

Abstract
AIM: To study the effect of phosphorylation of MAPK and Stat3 and the expression of c-fos and c-jun proteins on hepatocellular carcinogenesis and their clinical significance.
METHODS: SP immunohistochemistry was used to detect the expression of p42/44MAPK, p-Stat3, ca2fos and c-jun proteins in 55 hepatocellular carcinomas (HCC) and their surrounding liver tissues.
RESULTS: The positive rates and expression levels of p42/44MAPK, p-Stat3, c-fos and c-jun proteins in HCCs were significantly higher than those in pericarcinomatous liver tissues (PCLT). A positive correlation was observed between the expression of p42/44MAPK and c-fos proteins, and between pa2Stat3 and c-jun, but there was no significant correlation between p42/44MAPK and p-Stat3 in HCCs and their surrounding liver tissues.
CONCLUSION: The abnormalities of Ras/Raf/MAPK and JAKs/Stat3 cascade reaction may contribute to malignant transformation of hepatocytes.Hepatocytes which are positive for p42/44MAPK, c-fos or c-jun proteins may be potential malignant pre-cancerous cells. Activation of MAPK and Stat3 proteins may be an early event in hepatocellular carcinogenesis.
Keywords: liver neoplasms, mitogen-activated protein kinases, signal transduction, trans-activators, oncogenes, immunohistochemistry, precancerous conditions
INTRODUCTION
The regulation of cellular mitogenesis and proliferation under normal circumstances are dependent on a number of separate, yet integrated, signaling pathways responsible for the transduction of extracellular signals to the cell nucleus[1-3]. Similar to other tumorigenic cells, hepatocellular carcinoma (HCC) is characterized by the imbalance of the normal growth-promoting and growth-arresting signal transduction cascade reaction, the net result of which leads to uncontrolled hepatocyte growth [1,2]. Mitogen-activated protein kinase (MAPK) and signal transducer and activator of transcription 3 (Stat3) are important molecules in signal transduction cascades, which take part in cellular physiological growth, development, mitogenesis and differentiation, and play key roles in cellular malignant transformation[4,5]. In order to study the function of signal transduction cascade in hepatocarcinogenesis, the phosphorylation of MAPK (p42/44 MAPK, Erk1/Erk2) and Stat3 (p-Stat3), and the expression of c-fos and c-jun proteins were investigated by immunohistochemical method in HCCs and their surrounding liver tissues.
MATERIALS AND METHODS
Tissue samples
Fifty-five HCCs with pericarcinomatous tissue (PCLT) and 5 normal liver tissues were obtained by surgical resection in Xiang Ya Hospital and the Affiliated Second Hospital of Hunan Medical University, Changsha, People’s Republic of China. Of 55 HCC patients, 43 were men and 12 women. The age ranged from 36 to 69 years (mean, 51.8 years). The tumor sizes were less than 3cm in diameters in 3 cases, 2.1 cm-5 cm in 32 cases, 5.1 cm-10 cm in 18, and larger than 10 cm in 2 cases. All patients were HBsAg negative. Remote metastasis was not found, and preoperative radiological or chemical therapy was nor performed in all patients. All HCCs and PCLTs were fixed in 10% formalin and embedded in paraffin. Five μm serial sections were cut. According to Edmondson’s criteria[6], 10 HCCs were grade I, 21 grade II, 17 grade III, and 7 grade IV. Thirty-eight atypical hyperplasia cases were observed in 55 PCLTs.
Reagents
p42/44 MAPK (Erk1/Erk2, Thr202/Tyr204) and p-Stat3 (Ser727) MAbs, were used to detect the phosphorylation of MAPK and Stat3, and S-P detection kit were kindly donated by New England Biolab, USA. Anti-c-fos and anti-c-jun protein MAbs were purchased from GIB Com (Beijing, China).
Methods
Immunohistochemical staining Five μm tissue sections were deparaffinized, washed in 0.05 M PBS, and treated with 2% H2O2 for 20 min. According to the SP method, the tissues were detected with immunohistochemical technique. PBS was used as substitutes of MAbs for negative control groups[7].
Histological assessment Semi-quantity analysis was made as Formowitz described[8].
Statistical analysis
The difference between each group was analyzed by Chi-Square test and correlativity.
RESULTS
Expression of p42/44MAPK and p-Stat3 proteins in HCCs, PCLTs and normal liver tissues
p42/44MAPK was negative in normal liver tissues. The positive rate of p42/44MAPK in HCCs (87.3%, 48/55) was significantly higher than that in PCLTs (32.7%, 18/55, P < 0.01) (Figure 1). Positive signals were mainly located in the nucleus. The distribution of positive cells for p42/44MAPK was clustered and/or diffused in HCCs (Figure 2), and scattered in PCLTs, but also clustered in atypical hyperplasia regions of some PCLTs near cancer tissues. The expression level of p42/44MAPK in HCCs was higher than that in PCLTs (P < 0.01), but it was not correlated with the differentiation degree of cancer cells (Table 1). Positive signals of p-Stat3 were of nuclear-type. Of 9 normal liver tissues, 1 case was p-Stat3 positive. The positive rate of p-Stat3 in HCC (74.5%, 41/55) was significantly higher than that in PCLTs (23.6%, 13/55) (P < 0.01). The p-Stat3 positive cells were clustered in HCC (Figure 3), and scattered in PCLTs. The expression intensity of p-Stat3 in HCCs was also higher than that in PCLTs (P < 0.01), but it was not correlated with differentiation degree of cancer cells (P > 0.05) (Table 1).
Figure 1.
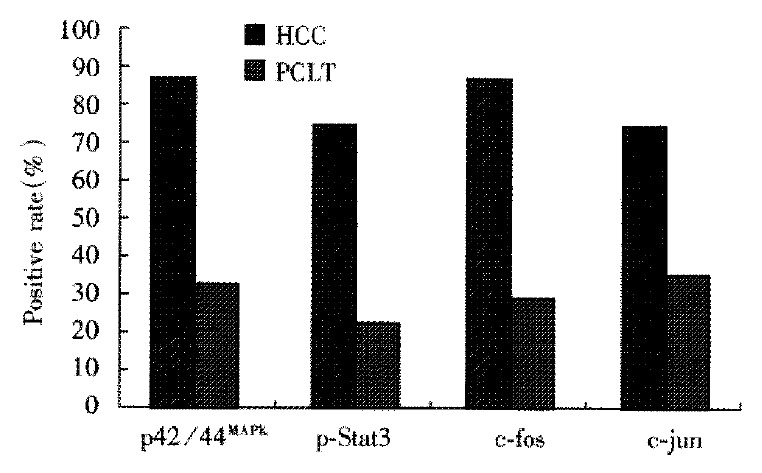
The positive rates of p42/44MAPK, p-Stat3, c-fos and c-jun proteins in HCCs and PCLTs.
Figure 2.
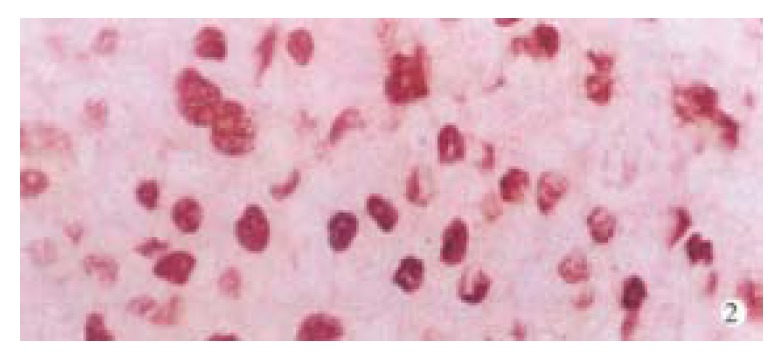
Localization of positive signal and distribution of positive cells for p42/44MAPK protein in HCC. SP × 400
Table 1.
Expressions of p42/44MAPK, p-Stat3, c-fos and c-jun proteins in HCCs, PCLTs and normal liver tissues
| Groups | No |
p42/44MAPK |
p-Stat3 |
c-fos |
c-jun |
||||||||||||
| - | + | ++ | +++ | - | + | ++ | +++ | - | + | ++ | +++ | - | + | ++ | +++ | ||
| Normal | 9 | 9 | 0 | 0 | 0 | 8 | 1 | 0 | 0 | 9 | 0 | 0 | 0 | 9 | 0 | 0 | 0 |
| PCLT | 55 | 37 | 9 | 7 | 2 | 42 | 4 | 7 | 2 | 36 | 12 | 7 | 0 | 39 | 13 | 3 | 0 |
| HCC | |||||||||||||||||
| Grade I | 10 | 1 | 4 | 4 | 1 | 5 | 1 | 2 | 2 | 8 | 1 | 1 | 0 | 4 | 4 | 1 | 1 |
| Grade II | 21 | 3 | 6 | 12 | 0 | 2 | 12 | 5 | 2 | 3 | 12 | 4 | 2 | 1 | 15 | 4 | 1 |
| Grade III | 17 | 1 | 9 | 2 | 5 | 4 | 3 | 6 | 4 | 2 | 4 | 8 | 3 | 2 | 8 | 5 | 2 |
| Grade IV | 7 | 2 | 1 | 3 | 1 | 3 | 1 | 3 | 0 | 0 | 1 | 2 | 4 | 0 | 0 | 2 | 5 |
Figure 3.
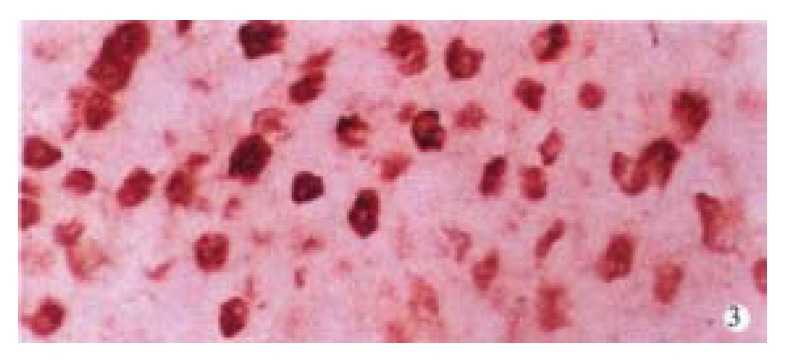
Localization of positive signal and distribution of positive cells for Stat3 protein in HCC. SP × 400
Expression of c-fos and c-jun proteins in HCCs, PCLTs and normal liver tissues
The expression of c-fos and c-jun proteins was not found in normal liver tissues. The rates of c-fos and c-jun protein expression in HCCs (76.4%, 87.3%) were significantly higher than those in PCLTs (34.5%, 29.1%) (P < 0.01). Their positive signals were mainly of nuclear type in HCCs (Figure 4, Figure 5) and nuclear-plasmic type in PCLTs. The positive intensity in HCCs was distinctly higher than that in PCLTs, and related to the differentiation degree of HCC cells. The poorer the differentiation of HCC cells, the stronger the expression of c-fos and c-jun proteins. The distribution of positive cells for c-fos or c-jun protein in HCCs was diffuse, and scattered in PCLTs.
Figure 4.
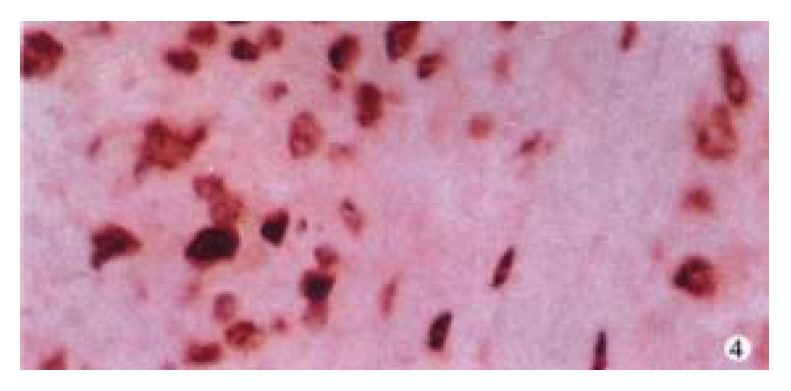
Positive expression of c-fos protein in HCC. SP × 400
Figure 5.
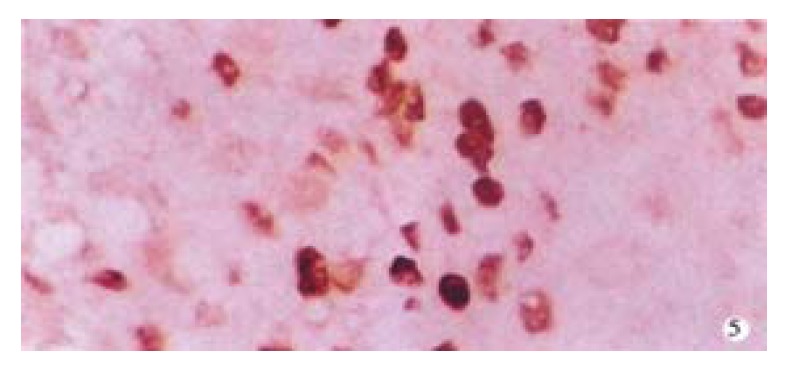
Positive expression of c-jun protein in HCC. SP × 400
Relativity of expression intensities of p42/44MAPK and p-Stat3 in HCCs and PCLTs
No relationship was found between p42/44MAPK and p-Stat3 signal intensities in HCCs and PCLTs (r = -0.167 and 0.0844, P > 0.05).
Relationship between expression strength of p42/44MAPK and c-fos proteins, between p42/44MAPK and c-jun in HCCs and PCLTs (Table 2)
Table 2.
Relationship in expressive strengthes of p42/44MAPK, c-fos and c-jun proteins between HCCs and PCLTs
| Groups | r | P |
| HCC | ||
| p42/44MAPK/c-fos | 0.44835 | < 0.01 |
| p42/44MAPK/c-jun | 0.3054 | < 0.01 |
| PCLT | ||
| p-MAPK/c-fos | 0.9433 | < 0.01 |
| p-MAPK/c-jun | 0.1472 | > 0.05 |
In HCCs, the expression of p42/44MAPK was positively related to the expression intensities of c-fos and c-jun proteins. The stronger the expression of p42/44MAPK, the higher the signal intensities of c-fos and c-jun proteins. In PCLTs, there was a positive relationship between the expression of p42/44MAPK and c-fos proteins, and no relationship between the expression of p42/44MAPK and c-jun proteins.
Relationship between expression strength of p-Stat3 and c-fos proteins, between p-Stat3 and c-jun in HCCs and PCLTs (Table 3)
Table 3.
Relationship in expression strength of p-Stat3, c-fos and c-jun proteins between HCCs and PCLTs
| Groups | r | P |
| HCC | ||
| p-Stat3/c-fos | 0.3967 | < 0.01 |
| p-Stat3/c-jun | 0.5444 | < 0.01 |
| PCLT | ||
| p-Stat3/c-fos | 0.0499 | > 0.05 |
| p-MAPK/c-jun | 0.3596 | < 0.01 |
In HCCs, the expression of p42/44MAPK was significantly positively related to expression intensities of c-fos and c-jun proteins. In PCLTs, there was a positive relationship between the expression of p-Stat3 and c-jun proteins, whereas no relationship between the expression of p-Stat3 and c-fos proteins was observed.
DISCUSSION
Ras/Raf/MAPK and Janus protein tyrosine kinases (JAKs)/Stat3 cascades are two major signaling pathways of growth factors and cytokines[1,2,4,5,9,10]. The former involves transient formation of ras-GTP and activation of raf kinase at the cell membrane to activate MAPK kinase which in turn activates MAPK through dual phosphorylation on threonine and tyrosine residues[11-14]. Activated MAPK then transduces into the nucleus and phosphorylates the ternary complex factor TCF which activates the expression of immediate-early genes, such as c-fos and egr-1[14,15]. There have been a few reports concerning MAPK activation in human cancers[1,2,16-21]. It was reported that MAPK activity level in HCC was significantly higher than that in the adjacent non-cancerous lesions[1,2,18]. Our results showed that the positive signal of p42/44MAPK was almost located in the nucleus, indicating that the detected MAPK is activated MAPK. The positive rate and intensity of p42/44MAPK in HCCs were all distinctly higher than those in PCLTs, which is consistent with the results reported in literature. Statistical analysis found that the expression intensity of p42/44MAPK in HCCs was significantly positively correlated with the expression strength of immediate-early genes (c-fos and c-jun), and the expressive intensity of p42/44MAPK in PCLTs was also significantly positively correlated with the expression strength of c-fos. These findings further suggest that the imbalance of Ras/Raf/MAPK cascade components may play a key role in hepatocarcinogenesis.
JAKs associate with cytokine receptors which lack intrinsic tyrosine kinase activity and become activated upon cytokine stimulation. Activated JAKs activate Stat3 through phosphorylation of Tyr-705[22]. Phosphorylated Stat3 (p-Stat3) then translocates into the nucleus and binds to DNA response elements and transactivates gene expression[14,23,24]. The data presented here showed that the positive signals of p-Stat3 were mainly of nuclear type, and the positive rate and strength of p-Stat3 in HCCs were significantly higher than those in PCLTs, and the signal intensity of p-Stat3 was positively correlated with the expression level of the c-jun protein in HCCs and PCLTs. These results suggest that JAKs signaling cascade may also contribute to malignant transformation of hepatocytes besides Ras/Raf/MAPK signaling pathway in hepatocellular carcinogenesis.
Results in literature were inconsistent on the correlation between Ras/Raf/MAPK and JAKs/Stat3 pathways in tumor genesis and development[14,25,26]. Chung et al[25] reported that MAPK activated Stat3 through phosphorylation of Ser-727. Leaman et al[26] found that JAKs may not be required for the activation of Stat3 by some growth factors such as EGF and PDGF. As the MAPK and Stat3 become activated upon many growth factors and cytokines, it was considered that MAPK may promote activation of Stat3, but in Jain’s results MAPK may inhibit transcription activity of Stat3[14]. Our data indicated that there was no correlation between expression intensities of p42/44MAPK and p-Stat3 in HCCs and PCLTs. The Stat3 phosphorylation is a complex event. Because the Stat3 can be phosphorylated through different protein kinases and pathways, detecting Stat3 at different phases of signaling cascades may produce entirely contradictory results about Stat3 activity. We think that both Ras/Raf/MAPK and JAKs/Stat3 pathways may play important roles in hepatocellular carcinogenesis, but their detailed mechanisms need to be further studied.
Up to now, the characteristics of pericar-cinomatous hepatocytes has not been clear[27]. The results showed that pericarcinomatous hepatocytes, especially proliferative or atypical hyperplasia hepatocytes, strongly express p42/44MAPK, p-Stat3, c-fos and c-jun proteins, which suggest that activation of MAPK and Stat3 may be an early event in hepatocellular carcinogenesis. It also implys that the hepatocytes which express p42/44MAPK and/or p-Stat3 may be potentially malignant precancerous cells. Therefore, in order to avoid local recurrence, the operative range should be as extended as possible in the resection of HCCs.
Footnotes
Supported by New England Biolab, MA, USA.
Edited by Ma JY
References
- 1.Ito Y, Sasaki Y, Horimoto M, Wada S, Tanaka Y, Kasahara A, Ueki T, Hirano T, Yamamoto H, Fujimoto J, et al. Activation of mitogen-activated protein kinases/extracellular signal-regulated kinases in human hepatocellular carcinoma. Hepatology. 1998;27:951–958. doi: 10.1002/hep.510270409. [DOI] [PubMed] [Google Scholar]
- 2.Schmidt CM, McKillop IH, Cahill PA, Sitzmann JV. Increased MAPK expression and activity in primary human hepatocellular carcinoma. Biochem Biophys Res Commun. 1997;236:54–58. doi: 10.1006/bbrc.1997.6840. [DOI] [PubMed] [Google Scholar]
- 3.Gu SQ, Liang YY, Fan LR, Li BY, Wang DS. Co-regulative effects of the cAMP/PKA and DAG/PKC signal pathways on human gas-tric cancer cells during differentiation induced by traditional Chi-nese medicines. China Natl J New Gastroen terol. 1997;3:50–53. doi: 10.3748/wjg.v3.i1.50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Hill CS, Treisman R. Transcriptional regulation by extracellular signals: mechanisms and specificity. Cell. 1995;80:199–211. doi: 10.1016/0092-8674(95)90403-4. [DOI] [PubMed] [Google Scholar]
- 5.Kim H, Baumann H. Dual signaling role of the protein tyrosine phosphatase SHP-2 in regulating expression of acute-phase plasma proteins by interleukin-6 cytokine receptors in hepatic cells. Mol Cell Biol. 1999;19:5326–5338. doi: 10.1128/mcb.19.8.5326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Edmondson HA, STEINER PE. Primary carcinoma of the liver: a study of 100 cases among 48900 necropsies. Cancer. 1954;7:462–503. doi: 10.1002/1097-0142(195405)7:3<462::aid-cncr2820070308>3.0.co;2-e. [DOI] [PubMed] [Google Scholar]
- 7.Feng DY, Chen RX, Peng Y, Zheng H, Yan YH. Effect of HCV NS(3) protein on p53 protein expression in hep atocarcinogenesis. World J Gastroenterol. 1999;5:45–46. doi: 10.3748/wjg.v5.i1.45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Fromowitz FB, Viola MV, Chao S, Oravez S, Mishriki Y, Finkel G, Grimson R, Lundy J. ras p21 expression in the progression of breast cancer. Hum Pathol. 1987;18:1268–1275. doi: 10.1016/s0046-8177(87)80412-4. [DOI] [PubMed] [Google Scholar]
- 9.Yu CL, Jove R, Burakoff SJ. Constitutive activation of the Janus kinase-STAT pathway in T lymphoma overexpressing the Lck protein tyrosine kinase. J Immunol. 1997;159:5206–5210. [PubMed] [Google Scholar]
- 10.Yu CL, Burakoff SJ. Involvement of proteasomes in regulating Jak-STAT pathways upon interleukin-2 stimulation. J Biol Chem. 1997;272:14017–14020. doi: 10.1074/jbc.272.22.14017. [DOI] [PubMed] [Google Scholar]
- 11.Leevers SJ, Paterson HF, Marshall CJ. Requirement for Ras in Raf activation is overcome by targeting Raf to the plasma membrane. Nature. 1994;369:411–414. doi: 10.1038/369411a0. [DOI] [PubMed] [Google Scholar]
- 12.Marshall CJ. MAP kinase kinase kinase, MAP kinase kinase and MAP kinase. Curr Opin Genet Dev. 1994;4:82–89. doi: 10.1016/0959-437x(94)90095-7. [DOI] [PubMed] [Google Scholar]
- 13.Stokoe D, Macdonald SG, Cadwallader K, Symons M, Hancock JF. Activation of Raf as a result of recruitment to the plasma membrane. Science. 1994;264:1463–1467. doi: 10.1126/science.7811320. [DOI] [PubMed] [Google Scholar]
- 14.Jain N, Zhang T, Fong SL, Lim CP, Cao X. Repression of Stat3 activity by activation of mitogen-activated protein kinase (MAPK) Oncogene. 1998;17:3157–3167. doi: 10.1038/sj.onc.1202238. [DOI] [PubMed] [Google Scholar]
- 15.Treisman R. Journey to the surface of the cell: Fos regulation and the SRE. EMBO J. 1995;14:4905–4913. doi: 10.1002/j.1460-2075.1995.tb00173.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hoshino R, Chatani Y, Yamori T, Tsuruo T, Oka H, Yoshida O, Shimada Y, Ari-i S, Wada H, Fujimoto J, et al. Constitutive activation of the 41-/43-kDa mitogen-activated protein kinase signaling pathway in human tumors. Oncogene. 1999;18:813–822. doi: 10.1038/sj.onc.1202367. [DOI] [PubMed] [Google Scholar]
- 17.Sivaraman VS, Wang H, Nuovo GJ, Malbon CC. Hyperexpression of mitogen-activated protein kinase in human breast cancer. J Clin Invest. 1997;99:1478–1483. doi: 10.1172/JCI119309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Toyoda M, Hashimoto N, Tokita K, Goldstein BJ, Yokosuka O, Kanatsuka A, Suzuki Y, Saito Y. Increased activity and expression of MAP kinase in HCC model rats induced by 3'-methyl-4-dimethylamino-azobenzene. J Hepatol. 1999;31:725–733. doi: 10.1016/s0168-8278(99)80354-7. [DOI] [PubMed] [Google Scholar]
- 19.McKillop IH, Schmidt CM, Cahill PA, Sitzmann JV. Altered expression of mitogen-activated protein kinases in a rat model of experimental hepatocellular carcinoma. Hepatology. 1997;26:1484–1491. doi: 10.1002/hep.510260615. [DOI] [PubMed] [Google Scholar]
- 20.Arbuthnot P, Capovilla A, Kew M. Putative role of hepatitis B virus X protein in hepatocarcinogenesis: effects on apoptosis, DNA repair, mitogen-activated protein kinase and JAK/STAT pathways. J Gastroenterol Hepatol. 2000;15:357–368. doi: 10.1046/j.1440-1746.2000.02069.x. [DOI] [PubMed] [Google Scholar]
- 21.Ostrowski J, Woszczynski M, Kowalczyk P, Wocial T, Hennig E, Trzeciak L, Janik P, Bomsztyk K. Increased activity of MAP, p70S6 and p90rs kinases is associated with AP-1 activation in spontaneous liver tumours, but not in adjacent tissue in mice. Br J Cancer. 2000;82:1041–1050. doi: 10.1054/bjoc.1999.1040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kaptein A, Paillard V, Saunders M. Dominant negative stat3 mutant inhibits interleukin-6-induced Jak-STAT signal transduction. J Biol Chem. 1996;271:5961–5964. doi: 10.1074/jbc.271.11.5961. [DOI] [PubMed] [Google Scholar]
- 23.Darnell JE, Kerr IM, Stark GR. Jak-STAT pathways and transcriptional activation in response to IFNs and other extracellular signaling proteins. Science. 1994;264:1415–1421. doi: 10.1126/science.8197455. [DOI] [PubMed] [Google Scholar]
- 24.Ihle JN. STATs: signal transducers and activators of transcription. Cell. 1996;84:331–334. doi: 10.1016/s0092-8674(00)81277-5. [DOI] [PubMed] [Google Scholar]
- 25.Chung J, Uchida E, Grammer TC, Blenis J. STAT3 serine phosphorylation by ERK-dependent and -independent pathways negatively modulates its tyrosine phosphorylation. Mol Cell Biol. 1997;17:6508–6516. doi: 10.1128/mcb.17.11.6508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Leaman DW, Pisharody S, Flickinger TW, Commane MA, Schlessinger J, Kerr IM, Levy DE, Stark GR. Roles of JAKs in activation of STATs and stimulation of c-fos gene expression by epidermal growth factor. Mol Cell Biol. 1996;16:369–375. doi: 10.1128/mcb.16.1.369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Feng D, Cheng R, Yan Y. [Comparative study of expression of p21 and c-myc protein in hepatocellular carcinoma with pericarcinomatous liver tissue] Hunan Yike Daxue Xuebao. 1997;22:310–312. [PubMed] [Google Scholar]