

Abstract
To determine the effects of plant density and row spacing on the mechanical harvesting of rapeseed (Brassica napus L.), field experiments were conducted. Higher plant density produced fewer pods and reduced the yield per plant. Wider row spacing at higher plant densities increased seeds per pod and the 1000-seed weight, resulting in a higher yield per plant. The highest yields were achieved at a density of 45 × 104 plants ha−1 (D45) in combination with 15 cm row spacing (R15) because mortality associated with competition increased as both the plant density and row spacing increased. The leaf area index (LAI) and pod area index (PAI) showed similar relations to the yield per hectare, and they were positively correlated with the percentage of intercepted light, whereas the radiation use efficiency (RUE) was positively correlated with population biomass. Reduced plant height and increased root/shoot ratios led to a decreased culm lodging index. Improved resistance to pod shattering was also observed as plant density and row spacing increased. The angle of the lowest 5 branches decreased as row spacing increased under D30 and D45. All of these structural changes influenced the mechanical harvesting operations, resulting in the highest yield of mechanically harvesting rapeseed under D45R15.
Conventional rapeseed (Brassica napus L.) production in China has been dependent on manual practices for decades. However, hand-harvesting of rapeseed is labor-intensive, and the supply of dependable, skilled labor is of concern to rapeseed producers in China because the proportion of domestic labor resources engaged in crop farming is sharply decreasing1. Mechanization is an important and efficient tool for enhancing crop yield; it also reduces the labor associated with crop production and ultimately increases farmers’ prosperity2. In fact, mechanized direct-seeding has increasingly been practiced in the major canola-producing regions of China, but the development of mechanized harvesting has lagged behind. Currently, the lower rate of mechanization and the greater emphasis on human labor for harvesting are major factors restricting rapeseed production in China3. China will not remain competitive in the world market for rapeseed with the current lack of research on mechanical harvesting. Therefore, there is an immediate need to develop a mechanized harvesting system for rapeseed production.
Rapeseed yield and mechanical harvesting efficiency have been widely affected by agronomic practices, among which, plant density and row spacing have played a vital role in obtaining higher yields4. Thus, it is important to determine the appropriate plant density and row spacing that optimize both the seed yield and mechanical harvesting efficiency. Numerous researchers have investigated the effects of plant density and row spacing on agronomic traits and the yield of rapeseed. Research to determine the optimal plant density in combination with row spacing for the maximum mechanized production of rapeseed has been inconclusive because the results vary depending on the location, cultivar, soil type and local climate.
Rapeseed growth, yield and yield components are directly affected by plant density5. In general, plant densities of 60–70 plants m−2 are considered to be optimal6 for rapeseed hybrids in Europe, whereas the typical plant density of hybrid rapeseed in China is approximately 30 plants m−2 7,8,9. As plant densities decline, the reduction in the number of plants per unit area is partially compensated for by an accompanying increase in the productivity of each plant, as a result of greater leaf area; more branches; and a greater number of pods per plant10. The rapeseed yield typically exhibits a quadratic response to plant density, with a near-linear increase across a range of low densities, a gradual decrease in the rate of yield increase, and finally, a maximum yield at the optimum plant density, which depends upon crop species, environmental conditions and agronomic factors5,7,10. Leach et al. (1999) found the maximum seed yield of winter rapeseed occurred at a density of 50–60 plants m−2 in a series of multi-factorial experiments10, whereas a field experiment by Momoh and Zhou (2001) found that the highest seed yield of transplanted winter rapeseed occurred at plant densities of 9.75 × 104 and 12.75 × 104 plant ha−1 7. Other studies have reported that different seeding rates (7–14 kg ha−1) had no significant effect on seed yield11.
Compared with conventional wide row spacing, rapeseed produced in narrow rows generally has superior yields when conditions are favorable. Narrow rows increase the total seasonal light interception, make more efficient use of available light and should allow for more rapid canopy closure and shading of the ground, thereby improving weed control12. Christensen and Drabble reported that both Brassica napus and Brassica rapa showed a higher grain yield with narrower row spacing (7.5 cm) compared with wider spacing (15 and 23 cm)11. Within a certain range, the yield of winter rapeseed increased with narrower row spacing from 30 to 15 cm or from 35 to 17.5 cm13,14. Similar results were obtained with cotton (Gossypium hirsutum L.)15, soybeans (Glycine max L.)16,17 and corn (Zea mays L.)12,18. However, the rapeseed yield will decrease when the row spacing is set much narrower than 15 cm. Shanhin and Valiollah (2009) showed that the seed yield of winter rapeseed grown at a 12 cm row spacing was lower than winter rapeseed grown at 24 cm. Moreover, mechanical cultivation, fertilizer application and post-emergence herbicide treatments are more difficult in narrow row cultivation19.
Rapeseed with greater resistance to lodging and pod shattering would permit the harvester to operate more efficiently. Lodging and pod-shatter resistance could be improved with appropriate plant densities and row spacing. At too high a density, rapeseed is often more susceptible to lodging10. However, Robinson (1986) noted more lodging at lower densities because of larger, heavier stems20. In general, higher plant densities are more suitable for the mechanical harvesting of grain amaranth compared to lower densities. This is because higher densities promote less branching, fewer secondary seed heads, smaller stalk diameters, and more uniform maturation21. However, high population densities may be less suitable in more arid climates due to greater competition for available soil moisture22. Lodging increased significantly with an increase in plant density at both 1.0 m and 0.5 m row spacings in soybeans (Glycine max L.), and more severe stem lodging and higher plant mortality were observed for high plant densities at the 1.0 m row spacing compared with the 0.5 m row spacing16, whereas other results have reported that lodging was reduced by increased row spacing in high-yielding irrigated wheat23. The use of narrow rows resulted in lower plant mortality and associated stand losses during the growing season and less plant lodging in grain amaranth (Amaranthus spp.)18. Information on changes in the pattern of pod shattering under different plant densities and row spacings, which are critical to the mechanical harvesting of rapeseed, is very limited.
There have been few published studies on the response of mechanical harvesting traits in rapeseed to changes in plant density and row spacing. Information on plant density, row spacing, and their possible interaction has not been well described. In addition, inconsistent results from previously published studies suggest that further research is needed. Growing rapeseed in alternating rows with different plant densities has the potential to increase seed yield and promote the mechanical harvesting efficiency. Consequently, the objective of this study was to determine the effects of plant density and row spacing on the rapeseed yield and mechanical harvesting properties. A balance was sought between conditions that facilitated mechanical harvesting, but did not adversely affect the yield of rapeseed by altering the plant density and row spacing required for maximizing yield.
Results
Weather data
The amount of monthly sunshine and the cumulative temperature of both the seedling to wintering stage and the flowering to pod-filling stage were higher in 2013–2014 than in 2012–2013, but monthly precipitation was lower in 2013–2014. A similar trend in monthly sunshine, cumulative temperature and monthly precipitation was observed over the entire growth period (Table 1).
Table 1. Meteorological conditions during the growing seasons of oilseed rape in 2012–2014.
| Year | Meteorological factors | Seedling to wintering stage | Wintering to flowering stage | Flowering to pod filling stage | The whole growth stages |
|---|---|---|---|---|---|
| 2012–2013 | Monthly Precipitation (mm) | 78.3 | 77.4 | 169.5 | 885.2 |
| Monthly sunshine duration (h) | 132.6 | 105.3 | 154.7 | 1188.4 | |
| Accumulated temperature (°C) | 1180.9 | 315.5 | 943.1 | 2439.5 | |
| 2013–2014 | Monthly Precipitation (mm) | 64.4 | 82.7 | 152.4 | 791.9 |
| Monthly sunshine duration (h) | 174.3 | 87.9 | 147.6 | 1342.3 | |
| Accumulated temperature (°C) | 1437.1 | 261.2 | 1162.2 | 2860.5 |
Yield and yield components
The mature plant rate was significantly affected by plant density and row spacing. In general, the mature plant rate decreased with increasing plant density and row spacing; the highest mature plant rate was observed under D15R25, where the plant spacing was 26.7 cm. At the same plant density, a smaller difference between row spacing and plant spacing produced a higher plant maturity rate (Table 2).
Table 2. The plant spacing and mature plant rate under different densities and row spacing arrangements during 2012–2014.
| Density | Row Spacing | Plant spacing (cm) | Maturity rate (%) |
|
|---|---|---|---|---|
| 2012–2013 | 2013–2014 | |||
| D15 | R15 | 44.5 | 93.07c | 90.09b |
| R25 | 26.7 | 98.10a | 96.84a | |
| R35 | 19.1 | 96.27b | 92.22b | |
| D30 | R15 | 22.2 | 90.67d | 86.55c |
| R25 | 13.3 | 84.99e | 81.12d | |
| R35 | 9.5 | 80.03f | 74.62e | |
| D45 | R15 | 14.8 | 85.20e | 78.48d |
| R25 | 8.9 | 78.63f | 71.70e | |
| R35 | 6.4 | 70.30g | 64.00f | |
Values followed by different letters within the same column are significantly different according to the least significant difference (LSD) test (P < 0.05); Each data represents the mean of three replications; D15, D30 and D45 mean densities of 15, 30 and 45 × 104 plants ha−1; R15, R25 and R35 mean inter-row spacing of 15, 25 and 35 cm.
The seed yield and yield components varied significantly with changes in plant density and row spacing. Under the same row spacing, the number of pods per plant, seeds per pod, 1000-seed weight and seed yield per plant all decreased with increasing plant density, whereas the seed yield per hectare increased. In contrast, under the same plant density (D30 and D45), the number of seeds per pod, the 1000-seed weight and the seed yield per plant increased with increasing row spacing, whereas the seed yield per hectare significantly decreased. Among all of the treatments, D45R15 consistently produced the highest seed yield, while D15R15 had the lowest seed yield in both growing seasons. Population biomass showed the same trend as seed yield. Increasing plant density significantly decreased the biomass per plant, whereas plant spacing had a smaller effect on biomass, particularly under higher plant densities (D30 and D45). Significant interactions between row spacing and plant density were found for seed yield and yield components (Table 3).
Table 3. Seed yield and yield components of winter rapeseed under different densities and row spacing arrangements during 2012–2014.
| Year | Density | Row Spacing | Biomass per plant (g) | Population biomass (kg ha−1) | Pods per plant | Seeds per pod | 1000-seed weight (g) | Seed yield per plant (g) | Manually seed yield (kg ha−1) |
|---|---|---|---|---|---|---|---|---|---|
| 2012–2013 | D15 | R15 | 84.74a | 11829.7e | 303.4a | 18.3bc | 3.48a | 19.31a | 2616.0h |
| R25 | 79.53b | 11702.8e | 290.8b | 18.9a | 3.40b | 18.68b | 2689.5g | ||
| R35 | 81.42b | 11757.0e | 295.1ab | 18.5b | 3.46a | 18.89ab | 2658.0g | ||
| D30 | R15 | 50.46c | 13725.1b | 201.9c | 17.7d | 3.36cd | 12.00d | 3190.5d | |
| R25 | 51.58c | 13151.9c | 204.3c | 18.0cd | 3.38bc | 12.43cd | 3088.5e | ||
| R35 | 52.78c | 12672.5d | 208.4c | 18.3b | 3.39b | 12.93c | 2997.0f | ||
| D45 | R15 | 38.35d | 14703.4a | 181.4d | 16.1c | 3.27f | 9.55f | 3543.0a | |
| R25 | 38.75d | 13711.7b | 183.5d | 16.4f | 3.31e | 9.96f | 3387.0b | ||
| R35 | 40.30d | 12748.9d | 186.6d | 17.0e | 3.34d | 10.59e | 3243.0c | ||
| Coefficient of variation (CV%) | 33.20 | 8.05 | 22.76 | 5.52 | 1.98 | 29.07 | 10.95 | ||
| Mean | 57.55 | 12889.2 | 228.4 | 17.7 | 3.38 | 13.82 | 3046.5 | ||
| 2013–2014 | D15 | R15 | 98.89a | 13363.9cd | 324.7a | 19.3bc | 3.33a | 20.78a | 2721.0h |
| R25 | 87.91c | 12770.2d | 307.0c | 20.1a | 3.22c | 19.86b | 2814.0g | ||
| R35 | 91.86b | 12706.9d | 312.9b | 19.6b | 3.28b | 20.10b | 2746.5h | ||
| D30 | R15 | 55.34e | 14369.0b | 213.8f | 18.8d | 3.18def | 12.78e | 3220.5d | |
| R25 | 57.39de | 13965.9bc | 222.5e | 19.0cd | 3.19cde | 13.48d | 3183.0e | ||
| R35 | 59.37d | 13290.6cd | 231.4d | 19.1cd | 3.21cd | 14.10c | 3094.5f | ||
| D45 | R15 | 42.92g | 15156.9a | 196.8h | 16.9f | 3.11g | 10.34h | 3612.0a | |
| R25 | 44.92fg | 14493.4b | 202.3g | 17.2f | 3.15f | 10.96g | 3465.0b | ||
| R35 | 47.36f | 13640.4c | 211.9f | 17.7e | 3.17ef | 11.88f | 3340.5c | ||
| CV% | 33.39 | 5.96 | 21.07 | 5.95 | 2.08 | 27.92 | 10.19 | ||
| Mean | 65.11 | 13750.8 | 247.0 | 18.6 | 3.20 | 14.92 | 3133.5 | ||
| Analysis of variance | Year (Y) | 125.3** | 32.4* | 41.4* | 180.9** | 477.3** | 46.3* | 1685.0** | |
| Density (D) | 4685.3** | 136.3** | 4473.9** | 305.4** | 269.1** | 5291.5** | 4650.4** | ||
| Row spacing (R) | 10.0** | 76.1** | 9.5** | 51.7** | 27.6** | 23.1** | 243.5** | ||
| Y × D | 18.9** | 3.5NS | 0.4NS | 1.6NS | 1.5NS | 1.6NS | 2.5NS | ||
| Y × R | 0.9NS | 0.1NS | 2.5NS | 1.4NS | 1.5NS | 0.6NS | 3.5* | ||
| D × R | 22.4** | 11.3** | 15.0** | 16.5** | 45.2** | 19.6** | 116.1** | ||
| Y × D × R | 3.0* | 1.8NS | 1.3NS | 1.0NS | 0.8NS | 0.8NS | 2.4NS |
Values followed by different letters within the same column are significantly different according to the least significant difference (LSD) test (P < 0.05); Each data represents the mean of three replications.
NS means not significant; *and **means significant differences at 0.05 and 0.01 probability levels, respectively; D15, D30 and D45 mean densities of 15, 30 and 45 × 104 plants ha−1; R15, R25 and R35 mean inter-row spacing of 15, 25 and 35 cm.
Leaf area index, pod area index and harvest index
The leaf area index (LAI), pod area index (PAI) and harvest index (HI) were all significantly affected by plant density and row spacing, but plant density had a greater impact on these variables (Fig. 1). The LAI and PAI increased significantly with increasing plant density. The effect of row spacing on the LAI and PAI was influenced by plant density. Under lower plant density (D15), the highest LAI and PAI were obtained in R25 followed by R35 and R15. Under higher plant densities (D30 and D45), the LAI and PAI both decreased with increasing row spacing. In all of the treatments, the combination of D45 plant density and R15 row spacing showed the highest LAI and PAI, whereas the combination of D15 plant density and R15 row spacing showed the lowest LAI and PAI in both growing seasons. A different trend was observed for the HI. In general, the HI increased significantly with increasing plant density and row spacing, with the highest and lowest values observed under D45R35 and D15R15, respectively.
Figure 1. LAI, PAI and HI for direct-seedling winter rapeseed under different densities and row spacing arrangements during 2012–2014.

D15, D30 and D45 mean densities of 15, 30 and 45 × 104 plants ha−1; R15, R25 and R35 mean inter-row spacing of 15, 25 and 35 cm. Different letters denote significant difference at P < 0.05 by LSD.
Light interception and radiation use efficiency (RUE)
The crop canopies intercepted approximately 80–90% of the incident radiation during the entire growing period. Generally, radiation interception increased with increasing plant densities and row spacing, reaching a peak of approximately 90% interception in 35 cm rows combined with 45 × 104 plants ha−1. Under D15, the RUE increased with increasing row spacing, whereas under D30 and D45 conditions, a higher RUE was found in 15 cm rows. A similar trend was observed in above-ground biomass per m2 (Table 4). Further analyses revealed that percent interception was positively correlated with the LAI (R2 = 0.5168**) and PAI (R2 = 0.5037**), whereas population biomass was positively correlated with the RUE (R2 = 0.4153**) (Fig. 2).
Table 4. Radiation use efficiency and its related parameters of winter rapeseed under different densities and row spacing arrangements during 2012–2014.
| Year | Density | Row Spacing | Incident radiation (MJ m−2) | Intercepted radiation (MJ m−2) | Intercepted percent (%) | Total aboveground biomass (g m−2) | Radiation use efficiency (g MJ−1) |
|---|---|---|---|---|---|---|---|
| 2012–2013 | D15 | R15 | 1312.5 | 1030.0cd | 78.5f | 1064.1f | 1.03f |
| R25 | 1218.4 | 964.7g | 79.2f | 1029.4h | 1.07d | ||
| R35 | 1292.1 | 996.2f | 80.6e | 1048.4g | 1.05e | ||
| D30 | R15 | 1266.9 | 1027.0cd | 81.1e | 1175.6b | 1.15b | |
| R25 | 1238.3 | 1009.1e | 84.1d | 1123.7d | 1.11c | ||
| R35 | 1226.7 | 1019.7de | 86.5b | 1079.1e | 1.06de | ||
| D45 | R15 | 1208.9 | 1032.5c | 85.4c | 1238.7a | 1.20a | |
| R25 | 1312.6 | 1138.0a | 87.2b | 1149.8c | 1.01g | ||
| R35 | 1243.3 | 1108.3b | 89.1a | 1062.7f | 0.96h | ||
| Mean | 1257.7 | 1036.2 | 83.5 | 1107.9 | 1.07 | ||
| 2013–2014 | D15 | R15 | 1337.7 | 951.6h | 82.7g | 1062.1h | 1.12c |
| R25 | 1389.5 | 1126.6b | 83.9f | 1186.1d | 1.05f | ||
| R35 | 1353.7 | 1125.6b | 83.2g | 1110.4g | 0.99h | ||
| D30 | R15 | 1280.9 | 986.3g | 84.2f | 1115.0f | 1.13b | |
| R25 | 1302.8 | 1105.5c | 89.7d | 1208.2b | 1.09d | ||
| R35 | 1259.5 | 1089.7d | 90.4c | 1168.3e | 1.07e | ||
| D45 | R15 | 1266.5 | 1014.6f | 87.1e | 1257.3a | 1.24a | |
| R25 | 1358.3 | 1032.6e | 91.0b | 1107.9g | 1.07e | ||
| R35 | 1284.4 | 1160.3a | 92.7a | 1199.4c | 1.03g | ||
| Mean | 1314.8 | 1065.9 | 87.2 | 1157.2 | 1.09 |
Values followed by different letters within the same column are significantly different according to the least significant difference (LSD) test (P < 0.05); Each data represents the mean of three replications.
D15, D30 and D45 mean densities of 15, 30 and 45 × 104 plants ha−1; R15, R25 and R35 mean inter-row spacing of 15, 25 and 35 cm.
Figure 2.

Relationships between (a) intercepted percent and LAI, PAI; (b) population biomass and RUE for direct-seedling winter rapeseed under different densities and row spacing arrangements (R20.05 = 0.2191, R20.01 = 0.3477).
Mechanical harvesting properties
Overall, higher plant density significantly reduced plant height, snapping resistance, angle of lodging and culm lodging index, whereas the pod-shatter resistance increased at higher plant densities. Under higher plant densities (D30 and D45), the plant height, angle of lodging and culm lodging index clearly decreased, but the snapping resistance, pod-shatter resistance and seed moisture increased with increasing row spacing. The lowest plant height, angle of lodging and culm lodging index were observed under the D45R35 combination. Root biomass increased with row spacing under higher plant densities (D30 and D45). The maximum root biomass was observed under D15R25, whereas the minimum root biomass was found under D45R15. The root/shoot ratio was maximized under the highest plant density and the widest row spacing (D45R35) and minimized under D15R15. Under the lowest plant density (D15), the branching angle of the five lowest branches did not change with row spacing; however, it significantly decreased with increasing row spacing under higher plant densities (D30 and D45). In all treatments, D15R15 produced the lowest mechanical harvesting yield in both seasons. The mechanical harvesting yield increased significantly with plant density and row spacing. The highest mechanical harvesting seed yield with the lowest yield loss was obtained with the combination of R15 row spacing and D45 plant density. An analysis of variance revealed that mechanical harvesting properties were markedly affected by year, plant density and row spacing, with density having the greatest impact. Interactions between plant density and row spacing were significant for all of the evaluated mechanical harvesting properties (Table 5).
Table 5. Mechanical harvesting yield and mechanical properties of winter rapeseed under different densities and row spacing arrangements during 2012–2014.
| Year | Density | Row spacing | Plant height (cm) | Root biomass (g) | Root/Shoot | Branching angle (°) |
Snapping resistance (N) | Angle of lodging (°) | Culm lodging Index (g.cm N−1) | Pod shatter resistance | Seed moisture (%) | Pod wall biomass (g) | Mechanical harvesting yield (kg ha−1) | Total yield loss (%) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| the 1st lowest branch | the 3rd lowest branch | the 5th lowest branch | ||||||||||||||
| 2012–2013 | D15 | R15 | 164.6bc | 8.56b | 0.101e | 44.65a | 42.65a | 36.59a | 181.1a | 23.8b | 103.4a | 0.029f | 16.10a | 0.036g | 2343.5i | 10.42a |
| R25 | 167.9a | 9.62a | 0.121d | 45.63a | 42.71a | 36.52a | 170.4b | 25.9a | 104.1a | 0.023i | 15.94a | 0.040f | 2434.0g | 9.49c | ||
| R35 | 166.3ab | 8.86b | 0.109e | 44.44a | 42.13a | 36.21a | 174.9b | 24.3b | 103.9a | 0.025h | 15.99a | 0.042ef | 2394.0h | 9.94b | ||
| D30 | R15 | 164.1bc | 7.27d | 0.144c | 44.38a | 41.78a | 35.06b | 129.4e | 19.4c | 89.3b | 0.028g | 14.61bc | 0.043de | 2927.5d | 8.24ef | |
| R25 | 162.3c | 7.53cd | 0.146c | 42.50b | 40.51b | 34.48bc | 136.9d | 18.4d | 87.1b | 0.032d | 14.79b | 0.044cd | 2825.0e | 8.53e | ||
| R35 | 159.2d | 7.86c | 0.149c | 41.33bc | 39.23cd | 33.29cd | 148.2c | 15.9e | 84.2c | 0.034c | 14.90b | 0.044cd | 2725.5f | 9.05d | ||
| D45 | R15 | 157.3d | 6.06f | 0.158b | 41.56bc | 39.41c | 33.34cd | 118.8f | 15.4e | 76.8d | 0.031e | 13.89d | 0.045bc | 3297.0a | 6.94h | |
| R25 | 154.1e | 6.28f | 0.162ab | 40.45c | 38.37d | 32.48d | 123.7ef | 7.3f | 73.1e | 0.038b | 14.10cd | 0.046ab | 3132.0b | 7.53g | ||
| R35 | 150.9f | 6.75e | 0.168a | 38.08d | 36.17e | 30.28e | 127.8e | 3.5g | 72.3e | 0.042a | 14.32bcd | 0.048a | 2983.5c | 8.01f | ||
| Mean | 160.7 | 7.64 | 0.14 | 42.56 | 40.33 | 34.25 | 145.7 | 17.1 | 88.2 | 0.031 | 14.96 | 14.96 | 14.96 | 8.68 | ||
| 2013–2014 | D15 | R15 | 177.1bc | 11.17b | 0.113g | 45.54a | 42.90a | 36.44a | 212.2a | 26.0b | 105.9b | 0.033f | 17.88a | 0.039ef | 2422.5i | 10.98a |
| R25 | 182.6a | 11.52a | 0.131e | 44.92a | 42.80a | 36.24a | 180.9c | 27.2a | 111.2a | 0.027h | 17.31bc | 0.036f | 2533.5g | 9.98c | ||
| R35 | 180.3ab | 11.3ab | 0.123f | 45.23a | 42.90a | 36.37a | 196.1b | 26.5b | 107.3b | 0.031g | 17.45ab | 0.039ef | 2460.5h | 10.42b | ||
| D30 | R15 | 175.7cd | 8.84e | 0.160d | 45.39a | 42.35ab | 35.76ab | 141.1f | 18.8c | 93.7c | 0.035e | 16.12de | 0.040de | 2942.5d | 8.63e | |
| R25 | 174.6cd | 9.41d | 0.164cd | 43.43b | 41.51bc | 35.39bc | 148.2e | 18.0d | 91.8c | 0.038cd | 16.38d | 0.044bc | 2894.5e | 9.08d | ||
| R35 | 172.1de | 9.91c | 0.167bcd | 43.01b | 40.99c | 34.76c | 165.6d | 15.0e | 88.2d | 0.039c | 16.90c | 0.045b | 2790.0f | 9.84c | ||
| D45 | R15 | 171.5de | 7.34h | 0.171abc | 43.12b | 41.10c | 35.04bc | 124.7h | 13.5e | 83.2e | 0.037d | 15.26f | 0.042cd | 3343.5a | 7.44g | |
| R25 | 170.1e | 7.77g | 0.173ab | 41.28c | 39.36d | 33.38d | 130.9g | 7.2f | 82.2ef | 0.041b | 15.74e | 0.046b | 3185.0b | 8.08f | ||
| R35 | 169.9e | 8.29f | 0.175a | 39.70d | 37.91e | 32.45e | 137.8f | 4.8g | 80.4f | 0.044a | 16.05de | 0.050a | 3049.0c | 8.74e | ||
| Mean | 174.9 | 9.56 | 0.153 | 43.51 | 41.31 | 35.09 | 159.7 | 17.4h | 93.8 | 0.036 | 16.57 | 16.57 | 2846.8 | 9.24 | ||
| Analysis of variance | Year (Y) | 1058.00** | 449.20** | 273.27** | 25.22** | 50.91** | 42.15** | 102.20** | 11.40NS | 198.40** | 130.50** | 695.70** | 695.70** | 764.01** | 3162.02** | |
| Density (D) | 163.60** | 704.70** | 515.71** | 177.93** | 279.26** | 253.29** | 1386.40** | 10092.60** | 3069.30** | 1695.70** | 203.70** | 203.70** | 5080.48** | 798.69** | ||
| Row spacing (R) | 5.76** | 62.80** | 32.84** | 42.18** | 57.71** | 43.69** | 42.90** | 785.20** | 13.60** | 150.10** | 5.70** | 5.70** | 315.86** | 75.74** | ||
| Y × D | 6.10* | 14.20** | 2.14NS | 2.79NS | 5.58** | 14.52** | 16.90** | 65.50** | 16.40** | 19.00** | 0.40NS | 0.40NS | 2.76NS | 0.27NS | ||
| Y × R | 1.70NS | 2.50NS | 0.03NS | 2.61NS | 2.66NS | 2.98NS | 5.90** | 8.80** | 2.80NS | 6.50** | 0.70NS | 0.70NS | 2.75NS | 1.67NS | ||
| D × R | 6.30** | 15.00** | 12.86** | 9.53** | 13.06** | 11.41** | 50.20** | 443.50** | 7.70** | 176.80** | 5.40** | 5.40** | 160.47** | 66.08** | ||
| Y × D × R | 0.50NS | 4.10* | 0.82NS | 0.47NS | 0.63NS | 0.58NS | 5.40** | 13.60** | 0.80NS | 8.60** | 1.20NS | 1.20NS | 1.66NS | 0.92NS | ||
Values followed by different letters within the same column are significantly different according to the least significant difference (LSD) test (P < 0.05); Each data represents the mean of three replications. NS means not significant; *and **means significant differences at 0.05 and 0.01 probability levels, respectively; D15, D30 and D45 mean densities of 15, 30 and 45 × 104 plants ha−1; R15, R25 and R35 mean inter-row spacing of 15, 25 and 35 cm.
Harvest loss rate
Loss from cleaning and threshing (CTL) accounted for the largest proportion of yield loss, whereas the loss from natural pod shattering (PSL) accounted for the smallest proportion of yield loss. Plant density and row spacing had little impact on PSL and the loss from the combine header (CHL), whereas the loss rate of un-threshed pods in the straw (CTL1) and the loss rate of shattered seeds in the straw (CTL2) showed marked changes under different plant densities and row spacings. Increasing plant density significantly reduced the PSL, the loss rate of the seed shattered by mechanical harvesting (CHL1) and the loss rate of seeds on the branches remaining in each mechanical harvesting area (CHL2), whereas the CTL2 loss rate increased with increasing plant density. Under higher plant densities (D30 and D45), increasing row spacing reduced the PSL and CTL2 loss rates, whereas the CHL1, CHL2 and CTL1 loss rates increased (Fig. 3).
Figure 3. Loss rates of PSL (pod shatter loss pre-harvesting), CHL1 (shatter seed loss caused by combine header), CHL2 (branches loss caused by combine header), CTL1 (loss of unthreshed pods) and CTL2 (loss of shattered seeds mixed up with the straw) for direct-seedling winter rapeseed under different densities and row spacing arrangements during 2012–2014.

D15, D30 and D45 mean densities of 15, 30 and 45 × 104 plants ha−1; R15, R25 and R35 mean inter-row spacing of 15, 25 and 35 cm. Different letters denote significant difference at P < 0.05 by LSD.
Discussion
Plant densities of 15 × 104 − 45 × 104 plant ha−1 and row spacings of 15–35 cm have long been known to improve the seed yield of rapeseed in the Yangzi River region. However, rapeseed plant spacing requirements vary depending on the plant density, which has always been neglected by farmers in China. The recommended combination of appropriate density and row spacing for mechanical harvesting has not been reported. Consequently, research related to the combined effects of plant density and row spacing are urgently needed.
Yield differences were observed between the two growing seasons. The seed yield in 2013–2014 was higher than in 2012–2013. More sunshine and higher temperatures during the seedling and pod-filling stages, as well as during the entire growth period, in 2013–2014 compared with 2012–2013 were favorable for seedling and seed development (Table 1). In addition, more precipitation during the flowering to pod-filling stages in 2012–2013 resulted in more severe pod shattering24, which was unfavorable for production.
Different spatial arrangements produced by changes in row spacing can affect resource competition relationships at both the intra-specific and the inter-specific levels. Plants growing in rows that are too wide may not efficiently utilize natural resources such as light, water and nutrients, whereas growing in rows that are too narrow may result in severe inter- and intra-row spacing competition. Intra-specific competition has three effects: (i) density-dependent mortality, (ii) trade-offs between size and density, and (iii) alterations to population size structure25. The first two effects can be inferred by changes in average yield components, whereas the distribution of plant sizes within the crop provides information about the population structure. In the present study, plant mortality increased with reduced plant spacing as plant density and inter-row spacing increased (Table 2), indicating serious intra-specific competition under these conditions. The rapeseed plants adjusted to growing conditions and stand differences by changing the number of pods per plant, the number of seeds per pod and the 1000-seed weight (Table 3). There was a significant interaction between plant density and row spacing for yield and yield components, and plant density had a significantly greater influence than row spacing. Among the yield components, the number of pods per plant was most affected by plant density and row spacing, as reflected by the high coefficient of variation. Higher plant density and wider row spacing produced fewer pods per plant, resulting in a reduced yield per plant. The number of seeds per pod and the 1000-seed weight decreased with plant density, but increased with row spacing; this may have occurred because the rapeseed compensated for a loss in number of pods per plant with more seeds and higher seed weight, as has been previously reported13. Although wider rows produced more seeds and larger seeds, these were not sufficient to compensate for the loss in yield per plant caused by a decreased number of pods per plant when row spacing increased at the same plant density.
Under the conditions of this study, yield superiority was due primarily to the size of the plant population, where each plant was allowed a specific growing area, rather than the intra- or inter-row spacing individually. Compared with wider rows, narrower row spacing under higher plant density increased the distance between the plants within the row, resulting in a more equidistant planting pattern that is expected to delay the initiation of intra-specific competition as well as reduce intra-row competition26 while increasing early crop growth27. Leach et al. (1999) found that, in a series of multi-factorial experiments, the seed yield of winter oilseed rape increased with plant densities up to 50–60 × 104 plants ha−1 10. Besides, cultivar Huayouza 62 was planted by mechanical seeding with five planting densities (15 × 104, 30 × 104, 45 × 104, 60 × 104, and 75 × 104 plant ha–1) in two growing seasons (2011–2012 and 2012–2013). Results showed that the direct seed rapeseed could obtain the highest yield under both densities of 30 × 104 plants ha−1 and 45 × 104 plants ha−1. Density higher than 45 × 104 plants ha−1 results in strong competition, increases the potential for cooperation and also increases the difficulty for mechanical production28. As expected, the highest yield was observed under D45 in combination with R15, where the plant spacing and row spacing were similar, indicating that higher yields were associated with a more even plant distribution and a lower degree of intra-row competition. These findings are in agreement with Morrison et al. (1990)13, Argadi et al. (2003)29 and Karchi and Rudich (1966)30, who reported that uniformly distributed plant populations had significant advantages in yield over plant populations that were not uniformly distributed.
The combination of plant density and row spacing defines the spatial distribution of the plants, which affects canopy structure, light interception and radiation use efficiency and consequently, biomass production31. By narrowing the row width, a nearly optimum canopy display of leaves could be achieved resulting in greater seed yields than in wide rows32. In the present study, the population biomass showed a response to plant density and row spacing similar to yield. Variation in the population biomass could arise as a result of differences in radiation intercepted by the canopy, RUE and partitioning among different tissues33. Previous studies reported that during the period of most intensive accumulation of dry matter in the seeds, the proportion of assimilated carbon delivered by the photosynthetic tissues to the growing seeds was 37% for the leaves, 32% for the fruit walls, 31% for the stem and 1% for the seeds34, indicating that the leaves and the pods made the most important contributions to the yield of rapeseed. The potential increase in yield under D45R15 was closely related to the dense plant canopy, as seen in LAI and PAI (Fig. 1). The PAI was greater than the LAI, indicating that pods may be very important in providing photosynthates for their own development because they receive much higher radiation intensities than the leaves, although their photosynthetic rate was lower than that of the leaf35. Changes in the population yield under varying plant density and row spacing are affected not only by the rate of photosynthesis per unit of leaf area and the total area of photosynthetically active surfaces, but are also affected by the penetration of photosynthetically active light into the canopy because changes in row spacing and plant density could alter the timing of canopy closure by changing leaf area or pod area. Early canopy closure at higher plant density and narrower row spacing not only maximizes light interception between rows, but it evidently allowed more photosynthetically available radiation (PAR) to be transmitted between plants within rows. In addition, it also decreases evaporation from the soil surface and inhibits weed growth36. As expected, in this study, the percentage of intercepted radiation increased as plant density and row spacing increased, with the maximum interception being observed under D45R35 (Table 4). Significant positive correlations were observed between the LAI and percentage of intercepted light (R2 = 0.5168**), as well as between the PAI and percentage of intercepted light (R2 = 0.5037**) (Fig. 2). Allen and Morgan (1972) observed that the number of pods and the number of seeds per pod were positively correlated with the LAI at the onset of flowering37. This indicated that the difference in light interception related to LAI and PAI was one reason for yield changes under varying plant densities and row spacings. However, biomass production in rapeseed is more closely related to the utilization of solar radiation than to its interception because the RUE was significantly correlated with population biomass (Fig. 2). The influence of row spacing on the RUE was dependent on plant density. It had a response similar to the yield, with the maximum RUE being observed under narrow row spacing (15 cm) and under higher plant density (45 × 104 plants ha−1) (Table 4). Overall, our values of RUE are lower than those of previous researchers as reported by Fletcher et al. (2012)38. There may be a genotypic difference among rapeseeds39 or lower air temperatures in winter could limit leaf photosynthesis40. The harvest index (HI) is one of the indices currently used to evaluate a crop’s partitioning efficiency. HI has been shown to improve with earlier maturity and to be negatively correlated with height and lodging score23, which is consistent with our results. HI clearly increased with plant density and row spacing (plant height and lodging were all reduced under this condition, as seen in Table 5), indicating that more photosynthate was distributed to the economic portions of the plant. Taken together, these results indicated that sowing rapeseed in narrow rows at higher plant density created a dense canopy and increased the light interception, RUE and distribution of photosynthetic production to the harvestable portion of the plant, leading to a yield increase in our study.
Stem lodging and pod shattering represent the most serious constraints to high yield during the mechanical harvesting of rapeseed. Experiments with natural and artificially induced lodging have shown that lodging-induced yield losses can range between 0 and 80%41, and stems lodged at 45° resulted in less yield loss than stems lodged at 80°42. Shattering can result in a yield loss of up to 50% by decreasing the overall dry matter production as well as all of the major yield components43. The combined effects of plant density and row spacing were studied here to find an optimum plant density and row spacing arrangement for the mechanical harvesting of rapeseed.
Because the plants lodged from the base of the stem, a weak stem or a shallow root system was presumed to be the cause of lodging. Most investigators have concluded that plots seeded at narrow row spacing and at high seeding rates produced plants with thinner stems that were less able to support the weight of the pods and the seeds and were more sensitive to lodging44. High plant density and narrow row spacing created a dense canopy where growing plants receive a different quality of light, enriched with far red (FR) and impoverished in red (R) radiation. This high FR/R ratio triggers many morphological changes in plant architecture, stimulating stem elongation, favoring apical dominance and reducing the stem diameter45. Such changes might make stems more susceptible to breakage before seeds reach physiological maturity. However, other results have suggested that the l5 cm row spacing produced significantly greater yields and lodged less frequently than the 30 cm row spacing at seeding rates of 1.5–12.0 kg ha−1 13. We found that the lodging index and the angle of lodging both decreased at higher plant density and wider row spacing, with the minimum values being observed under D45R35 (Table 5). The modern hybrid morphological traits may have helped to mitigate stem lodging at high plant population densities. Plant height is important when considering harvest equipment and has been targeted in efforts to improve lodging resistance46. Lodging resistance was greatly influenced by plant density and row spacing and was higher in wide-row compared with narrow-row treatments in soybeans [Glycine max (L.) Merr.]35. In contrast, in our research, there was decreased plant height of rapeseed under higher plant density and wider row spacing, possibly because the field was rain-fed, without any irrigation during the entire development stage, and thus intra-specific competition existed for water, which limited stem elongation. At sites with adequate soil moisture, mature plant heights increased as seeding rates increased, whereas at moisture-deficient sites, plant height was not significantly affected by increased seeding rates47. In addition, closer spacing within the row resulted in partial shading at earlier growth stages, and partial shading can change the light quality in the canopy, triggering plants to develop longer stem internodes48. Therefore, another reason for decreased plant height in this study might be that higher plant density and appropriate row spacing lowered the number of upright branches, and the pod numbers may have reduced the selective absorption of red light by the upper canopy, improving light quality and decreasing stem elongation. Higher plant density reduced the root biomass, but an irregular trend in root biomass was affected by row spacing in both seasons. However, the root-shoot ratio clearly increased as plant density and row spacing increased (Table 5), suggesting relatively better root growth under these conditions, which enhanced water and nutrient uptake. The short plant height and increased root-shoot ratio were probably instrumental in maintaining the plant’s gravitational center close to the ground, a key feature in sustaining the canopy weight until harvesting. Similar results were reported by Sangoi et al.49. Altering the pattern of plant spacing by changing plant density and row spacing also had impact on pod-shatter resistance. The increased HI improved the dry matter accumulation of the pod wall under the highest plant density and widest row spacing, and the same trend was observed for pod-shatter resistance (Table 5). In a previous study, we found that there was a significant correlation between pod-shatter resistance and pod dry weight50. Consequently, the increased biomass led to a more compact pod wall, which improved the ability of the pod to resist shattering.
Changes in agronomic traits, especially in lodging, pod shattering and canopy architecture, could affect the mechanical harvesting yield. At low seeding rates, a large proportion of the total yield is produced on the branches, and as the seeding rate increases, the yield from the branches decreases13 indicating that branch growth is inhibited as plant density increased. Similar results were observed in the present study. The angle of the five lower branches significantly decreased with increasing row spacing under higher plant densities (D30 and D45) (Table 5). Rapeseed, with a greater capacity for altering branch display angles, was well-suited for efficient light interception and mechanical harvesting operations in narrow rows under higher density, which ultimately reduced mechanical harvesting yield loss. The highest mechanical seed yield was obtained for the combination of 15.0 cm row spacing and 45 × 104 plants ha−1 plant density without a serious yield loss resulting from lodging and pod shattering, with decreased branching angles (Table 5) and with a higher theoretical seed yield. Similar results were reported by Bilgili et al. (2003)44.
Previous studies have shown that seed loss of rapeseed decreased by 5%–10% under direct harvesting with a combine harvester43, which was slightly lower than our results. The yield loss in the present study was between 7.0% and 11.0% (Table 5). Total yield loss consisted of loss from natural pod shattering, loss from the combine header and loss from cleaning and threshing. An optimal level of moisture was present in the seeds during harvesting with the combine harvester24,51, decreasing seed loss and damage. It was shown that a seed moisture content of 15%–20% could limit seed loss to within 11% (Table 5). The rate of PSL and CHL loss was largely influenced by weather conditions and combine header operation, respectively, while the loss from CTL was primarily affected by agronomic practices. According to the loss rates in the current study, the minimum seed loss was associated with PSL and the maximum loss was observed in CTL, which was caused by cleaning and threshing. This indicates that optimum plant density combined with appropriate row spacing helped to maintain the uniformity of the canopy and could significantly reduce yield loss caused by mechanical harvesting. Among the different types of loss rates, CTL1 was most significantly affected by plant density and row spacing (Fig. 3). Under the combination of plant density and row spacing, the minimum yield loss under D45R15 was primarily due to the low CTL1 loss rate. Plants at low population densities produce more branches that carry fertile pods, thus prolonging the seed development phase. This results in a range of seed maturities at harvest, which may affect seed quality, increase the risk of seed loss through pod shatter and impede harvesting. Decreased yield loss was observed with increasing plant density and narrowing row spacing because of an improvement in resistance to stem lodging and pod shattering, decreased branching angles and the presence of fewer pod-bearing branches that produced more synchronous pods and seed development and resulted in more uniform seed maturation, which facilitated harvesting10.
Conclusion
Uniform row spacing and plant spacing (a combination of 15.0 cm row spacing and 45 × 104 plants ha−1 plant density) could improve the seed yield and mechanical harvesting efficiency for direct-seeded rapeseed. This combination produced a more compact canopy architecture, leading to a higher LAI and PAI, enhanced solar radiation and RUE, and subsequently, higher crop biomass. All of these factors contributed to a high yield. In addition, this combination also led to shorter plants, increased root-shoot ratios, reduced branching angles and promoted greater resistance to stem lodging and pod shattering. These various agronomic traits facilitated mechanical harvesting. Overall, these results suggest that producers should be encouraged to seed rapeseed in narrowly-spaced rows with high plant density. The combination of 15.0 cm row spacing and 45 × 104 plants ha−1 plant density could be considered an optimum cultivation practice for mechanized production of direct-seeded rapeseed in Central China.
Methods
Experimental site
A replicated field experiment was conducted during two seasons (2012–2013 and 2013–2014) at the Huazhong Agricultural University Experimental Farm (30°28′12″N, 114°21′05″E) in Wuhan, China. The previous crop, rice, was harvested in September. The initial soil status (0–20 cm) of the field in terms of available nitrogen (N), phosphorus (P) and potassium (K) concentrations was as follows: 101.26 mg kg–1, 13.84 mg kg–1 and 146.28 mg kg–1, respectively, in 2012–2013 and 103.63 mg kg–1, 14.47 mg kg–1 and 150.38 mg kg–1, respectively, in 2013–2014. Table 1 shows the rainfall and temperature during the two growing seasons; these data were provided by the National Meteorological Information Center of the China Meteorological Administration.
Experimental design
Huayouza 62, a popular winter rapeseed hybrid cultivar in Central China, was used in this study. The experiment had a split-plot design with three plant densities (D1, 15 × 104 plant ha−1; D2, 30 × 104 plant ha−1; D3, 45 × 104 plant ha−1) as the primary plots and three row spacings (R1, 15 cm; R2, 25 cm; R3, 35 cm) as the split plots. The row arrangements are illustrated in Fig. 4; inter-row spacings and plant densities varied in each plot (Table 2). The plant densities of 15 × 104–45 × 104 plant ha−1 and row spacings from 15–35 cm used in the study are widely used for rapeseed production in the Yangzi River region. Each treatment was performed in three replicate plots.
Figure 4. The schematic diagram for direct-seedling winter rapeseed under different densities and row spacing arrangements during 2012–2014 (unit: cm).

D15, D30 and D45 mean densities of 15, 30 and 45 × 104 plants ha−1; R15, R25 and R35 mean inter-row spacing of 15, 25 and 35 cm.
Rapeseed was manually sown on the 23rd of September and the 26th of September in 2012 and 2013, respectively. The seed density was evaluated directly after seedling emergence and adjusted for precise planting density at the five-leaf growth stage for all plots. Before sowing, 900 kg ha−1 of N, P and K compound fertilizer (N:P:K = 15:15:15) and 7.5 kg ha−1 of borax (for boro) were applied as a basal fertilizer. During the wintering stage, 135 kg ha−1 nitrogen from urea was applied. Pest, disease and weed control were performed according to local management practices.
LAI, PAI and RUE
The Leaf area was measured on 20 plants in each plot at the bolting stage using a leaf area-meter (Li-3100c, Li-COR Inc., USA). The leaf area index (LAI) was defined as the ratio of total one-sided leaf area to ground surface area52. The pod area was measured on 20 plants in each plot 30 days after the end of flowering according to the formula: 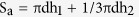 (where h1 = 0.8 H, h2 = 0.2 H, H is pod length and d is pod width)53. The pod area index (PAI) was defined as the ratio of total pod area to ground surface area. Canopy radiation interception was measured using a SunScan Canopy Analysis System (Delta-T Devices Ltd., UK) at the seedling, wintering, bolting, flowering, pod-filling and maturity stages as suggested by Wang et al. (2015), with some modification54. Canopy radiation interception was calculated as [100 × (incoming radiation intensity– radiation intensity inside canopy)/incoming radiation intensity]. Intercepted radiation was calculated as [1/2 × (canopy radiation interception at the beginning of the growth period + canopy light interception at the end of the growth period) × accumulated incoming radiation during the growth period]55. The radiation use efficiency (RUE) was calculated as the ratio of above-ground total dry weight at maturity to intercepted radiation during the entire growing season52.
(where h1 = 0.8 H, h2 = 0.2 H, H is pod length and d is pod width)53. The pod area index (PAI) was defined as the ratio of total pod area to ground surface area. Canopy radiation interception was measured using a SunScan Canopy Analysis System (Delta-T Devices Ltd., UK) at the seedling, wintering, bolting, flowering, pod-filling and maturity stages as suggested by Wang et al. (2015), with some modification54. Canopy radiation interception was calculated as [100 × (incoming radiation intensity– radiation intensity inside canopy)/incoming radiation intensity]. Intercepted radiation was calculated as [1/2 × (canopy radiation interception at the beginning of the growth period + canopy light interception at the end of the growth period) × accumulated incoming radiation during the growth period]55. The radiation use efficiency (RUE) was calculated as the ratio of above-ground total dry weight at maturity to intercepted radiation during the entire growing season52.
Measurement of lodging behavior
The lodging degree, snapping resistance, culm lodging index and pod-shatter resistance were measured as previously reported by Kuai et al. (2015)50.
 |
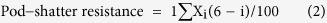 |
where i is the frequency of agitation and 1 ≤ i ≤ 5 and Xi is the number of broken pods.
After the random impact tests, the dry weight of the pod was recorded.
Assessment of manually harvesting seed yield and yield components
Plots were harvested when approximately 2/3 of the seed was brown. Ten plants from each plot were randomly sampled and slowly uprooted and the taproot and large lateral roots were retained. Next, the yield components and seed yield per plant were determined. On each plant, the following measurements and observations were made: the plant height (cm), the angle of the lowest 5 branches, pods per plant, seeds per pod and the 1000-seed-weight (g). Then, the remaining plants of 8 m in each plot were manually harvested to measure the manually harvesting seed yield (Ymn, kg ha−1). The heights of the plants were measured from the cotyledonary node to the ligule of the uppermost fully expanded leaf, and the average height was calculated from all plants measured. Plant tissue samples were separated from the cotyledonary node into roots and aboveground tissues. After the fresh weight was determined, the roots and the aboveground tissues were dried in an oven for 30 min at 105 °C to deactivate enzymes and then dried at 70 °C until a constant weight was reached for dry weight determination.
Assessment of yield loss and mechanical harvesting yield
A combine harvester (4LL-2.0D, Huzhou, China) was used for mechanical harvesting when the seed moisture of pods from the main inflorescences was 12%–13%. Each plot was harvested over a length of 40 m, maintaining a 0.4 m cutting height. Three sources of losses were assessed as shown in Fig. 5.
Figure 5. Schematic operation for determination yield losses in the study.

(A) loss of naturally pod shattering pre-harvesting; (B) loss of combine header; (C) loss of cleaning and threshing.
A. Loss from natural pod shattering (PSL): The pre-harvest losses due to natural dehiscence and environmental conditions, e.g., rainfall, wind, or birds. Because the direct collection of seeds that have fallen to the soil surface is impracticable, 10 plastic trays (25 cm × 15 cm × 5 cm) were positioned inside each plot, as described below, when approximately 2/3 of the seeds in the field were brown (the manual harvesting date)43,56. These trays were removed just prior to harvesting and the seed was weighed (WPSL). The pre-harvesting loss rate (PSL) (%) = WPSL/Ymn × 100.
B. Loss from combine header (CHL): The shattered seed loss and branch loss attributable to the combine header. The measurement of shattered seed loss during mechanical harvesting was conducted by placing 10 trays on the ground in each plot. The trays were placed 8 m before the end of the plot (the plot was 40 m in length) to allow for the combine to pass over them while operating (threshing, separating and cleaning). The distance before the trays (32 m) was also sufficient to allow the combine to transit above the trays in full operation. The weight of the seed shattered by mechanical harvesting was recorded. The loss rate of these seeds was represented by (CHL1) (%) = WCHL1/Ymn × 100. The branches remaining in each mechanical harvesting area (40 m in length) after harvest were collected, and the seeds on these branches were weighed. The loss rate of seeds on these branches was represented by (CHL2) (%) = W CHL2/Ymn × 100.
C. Loss from cleaning and threshing (CTL). The rear of the harvester was covered with a nylon membrane 3 m in width; the straw was collected for 20 m along each plot. The seeds of un-threshed pods and shattered seeds in the straw were collected and weighed separately. The loss rate of un-threshed pods in the straw was represented by (CTL1) (%) = WCTL1/Ymn × 100. The loss rate of shattered seeds in the straw was represented by (CTL2) (%) = WCTL2/Ymn × 100. All seeds freely falling from the frame of the harvester were collected and weighed to determine the mechanical harvesting yield.
The total loss rate (%) was determined as (Ymn −Ym)/Ymn × 100, where Ym = mechanical harvesting seed yield and Ymn = manually harvesting seed yield.
Statistics analysis
Two-way analysis of variance (ANOVA) was performed using Duncan’s multiple range test. Significant differences in means between the treatments were compared by the protected least significant difference (LSD) procedure at P < 0.05. ANOVA and the LSD test were conducted using the SPSS 17.0 software program. Figures were prepared using the Origin 9.0 software program.
Additional Information
How to cite this article: Kuai, J. et al. The yield of mechanically harvested rapeseed (Brassica napus L.) can be increased by optimum plant density and row spacing. Sci. Rep. 5, 18835; doi: 10.1038/srep18835 (2015).
Acknowledgments
This research was funded by the National Key Technology Research and Development Program (2013BAD20B06; 2014BAD11B03), the Industry Technology Research Institute (CARS-13), the Special Funds for Public Welfare Industry (Agriculture) Study of China (201203096) and the Fundamental Research Funds for the Central Universities (2013PY001).
Footnotes
Author Contributions All authors initiated and designed the experiments, K.J., S.Y. and H.H. conducted the experiments, K.J., L.Q. and Z.Q. analyzed the data and wrote the manuscript, W.C., L.J., W.J. and Z.G. revised and edited the manuscript and also provided advice on the experiments.
References
- Yin Y. & Wang H. Z. Achievement, problem and scientific policy of rapeseed industry development in China. J. Agr. Sci. Tech. 14, 1–7 (2012). [Google Scholar]
- Shahid L., Saeed M. & Amjad N. Present status and future prospects of mechanized production of oilseed crops in Pakistan- a review. Pakistan J. Agric. Res. 23, 83–93 (2010). [Google Scholar]
- Yin Y. &Wang H. Z. Production status and development trends of rapeseed in China. Agr. Outlook. 43–45 (2011). [Google Scholar]
- Ozer H. The effect of plant population densities on growth, yield and yield components of two spring rapeseed cultivars. Plant Soil Environ. 49, 422–426 (2003). [Google Scholar]
- Diepenbrock W. Yield analysis of winter oilseed rape (Brassica napus L.): a review. Field Crops Res. 67, 35–49 (2000). [Google Scholar]
- Rathke G. W., Behrens T. & Diepenbrock W. Integrated nitrogen management strategies to improve seed yield, oil content and nitrogen efficiency of winter oilseed rape (Brassica napus L.): A review. Agr. Ecosyst. Environ. 117, 80–108 (2006). [Google Scholar]
- Momoh E. J. J. & Zhou W. Growth and Yield Responses to plant density and stage of Transplanting in winter oilseed rape (Brassica napus L.). J Agron. Crop Sci. 186, 253–259 (2001). [Google Scholar]
- Yang G. et al. Cotton stubble mulching helps in the yield improvement of subsequent winter canola (Brassica napus L.) crop. Ind. Crop Prod. 50, 190–196 (2013). [Google Scholar]
- Zhang S. et al. Identification of the para- nitrophenol catabolic pathway, and characterization of three enzymes involved in the hydroquinone pathway, in pseudomonas sp. 1-7. BMC Microbiol. 12, 27–38 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leach J. E., Stevenson H. J., Rainbow A. J. & Mullen L. A. Effects of high plant populations on the growth and yield of winter oilseed rape (Brassica napus L.). J. Agric. Sci. Camb. 132, 173–180 (1999). [Google Scholar]
- Christensen J. V. & Drabble J. C. Effect of row spacing and seeding rate on rapeseed yield in northwest Alberta. Can. J. Plant Sci. 64, 1011–1013 (1984). [Google Scholar]
- Turgut I. et al. Alternate row spacing and plant density effects on forage and dry matter yield of corn hybrids (Zea mays L.). J Agron. Crop Sci. 191, 146–151 (2005). [Google Scholar]
- Morrison M. J., McVetty P. B. E. & Scarth R. Effect of row spaicng and seeding rates on summer rape in Southern Manitoba. Can. J. Plant Sci. 70, 127–137 (1990). [Google Scholar]
- Sincik M., Goksoy A. T. & Turan Z. M. Influence of sowing properties on winter oilseed rape in a sub-humid Mediterranean environment. Not. Bot. Hort. Agrobot. Cluj. 38, 171–175 (2010). [Google Scholar]
- Boquet D. J. Cotton in ultra-narrow row spacing: plant density and nitrogen fertilizer rates. Agron. J. 97, 279–287 (2005). [Google Scholar]
- Boquet D. J. Plant population density and row spacing effects on soybean at post-optimal planting dates. Agron. J. 82, 59–64 (1990). [Google Scholar]
- De Bruin J. L. & Pedersen P. Effect of row spacing and seeding rate on soybean yield. Agron. J. 100, 704–710 (2008). [Google Scholar]
- Henderson T. L., Johnson B. L. & Schneiter A. A. Row spacing, plant population, and cultivar effects on grain amaranth in the Northern Great Plains. Semigroup Forum. 92, 329–336 (2000). [Google Scholar]
- Shahin Y. &Valiollah R. Effects of row spacing and seeding rates on some agronomical traits of spring canola (Brassica napus L.) cultivars. J. Cent. Eur. Agric. 10, 115–122 (2009). [Google Scholar]
- Robinson R. G. Amaranth, quinoa, ragi, tef, and niger: Tiny varying plant density on the growth and yield of grain amaranth seeds of ancient history and modern interest, 1-23 (Stn. Bull. AD-SB-2949. Agric. Exp. Stn., Univ. of Minnesota, St. Paul, 1986).
- Haas P. W. Amaranth density report (Rodale Research Center Report No. RRC/NC-83-8, Rodale Press, Emmaus, Pennsylvania, USA, 1983).
- Haas P. W. & Kauffman C. S. Grain amaranth: An overview of research and production methods (Rodale Research Center Report No. NC-84/6, Rodale Press, Emmaus, Pennsylvania, USA, 1984).
- Stapper M. & Fischer R. Genotype, sowing date and plant spacing influence on high-yielding irrigated wheat in southern New South Wales. II. Growth, yield and nitrogen use. Aust. J. Agr. Res. 41, 1021–1041 (1990). [Google Scholar]
- Ma N. et al. Mechanical harvesting effects on seed yield loss, quality traits and profitability of winter oilseed rape (Brassica Napus L.). J. Integr. Agr. 11, 1297–1304 (2012). [Google Scholar]
- Park S. E., Benjamin L. R. & Watkinson A. R. The theory and application of plant competition models: an agronomic perspective. Ann. Bot. 92, 741–748 (2003). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duncan W. A theory to explain the relationship between corn population and grain yield. Crop Sci. 24, 1141–1145 (1984). [Google Scholar]
- Bullock D., Nielsen R. & Nyquist W. A growth analysis comparison of corn grown in conventional and equidistant plant spacing. Crop Sci. 28, 254–258 (1988). [Google Scholar]
- Zuo Q. et al. Effects of nitrogen fertilizer and planting density on canopy structure and population characteristic of rapeseed with direct seeding treatment. Acta Agronomica Sinica, 41, 758–765 (2015). [Google Scholar]
- Angadi S. V. et al. Yield adjustment by canola grown at different plant populations under semiarid conditions. Crop Sci. 43, 1358–1366 (2003). [Google Scholar]
- Karchi Z. & Rudich Y. Effects of row width and seedling spacing on yield and its components in grain sorghum grown under dryland conditions. Agron. J. 58, 602–604 (1966). [Google Scholar]
- Mattera J. et al. Yield components, light interception and radiation use efficiency of lucerne (Medicago sativa L.) in response to row spacing. Eur. J. Agron. 45, 87–95 (2013). [Google Scholar]
- Herbert S. J. & Litchfield G. V. Growth response of short-season soybean to variations in row spacing and density. Field Crops Res. 9, 163–171 (1984). [Google Scholar]
- Hamzei J. & Soltani J. Deficit irrigation of rapeseed for water-saving: Effects on biomass accumulation, light interception and radiation use efficiency under different N rates. Agr. Ecosyst. Environ. 155, 153–160 (2012). [Google Scholar]
- Brar G. & Thies W. Contribution of leaves, stem, siliques and seeds to dry matter accumulation in ripening seeds of rapeseed, Brassica napus L. Zeitschrift für Pflanzenphysiologie. 82, 1–13 (1977). [Google Scholar]
- Gammelvind L. et al. Photosynthesis in leaves and siliques of winter oilseed rape (Brassica napus L.). Plant soil. 186, 227–236 (1996). [Google Scholar]
- Seiter S.,. Altemose C. E. & Davis M. H. Forage soybean yield and quality responses to plant density and row distance. Agron. J. 96, 966–970 (2004). [Google Scholar]
- Allen E. J. & Morgan D. G. A quantitative analysis of the effects of nitrogen on the growth, development and yield of oilseed rape. J. Agr. Sci. 78, 315–324 (1972). [Google Scholar]
- Fletcher A. L. et al. Canopy development and radiation use efficiency of four forage brassica crops. In:16th Australian Agronomy Conference, Armidale, Australia (2012).
- Fletcher A. L. et al. Radiation capture and radiation use efficiency in response to N supply for crop species with contrasting canopies. Field Crops Res. 150, 126–134 (2013). [Google Scholar]
- Sinclair T. R. & Muchow R. C. Radiation use efficiency. Advances in agronomy. 65, 215–265 (1999). [Google Scholar]
- Berry P. M. & Spink J. Predicting yield losses caused by lodging in wheat. Field Crops Res. 137, 19–26 (2012). [Google Scholar]
- Fischer R. & Stapper M. Lodging effects on high-yielding crops of irrigated semidwarf wheat. Field Crops Res. 17, 245–258 (1987). [Google Scholar]
- Price J. et al. Seed losses in commercial harvesting of oilseed rape. J. Agr. Eng. Res. 65, 183–191 (1996). [Google Scholar]
- Bilgili U. et al. The influence of row spacing and seeding rate on seed yield and yield components of Forage Turnip (Brassica rapa L.). J. Agron. Crop Sci. 189, 250–254 (2003). [Google Scholar]
- Rajcan I. & Swanton C. J. Understanding maize–weed competition: resource competition, light quality and the whole plant. Field Crops Res. 71, 139–150 (2001). [Google Scholar]
- Peng D. et al. Lodging resistance of winter wheat (Triticum aestivum L.): Lignin accumulation and its related enzymes activities due to the application of paclobutrazol or gibberellin acid. Field Crops Res. 157, 1–7 (2014). [Google Scholar]
- Devlin D. L. et al. Row spacing and seeding rates for soybean in low and high yielding environments. J. Prod. Agric. 8, 215–222 (1995). [Google Scholar]
- Kasperbauer M. J. Spectral distribution of light in a tobacco canopy and effects of end-of-day light quality on growth and development. Plant Physiol. 47, 775–778 (1971). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sangoi L. et al. Response of Brazilian maize hybrids from different eras to changes in plant density. Field Crops Res. 79, 39–51 (2002). [Google Scholar]
- Kuai J. et al. Paclobutrazol increases canola seed yield by enhancing lodging and pod shatter resistance in Brassica napus L. Field Crops Res. 180, 10–20 (2015). [Google Scholar]
- Alizadeh M. R. & Payman B. M. H. Evaluation of a rice reaper used for rapeseed harvesting. Agricultural and biosystems engineering for a sustainable world. International Conference on Agricultural Engineering, Hersonissos, Crete, Greece, 23–25 June, 2008, 4, 388–394.
- Behrens T. & Diepenbrock W. Using hemispherical radiation measurements to predict weight‐related growth traits in oilseed rape (Brassica napus L.) and barley (Hordeum vulgare L.) Canopies. J. Agron. Crop Sci. 192, 465–474 (2006). [Google Scholar]
- Zhou W. & Xi H. Effects of mixtalol and paclobutrazol on photosynthesis and yield of rape (Brassica napus L.). J. Plant Rrowth Regul. 12, 157–161 (1993). [Google Scholar]
- Wang R., Cheng T. & Hu L. Effect of wide–narrow row arrangement and plant density on yield and radiation use efficiency of mechanized direct-seeded canola in Central China. Field Crops Res. 172, 42–52 (2015). [Google Scholar]
- Zhang Y. et al. Yield potential and radiation use efficiency of “super” hybrid rice grown under subtropical conditions. Field Crops Res. 114, 91–98 (2009). [Google Scholar]
- Pari L. et al. The harvest of oilseed rape (Brassica napus L.): The effective yield losses at on-farm scale in the Italian area. Biomass Bioenerg. 46, 453–458 (2012). [Google Scholar]