas HR, Hodgkin J. The ERK MAP kinase cascade mediates tail swelling and a protective response to rectal infection in C. elegans. Curr Biol 2004; 14:1256-61; PMID:15268855; http://dx.doi.org/ 10.1016/j.cub.2004.07.022 [DOI] [PubMed] [Google Scholar]
- 39.Hodgkin J, Kuwabara PE, Corneliussen B. A novel bacterial pathogen, Microbacterium nematophilum, induces morphological change in the nematode C. elegans. Curr Biol 2000; 10:1615-8; PMID:11137017; http://dx.doi.org/ 10.1016/S0960-9822(00)00867-8 [DOI] [PubMed] [Google Scholar]
- 40.Gravato-Nobre MJ, Stroud D, O'Rourke D, Darby C, Hodgkin J. Glycosylation genes expressed in seam cells determine complex surface properties and bacterial adhesion to the cuticle of Caenorhabditis elegans. Genetics 2011; 187:141-55; PMID:20980242; http://dx.doi.org/ 10.1534/genetics.110.122002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Palaima E, Leymarie N, Stroud D, Mizanur RM, Hodgkin J, Gravato-Nobre MJ, Costello CE, Cipollo JF. The Caenorhabditis elegans bus-2 mutant reveals a new class of O-glycans affecting bacterial resistance. J Biol Chem 2010; 285:17662-72; PMID:20385555; http://dx.doi.org/ 10.1074/jbc.M109.065433 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Rouger V, Bordet G, Couillault C, Monneret S, Mailfert S, Ewbank JJ, Pujol N, Marguet D. Independent Synchronized Control and Visualization of Interactions between Living Cells and Organisms. Biophys J 2014; 106:2096-104; PMID:24853738; http://dx.doi.org/ 10.1016/j.bpj.2014.03.044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Gravato-Nobre MJ, Nicholas HR, Nijland R, O'Rourke D, Whittington DE, Yook KJ, Hodgkin J. Multiple genes affect sensitivity of Caenorhabditis elegans to the bacterial pathogen Microbacterium nematophilum. Genetics 2005; 171:1033-45; PMID:16079230; http://dx.doi.org/ 10.1534/genetics.105.045716 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Olsen DP, Phu D, Libby LJ, Cormier JA, Montez KM, Ryder EF, Politz SM. Chemosensory control of surface antigen switching in the nematode Caenorhabditis elegans. Genes Brain Behav 2007; 6:240-52; PMID:16879619; http://dx.doi.org/ 10.1111/j.1601-183X.2006.00252.x [DOI] [PubMed] [Google Scholar]
- 45.Grenache DG, Caldicott I, Albert PS, Riddle DL, Politz SM. Environmental induction and genetic control of surface antigen switching in the nematode Caenorhabditis elegans. Proc Natl Acad Sci U S A 1996; 93:12388-93; PMID:8901591; http://dx.doi.org/ 10.1073/pnas.93.22.12388 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Labed S, Pujol N. Caenorhabditis elegans Antifungal Defense Mechanisms. J Invasive Fungal Infection 2011; 5:110-7 [Google Scholar]
- 47.Fekete C, Tholander M, Rajashekar B, Ahren D, Friman E, Johansson T, Tunlid A. Paralysis of nematodes: shifts in the transcriptome of the nematode-trapping fungus Monacrosporium haptotylum during infection of Caenorhabditis elegans. Environ Microbiol 2008; 10:364-75; PMID:18028414; http://dx.doi.org/ 10.1111/j.1462-2920.2007.01457.x [DOI] [PubMed] [Google Scholar]
- 48.Couillault C, Pujol N, Reboul J, Sabatier L, Guichou JF, Kohara Y, Ewbank JJ. TLR-independent control of innate immunity in Caenorhabditis elegans by the TIR domain adaptor protein TIR-1, an ortholog of human SARM. Nat Immunol 2004; 5:488-94; PMID:15048112; http://dx.doi.org/ 10.1038/ni1060 [DOI] [PubMed] [Google Scholar]
- 49.Engelmann I, Griffon A, Tichit L, Montanana-Sanchis F, Wang G, Reinke V, Waterston RH, Hillier LW, Ewbank JJ. A comprehensive analysis of gene expression changes provoked by bacterial and fungal infection in C. elegans. PLoS One 2011; 6:e19055; PMID:21602919; http://dx.doi.org/ 10.1371/journal.pone.0019055 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Ahren D, Tholander M, Fekete C, Rajashekar B, Friman E, Johansson T, Tunlid A. Comparison of gene expression in trap cells and vegetative hyphae of the nematophagous fungus Monacrosporium haptotylum. Microbiology 2005; 151:789-803; PMID:15758225; http://dx.doi.org/ 10.1099/mic.0.27485-0 [DOI] [PubMed] [Google Scholar]
- 51.Pujol N, Davis PA, Ewbank JJ. The Origin and Function of Anti-Fungal Peptides in C. elegans: Open Questions. Front Immunol 2012; 3:237; PMID:22870075; http://dx.doi.org/ 10.3389/fimmu.2012.00237 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Ewbank JJ. Signaling in the Immune Response. WormBook. http://www.wormbook.org: The C. elegans Research Community ed, 2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Zugasti O, Ewbank JJ. Neuroimmune regulation of antimicrobial peptide expression by a noncanonical TGF-β signaling pathway in Caenorhabditis elegans epidermis. Nat Immunol 2009; 10:249-56; PMID:19198592; http://dx.doi.org/ 10.1038/ni.1700 [DOI] [PubMed] [Google Scholar]
- 54.Zugasti O, Bose N, Squiban B, Belougne J, Kurz CL, Schroeder FC, Pujol N, Ewbank JJ. Activation of a G protein-coupled receptor by its endogenous ligand triggers the innate immune response of Caenorhabditis elegans. Nat Immunol 2014; 15:833-8; PMID:25086774; http://dx.doi.org/ 10.1038/ni.2957 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Mrochek JE, Dinsmore SR, Ohrt DW. Monitoring phenylalanine-tyrosine metabolism by high-resolution liquid chromatography of urine. Clin Chem 1973; 19:927-36; PMID:4720815 [PubMed] [Google Scholar]
- 56.Mayatepek E, Seppel CK, Hoffmann GF. Increased urinary excretion of dicarboxylic acids and 4-hydroxyphenyllactic acid in patients with Zellweger syndrome. Eur J Pediatr 1995; 154:755-6; PMID:8582432; http://dx.doi.org/ 10.1007/BF02276727 [DOI] [PubMed] [Google Scholar]
- 57.Spaapen LJ, Ketting D, Wadman SK, Bruinvis L, Duran M. Urinary D-4-hydroxyphenyllactate, D-phenyllactate and D-2-hydroxyisocaproate, abnormalities of bacterial origin. J Inherited Metabolic Dis 1987; 10:383-90; PMID:3126358; http://dx.doi.org/ 10.1007/BF01799981 [DOI] [PubMed] [Google Scholar]
- 58.Ziegler K, Kurz CL, Cypowyj S, Couillault C, Pophillat M, Pujol N, Ewbank JJ. Antifungal innate immunity in C. elegans: PKCdelta links G protein signaling and a conserved p38 MAPK cascade. Cell Host Microbe 2009; 5:341-52; PMID:19380113; http://dx.doi.org/ 10.1016/j.chom.2009.03.006 [DOI] [PubMed] [Google Scholar]
- 59.Imler JL, Hoffmann JA. Toll signaling: the TIReless quest for specificity. Nat Immunol 2003; 4:105-6; PMID:12555093; http://dx.doi.org/ 10.1038/ni0203-105 [DOI] [PubMed] [Google Scholar]
- 60.Dierking K, Polanowska J, Omi S, Engelmann I, Gut M, Lembo F, Ewbank JJ, Pujol N. Unusual regulation of a STAT protein by an SLC6 family transporter in C. elegans epidermal innate immunity. Cell Host Microbe 2011; 9:425-35; PMID:21575913; http://dx.doi.org/ 10.1016/j.chom.2011.04.011 [DOI] [PubMed] [Google Scholar]
- 61.Kiss-Toth E. Tribbles: ‘puzzling’ regulators of cell signalling. Biochem Soc Trans 2011; 39:684-7; PMID:21428962; http://dx.doi.org/ 10.1042/BST0390684 [DOI] [PubMed] [Google Scholar]
- 62.Gumienny TL, Savage-Dunn C. TGF-β signaling in C. elegans. WormBook 2013:1-34; PMID:23908056; http://dx.doi.org/ 10.1895/wormbook.1.22.2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Tan MW. Genetic and genomic dissection of host-pathogen interactions using a P. aeruginosa-C. elegans pathogenesis model. Pediatr Pulmonol 2001; 32:96-7 [Google Scholar]
- 64.Mallo GV, Kurz CL, Couillault C, Pujol N, Granjeaud S, Kohara Y, Ewbank JJ. Inducible antibacterial defense system in C. elegans. Curr Biol 2002; 12:1209-14; PMID:12176330; http://dx.doi.org/ 10.1016/S0960-9822(02)00928-4 [DOI] [PubMed] [Google Scholar]
- 65.Duclos S, Clavarino G, Rousserie G, Goyette G, Boulais J, Camossetto V, Gatti E, LaBoissiere S, Pierre P, Desjardins M. The endosomal proteome of macrophage and dendritic cells. Proteomics 2011; 11:854-64; PMID:21280226; http://dx.doi.org/ 10.1002/pmic.201000577 [DOI] [PubMed] [Google Scholar]
- 66.Blasius AL, Arnold CN, Georgel P, Rutschmann S, Xia Y, Lin P, Ross C, Li X, Smart NG, Beutler B. Slc15a4, AP-3, and Hermansky-Pudlak syndrome proteins are required for Toll-like receptor signaling in plasmacytoid dendritic cells. Proc Natl Acad Sci U S A 2010; 107:19973-8; PMID:21045126; http://dx.doi.org/ 10.1073/pnas.1014051107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Kobayashi T, Shimabukuro-Demoto S, Yoshida-Sugitani R, Furuyama-Tanaka K, Karyu H, Sugiura Y, Shimizu Y, Hosaka T, Goto M, Kato N, et al.. The Histidine Transporter SLC15A4 Coordinates mTOR-Dependent Inflammatory Responses and Pathogenic Antibody Production. Immunity 2014; 41:375-88; PMID:25238095; http://dx.doi.org/ 10.1016/j.immuni.2014.08.011 [DOI] [PubMed] [Google Scholar]
- 68.Wang C, Ahlford A, Jarvinen TM, Nordmark G, Eloranta ML, Gunnarsson I, Svenungsson E, Padyukov L, Sturfelt G, Jonsen A, et al.. Genes identified in Asian SLE GWASs are also associated with SLE in Caucasian populations. Eur J Hum Genet 2013; 21:994-9; PMID:23249952; http://dx.doi.org/ 10.1038/ejhg.2012.277 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Labed SA, Omi S, Gut M, Ewbank JJ, Pujol N. The pseudokinase NIPI-4 is a novel regulator of antimicrobial peptide gene expression. PLoS One 2012; 7:e33887; PMID:22470487; http://dx.doi.org/ 10.1371/journal.pone.0033887 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Boudeau J, Miranda-Saavedra D, Barton GJ, Alessi DR. Emerging roles of pseudokinases. Trends Cell Biol 2006; 16:443-52; PMID:16879967; http://dx.doi.org/ 10.1016/j.tcb.2006.07.003 [DOI] [PubMed] [Google Scholar]
- 71.Zeqiraj E, Filippi BM, Deak M, Alessi DR, van Aalten DM. Structure of the LKB1-STRAD-MO25 complex reveals an allosteric mechanism of kinase activation. Science 2009; 326:1707-11; PMID:19892943; http://dx.doi.org/ 10.1126/science.1178377 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Zhang Y, Li W, Li L, Li Y, Fu R, Zhu Y, Li J, Zhou Y, Xiong S, Zhang H. Structural Damage in the C. elegans Epidermis Causes Release of STA-2 and Induction of an Innate Immune Response. Immunity 2015; 42:309-20; PMID:25692704; http://dx.doi.org/ 10.1016/j.immuni.2015.01.014 [DOI] [PubMed] [Google Scholar]
- 73.Francis R, Waterston RH. Muscle cell attachment in Caenorhabditis elegans. J Cell Biol 1991; 114:465-79; PMID:1860880; http://dx.doi.org/ 10.1083/jcb.114.3.465 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Hahn BS, Labouesse M. Tissue integrity: hemidesmosomes and resistance to stress. Curr Biol 2001; 11:R858-61; PMID:11696341; http://dx.doi.org/ 10.1016/S0960-9822(01)00516-4 [DOI] [PubMed] [Google Scholar]
- 75.Zhang H, Landmann F, Zahreddine H, Rodriguez D, Koch M, Labouesse M. A tension-induced mechanotransduction pathway promotes epithelial morphogenesis. Nature 2011; 471:99-103; PMID:21368832; http://dx.doi.org/ 10.1038/nature09765 [DOI] [PubMed] [Google Scholar]
- 76.Ogawa R, Hsu CK. Mechanobiological dysregulation of the epidermis and dermis in skin disorders and in degeneration. J Cell Mol Med 2013; 17:817-22; PMID:23672502; http://dx.doi.org/ 10.1111/jcmm.12060 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Laval S, Laklai H, Fanjul M, Pucelle M, Laurell H, Billon-Gales A, Le Guellec S, Delisle MB, Sonnenberg A, Susini C, et al.. Dual roles of hemidesmosomal proteins in the pancreatic epithelium: the phosphoinositide 3-kinase decides. Oncogene 2014; 33:1934-44; PMID:23624916; http://dx.doi.org/ 10.1038/onc.2013.146 [DOI] [PubMed] [Google Scholar]