

Abstract
Many important psychological characteristics show sex differences, and are influenced by sex hormones at different developmental periods. We focus on the role of sex hormones in early development, particularly the differential effects of prenatal androgens on aspects of gender development. Increasing evidence confirms that prenatal androgens have facilitative effects on male-typed activity interests and engagement (including child toy preferences and adult careers), and spatial abilities, but relatively minimal effects on gender identity. Recent emphasis has been directed to the psychological mechanisms underlying these effects (including sex differences in propulsive movement, and androgen effects on interest in people versus things), and neural substrates of androgen effects (including regional brain volumes, and neural responses to mental rotation, sexually arousing stimuli, emotion, and reward). Ongoing and planned work is focused on understanding the ways in which hormones act jointly with the social environment across time to produce varying trajectories of gender development, and clarifying mechanisms by which androgens affect behaviors. Such work will be facilitated by applying lessons from other species, and by expanding methodology. Understanding hormonal influences on gender development enhances knowledge of psychological development generally, and has important implications for basic and applied questions, including sex differences in psychopathology, women’s underrepresentation in science and math, and clinical care of individuals with variations in gender expression.
Why are the sexes different? How does the prenatal environment set the stage for postnatal development? How does behavior result from transactions between the brain and the social world? All three questions are the focus of contemporary work in the behavioral sciences, and they converge in questions regarding prenatal sex hormone effects on gender development, which includes characteristics that show sex differences and that relate to being female or male.
Foundation
Human work linking hormones and behavior borrows heavily from work in nonhuman species showing that exposure to sex hormones early in development has permanent effects on sex-related behavior, and reproductive anatomy and function [reviewed in 1,2,3]. Thus, early development represents a sensitive period for hormones to organize the brain.1 Studying prenatal hormonal influences on gender development is challenging: hormones cannot be manipulated experimentally, and human behavior is strongly influenced by social context.
Fortunately, several methods are available to meet these challenges, as shown in Table 1. Evidence using those methods has accrued to demonstrate that levels of androgens during prenatal development are related (in varying degrees) to characteristics that show sex differences; this evidence is described in recent reviews [4–6], and summarized in Table 2 (Column 1). Recent progress, discussed below and summarized in Table 2 (Column 2), has examined the limits of the effects, and considered the psychological and neural mechanisms mediating them. Ongoing and planned work involves expansion of those themes, novel methods, and incorporation of recent lessons from animal studies. Given the challenges of studying these questions, this field moves more slowly than many others, so we extend our review beyond the past two years.
Table 1.
Key Methods for Studying Androgen Effects on Gender Development
| Natural Experiments | Strengths | Limitations | Used to Study |
| Congenital Adrenal Hyperplasia (CAH) | Separation of prenatal androgens and rearing (social) sex in girls | Physical virilization Abnormalities in other hormones (e.g., glucocorticoids) | Prenatal androgens versus rearing in girls and women |
| Complete Androgen Insensitivity Syndrome (CAIS) | Separation of prenatal androgens and sex chromosomes | Confounding of androgens and rearing (social) sex (both female-typical) | Prenatal androgens versus genes on the sex chromosomes |
| Typical Samples | Strengths | Limitations | Used to Study |
| Amniotic Hormones | Natural variations in hormones No confounding disease factors | Single sample of hormones at varying gestational ages Selected sample | Effects of within-sex variations in prenatal hormones |
| Opposite-Sex Twins | Natural variations in hormones No confounding disease factors | Unclear whether and how hormones transferred between fetuses Twins share postnatal social environment | Effects of within-sex variations in prenatal hormones |
| Genital Anatomy | Natural variations in androgens Measured postnatally | No direct evidence linking anatomy to prenatal androgens in human beings | Effects of within-sex variations in prenatal and early postnatal androgens |
| Digit Ratio | Easy to measure Differs in groups known to differ in prenatal androgen exposure | Does not reflect within-sex variation in prenatal androgen exposure | Should not be used |
Table 2.
Summary of Prenatal Androgen Effects on Gender Development
| Reviewed in 2011 | Confirmed in Recent Studies? | ||||||
|---|---|---|---|---|---|---|---|
| Effect Size | Evidence | Effect Size | Evidence | ||||
| Source | Strength | Source | Strength | ||||
| Activity Interests & Participation | large | Natural Expt Amniotic T | +++ ++ |
large | Natural Expt Amniotic T | +++ --- |
|
| Gender Identity | small | Natural Expt | +++ | small | Natural Expt | +++ | |
| Sexual Orientation | moderate | Natural Expt | +++ | Natural Expt | --- | ||
| Spatial Abilities | small-moderate | Natural Expt OS twins |
++ + |
small-moderate | Natural Expt OS Twins Amniotic T |
+++ ++ + |
|
| Psychopathology: | |||||||
| “Autistic” Traits | moderate | Natural Expt Amniotic T |
+ + |
large | Amniotic T | + | |
| Autism Diagnosis | no effect | Natural Expt | + | ||||
| Substance Use | small | OS Twins | + | ||||
| Disordered Eating | small-moderate | OS Twins | + | ||||
Modified from [4].
Source (of evidence): Natural Expt: Natural experiments (e.g., CAH); Amniotic T: Typical samples with direct measure of testosterone in amniotic fluid; OS Twins: Typical samples of opposite versus same-sex twins
Strength (of evidence), based on number of studies and ability to infer causation about androgen from design (e.g., more weight given to data from natural experiments than twins): + weak; ++ moderate; +++ strong; --- insufficient new evidence
Nature and Psychological Mechanisms of Prenatal Androgen Effects on Gendered Behavior
Confidence has increased that early androgens affect gender development, in light of recent studies that confirm, extend, and clarify previous findings. Most promising, research has moved from asking whether hormones influence human behavior to asking how they do so.
Activity interests and participation – from childhood toy preferences to adult hobbies and occupations – continue to be strongly linked to prenatal androgen exposure [e.g., 7,8], with two notable recent findings. First, androgen effects on interest and engagement in male-typed occupations was seen to have economic consequences: women with exposure to high levels of prenatal androgens due to congenital adrenal hyperplasia (CAH) were more likely than controls to have income in the top 20th percentile, reflecting employment in male-typical, higher-paying jobs (despite having lower education, and more psychosocial problems) [9]. Second, the sensitive period for androgen effects on activity interests was extended to the early postnatal months, as seen in links between parent-reported child play and urinary testosterone during the first six postnatal months [10], and penile length at a similar time (marking the postnatal testosterone surge also known as mini-puberty) [11].
Speculations about the affective and cognitive processes that underlie children’s sextyped toy preferences have been stimulated by evidence that those preferences are paralleled in rhesus monkeys (male monkeys, like boys, strongly preferred wheeled toys, whereas female monkeys, like girls, had variable preferences, leading to sex differences in preference for wheeled versus plush toys) [12, 13]. Recent work has documented early sex differences in propulsive movement (hitting versus cradling an object), with sex-typed activities suggested to develop “from socialization mechanisms that build on a male predisposition to imitate propulsive motion” [14, p. 262]. Furthermore, sex differences in occupational choices (e.g., male-predominance in science and engineering, female-predominance in social service) appear to be driven in part by androgen effects on interest in things versus people [15].
Spatial abilities are facilitated by exposure to high levels of prenatal androgens. Performance on several spatial tests was higher in two samples of females with CAH compared to typical females [16,17]. Inconsistencies in early studies were clarified: androgen facilitation of spatial abilities may be countered by adverse effects of the disease during early life [17]. Converging evidence comes from typical samples. Spatial abilities were positively related to amniotic testosterone in girls for one measure, but not two others, perhaps due to small sample [18]. Spatial performance was also higher in females with a male co-twin than a female co-twin, thought to reflect transfer of testosterone during gestation [19,20]; importantly, one study controlled for postnatal socialization effects by showing that females with a (non-twin) brother did not have better spatial ability than those with a sister [19].
Sex-related psychiatric disorders are suggested to result, in part, from sex hormones. Prenatal androgens have been invoked to explain male predominance of externalizing disorders, such as attention-deficit hyperactivity disorder (ADHD) [e.g., 21] and substance use, and of autism [e.g., 22]; it is not specified when prenatal androgens trigger ADHD or autism, although genes presumably play a role. Population registry-based studies of psychiatric morbidity in individuals with CAH in Sweden produced results not easily reconciled with early androgen effects. Although girls and women with CAH had a higher rate of substance misuse than female controls, the rate was also higher than male controls; moreover, other conditions that show female predominance (stress and adjustment disorders) were increased, but ADHD and autism spectrum disorders were not [23]. Boys and men with CAH also had higher rates of psychiatric disorder than controls [24].
Overall, the increased psychopathology seen in males and females with CAH reflects the limitations of natural experiments (i.e., other disease factors contribute to behavior), so it is important to note indirect evidence for prenatal androgen effects on some forms of sex-related psychopathology. Consistent with male predominance of substance use and female predominance of disordered eating, females with a male co-twin had more alcohol use [25,26], and less disordered eating [27] than females with a female co-twin; importantly, these studies included siblings to control for postnatal environmental effects. “Autistic” traits were associated with amniotic testosterone [28], but interpretation is not simple because some traits show sex differences in the normal range and would be expected to relate to prenatal hormones for that reason.
Neural Substrates of Prenatal Androgen Effects on Gendered Behavior
Neuroimaging has come to human behavioral neuroendocrinology, as to other areas of behavioral science, with increasing interest in aspects of brain anatomy and activation that are related to sex and sex hormones [reviewed and discussed in 29]. In a magnetic resonance imaging (MRI) study of typical boys aged 8–11, brain structures that show sex differences were examined in relation to amniotic testosterone, with effects for regional gray matter volume in some areas, but not to midsaggital corpus callosum size [30,31]. The behavioral significance of the effects is not clear (anatomy was not examined in relation to behavior). It is also unclear why girls were not studied since they were included in other aspects of the study linking amniotic testosterone to behavior.
Brain activation patterns during cognitive and affective tasks were also examined in relation to prenatal androgens in natural experiments and a typical sample, with most using functional MRI. (Few studies considered potential confounding effects of postnatal androgens.) Evidence for androgen effects on two aspects of sex-typed brain responses comes from women with complete androgen insensitivity syndrome (CAIS) who have a Y chromosome but do not have effective androgen exposure (due to lack of functional androgen receptors). In two separate samples, women with CAIS had female-typical brain responses to mental rotation [32] and to sexually-arousing stimuli [33]; their brain activation patterns were different than those of men and similar to those of typical women, consistent with their low androgen responsiveness and not with possessing a Y chromosome.
Other studies linking androgens to brain activation were not as clear. For example, in a positron emission tomography study, women with and without CAH did not differ in neural response to olfactory stimuli [34].
Several studies focused on characteristics related to psychopathology, consistent with the approach of understanding psychiatric disorders in terms of underlying dimensions of observable behavior and neurobiology [35–37]. For example, the male predominance of childhood-onset externalizing problems suggests that early androgens masculinize reward systems. In one set of studies focused on emotion in faces [38,39], girls with CAH had greater amygdala activation while viewing negative facial emotions and less hippocampal activation while recalling emotional faces than did typical girls, but the groups also differed in performance. In a study of typical boys (those whose structure was described above), amniotic testosterone was linked to striatal responses to valenced facial cues, and to behavioral approach (on a questionnaire measure of impulsivity/fun-seeking, drive, and reward responsivity) through striatal activity (but not directly to behavioral approach, perhaps due to low statistical power) [40]. Interpretation would have been enhanced with data from girls to evaluate sex differences and because testosterone is often seen to have within-sex effects in females [4].
Overall, the zeal for neuroimaging studies has yet to be matched by findings, with few clear and consistent patterns regarding androgen effects on brain structure and activation. Advances will likely come from studying neural systems underlying sex-related behavior and likely to be influenced by androgens, and directly linking brain measures to behavior. But, links between hormones and neural systems do not necessarily imply causality; for example, a brain region may be larger or more active during a specific task in women with versus without CAH simply because that region changes in response to behavior or environmental input that differs between the groups. Furthermore, a given behavioral endpoint may emerge through a variety of trajectories; androgens may affect the path to an outcome but not necessarily the outcome itself (the sexes may get to the same outcome through different paths) [e.g., 41].
Ongoing Work & Future Directions
This is an exciting time to study how hormones shape gender development. The field is poised for some significant advances, and we highlight topics that represent opportunities based on animal studies, tantalizing recent findings noted above, and related trends in other areas of science.
First, there is need to understand variations in androgen effects across behaviors and across people. For example, why is gender identity less affected by early androgens than are activity interests; what differentiates the women with CAH who are bisexual from the majority who are heterosexual; why would high prenatal androgen levels produce autism in some children, ADHD in others, and normal development in most?
Second, findings from animal studies [e.g., 41–46] can be used to guide work in human beings, as illustrated with some examples. The importance of later sensitive periods for hormone effects, such as puberty and pregnancy [e.g., 43, 46] might be tested in studies of cognition and affect in children receiving drugs to suppress puberty because of precocious puberty or gender identity concerns, and in women raising biological versus adopted children (the latter differentiates changes due to pregnancy hormones from childrearing). The evidence for a behavioral role of genes on the sex chromosomes [e.g., 42,45] can be extended to human behavior by studying women with CAIS. The notion that the sexes use different paths to get to the same outcome [e.g., 41] might be understood through studies combining neuroimaging and behavioral measures, e.g., identifying sex differences in links between brain activation during a cognitive task and performance on a related task outside the scanner.
Third, it is important to pursue additional windows into prenatal androgen exposure. Natural experiments, particularly CAH and CAIS, have yielded valuable data, but they are not perfect. (Unfortunately, most criticisms [e.g., 47] are narrow, ignore the consistency of the evidence, and fail to capture the difference between cause and effect [discussed in 48]). Methods involving direct measures of prenatal hormones (e.g., from amniotic fluid) have both promise and pitfalls [49]. Importantly, most methods relying on indirect indicators of prenatal hormones create more confusion than clarification. It is time to stop using digit ratio to mark variations in prenatal androgen exposure because such use is not supported by evidence [as detailed elsewhere, 50,51,52]. For example, in one study [50], women with CAIS who have no effective androgen exposure had only moderately feminized digit ratio compared to men, were not significantly different from typical women (who have some effective androgen exposure), and showed variability in digit ratio despite minimal variability in androgen exposure; furthermore, digit ratios did not even provide high discrimination between control men and women, despite the marked sex difference in prenatal androgen exposure. It is necessary to obtain validity data on other purported markers, such as otoacoustic emissions, before using them on a large scale [53].
Promising measures include aspects of genital anatomy, which reflect early androgen action, and thus are likely to relate to later behavior in people as they do in nonhuman animals. Anogenital distance reflects prenatal androgen exposure [54–57], although it may be modified by postnatal androgen [58], and has been linked to boys’ parent-reported play in one study [11]. Penile length, especially change during the first months of life, may mark the early postnatal testosterone surge in boys, and has also been linked to parent-reported play [5,11].
Fourth, we need to understand how hormones work jointly with socialization to influence gender development [48]. Children are socialized in ways that reflect their characteristics, and the social world does not affect all children equally with regard to gender development [e.g., 59,60]. Girls with CAH provide a unique opportunity to uncouple effects on gender development of biological and rearing sex, to ask how girls with CAH are socialized (e.g., whether they are socialized in female-typical ways consistent with their rearing sex and identity, or in atypical ways in relation to their masculinized activity interests), and whether socialization has different effects on girls with and without CAH [8,48,61]. This question is ideally suited for biomarkers: imagine what we could learn if we could easily identify children at birth in terms of their early androgen exposure and then study how they elicit different socialization and respond differently to the same socialization.
These questions represent pieces of the large puzzle to identify the neural, psychological, and developmental paths linking prenatal androgens to behavior, and studying how varying trajectories develop from the interplay of hormones and the social environment at different sensitive periods, as represented in Figure 1. The figure highlights the hypothesized biological and psychosocial contributors to gender development, and how different paths can lead to similar endpoints; for example, male-typed activity interests may result primarily from increased exposure to prenatal androgens for some girls (such as those with CAH), but from gendered socialization experiences for other girls. The figure also calls attention to the interplay between biological and social processes, and the psychological and neural mechanisms that mediate links between causes and outcomes. But, empirical evidence is needed to test most of the paths in the figure; most extant data concern simple links between behavior and prenatal androgens or gendered socialization.
Figure 1.
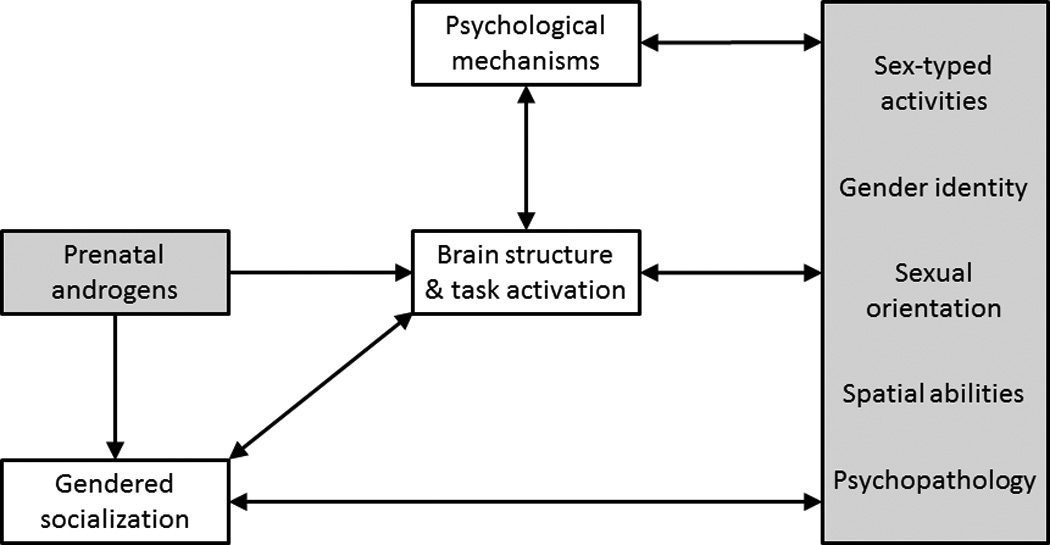
Simple process model delineating the link between prenatal androgens and sex-typed behaviors (gray boxes) by considering neural and psychological mediators of the link and including the influence of gendered socialization (white boxes) on the process. Path magnitudes change with development and vary for different behaviors, contexts, and individuals.
Relevance to Important Psychological Questions
There are broad implications of identifying how hormones shape gender development, as illustrated in Figure 1. Sex and gender are crucial to identity and a range of characteristics, psychological and physical, in health and disease [62,63]. Prenatal programming provides a window into development and, ultimately, an opportunity to facilitate optimal development [e.g., 64,65]. Gender development clearly represents the interplay of biology and socialization, so provides a nice model for psychological development more broadly. We illustrate with several examples.
First, sex matters for mental illness susceptibility, prevalence, age of onset, and form. It is likely that understanding how hormones contribute to sex-related psychopathology will provide information fundamental to the development of personalized interventions [35–37].
Second, controversy surrounds the causes of women’s underrepresentation in science, math, engineering, and technology (STEM) careers [e.g., 66,67]. There is little doubt that social structure (e.g., discrimination, child care policies) contributes to the problem [e.g., 68], but it is likely that sex differences in interests also play a role. Prenatal androgen effects on the tendency to prefer careers that involve things versus people [15] reinforce other suggestions [69,70] that women might be engaged by STEM when emphasis is placed on its social relevance.
Third, optimal care of children with variations in gender expression (e.g., disorders of sex development, transgender identity) requires more evidence on the ways that hormones are modified by genes and social factors, best studied in systematic, long-term follow-up studies [71]. A pressing question concerns the development and causes of gender identity: it is not simply related to prenatal androgens [as confirmed recently, 72,73], and appears to be plastic, with adolescence a key period for the development of nonnormative identity [74].
Conclusions
Work on hormonal influences on gender development provides a nice model for understanding psychological development in general. Identifying mechanisms by which sex and gender matter can tell us about the ways that the prenatal environment primes us to elicit and respond to our social worlds, and how our biology and experiences transact across development to shape brain structure and function that guide behavior.
Highlights.
Prenatal androgens influence sex-related characteristics to varying degrees
Androgens facilitate male-typed activities through interest in things vs. people
Androgens are associated with some aspects of brain structure and activation
Current work is focused on interplay of hormones and social environment
Relevant to questions regarding sex-related psychopathology, prenatal programming
Acknowledgements
Funding for our research was provided by grants from the National Institutes of Health HD19644, HD044398, HD057930, and MH099617. Adriene Beltz is currently supported by National Science Foundation grant 1157220.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
In nonhuman and human primates, androgens facilitate masculinization during the prenatal period [reviewed in 3,75]. Estrogens probably do not have effects during prenatal development because both sexes are exposed to estrogens from the mother [76], but they may have effects at later sensitive periods. Therefore, our terminology reflects the focus on androgens, while acknowledging the potential role for estrogens (and perhaps other hormones) at other periods.
Conflict of Interest
Nothing declared
References
- 1.de Vries GJ, Fields CT, Peters NV, Whylings J, Paul MJ. Sensitive periods for hormonal programming of the brain. Current Topics in the Behavioral Neurosciences. 2014;16:79–108. doi: 10.1007/7854_2014_286. [DOI] [PubMed] [Google Scholar]
- 2.McCarthy MM. A lumpers versus splitters approach to sexual differentiation of the brain. Frontiers in Neuroendocrinology. 2011;32:114–123. doi: 10.1016/j.yfrne.2011.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Wallen K. The Organizational Hypothesis: Reflections on the 50th anniversary of the publication of Phoenix Goy, Gerall, and Young (1959) Hormones and Behavior. 2009;55:561–565. doi: 10.1016/j.yhbeh.2009.03.009. [DOI] [PubMed] [Google Scholar]
- 4. Berenbaum SA, Beltz AM. Sexual differentiation of human behavior: Effects of prenatal and pubertal organizational hormones. Frontiers in Neuroendocrinology. 2011;32:183–200. doi: 10.1016/j.yfrne.2011.03.001. **Critically reviewed theory and evidence regarding the ways in which human sex-related behavior and brain are affected by sex hormones during early development and again in adolescence; provided recommendations for studying organizational effects of pubertal hormones in human beings.
- 5.Hines M, Constantinescu M, Spencer D. Early androgen exposure and human gender development. Biology of Sex Differences. 2015;6 doi: 10.1186/s13293-015-0022-1. article 3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Meyer-Bahlburg HFL. Brain development and cognitive, psychosocial, and psychiatric functioning in classical 21-hydroxylase deficiency. In: Ghizzoni L, Cappa M, Chrousos G, Loche S, Maghnie M, editors. Endocrine Development: Pediatric Adrenal Diseases. 2011. pp. 88–95. Endocrine Development, vol 20. [DOI] [PubMed] [Google Scholar]
- 7.Auyeung B, Baron-Cohen S, Ashwin E, Knickmeyer R, Taylor K, Hackett G, Hines M. Fetal testosterone predicts sexually differentiated childhood behavior in girls and in boys. Psychological Science. 2009;20:144–148. doi: 10.1111/j.1467-9280.2009.02279.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wong WI, Pasterski V, Hindmarsh PC, Geffner ME, Hines M. Are there parental socialization effects on the sex-typed behavior of individuals with congenital adrenal hyperplasia? Archives of Sexual Behavior. 2013;42:381–391. doi: 10.1007/s10508-012-9997-4. [DOI] [PubMed] [Google Scholar]
- 9.Strandqvist A, Falhammar H, Lichtenstein P, Hirschberg AL, Wedell A, Norrby C, Nordenskjöld A, Frisén L, Nordenström A. Suboptimal psychosocial outcomes in patients with congenital adrenal hyperplasia: Epidemiological studies in a nonbiased national cohort in Sweden. Journal of Clinical Endocrinology & Metabolism. 2014;99:1425–1432. doi: 10.1210/jc.2013-3326. [DOI] [PubMed] [Google Scholar]
- 10.Lamminmäki A, Hines M, Kuiri-Hänninen T, Kilpeläinen L, Dunkel L, Sankilampi U. Testosterone measured in infancy predicts subsequent sex-typed behavior in boys and in girls. Hormones and Behavior. 2012;61:611–616. doi: 10.1016/j.yhbeh.2012.02.013. [DOI] [PubMed] [Google Scholar]
- 11. Pasterski V, Acerini CL, Dunger DB, Ong KK, Hughes IA, Thankamony A, Hines M. Postnatal penile growth concurrent with mini-puberty predicts later sex-typed play behavior: Evidence for neurobehavioral effects of the postnatal androgen surge in typically developing boys. Hormones and Behavior. 2015;69:98–105. doi: 10.1016/j.yhbeh.2015.01.002. **Using genital markers of hormones, showed that prenatal and early postnatal testosterone in boys is related to childhood sex-typed activities; important for providing suggestive evidence regarding mini-puberty as another sensitive period in boys.
- 12.Hassett JM, Siebert ER, Wallen K. Sex differences in rhesus monkey toy preferences parallel those of children. Hormones and Behavior. 2008;54:359–364. doi: 10.1016/j.yhbeh.2008.03.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Williams CL, Pleil KE. Toy story: Why do monkey and human males prefer trucks? Comment on "Sex differences in rhesus monkey toy preferences parallel those of children" by Hassett, Siebert and Wallen. Hormones and Behavior. 2008;54:355–358. doi: 10.1016/j.yhbeh.2008.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Benenson JF, Tennyson R, Wrangham RW. Male more than female infants imitate propulsive motion. Cognition. 2011;121:262–267. doi: 10.1016/j.cognition.2011.07.006. *Provided a hypothesis and evidence regarding propulsive movement as a mechanism underlying sex differences (and early hormone effects) on sex-related activity interests and participation.
- 15. Beltz AM, Swanson JL, Berenbaum SA. Gendered occupational interests: Prenatal androgen effects on psychological orientation to Things versus People. Hormones and Behavior. 2011;60:313–317. doi: 10.1016/j.yhbeh.2011.06.002. *Provided evidence that sex differences in occupational preferences are driven in part by prenatal androgen effects on interest in things versus people.
- 16.Berenbaum SA, Bryk KLK, Beltz AM. Early androgen effects on spatial and mechanical abilities: Evidence from congenital adrenal hyperplasia. Behavioral Neuroscience. 2012;126:86–96. doi: 10.1037/a0026652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Hampson E, Rovet JF. Spatial function in adolescents and young adults with congenital adrenal hyperplasia: Clinical phenotype and implications for the androgen hypothesis. Psychoneuroendocrinology. 2015;54:60–70. doi: 10.1016/j.psyneuen.2015.01.022. *Provided an explanation, supported by evidence, for inconsistencies in previous work examining prenatal androgen effects on spatial abilities by showing that neurological status plays a role; enhanced abilities were found in females with CAH with the severe (salt-wasting) form of the disorder who had no evidence of neurological insult.
- 18.Auyeung B, Knickmeyer R, Ashwin E, Taylor K, Hackett G, Baron-Cohen S. Effects of fetal testosterone on visuospatial ability. Archives of Sexual Behavior. 2012;41:571–581. doi: 10.1007/s10508-011-9864-8. [DOI] [PubMed] [Google Scholar]
- 19.Heil M, Kavšek M, Rolke B, Beste C, Jansen P. Mental rotation in female fraternal twins: Evidence for intra-uterine hormone transfer? Biological Psychology. 2011;86:90–93. doi: 10.1016/j.biopsycho.2010.11.002. [DOI] [PubMed] [Google Scholar]
- 20.Vuoksimaa E, Kaprio J, Kremen WS, Hokkanen L, Viken RJ, Tuulio-Henriksson A, Rose RJ. Having a male co-twin masculinizes mental rotation performance in females. Psychological Science. 2010;21:1069–1071. doi: 10.1177/0956797610376075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Martel MM, Klump K, Nigg JT, Breedlove SM, Sisk CL. Potential hormonal mechanisms of Attention-Deficit/Hyperactivity Disorder and Major Depressive Disorder: A new perspective. Hormones and Behavior. 2009;55:465–479. doi: 10.1016/j.yhbeh.2009.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Baron-Cohen S, Lutchmaya S, Knickmeyer R. Prenatal testosterone in mind. Cambridge, MA: MIT Press; 2004. [Google Scholar]
- 23. Engberg H, Butwicka A, Nordenström A, Hirschberg AL, Falhammar H, Lichtenstein P, Nordenskjöld A, Frisén L, Landén M. Congenital adrenal hyperplasia and risk for psychiatric disorders in girls and women born between 1915 and 2010: A total population study. Psychoneuroendocrinology. 2015;60:195–205. doi: 10.1016/j.psyneuen.2015.06.017. *In the only population-based study of psychopathology in females with CAH, failed to show that androgens influence autism or ADHD.
- 24.Falhammar H, Butwicka A, Landen M, Lichtenstein P, Nordenskjöld A, Nordenström A, Frisén L. Increased psychiatric morbidity in men with congenital adrenal hyperplasia due to 21-hydroxylase deficiency. Journal of Clinical Endocrinology & Metabolism. 2014;99:E554–E560. doi: 10.1210/jc.2013-3707. [DOI] [PubMed] [Google Scholar]
- 25.Ellingson JM, Slutske WS, Richmond-Rakerd LS, Martin NG. Investigating the influence of prenatal androgen exposure and sibling effects on alcohol use and alcohol use disorder in females from opposite-sex twin pairs. Alcoholism: Clinical and Experimental Research. 2013;37:868–876. doi: 10.1111/acer.12035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Meyers JL, Salvatore JE, Vuoksimaa E, Korhonen T, Pulkkinen L, Rose RJ, Kaprio J, Dick DM. Genetic influences on alcohol use behaviors have diverging developmental trajectories: A prospective study among male and female twins. Alcoholism: Clinical and Experimental Research. 2014;38:2869–2877. doi: 10.1111/acer.12560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Culbert KM, Breedlove SM, Sisk CL, Keel PK, Neale MC, Boker SM, Burt SA, Klump KL. Age differences in prenatal testosterone's protective effects on disordered eating symptoms: Developmental windows of expression? Behavioral Neuroscience. 2015;129:18–36. doi: 10.1037/bne0000034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Auyeung B, Ahluwalia J, Thomson L, Taylor K, Hackett G, O'Donnell KJ, Baron-Cohen S. Prenatal versus postnatal sex steroid hormone effects on autistic traits in children at 18 to 24 months of age. Molecular Autism. 2012;3 doi: 10.1186/2040-2392-3-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Beltz AM, Blakemore JEO, Berenbaum SA. Sex differences in brain and behavioral development. In: Rubenstein JLR, Rakic P, editors. Comprehensive Developmental Neuroscience: Neural Circuit Development and Function in the Healthy and Diseased Brain. 2013. pp. 467–499. *Critically evaluated conceptual frameworks, evidence, and issues regarding human sex and sex hormone effects on brain structure and activation.
- 30.Chura LR, Lombardo MV, Ashwin E, Auyeung B, Chakrabarti B, Bullmore ET, Baron-Cohen S. Organizational effects of fetal testosterone on human corpus callosum size and asymmetry. Psychoneuroendocrinology. 2010;35:122–132. doi: 10.1016/j.psyneuen.2009.09.009. [DOI] [PubMed] [Google Scholar]
- 31.Lombardo MV, Ashwin E, Auyeung B, Chakrabarti B, Taylor K, Hackett G, Bullmore ET, Baron-Cohen S. Fetal testosterone influences sexually dimorphic gray matter in the human brain. Journal of Neuroscience. 2012;32:674–680. doi: 10.1523/JNEUROSCI.4389-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. van Hemmen J, Veltman DJ, Hoekzema E, Cohen-Kettenis PT, Dessens AB, Bakker J. Neural activation during mental rotation in complete androgen insensitivity syndrome: The influence of sex hormones and sex chromosomes. Cerebral Cortex. 2015 doi: 10.1093/cercor/bhu280. **A compelling study documenting androgen effects on neural processing of spatial ability; the first neuroimaging study of women with complete androgen insensitivity syndrome.
- 33.Hamann S, Stevens J, Vick JH, Bryk K, Quigley CA, Berenbaum SA, Wallen K. Brain responses to sexual images in 46, XY women with complete androgen insensitivity syndrome are female-typical. Hormones and Behavior. 2014;66:724–730. doi: 10.1016/j.yhbeh.2014.09.013. [DOI] [PubMed] [Google Scholar]
- 34.Ciumas C, Hirschberg AL, Savic I. High fetal testosterone and sexually dimorphic cerebral networks in females. Cerebral Cortex. 2009;19:1167–1174. doi: 10.1093/cercor/bhn160. [DOI] [PubMed] [Google Scholar]
- 35.Cuthbert BN. The RDoC framework: Facilitating transition from ICD/DSM to dimensional approaches that integrate neuroscience and psychopathology. World Psychiatry. 2014;13:28–35. doi: 10.1002/wps.20087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Cuthbert BN. Research Domain Criteria: Toward future psychiatric nosologies. Dialogues in Clinical Neuroscience. 2015;17:89–97. doi: 10.31887/DCNS.2015.17.1/bcuthbert. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Sanislow CA, Pine DS, Quinn KJ, Kozak MJ, Garvey MA, Heinssen RK, Wang PSE, Cuthbert BN. Developing constructs for psychopathology research: Research Domain Criteria. Journal of Abnormal Psychology. 2010;119:631–639. doi: 10.1037/a0020909. [DOI] [PubMed] [Google Scholar]
- 38.Ernst M, Maheu FS, Schroth E, Hardin J, Golan LG, Cameron J, Allen R, Holzer S, Nelson E, Pine DS, et al. Amygdala function in adolescents with congenital adrenal hyperplasia: A model for the study of early steroid abnormalities. Neuropsychologia. 2007;45:2104–2113. doi: 10.1016/j.neuropsychologia.2007.01.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Mazzone L, Mueller SC, Maheu F, VanRyzin C, Merke DP, Ernst M. Emotional memory in early steroid abnormalities: An fMRI study of adolescents with congenital adrenal hyperplasia. Developmental Neuropsychology. 2011;36:473–492. doi: 10.1080/87565641.2010.549866. [DOI] [PubMed] [Google Scholar]
- 40.Lombardo MV, Ashwin E, Auyeung B, Chakrabarti B, Lai MC, Taylor K, Hackett G, Bullmore ET, Baron-Cohen S. Fetal programming effects of testosterone on the reward system and behavioral approach tendencies in humans. Biological Psychiatry. 2012;72:839–847. doi: 10.1016/j.biopsych.2012.05.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.De Vries GJ. Minireview: Sex differences in adult and developing brains: Compensation, compensation, compensation. Endocrinology. 2004;145:1063–1068. doi: 10.1210/en.2003-1504. [DOI] [PubMed] [Google Scholar]
- 42.Arnold AP, Chen X. What does the "four core genotypes" mouse model tell us about sex differences in the brain and other tissues? Frontiers in Neuroendocrinology. 2009;30:1–9. doi: 10.1016/j.yfrne.2008.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Macbeth AH, Luine VN. Changes in anxiety and cognition due to reproductive experience: A review of data from rodent and human mothers. Neuroscience and Biobehavioral Reviews. 2010;34:452–467. doi: 10.1016/j.neubiorev.2009.08.011. [DOI] [PubMed] [Google Scholar]
- 44. McCarthy MM, Arnold AP. Reframing sexual differentiation of the brain. Nature Neuroscience. 2011;14:677–683. doi: 10.1038/nn.2834. **Important discussion expanding early ideas about sexual differentiation, including concepts and evidence showing complex interplay of genes, hormones, and environment; focused on work in rodents, but has important implications for human gender development.
- 45.Ngun TC, Ghahramani N, Sánchez FJ, Bocklandt S, Vilain E. The genetics of sex differences in brain and behavior. Frontiers in Neuroendocrinology. 2011;32:227–246. doi: 10.1016/j.yfrne.2010.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Schulz KM, Molenda-Figueira HA, Sisk CL. Back to the future: The organizational-activational hypothesis adapted to puberty and adolescence. Hormones and Behavior. 2009;55:597–604. doi: 10.1016/j.yhbeh.2009.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Jordan-Young RM. Hormones, context, and"brain gender": A review of evidence from congenital adrenal hyperplasia. Social Science & Medicine. 2012;74:1738–1744. doi: 10.1016/j.socscimed.2011.08.026. [DOI] [PubMed] [Google Scholar]
- 48. Berenbaum SA, Blakemore JEO, Beltz AM. A role for biology in gender-related behavior. Sex Roles. 2011;64:804–825. *Discussed the ways in which evidence about hormonal influences on behavior and the brain is misunderstood and misused; highlighted opportunities for studying how the interplay between biology and socialization shapes gender development.
- 49. Constantinescu M, Hines M. Relating prenatal testosterone exposure to postnatal behavior in typically developing children: Methods and findings. Child Development Perspectives. 2012;6:407–413. *Reviewed methods and evidence regarding behavioral effects of normal variations in prenatal hormones in typical samples.
- 50. Berenbaum SA, Bryk KK, Nowak N, Quigley CA, Moffat S. Fingers as a marker of prenatal androgen exposure. Endocrinology. 2009;150:5119–5124. doi: 10.1210/en.2009-0774. **Using data from women with absence of effective androgen exposure, showed that digit ratio is not a good index of individual differences in prenatal androgen levels; provides strong evidence against the validity of digit ratio as a marker of androgen exposure in typical samples.
- 51.Voracek M. No effects of androgen receptor gene CAG and GGC repeat polymorphisms on digit ratio (2D:4D): A comprehensive meta-analysis and critical evaluation of research. Evolution and Human Behavior. 2014;35:430–437. [Google Scholar]
- 52.Wallen K. Does finger fat produce sex differences in second to fourth digit ratios? Endocrinology. 2009;150:4819–4822. doi: 10.1210/en.2009-0986. [DOI] [PubMed] [Google Scholar]
- 53.Wisniewski AB, Espinoza-Varas B, Aston CE, Edmundson S, Champlin CA, Pasanen EG, McFadden D. Otoacoustic emissions, auditory evoked potentials and self-reported gender in people affected by disorders of sex development (DSD) Hormones and Behavior. 2014;66:467–474. doi: 10.1016/j.yhbeh.2014.07.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Barrett ES, Parlett LE, Sathyanarayana S, Liu F, Redmon JB, Wang C, Swan SH. Prenatal exposure to stressful life events is associated with masculinized anogenital distance (AGD) in female infants. Physiology & Behavior. 2013;114:14–20. doi: 10.1016/j.physbeh.2013.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Barrett ES, Parlett LE, Redmon JB, Swan SH. Evidence for sexually dimorphic associations between maternal characteristics and anogenital distance, a marker of reproductive development. American Journal of Epidemiology. 2014;179:57–66. doi: 10.1093/aje/kwt220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Dean A, Sharpe RM. Anogenital distance or digit length ratio as measures of fetal androgen exposure: Relationship to male reproductive development and its disorders. Journal of Clinical Endocrinology & Metabolism. 2013;98:2230–2238. doi: 10.1210/jc.2012-4057. **Discussed the value of anogenital distance as an indicator of prenatal androgen exposure useful for studies of postnatal reproductive health; principles can be extended for use in studies of behavior.
- 57.Sathyanarayana S, Grady R, Redmon JB, Ivicek K, Barrett E, Janssen S, Nguyen R, Swan SH, Team TS. Anogenital distance and penile width measurements in The Infant Development and the Environment Study (TIDES): Methods and predictors. Journal of Pediatric Urology. 2015;11:76.e1–76.e6. doi: 10.1016/j.jpurol.2014.11.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Mitchell RT, Mungall W, McKinnell C, Sharpe RM, Cruickshanks L, Milne L, Smith LB. Anogenital distance plasticity in adulthood: Implications for its use as a biomarker of fetal androgen action. Endocrinology. 2015;156:24–31. doi: 10.1210/en.2014-1534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Galambos NL, Berenbaum SA, McHale SM. Gender development in adolescence. In: Lerner RM, Steinberg L, editors. Handbook of adolescent psychology: Vol. 1: Individual bases of adolescent development. edn 3rd. Wiley; 2009. pp. 305–357. [Google Scholar]
- 60.McHale SM, Kim JY, Dotterer AM, Crouter AC, Booth A. The development of gendered interests and personality qualities from middle childhood through adolescence: A biosocial analysis. Child Development. 2009;80:482–495. doi: 10.1111/j.1467-8624.2009.01273.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Beltz AM, Bryk KL, Speiser P, Berenbaum SA. Dose-dependent prenatal androgen influences on gender development in girls with congenital adrenal hyperplasia. Society for Behavioral Neuroendocrinology annual meeting; 2012, June; Madison, WI. [Google Scholar]
- 62.Blakemore JEO, Berenbaum SA, Liben LS. Gender development. New York: Psychology Press / Taylor & Francis; 2009. [Google Scholar]
- 63.Wizemann TM, Pardue ML. Exploring the biological contributions to human health: Does sex matter? Washington, DC: National Academy Press; 2001. Committee on Understanding the Biology of Sex and Gender Differences (Ed) [PubMed] [Google Scholar]
- 64.Birnbaum LS, Miller MF. Prenatal programming and toxicity (PPTOX) introduction. Endocrinology. 2015 doi: 10.1210/en.2015-1458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Grandjean P, Barouki R, Bellinger DC, Casteleyn L, Chadwick LH, Cordier S, Etzel RA, Gray KA, Ha EH, Junien C, et al. Life-long implications of developmental exposure to environmental stressors: New perspectives. Endocrinology. 2015 doi: 10.1210/EN.2015-1350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Ceci SJ, Williams WM. Sex differences in math-intensive fields. Current Directions in Psychological Science. 2010;19:275–279. doi: 10.1177/0963721410383241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Wang MT, Eccles JS, Kenny S. Not lack of ability but more choice: Individual and gender differences in choice of careers in science, technology, engineering, and mathematics. Psychological Science. 2013;24:770–775. doi: 10.1177/0956797612458937. [DOI] [PubMed] [Google Scholar]
- 68.Moss-Racusin CA, Dovidio JF, Brescoll VL, Graham MJ, Handelsman J. Science faculty's subtle gender biases favor male students. Proceedings of the National Academy of Sciences of the United States of America. 2012;109:16474–16479. doi: 10.1073/pnas.1211286109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Diekman AB, Brown ER, Johnston AM, Clark EK. Seeking congruity between goals and roles: A new look at why women opt out of science, technology, engineering, and mathematics careers. Psychological Science. 2010;21:1051–1057. doi: 10.1177/0956797610377342. [DOI] [PubMed] [Google Scholar]
- 70.Diekman AB, Clark EK, Johnston AM, Brown ER, Steinberg M. Malleability in communal goals and beliefs influences attraction to STEM careers: Evidence for a goal congruity perspective. Journal of Personality and Social Psychology. 2011;101:902–918. doi: 10.1037/a0025199. [DOI] [PubMed] [Google Scholar]
- 71.Freund LS, Taymans S, Wehr E, Stratakis CA. Developmental focus on disorders of sex development. Hormone and Metabolic Research. 2015;47:309–400. doi: 10.1055/s-0035-1549880. [DOI] [PubMed] [Google Scholar]
- 72.Pasterski V, Zucker KJ, Hindmarsh PC, Hughes IA, Acerini C, Spencer D, Neufeld S, Hines M. Increased cross-gender identification independent of gender role behavior in girls with congenital adrenal hyperplasia: Results from a standardized assessment of 4-to 11-year-old children. Archives of Sexual Behavior. 2015;44:1363–1375. doi: 10.1007/s10508-014-0385-0. [DOI] [PubMed] [Google Scholar]
- 73.Jürgensen M, Kleinemeier E, Lux A, Steensma TD, Cohen-Kettenis PT, Hiort O, Thyen U, Köhler B DSD Network Working Group. Psychosexual development in adolescents and adults with disorders of sex development: Results from the German Clinical Evaluation Study. Journal of Sexual Medicine. 2013;10:2703–2714. doi: 10.1111/j.1743-6109.2012.02751.x. [DOI] [PubMed] [Google Scholar]
- 74.Steensma TD, Kreukels BPC, de Vries ALC, Cohen-Kettenis PT. Gender identity development in adolescence. Hormones and Behavior. 2013;64:288–297. doi: 10.1016/j.yhbeh.2013.02.020. [DOI] [PubMed] [Google Scholar]
- 75.Cohen-Bendahan CCC, van de Beek C, Berenbaum SA. Prenatal sex hormone effects on child and adult sex-typed behavior: Methods and findings. Neuroscience and Biobehavioral Reviews. 2005;29:353–384. doi: 10.1016/j.neubiorev.2004.11.004. [DOI] [PubMed] [Google Scholar]
- 76.Grumbach MM, Hughes IA, Conte FA. Disorders of sex differentiation. In: Larsen PR, Kronenberg HM, Melmed S, Polonsky KS, editors. Williams textbook of endocrinology. edn 10th. W.B. Saunders; 2003. pp. 842–1002. [Google Scholar]