

Abstract
There is a critical shortage in the number of deceased human organs that become available for purposes of clinical transplantation. This problem might be resolved by the transplantation or organs from pigs genetically-engineered to protect them from the human immune response. The pathobiological barriers to successful pig organ transplantation in primates include activation of the innate and adaptive immune systems, coagulation dysregulation, and inflammation. Genetic engineering of the pig as an organ source has increased the survival of the transplanted pig heart, kidney, islet and corneal graft in nonhuman primates (NHP) from minutes to months or occasionally years. Genetic engineering may also contribute to any physiological barriers that might be identified as well as to reducing the risks of transfer of a potentially infectious micro-organism with the organ. There are now an estimated 40 or more genetic alterations that have been carried out in pigs, with some pigs expressing 5 or 6 manipulations. With the new technology now available, it will become increasingly common for a pig to express even more genetic manipulations, and these could be tested in the pig-to-NHP models to assess their efficacy and benefit. It is therefore likely that clinical trials of pig kidney, heart, and islet transplantation will become feasible in the near future.
Keywords: Pig, genetically-engineered, Nonhuman primate Xenotransplantation, islets Xenotransplantation, organs
Introduction
The current issue of this journal is directed towards animal models of human disease. This present review is not strictly on this topic, but it is in the closely-related field of genetically-engineering a large animal to overcome pathobiological barriers that currently prevent cross-species organ or cell transplantation.
There is a critical shortage in the number of deceased human organs that become available for purposes of clinical transplantation. For example, in the USA alone, there are currently more than 124,000 patients waiting for transplant and yet only approximately 28,000 organ transplants are carried annually using organs from deceased donors. Despite enormous efforts over the past 50 years, this problem has not been resolved, and in fact is increasing. This is despite the fact that transplant surgeons have liberalized their criteria for selection of the donor; they now use organs from what are termed ‘high risk’, ‘marginal’, or ‘extended criteria’ deceased donors, which are organs from less-than-ideal donors that would not have been used a few years ago. Furthermore, organs from living donors are increasingly being used. With regard to kidney donation, the risk to the living donor is small, though there are some small long-term detrimental sequelae. With regard to donors of partial livers, there is a significant risk of both mortality and morbidity. We are, therefore, putting healthy altruistic humans at some risk in an effort to provide organs for those with end-stage organ failure [1].
Background
Throughout the 20th century, there were attempts to use animals as sources of organs for clinical transplantation [2,3]. These generally involved non-human primates (NHPs), although a few efforts were made using non-primate mammals. In these latter cases, graft survival was extremely short, often measured in minutes, rather than hours or days. Transplantation between immunologically ‘discordant’ species, e.g., pig-to-human, therefore represents a major immunological barrier. However, it was soon realized that NHPs would not be the ideal source of organs or cells for clinical transplantation. Therefore, during the latter part of the 20th century, attention was directed towards using the pig as the potential source of organs and cells. The pig has a number of advantages in this respect although the immunological barriers are far greater than those associated with NHP species.
Without the genetic-engineering of pigs to protect their organs and cells from the primate’s immune response, little progress would have been made. The transplantation of organs from wild-type (genetically-unmodified) pigs into NHPs (or even humans) resulted in rapid antibody-dependent complement-mediated rejection (hyperacute rejection) (Figures 1 and 2), similar to that which occurs when an ABO-incompatible allograft is transplanted between humans. In allotransplantation, hyperacute rejection may also result when the recipient is sensitized to donor human leukocyte antigens (HLA, i.e., has a high level of panel-reactive antibodies [PRA]). The standard pharmacologic immunosuppressive therapy administered to prolong allograft survival has no or little effect in protecting pig organs from hyperacute rejection.
Figure 1.

Macroscopic appearance of a wild-type pig kidney immediately after transplantation and reperfusion in a baboon (A) and 10 minutes later when hyperacute rejection had occurred (B).
Figure 2.
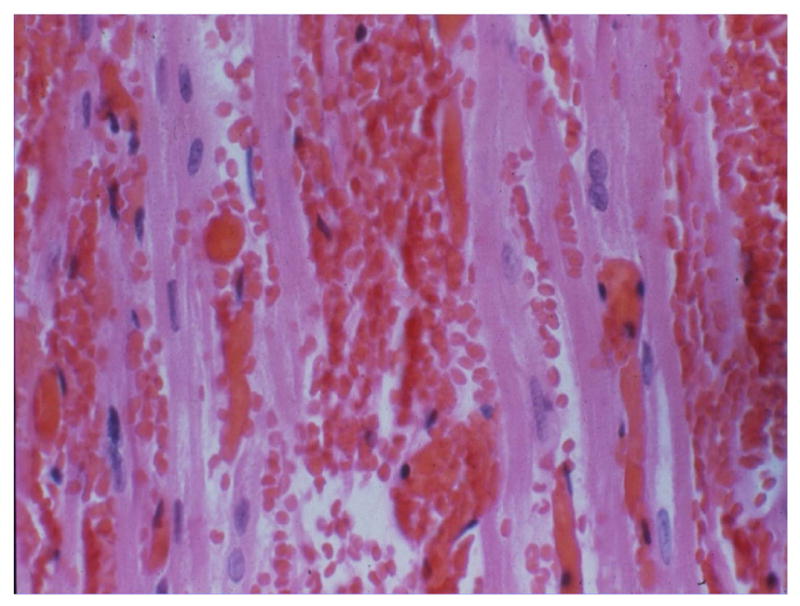
Histopathology of hyperacute rejection in a wild-type pig heart graft. Complement-mediated injury associated with the binding of baboon natural preformed anti-pig antibodies to antigens expressed on the vascular endothelium of the pig organ results in intravascular thrombosis and interstitial haemorrhage. Acute humoral xenograft rejection, a delayed antibody-mediated response, is often, but not always, associated with the production of elicited antibodies, and has a similar histopathological appearance but possibly with the presence of rather more innate immune cells, such as macrophages and neutrophils. Reproduced with permission from Byrne GW, et al. Xenotransplantation 2013;20:292–307 (AQ : ensure that this reference is cited in sequence in the References list and the correct citation [number] used in this legend.)
As the identity of the antigens on the vascular endothelium of the pig organ was unknown in the 1980s, efforts were made to prolong graft survival by removing all immunoglobulins (which included anti-pig antibodies) from the recipient blood by plasmapheresis [4]. The temporary removal of immunoglobulins (antibodies) directed to ABO blood group antigens had indeed proved successful in allowing permanent ABO-incompatible allograft survival [5–7], and so it was hoped that this approach might also suffice in xenotransplantation [8,9]. As the pathobiological differences between pig and primate are so much more complex than between ABO-incompatible primates, this proved not to be the case. The continuing production of anti-pig antibodies resulted in graft destruction (a form of rejection variously termed acute humoral xenograft rejection [AHXR], delayed xenograft rejection, or acute vascular rejection).
Gal antigen expression in pigs
A major step forward was taken when the major carbohydrate antigen on pig vascular endothelial cells, against which humans have anti-pig antibodies, was identified as galactose-α1,3-galactose (Gal) (reviewed in [10]). This resulted in efforts to remove specifically anti-Gal antibodies from the potential recipient using either immunoadsorption with immunoaffinity columns of synthetic Gal oligosaccharides or by the intravenous infusion of the Gal oligosaccharides (which were then bound by anti-Gal antibodies, and the resulting antigen-antibody complexes excreted) (reviewed in [11] and [12]). These approaches proved no more successful than plasmapheresis. Although delaying graft rejection, they were not associated with truly prolonged graft survival even when combined with immunosuppressive therapy that prevented a T cell-dependent elicited antibody response.
Pig genetic engineering to control human complement-mediated injury
Attention then turned to genetic engineering to try to overcome these barriers. The first approach was directed towards protecting the graft from the effects of primate complement. When the complement cascade is activated in humans, for example in response to the presence of pathogenic bacteria in the blood, complement activation generally does not damage the host’s tissues. This is because of the expression of human complement-regulatory proteins (CRPs) on the surface of vascular endothelial cells. Pigs have similar CRPs, but these are relatively inefficient at protecting the pig cells from human complement-mediated injury [13]. It was therefore suggested that pigs should be genetically-manipulated to introduce transgenes for human CRPs, of which there are several (e.g., CD46, CD55, CD59).
At least two independent groups made this original suggestion [14,15], and the first pig expressing a human CRP (CD55, decay-accelerating factor), produced by microinjection of DNA directly into the pronucleus of a fertilized egg, was born in the early 1990s (Table 1) [16–19]. The U.K. group of White and his colleagues performed numerous studies in the pig-to-baboon or pig-to-monkey model and demonstrated that the expression of CD55 prolonged pig heart and kidney graft survival to days or weeks (reviewed in [11]). This was the first demonstration of the benefit of genetically-engineering the pig in overcoming the barriers of xenotransplantation. However, this manipulation alone did not enable truly long-term graft survival.
Table 1.
Timeline for application of evolving techniques for genetic engineering of pigs employed in xenotransplantation
| Year | Technique |
|---|---|
| 1992 | Microinjection of randomly-integrating transgenes |
| 2000 | Somatic cell nuclear transfer (SCNT) |
| 2002 | Homologous recombination |
| 2011 | Zinc finger nucleases (ZFNs) |
| 2013 | Transcription activator-like effector nucleases (TALENs) |
| 2014 | CRISPR/Cas9* |
CRISPR/Cas9 = clustered randomly interspaced short palindromic repeats and the associated protein 9
Pig genetic engineering to delete Gal antigens
When it became possible to delete a gene from a pig, which was not possible until nuclear transfer (cloning) technology was introduced [20,21], efforts were made to delete the gene for the enzyme that attaches the terminal Gal saccharides to the underlying carbohydrates on the pig vascular endothelium [22–24]. Nuclear transfer was combined with homologous recombination technology, and the resulting α1,3-galactosyltransferase gene-knockout (GTKO) pigs were then used as sources of organs for experimental transplantation in NHPs. In baboons selected for low levels of the remaining anti-pig antibodies (so-called anti-nonGal antibodies), the transplantation of GTKO pig hearts and kidneys prolonged graft survival significantly [25–30].
The transplantation of organs from GTKO/hCRP pigs had a further beneficial effect over GTKO or CRP alone [31–33].
Coagulation dysregulation
However, vascular endothelial cell activation by anti-nonGal antibodies (the antigenic targets of which were unknown) (or by anti-Gal antibodies), complement deposition, and the activation of the endothelium by innate immune cells (e.g., neutrophils, monocytes, macrophages) resulted in the development of a thrombotic microangiopathy and graft failure (Figure 3) [25,30,34]. This was a result of fibrin deposition and platelet aggregation in the vessels of the graft, which, when advanced (i.e., when all the recipient coagulation factors had been exhausted) could result in a consumptive coagulopathy. This lead to spontaneous bleeding (e.g., in the gastrointestinal tract) and could be fatal [25,35–37]. Excision of the graft reversed this process, confirming that it was the changes taking place in the graft that was the source of the problem.
Figure 3.
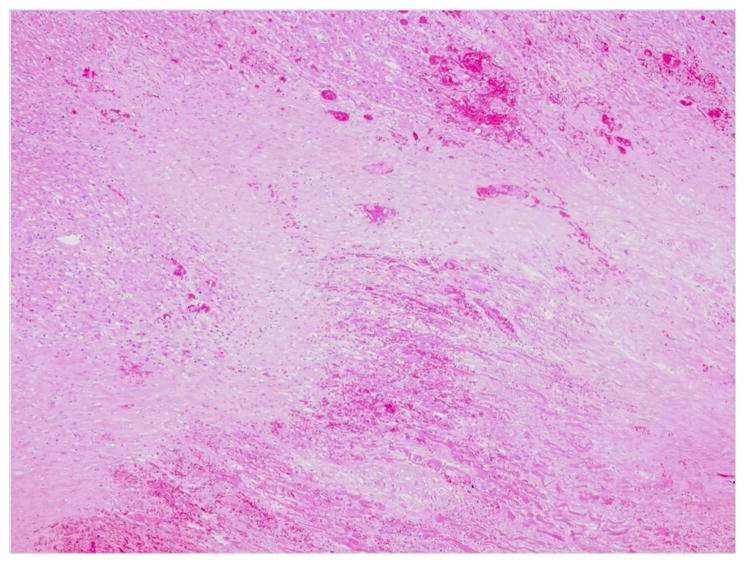
Thrombotic microangiopathy in a GTKO pig heart graft. Occlusion of small vessels by fibrin deposition and platelet aggregation results in ischemic injury with replacement fibrosis.
This development of thrombotic microangiopathy from vascular endothelial activation was enhanced by molecular incompatibilities in the coagulation systems between pig and primate. For example, pig tissue factor pathway inhibitor does not successfully inhibit primate factor Xa, pig thrombomodulin does not catalyze primate protein C, and pig von Willebrand factor is associated with excessive primate platelet aggregation. The mechanism is complicated in that not only do the activated porcine endothelial cells express high levels of tissue factor (a procoagulant molecule) and increased tissue factor activity, but direct exposure of primate platelets and monocytes to porcine endothelial cells results in increased tissue factor activity on these primate structures also [37]. Coagulation-anticoagulation dysregulation of relevance to xenotransplantation has been discussed previously by several authors [38–40].
Pig genetic engineering to correct coagulation dysfunction
It was therefore determined that the pig should be genetically-engineered to protect it from these coagulation discrepancies [41]. As with CRPs, efforts were made to introduce transgenes of human coagulation regulatory proteins (e.g., thrombomodulin, endothelial protein C receptor, tissue factor pathway inhibitor, CD39, CD73) [42–45]. An alternative approach is to reduce expression of tissue factor by small interfering RNA [46].
Multi-gene transgenic pigs have been produced that express not only GTKO and CRPs, but multiple anticoagulant transgenes, including various combinations of thrombomodulin, endothelial protein C receptor, tissue factor pathway inhibitor, and CD39 [41]. The expression of one or more human coagulation-regulatory proteins increases graft survival further. For example, in pig heterotopic heart transplantation (i.e., non-life-supporting grafts), grafts have beat strongly for more than one or even two years [47–49]. After orthotopic heart transplantation, where the pig heart actually replaces the NHP heart and is therefore entirely responsible for support of the circulation, recipient survival has been approximately 2 months (but the demise of the recipient has more commonly been related to complications in the management of the animal than to graft failure) (reviewed in [12]). With pig life-supporting kidney transplantation, survival has been for >6 months in at least one recipient animal [50,51].
McGregor and his colleagues have demonstrated that increased immunosuppressive therapy, thus reducing the immune activation of the pig vascular endothelium, is at least as effective as the administration of anticoagulant agents in delaying the development of thrombotic microangiopathy [52–54]. This adds support to the conclusion that the development of this complication is primarily related to immune activation and injury of the endothelium. Nevertheless, since at present immune activation of the vascular endothelial cells cannot be totally prevented, reducing the problem by correcting the coagulation dysfunction between pig and primate provides an alternative approach towards a solution and may even be essential when there is no immune response.
NonGal antigen expression in pigs
Efforts have been made to identify the structure of nonGal antigens. It has been known since the 1990s that pigs express N-glycolylneuraminic acid (NeuGc), which is not expressed in humans, although it is expressed in all apes and Old World monkeys [55]. As humans do not express NeuGc, they make anti-NeuGc antibodies, just as they make anti-Gal antibodies (reviewed in [56]). It has therefore been predicted for many years that expression of NeuGc might be a factor when clinical xenotransplantation is carried out even if the donor organs are from GTKO pigs. However, NeuGc does not play a role in the experimental laboratory model as both pigs and NHPs express this saccharide, and therefore anti-NeuGc antibodies are not involved in the immune response.
Recently, pigs that express neither Gal nor NeuGc have been produced [57,58]. In vitro studies using human serum have demonstrated that there is reduced human antibody binding to cells from these GTKO/NeuGcKO pigs compared to binding to GTKO pig cells [59]. It is therefore likely to be important to utilize organs from pigs that express neither Gal nor NeuGc (i.e., GTKO/NeuGcKO pigs) for clinical organ xenotransplantation.
A second nonGal antigenic target for primate anti-pig antibodies has recently been identified, β1,4 N-acetylgalactosaminyltransferase [60]. Baboons have pre-formed antibodies to this glycan, as do most humans [59]. There is preliminary evidence that the absence of this antigen on pig cells also reduces human serum antibody binding [59]. As this antigen (like Gal) is not expressed in NHPs, the pig-to-NHP model will be valuable in determining the relative importance of its effect. This pig antigen may also need to be deleted in pigs used as organ-sources in clinical trials of xenotransplantation. Tector’s group has produced pigs that express neither Gal, NeuGc, nor β1,4 N-acetylgalactosaminyltransferase [59]. In vitro evidence therefore suggests that pigs that do not express NeuGc or Gal (and possibly β1,4 N-acetylgalactosaminyltransferase), but do express one or more human CRPs and one or more human coagulation-regulatory proteins that (AQ: as intended?) may be beneficial for clinical xenotransplantation.
Pig genetic engineering to suppress the inflammatory response
There are other genetic modifications that may provide further benefit. For example, there is increasing evidence that a primate inflammatory response to pig grafts is playing a significant role in graft failure, a condition that Ezzelarab has called a ‘systemic inflammatory response in xenograft recipients’ (SIXR) [61,62]. It is known that interleukin-6 (IL-6) plays a significant role in this response, but other cytokines and chemokines are also involved. It may therefore be beneficial to develop pigs that are transgenic for one or more human anti-inflammatory gene(s), e.g., hemeoxygenase-1 or A20. Transgenic pigs expressing either A20 or HO1 are available [63–65], and these have recently become available on a GTKO/hCRP background (Ayares D, unpublished). While there is preliminary evidence, from ex vivo organ perfusion experiments, of a functional effect of these transgenes, it has not yet been clarified in vivo.
The adaptive (T cell) immune response
Whatever genetically-engineered pig is used as the organ source, the NHP recipient will develop a T cell-dependent (adaptive) immune response that will lead to graft infiltration with T cells and other immune cells as well as a major increase in elicited anti-pig (largely IgG) antibodies. This response will inevitably result in graft rejection [66]. Therefore, an effective immunosuppressive regimen has to be administered to the recipient animal to prevent this response. The standard pharmacologic immunosuppressive agents, for example tacrolimus, cyclosporine, mycophenolate mofetil, rapamycin, even when employed in combinations that are effective in controlling allotransplant rejection, are not particularly successful in xenotransplantation, unless given in high doses that are inevitably associated with an increase in complications, particularly infection [67,68]. Newer agents that prevent T cell costimulation (and are therefore known as costimulation blockade agents) have proven more successful.
The initial agent (directed towards blocking the CD40/CD154 costimulation pathway) was an anti-CD154 monoclonal antibody (mAb), first introduced into xenotransplantation studies by Buhler and colleagues in 2000 [69]. This proved particularly effective in preventing an elicited antibody response, but has been shown to be associated with some thrombogenic effects (both in xenotransplantation and allotransplantation) and therefore may not be available for clinical use. An alternative is an anti-CD40 mAb, that also inhibits the CD40/CD154 costimulation pathway, and this has been associated with excellent results, particularly by the groups of Mohiuddin [47–49] and Iwase [51,70].
An alternative costimulation pathway, namely the B7/CD28 pathway, can be blocked by commercially-available agents, such as CTLA4-Ig (abatacept) and a modified version (belatacept), but this has proved less successful. Belatacept prevents a T cell proliferative response to a pig graft in vivo, but it does not prevent an elicited antibody response [50,70–72]. Ezzelarab has suggested that other mechanisms of inducing sensitization may be involved [61,73]. There may be a case for blockade of both the CD40/CD154 and B7/CD28 pathways [70].
The less systemic immunosuppressive therapy that is administered to the recipient, then the less likelihood there will be of major complications related to this therapy. Attention is therefore being directed to protect the graft from the T cell response by genetic manipulation of the organ-source pig.
Pig genetic engineering to suppress the adaptive immune response
Surprisingly, but fortunately, genetic engineering of the organ-source pigs directed towards protection from the innate response in some cases also reduces the adaptive immune response [74]. Deletion of Gal antigens [75] or expression of a human CRP [76,77] significantly reduces the in vitro T cell response to pig cells, though the nature of the experiments is such that it is difficult to measure this in vivo.
The gene for CTLA4-Ig has been introduced into the pig; this was carried out successfully by Phelps and colleagues in 2009 [78] (AQ: please check your meaning has been retained). These pigs expressed CTLA4-Ig to such an extent that the levels of soluble CTLA4-Ig in the blood were on occasions many times higher than the therapeutic levels required after administration of the agent to a patient with an allograft. The genetic manipulation, therefore, had been extremely successful. However, once weaned from the sow, and thus no longer receiving immunoglobulin in the milk, these piglets became immunocompromised and susceptible to infection, precluding their survival to an age at which they could reproduce.
Since then, methods to express the transgene only in specific cell types have been developed, and pigs have been produced that express CTLA4-Ig only in the pancreatic islet beta cells [79,80], vascular endothelial cells, or in certain neuronal cells (that have been used to try to correct Parkinson-like condition in monkeys [81,82]. Whether expression confined to the endothelial cells of an organ graft or to the beta cells of the islets will be sufficient to protect the organ or islets from the adaptive immune response remains uncertain, although it is likely to have some beneficial effect and may allow reduced systemic immunosuppressive therapy to be administered. It is difficult to determine this in the current animal models of xenotransplantation.
An alternative approach has been to introduce a dominant-negative mutant major histocompatibility complex (MHC) class II transactivator gene that results in reduced expression of swine leukocyte antigen (SLA) class II on the vascular endothelium of the pig organs [83]. This has been clearly demonstrated to reduce the T cell response in vitro [84], and there is a report indicating a modest effect in vivo [72]. This would therefore hopefully allow a reduced intensity of systemic immunosuppressive therapy.
Deletion by knockout of MHC class I expression in the pig has recently been achieved and its effect is being explored [85].
Over-expression of human CD47 in transgenic pigs, with potential for ‘immune cloaking’ that prevents macrophage activation and phagocytosis of CD47 transgenic pig cells, is also being tested [86]. It is possible that peripheral tolerance can be induced in the host if mixed hematopoietic chimaerism can be established using CD47 transgenic pig progenitor cells. Three-gene transgenic pigs (GTKO/CD46/CD47) have been generated that robustly express both CD46 and CD47 for use in ex vivo lung perfusion and in vivo lung transplantation NHP models (Phelps C, unpublished), as well as in pig-to-NHP studies of tolerance induction.
Natural killer cells also play a role in xenograft rejection [87–91], and here again genetic engineering of the pig (to express HLA-E and/or G and/or Cw3) may prevent this [92–99].
Pig genetic engineering to correct physiological incompatibilities
If problems related to the immune response can be completely resolved, then attention can be directed to whether pig organs will function normally in primate hosts. There have been few studies in this respect [100,101]. These indicate that some problems initially perceived to be related to physiological differences between pig and primate, were in fact related to the effects of the immune response. When the immune response had been adequately controlled, organ function remained normal.
For example, early pig kidney transplants were associated with high levels of proteinuria that resulted in hypoalbuminemia [27,100]. When protection from the immune response was increased by the genetic engineering of the pig, this complication was greatly reduced or not seen. The fact that heterotopic pig hearts have functioned well in NHPs for >12 months, and life-supporting hearts for almost 2 months, suggests that pig myocardial function will not be compromised in a primate host.
With regard to genetically-engineered pig islet transplantation in diabetic NHPs, normoglycemia has been maintained for >1 year, confirming that pig insulin (which has only one amino acid difference from human insulin, and was used therapeutically in diabetic patients for decades) will be sufficient to correct diabetes after clinical islet xenotransplantation [79,102].
Pig livers have only functioned in NHPs for periods of a few days (largely associated with specific coagulation problems that have been identified after pig liver transplantation, particularly relating to the phagocytosis of the host platelets by the pig vascular endothelial cells and Kupffer cells in the graft [103–105]. These problems are also being addressed by genetic engineering of the pigs [106,107]. However, observations have been made that indicate that pig coagulation factors are at least partially functional in NHPs [108].
Nevertheless, it is unlikely that all of the myriad products of a pig liver will be able to fulfil the requirements of a primate. This problem can also be resolved by genetic engineering of the pig. For example, if pig albumin is found to be inadequate in a human host, then the pig could be genetically manipulated to produce human albumin.
Pig genetic engineering to ensure the safety of xenotransplantation
One final area that has received considerable attention over a number of years is the potential risk of transferring a porcine virus or micro-organism to the recipient of the graft and, of greater importance, the potential for that micro-organism to be transferred to close contacts of the recipient, e.g., family members, medical or nursing staff. This could provide a potential risk to the community, particularly as humans may not have a natural immunity to this microorganism. The organ-source pigs will be housed under isolation conditions and will be monitored closely, and so it will be known if they are carrying any potentially infectious micro-organisms that could cause problems in the transplant recipient. It should be noted that this is often not the case in allotransplantation when viruses or micro-organisms are knowingly transferred (such as cytomegalovirus and Epstein-Barr virus), and on occasions may unknowingly be transferred, for example human immunodeficiency (HIV), West Nile, or rabies viruses.
Expert opinion is that, by breeding, housing, and management of the donor pigs in designated pathogen-free (DPF) indoor facilities, it should be possible to prevent all significant pathogenic micro-organisms from being transferred with the pig organ, though endogenous retroviruses may be one exception [109–111].
The genome of every human cell contains viruses and virus particles known as human endogenous retroviruses (HERVs). It is not believed that these have any pathogenic effect and do not play any role in diseases that develop in humans. Similarly, pig genomes contain porcine endogenous retroviruses (PERVs), which are also thought not to be associated with any disease process in pigs. Concern has been raised, however, that the transfer of PERVs to humans might be pathogenic, or that recombination between PERVs and HERVs may give rise to a new virus that may be pathogenic [111,112]. Current opinion is that PERVs are not likely to be pathogenic to humans, but it would be possible to prevent activation of these viruses by genetic engineering of the pig if it were deemed necessary. For example, PERV activation can be suppressed by small interfering RNA technology [113–115].
The present status of pig genetic engineering for xenotransplantation
Ten years ago, developing a pig with a single genetic modification was a prolonged process, involving either pronuclear injection to add genes or homologous recombination for gene-knockout. Techniques by which this can be achieved more rapidly, including the addition of co-expressed strings of transgenes using multi-cistronic 2A technology, have been introduced [41].
Recently, new methodologies have been developed using gene editing nuclease technology, and it is now possible to produce pigs with multiple genetic modifications simultaneously (Table 1) [116,117]. The techniques of zinc finger nucleases (ZFNs) [118–120], transcription activator-like effector nucleases (TALENS) [121,122] and genome editing by RNA-guided endonucleases (also known as clustered regularly interspaced short palindromic repeat [CRISPR]-Cas [CRISPR-associated]) significantly increase gene-editing efficiency.
CRISPRs are short, directly-repeating nucleotide sequences that alternate with small unique DNA fragments acquired from invading bacteriophages or plasmids, and are transcribed into target-specific RNA. The CRISPRs hybridize and form a complex with Cas9 nuclease that recognizes and cleaves foreign genetic material matching the CRISPR-derived RNA [123]. This complex CRISPR/Cas9 system has been developed into an elegant genome editing tool that allows the rapid and efficient production of pigs with multiple genetic modifications [116,124–126], which is having an impact on developing new pigs for xenotransplantation.
Recent studies by Tector’s group have shown that it is possible to produce pigs in which there are variable genetic manipulations demonstrated in different members of the litter [58,59]. These piglets may demonstrate 1, 2, or 3 genetic manipulations, thus providing a range of piglets from the same sow that can be tested in vitro and in vivo. These new technologies have increased the speed at which pigs with multiple genetic manipulations can be produced, and have also reduced the costs. It is likely, therefore, that more rapid progress will be achieved in the genetic manipulation of pigs for the specific purposes of xenotransplantation.
However, the ability to introduce multiple genetic manipulations simultaneously could lead to increased difficulty in determining exactly which manipulation is essential and which superfluous. Obviously, if more than one manipulation has been introduced, it may be difficult to determine which genetic alteration promoted a beneficial outcome following transplantation of a pig organ into a NHP. Furthermore, some manipulations have already been shown to be lethal or detrimental to the pig’s health, and it remains unknown whether there will be a maximum number of manipulations that can be tolerated by a pig.
There are now an estimated 40 or more genetic manipulations that have been carried out in pigs (Table 2), with some pigs expressing 5 or 6 manipulations [70,79,80]. With the technology now available, it will become increasingly common for a pig to express even more genetic manipulations, and these could be tested in the pig-to-NHP models to assess their efficacy and benefit [127]. It is therefore very likely that progress will be rapid during the next few years, and clinical trials of pig kidney, heart, and islet transplantation will become feasible.
Table 2.
Selected genetically-modified pigs currently available for xenotransplantation research
| Complement regulation by human complement-regulatory gene expression |
| CD46 (membrane cofactor protein) |
| CD55 (decay-accelerating factor) |
| CD59 (protectin or membrane inhibitor of reactive lysis) |
| Gal or nonGal antigen ‘masking’ or deletion |
| Human H-transferase gene expression (expression of blood type O antigen) |
| Endo-beta-galactosidase C (reduction of Gal antigen expression) |
| α1,3-galactosyltransferase gene-knockout (GTKO) |
| Cytidine monophosphate-N-acetylneuraminic acid hydroxylase (CMAH) gene-knockout (NeuGcKO) |
| β4GalNT2 (β1,4 N-acetylgalactosaminyltransferase) gene-knockout (β4GalNT2KO) |
| Suppression of cellular immune response by gene expression or downregulation |
| CIITA-DN (MHC class II transactivator knockdown, resulting in swine leukocyte antigen class II knockdown) |
| Class I MHC-knockout (MHC-IKO) |
| HLA-E/human β2-microglobulin (inhibits human natural killer cell cytotoxicity) |
| Human FAS ligand (CD95L) |
| Human GnT-III (N-acetylglucosaminyltransferase III) gene |
| Porcine CTLA4-Ig (Cytotoxic T-Lymphocyte Antigen 4 or CD152) |
| Human TRAIL (tumor necrosis factor-alpha-related apoptosis-inducing ligand) |
| Anticoagulation and anti-inflammatory gene expression or deletion |
| von Willebrand factor (vWF)-deficient (natural mutant) |
| Human tissue factor pathway inhibitor (TFPI) |
| Human thrombomodulin |
| Human endothelial protein C receptor (EPCR) |
| Human CD39 (ectonucleoside triphosphate diphosphohydrolase-1) |
| Anticoagulation, anti-inflammatory, and anti-apoptotic gene expression |
| Human A20 (tumor necrosis factor-alpha-induced protein 3) |
| Human heme oxygenase-1 (HO-1) |
| Human CD47 (species-specific interaction with SIRP-α inhibits phagocytosis) |
| Porcine asialoglycoprotein receptor 1 gene-knockout (ASGR1-KO) (decreases platelet phagocytosis) |
| Human signal regulatory protein α (SIRPα) (decreases platelet phagocytosis by ‘self’ recognition) |
| Prevention of porcine endogenous retrovirus (PERV) activation |
| PERV siRNA |
Clinical trials of xenotransplantation
There are already clinical trials taking place of encapsulated pig islet transplantation [128] and of pig corneal transplantation. In the former case, results have to date been disappointing, but pig corneal transplantation has shown encouraging results [129]. Organ transplantation will follow when there is sufficient evidence in NHPs of consistent survival of the recipients of a life-supporting graft for periods of 6 or 12 months in the absence of complications from over-intensive immunosuppression therapy. Consideration is already being given to the selection of potential patients for these early clinical trials.
For example, patients awaiting kidney allotransplants who have a high level of anti-HLA antibody (with panel reactive antibodies [PRA] that may be 100%, indicating that it would be exceedingly difficult to find a human donor against whom there will not be a high risk of hyperacute rejection) may be suitable candidates for an initial clinical trial of pig kidney transplantation. Current limited evidence is that patients with a high PRA are at no greater risk of rejecting a pig graft than patients with a low level or no PRA [130–132]. Furthermore, there is limited evidence that, even if a patient with a pig graft became sensitized to pig antigens, this would not preclude him or her from undergoing a subsequent allotransplant if a suitable donor could be found [132–134].
Other potential patients include those who might be ‘bridged’ by a pig heart while they await a human heart, although left ventricular assist devices can support a patient for several months or even 1–2 years. In the case of patients with fulminant liver failure, however, there is no similar device to support them and they might benefit from bridging with a pig liver (if the coagulation problems referred to above can be overcome). In an emergency, a human liver can often become available within days, and bridging with a pig organ would be short-term, but potentially life-saving.
Alternative approaches to resolving the shortage of deceased human organs for transplantation include regenerative medicine approaches, stem cell technology, and blastocyst complementation, which have been discussed elsewhere (135). None of these approaches has yet reached the stage of testing in NHPs, and so remain less advanced than xenotransplantation. Mechanical assist devices to support or replace a failing heart, however, are in a much more advanced state than xenotransplantation (135), though a biological heart will have some advantages.
Comment
In 1982, our senior colleague, Thomas Starzl, one of the great pioneers of allotransplantation, wrote, “History tells us that procedures which were inconceivable yesterday, and are barely achievable today, often become routine tomorrow.” We believe that, with the novel genetically-engineered pigs becoming available to us, clinical pig organ and cell transplantation, inconceivable only a few years ago, and barely achievable today, will become routine tomorrow. Indeed, allotransplantation will eventually become of historic interest only.
Acknowledgments
Work on xenotransplantation in the Thomas E. Starzl Transplantation Institute at the University of Pittsburgh has been supported in part by NIH grants U01 AI068642, R21 AI074844, and U19 AI090959, and by Sponsored Research Agreements between the University of Pittsburgh and Revivicor, Inc., Blacksburg, VA.
Abbreviations
- Gal
galactose-α1,3-galactose
- GTKO
α1,3-galactosyltransferase gene-knockout
- HERV
human endogenous retrovirus
- NeuGc
N-glycolylneuraminic acid
- NeuGcKO
NeuGc gene-knockout
- NHP
nonhuman primate
- PERV
porcine endogenous retrovirus
Footnotes
Conflicts of interest: DKCC and BE have no conflict of interest. JR, CP, and DA are employees of Revivicor, Inc.
Authors’ Contributions
DKCC wrote the manuscript; and BE, JR, CP, and DA edited the final version.
References
- 1.Cooper DK. The case for xenotransplantation. Clin Transplant. 2015;29:288–293. doi: 10.1111/ctr.12522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Taniguchi S, Cooper DK. Clinical xenotransplantation: past, present and future. Ann R Coll Surg Engl. 1997;79:13–19. [PMC free article] [PubMed] [Google Scholar]
- 3.Cooper DK. A brief history of cross-species organ transplantation. Proc (Bayl Univ Med Cent) 2012;25:49–57. doi: 10.1080/08998280.2012.11928783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Alexandre GPJ, Gianello P, Latinne D, et al. Plasmapheresis and splenectomy in experimental renal xenotransplantation Xenograft 25. In: Hardy M, editor. Excerpta Medica. Amsterdam, New York, Oxford: 1989. pp. 259–266. [Google Scholar]
- 5.Alexandre GP, De Bruyere M, Squifflet JP, et al. Human ABO-incompatible living donor renal homografts. Neth J Med. 1985;28:231–234. [PubMed] [Google Scholar]
- 6.Alexandre GP, Squifflet JP, De Bruyere M, et al. Present experiences in a series of 26 ABO-incompatible living donor renal allografts. Transplant Proc. 1987;19:4538–4542. [PubMed] [Google Scholar]
- 7.Cooper DK, Ye Y, Niekrasz M, et al. Specific intravenous carbohydrate therapy. A new concept in inhibiting antibody-mediated rejection--experience with ABO-incompatible cardiac allografting in the baboon. Transplantation. 1993;56:769–777. doi: 10.1097/00007890-199310000-00001. [DOI] [PubMed] [Google Scholar]
- 8.Alexandre GP. From ABO-incompatible human kidney transplantation to xenotransplantation. Xenotransplantation. 2004;11:233–236. doi: 10.1111/j.1399-3089.2004.00105.x. [DOI] [PubMed] [Google Scholar]
- 9.Stussi G, West L, Cooper DK, et al. ABO-incompatible allotransplantation as a basis for clinical xenotransplantation. Xenotransplantation. 2006;13:390–399. doi: 10.1111/j.1399-3089.2006.00324.x. [DOI] [PubMed] [Google Scholar]
- 10.Kobayashi T, Cooper DK. Anti-Gal, alpha-Gal epitopes, and xenotransplantation. Subcell Biochem. 1999;32:229–257. doi: 10.1007/978-1-4615-4771-6_10. [DOI] [PubMed] [Google Scholar]
- 11.Lambrigts D, Sachs DH, Cooper DK. Discordant organ xenotransplantation in primates: world experience and current status. Transplantation. 1998;66:547–561. doi: 10.1097/00007890-199809150-00001. [DOI] [PubMed] [Google Scholar]
- 12.Cooper DK, Satyananda V, Ekser B, et al. Progress in pig-to-nonhuman primate transplantation models (1998–2013): a comprehensive review of the literature. Xenotransplantation. 2014;21:397–419. doi: 10.1111/xen.12127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Atkinson JP, Oglesby TJ, White D, et al. Separation of self from non-self in the complement system: a role for membrane cofactor protein and decay accelerating factor. Clin Exp Immunol. 1991;86(Suppl 1):27–30. doi: 10.1111/j.1365-2249.1991.tb06203.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Dalmasso AP, Vercellotti GM, Platt JL, et al. Inhibition of complement-mediated endothelial cell cytotoxicity by decay-accelerating factor. Potential for prevention of xenograft hyperacute rejection. Transplantation. 1991;52:530–533. doi: 10.1097/00007890-199109000-00029. [DOI] [PubMed] [Google Scholar]
- 15.White DJ, Oglesby T, Liszewski MK, et al. Expression of human decay accelerating factor or membrane cofactor protein genes on mouse cells inhibits lysis by human complement. Transpl Int. 1992;5(Suppl 1):S648–650. doi: 10.1007/978-3-642-77423-2_190. [DOI] [PubMed] [Google Scholar]
- 16.White DJG, Langford GA, Cozzi E, et al. Production of pigs trangenic for human DAF: a strategy for xenotransplantation. Xenotransplantation. 1995;2:213–217. [Google Scholar]
- 17.Rosengard AM, Cary NR, Langford GA, et al. Tissue expression of human complement inhibitor, decay-accelerating factor, in transgenic pigs. A potential approach for preventing xenograft rejection. Transplantation. 1995;59:1325–1333. [PubMed] [Google Scholar]
- 18.Cozzi E, White DJ. The generation of transgenic pigs as potential organ donors for humans. Nat Med. 1995;1:964–966. doi: 10.1038/nm0995-964. [DOI] [PubMed] [Google Scholar]
- 19.Cozzi E, Yannoutsos N, Langford GA, et al. Effects of transgenic expression of human decay accelerating factor on the inhibition of hyperacute rejection of pig organs. In: Cooper D, Kemp E, Platt JL, White DJG, editors. Xenotransplantation - the transplantation of organs and tissues betwen species. 2. Springer; Berlin: 1997. pp. 665–682. [Google Scholar]
- 20.Campbell KH, McWhir J, Ritchie WA, et al. Sheep cloned by nuclear transfer from a cultured cell line. Nature. 1996;380:64–66. doi: 10.1038/380064a0. [DOI] [PubMed] [Google Scholar]
- 21.Polejaeva IA, Chen SH, Vaught TD, et al. Cloned pigs produced by nuclear transfer from adult somatic cells. Nature. 2000;407:86–90. doi: 10.1038/35024082. [DOI] [PubMed] [Google Scholar]
- 22.Cooper DK, Koren E, Oriol R. Genetically engineered pigs. Lancet. 1993;342:682–683. doi: 10.1016/0140-6736(93)91791-j. [DOI] [PubMed] [Google Scholar]
- 23.Phelps CJ, Koike C, Vaught TD, et al. Production of alpha 1,3- galactosyltransferase-deficient pigs. Science. 2003;299:411–414. doi: 10.1126/science.1078942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kolber-Simonds D, Lai L, Watt SR, et al. Production of alpha-1,3-galactosyltransferase null pigs by means of nuclear transfer with fibroblasts bearing loss of heterozygosity mutations. Proc Natl Acad Sci U S A. 2004;101:7335–7340. doi: 10.1073/pnas.0307819101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kuwaki K, Tseng YL, Dor FJ, et al. Heart transplantation in baboons using alpha1,3-galactosyltransferase gene-knockout pigs as donors: initial experience. Nat Med. 2005;11:29–31. doi: 10.1038/nm1171. [DOI] [PubMed] [Google Scholar]
- 26.Tseng YL, Kuwaki K, Dor FJ, et al. alpha1,3-Galactosyltransferase gene-knockout pig heart transplantation in baboons with survival approaching 6 months. Transplantation. 2005;80:1493–1500. doi: 10.1097/01.tp.0000181397.41143.fa. [DOI] [PubMed] [Google Scholar]
- 27.Yamada K, Yazawa K, Shimizu A, et al. Marked prolongation of porcine renal xenograft survival in baboons through the use of alpha1,3-galactosyltransferase gene-knockout donors and the cotransplantation of vascularized thymic tissue. Nat Med. 2005;11:32–34. doi: 10.1038/nm1172. [DOI] [PubMed] [Google Scholar]
- 28.Hisashi Y, Yamada K, Kuwaki K, et al. Rejection of cardiac xenografts transplanted from alpha1,3-galactosyltransferase gene-knockout (GalT-KO) pigs to baboons. Am J Transplant. 2008;8:2516–2526. doi: 10.1111/j.1600-6143.2008.02444.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Shimizu A, Yamada K, Yamamoto S, et al. Thrombotic microangiopathic glomerulopathy in human decay accelerating factor-transgenic swine-to-baboon kidney xenografts. J Am Soc Nephrol. 2005;16:2732–2745. doi: 10.1681/ASN.2004121148. [DOI] [PubMed] [Google Scholar]
- 30.Shimizu A, Hisashi Y, Kuwaki K, et al. Thrombotic microangiopathy associated with humoral rejection of cardiac xenografts from alpha1,3-galactosyltransferase gene-knockout pigs in baboons. Am J Pathol. 2008;172:1471–1481. doi: 10.2353/ajpath.2008.070672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.McGregor CG, Ricci D, Miyagi N, et al. Human CD55 expression blocks hyperacute rejection and restricts complement activation in Gal knockout cardiac xenografts. Transplantation. 2012;93:686–692. doi: 10.1097/TP.0b013e3182472850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Azimzadeh A, Kelishadi S, Ezzelarab M, et al. Early graft failure of GTKO pig organs in baboons is reduced in hCPRP expression. Xenotransplantation. 2009;16:356. doi: 10.1111/xen.12176. (Abstract) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Azimzadeh A, Kelishadi S, Ezzelarab MB, et al. Early graft failure of GalTKO pig organs in baboons is reduced by expression of a human complement-regulatory protein. Xenotransplantation. 2015 doi: 10.1111/xen.12176. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Houser SL, Kuwaki K, Knosalla C, et al. Thrombotic microangiopathy and graft arteriopathy in pig hearts following transplantation into baboons. Xenotransplantation. 2004;11:416–425. doi: 10.1111/j.1399-3089.2004.00155.x. [DOI] [PubMed] [Google Scholar]
- 35.Buhler L, Basker M, Alwayn IP, et al. Coagulation and thrombotic disorders associated with pig organ and hematopoietic cell transplantation in nonhuman primates. Transplantation. 2000;70:1323–1331. doi: 10.1097/00007890-200011150-00010. [DOI] [PubMed] [Google Scholar]
- 36.Cowan PJ, Aminian A, Barlow H, et al. Renal xenografts from triple-transgenic pigs are not hyperacutely rejected but cause coagulopathy in non-immunosuppressed baboons. Transplantation. 2000;69:2504–2515. doi: 10.1097/00007890-200006270-00008. [DOI] [PubMed] [Google Scholar]
- 37.Lin CC, Ezzelarab M, Shapiro R, et al. Recipient tissue factor expression is associated with consumptive coagulopathy in pig-to-primate kidney xenotransplantation. Am J Transplant. 2010;10:1556–1568. doi: 10.1111/j.1600-6143.2010.03147.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Robson SC, Cooper DK, d’Apice AJ. Disordered regulation of coagulation and platelet activation in xenotransplantation. Xenotransplantation. 2000;7:166–176. doi: 10.1034/j.1399-3089.2000.00067.x. [DOI] [PubMed] [Google Scholar]
- 39.Cowan PJ, Roussel JC, d’Apice AJ. The vascular and coagulation issues in xenotransplantation. Curr Opin Organ Transplant. 2009;14:161–167. doi: 10.1097/mot.0b013e3283279591. [DOI] [PubMed] [Google Scholar]
- 40.Cowan PJ, Robson SC, d’Apice AJ. Controlling coagulation dysregulation in xenotransplantation. Curr Opin Organ Transplant. 2011;16:214–221. doi: 10.1097/MOT.0b013e3283446c65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ayares D, Vaught T, Ball S, et al. Genetic engineering of source pigs for xenotransplantation: progress and prospects. Xenotransplantation. 2013;20:361. (Abstract 408) [Google Scholar]
- 42.Roussel JC, Moran CJ, Salvaris EJ, et al. Pig thrombomodulin binds human thrombin but is a poor cofactor for activation of human protein C and TAFI. Am J Transplant. 2008;8:1101–1112. doi: 10.1111/j.1600-6143.2008.02210.x. [DOI] [PubMed] [Google Scholar]
- 43.Petersen B, Ramackers W, Tiede A, et al. Pigs transgenic for human thrombomodulin have elevated production of activated protein C. Xenotransplantation. 2009;16:486–495. doi: 10.1111/j.1399-3089.2009.00537.x. [DOI] [PubMed] [Google Scholar]
- 44.Miwa Y, Yamamoto K, Onishi A, et al. Potential value of human thrombomodulin and DAF expression for coagulation control in pig-to-human xenotransplantation. Xenotransplantation. 2010;17:26–37. doi: 10.1111/j.1399-3089.2009.00555.x. [DOI] [PubMed] [Google Scholar]
- 45.Lee KF, Salvaris EJ, Roussel JC, et al. Recombinant pig TFPI efficiently regulates human tissue factor pathways. Xenotransplantation. 2008;15:191–197. doi: 10.1111/j.1399-3089.2008.00476.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Ahrens HE, Petersen B, Herrmann D, et al. siRNA mediated knockdown of tissue factor expression in pigs for xenotransplantation. Am J Transplant. 2015;15:1407–1414. doi: 10.1111/ajt.13120. [DOI] [PubMed] [Google Scholar]
- 47.Mohiuddin MM, Singh AK, Corcoran PC, et al. Genetically engineered pigs and target-specific immunomodulation provide significant graft survival and hope for clinical cardiac xenotransplantation. J Thorac Cardiovasc Surg. 2014;148:1106–1113. doi: 10.1016/j.jtcvs.2014.06.002. discussion 1113–1104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Mohiuddin MM, Singh AK, Corcoran PC, et al. Role of anti-CD40 antibody-mediated costimulation blockade on non-Gal antibody production and heterotopic cardiac xenograft survival in a GTKO. hCD46Tg pig-to-baboon model. Xenotransplantation. 2014;21:35–45. doi: 10.1111/xen.12066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Mohiuddin MM, Singh AK, Corcoran PC, et al. One-year heterotopic cardiac xenograft survival in a pig to baboon model. Am J Transplant. 2014;14:488–489. doi: 10.1111/ajt.12562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Higginbotham L, Mathews D, Breeden CA, et al. Pre-transplant antibody screening and anti-CD154 costimulation blockade promote long-term xenograft survival in a pig-to-primate kidney transplant model. Xenotransplantation. 2015;22:221–230. doi: 10.1111/xen.12166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Iwase H, Liu H, Wijkstrom M, et al. Pig kidney graft survival in a baboon for 136 days: longest life-supporting organ graft survival to date. Xenotransplantation. 2015 doi: 10.1111/xen.12174. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Schirmer JM, Fass DN, Byrne GW, et al. Effective antiplatelet therapy does not prolong transgenic pig to baboon cardiac xenograft survival. Xenotransplantation. 2004;11:436–443. doi: 10.1111/j.1399-3089.2004.00159.x. [DOI] [PubMed] [Google Scholar]
- 53.Byrne GW, Schirmer JM, Fass DN, et al. Warfarin or low-molecular-weight heparin therapy does not prolong pig-to-primate cardiac xenograft function. Am J Transplant. 2005;5:1011–1020. doi: 10.1111/j.1600-6143.2005.00792.x. [DOI] [PubMed] [Google Scholar]
- 54.Byrne GW, Davies WR, Oi K, et al. Increased immunosuppression, not anticoagulation, extends cardiac xenograft survival. Transplantation. 2006;82:1787–1791. doi: 10.1097/01.tp.0000251387.40499.0f. [DOI] [PubMed] [Google Scholar]
- 55.Bouhours D, Pourcel C, Bouhours JE. Simultaneous expression by porcine aorta endothelial cells of glycosphingolipids bearing the major epitope for human xenoreactive antibodies (Gal alpha 1–3Gal), blood group H determinant and N-glycolylneuraminic acid. Glycoconj J. 1996;13:947–953. doi: 10.1007/BF01053190. [DOI] [PubMed] [Google Scholar]
- 56.Padler-Karavani V, Varki A. Potential impact of the non-human sialic acid N- glycolylneuraminic acid on transplant rejection risk. Xenotransplantation. 2011;18:1–5. doi: 10.1111/j.1399-3089.2011.00622.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Lutz AJ, Li P, Estrada JL, et al. Double knockout pigs deficient in N-glycolylneuraminic acid and galactose alpha-1,3-galactose reduce the humoral barrier to xenotransplantation. Xenotransplantation. 2013;20:27–35. doi: 10.1111/xen.12019. [DOI] [PubMed] [Google Scholar]
- 58.Li P, Estrada JL, Burlak C, et al. Efficient generation of genetically distinct pigs in a single pregnancy using multiplexed single-guide RNA and carbohydrate selection. Xenotransplantation. 2015;22:20–31. doi: 10.1111/xen.12131. [DOI] [PubMed] [Google Scholar]
- 59.Estrada JL, Martens G, Li P, et al. Evaluation of human and non-human primate antibody binding to pig cells lacking GGTA1/CMAH/beta4GalNT2 genes. Xenotransplantation. 2015;22:194–202. doi: 10.1111/xen.12161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Byrne GW, Du Z, Stalboerger P, et al. Cloning and expression of porcine beta1,4 N-acetylgalactosaminyl transferase encoding a new xenoreactive antigen. Xenotransplantation. 2014;21:543–554. doi: 10.1111/xen.12124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Ezzelarab MB, Ekser B, Azimzadeh A, et al. Systemic inflammation in xenograft recipients precedes activation of coagulation. Xenotransplantation. 2015;22:32–47. doi: 10.1111/xen.12133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Iwase H, Ekser B, Zhou H, et al. Further evidence for a sustained systemic inflammatory response in xenograft recipients (SIXR) 2015 doi: 10.1111/xen.12182. Submitted. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Oropeza M, Petersen B, Carnwath JW, et al. Transgenic expression of the human A20 gene in cloned pigs provides protection against apoptotic and inflammatory stimuli. Xenotransplantation. 2009;16:522–534. doi: 10.1111/j.1399-3089.2009.00556.x. [DOI] [PubMed] [Google Scholar]
- 64.Petersen B, Lucas-Hahn A, Lemme E, et al. Generation and characterization of pigs transgenic for human hemeoxygenase-1 (hHO-1) Xenotransplantation. 2010;17:102–103. [Google Scholar]
- 65.Petersen B, Ramackers W, Lucas-Hahn A, et al. Transgenic expression of human heme oxygenase-1 in pigs confers resistance against xenograft rejection during ex vivo perfusion of porcine kidneys. Xenotransplantation. 2011;18:355–368. doi: 10.1111/j.1399-3089.2011.00674.x. [DOI] [PubMed] [Google Scholar]
- 66.Chen G, Qian H, Starzl T, et al. Acute rejection is associated with antibodies to non-Gal antigens in baboons using Gal-knockout pig kidneys. Nat Med. 2005;11:1295–1298. doi: 10.1038/nm1330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.McGregor CG, Davies WR, Oi K, et al. Cardiac xenotransplantation: recent preclinical progress with 3-month median survival. J Thorac Cardiovasc Surg. 2005;130:844–851. doi: 10.1016/j.jtcvs.2005.04.017. [DOI] [PubMed] [Google Scholar]
- 68.Teotia SS, Walker RC, Schirmer JM, et al. Prevention, detection, and management of early bacterial and fungal infections in a preclinical cardiac xenotransplantation model that achieves prolonged survival. Xenotransplantation. 2005;12:127–133. doi: 10.1111/j.1399-3089.2005.00205.x. [DOI] [PubMed] [Google Scholar]
- 69.Buhler L, Awwad M, Basker M, et al. High-dose porcine hematopoietic cell transplantation combined with CD40 ligand blockade in baboons prevents an induced anti-pig humoral response. Transplantation. 2000;69:2296–2304. doi: 10.1097/00007890-200006150-00013. [DOI] [PubMed] [Google Scholar]
- 70.Iwase H, Ekser B, Satyananda V, et al. Pig-to-baboon heart transplantation - first experience with pigs transgenic for human thrombomodulin and comparison of three costimulation blockade-based regimens. Xenotransplantation. 2015;22:211–220. doi: 10.1111/xen.12167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Ezzelarab MB, Ekser B, Echeverri G, et al. Costimulation blockade in pig artery patch xenotransplantation - a simple model to monitor the adaptive immune response in nonhuman primates. Xenotransplantation. 2012;19:221–232. doi: 10.1111/j.1399-3089.2012.00711.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Iwase H, Satyananda V, Zhou H, et al. Initial in vivo experience of pig artery patch transplantation in baboons using mutant MHC (CIITA-DN) pigs. Transpl Immunol. 2015;32:99–108. doi: 10.1016/j.trim.2015.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Ezzelarab C, Ayares D, Cooper DK, et al. Human T-cell proliferation in response to thrombin-activated GTKO pig endothelial cells. Xenotransplantation. 2012;19:311–316. doi: 10.1111/j.1399-3089.2012.00718.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Satyananda V, Hara H, Ezzelarab MB, et al. New concepts of immune modulation in xenotransplantation. Transplantation. 2013;96:937–945. doi: 10.1097/TP.0b013e31829bbcb2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Wilhite T, Ezzelarab C, Hara H, et al. The effect of Gal expression on pig cells on the human T-cell xenoresponse. Xenotransplantation. 2012;19:56–63. doi: 10.1111/j.1399-3089.2011.00691.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Ezzelarab M, Ezzelarab C, Wilhite T, et al. Genetically-modified pig mesenchymal stromal cells: xenoantigenicity and effect on human T-cell xenoresponses. Xenotransplantation. 2011;18:183–195. doi: 10.1111/j.1399-3089.2011.00635.x. [DOI] [PubMed] [Google Scholar]
- 77.Ezzelarab MB, Ayares D, Cooper DKC. Transgenic expression of human CD46 reduces primate T cell responses to pig cells in vitro. Xenotransplantation. 2015 doi: 10.1111/xen.12209. (Submitted) [DOI] [PubMed] [Google Scholar]
- 78.Phelps CJ, Ball SF, Vaught TD, et al. Production and characterization of transgenic pigs expressing porcine CTLA4-Ig. Xenotransplantation. 2009;16:477–485. doi: 10.1111/j.1399-3089.2009.00533.x. [DOI] [PubMed] [Google Scholar]
- 79.Bottino R, Wijkstrom M, van der Windt DJ, et al. Pig-to-monkey islet xenotransplantation using multi-transgenic pigs. Am J Transplant. 2014;14:2275–2287. doi: 10.1111/ajt.12868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Wijkstrom M, Bottino R, Iwase H, et al. Glucose metabolism in pigs expressing human genes under an insulin promoter. Xenotransplantation. 2015;22:70–79. doi: 10.1111/xen.12145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Martin C, Plat M, Nerriere-Daguin V, et al. Transgenic expression of CTLA4-Ig by fetal pig neurons for xenotransplantation. Transgenic Res. 2005;14:373–384. doi: 10.1007/s11248-004-7268-4. [DOI] [PubMed] [Google Scholar]
- 82.Leveque X, Cozzi E, Naveilhan P, et al. Intracerebral xenotransplantation: recent findings and perspectives for local immunosuppression. Curr Opin Organ Transplant. 2011;16:190–194. doi: 10.1097/MOT.0b013e32834494b5. [DOI] [PubMed] [Google Scholar]
- 83.Phelps C, Dai Y, Ball S, et al. Production of transgenic pigs with down-regulation of SLA Class II on a GTKO/CD46 genetic background. 12th Congress of the International Xenotransplantation Association; Osaka, Japan. 2013; Abstract 262. [Google Scholar]
- 84.Hara H, Witt W, Crossley T, et al. Human dominant-negative class II transactivator transgenic pigs - effect on the human anti-pig T-cell immune response and immune status. Immunology. 2013;140:39–46. doi: 10.1111/imm.12107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Reyes LM, Estrada JL, Wang ZY, et al. Creating class I MHC-null pigs using guide RNA and the Cas9 endonuclease. J Immunol. 2014;193:5751–5757. doi: 10.4049/jimmunol.1402059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Tena A, Kurtz J, Leonard DA, et al. Transgenic expression of human CD47 markedly increases engraftment in a murine model of pig-to-human hematopoietic cell transplantation. Am J Transplant. 2014;14:2713–2722. doi: 10.1111/ajt.12918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Inverardi L, Clissi B, Stolzer AL, et al. Human natural killer lymphocytes directly recognize evolutionarily conserved oligosaccharide ligands expressed by xenogeneic tissues. Transplantation. 1997;63:1318–1330. doi: 10.1097/00007890-199705150-00021. [DOI] [PubMed] [Google Scholar]
- 88.Baumann BC, Forte P, Hawley RJ, et al. Lack of galactose-alpha-1,3-galactose expression on porcine endothelial cells prevents complement-induced lysis but not direct xenogeneic NK cytotoxicity. J Immunol. 2004;172:6460–6467. doi: 10.4049/jimmunol.172.10.6460. [DOI] [PubMed] [Google Scholar]
- 89.Rieben R, Seebach JD. Xenograft rejection: IgG1, complement and NK cells team up to activate and destroy the endothelium. Trends Immunol. 2005;26:2–5. doi: 10.1016/j.it.2004.11.011. [DOI] [PubMed] [Google Scholar]
- 90.Horvath-Arcidiacono JA, Porter CM, Bloom ET. Human NK cells can lyse porcine endothelial cells independent of their expression of Galalpha(1,3)-Gal and killing is enhanced by activation of either effector or target cells. Xenotransplantation. 2006;13:318–327. doi: 10.1111/j.1399-3089.2006.00316.x. [DOI] [PubMed] [Google Scholar]
- 91.Kennett SB, Porter CM, Horvath-Arcidiacono JA, et al. Characterization of baboon NK cells and their xenogeneic activity. Xenotransplantation. 2010;17:288–299. doi: 10.1111/j.1399-3089.2010.00591.x. [DOI] [PubMed] [Google Scholar]
- 92.Seebach JD, Comrack C, Germana S, et al. HLA-Cw3 expression on porcine endothelial cells protects against xenogeneic cytotoxicity mediated by a subset of human NK cells. J Immunol. 1997;159:3655–3661. [PubMed] [Google Scholar]
- 93.Dorling A, Monk NJ, Lechler RI. HLA-G inhibits the transendothelial migration of human NK cells. Eur J Immunol. 2000;30:586–593. doi: 10.1002/1521-4141(200002)30:2<586::AID-IMMU586>3.0.CO;2-Y. [DOI] [PubMed] [Google Scholar]
- 94.Forte P, Pazmany L, Matter-Reissmann UB, et al. HLA-G inhibits rolling adhesion of activated human NK cells on porcine endothelial cells. J Immunol. 2001;167:6002–6008. doi: 10.4049/jimmunol.167.10.6002. [DOI] [PubMed] [Google Scholar]
- 95.Crew MD. Play it in E or G: utilization of HLA-E and -G in xenotransplantation. Xenotransplantation. 2007;14:198–207. doi: 10.1111/j.1399-3089.2007.00395.x. [DOI] [PubMed] [Google Scholar]
- 96.Weiss EH, Lilienfeld BG, Muller S, et al. HLA-E/human beta2-microglobulin transgenic pigs: protection against xenogeneic human anti-pig natural killer cell cytotoxicity. Transplantation. 2009;87:35–43. doi: 10.1097/TP.0b013e318191c784. [DOI] [PubMed] [Google Scholar]
- 97.Maeda A, Kawamura T, Ueno T, et al. The suppression of inflammatory macrophage-mediated cytotoxicity and proinflammatory cytokine production by transgenic expression of HLA-E. Transpl Immunol. 2013;29:76–81. doi: 10.1016/j.trim.2013.08.001. [DOI] [PubMed] [Google Scholar]
- 98.Esquivel EL, Maeda A, Eguchi H, et al. Suppression of human macrophage-mediated cytotoxicity by transgenic swine endothelial cell expression of HLA-G. Transpl Immunol. 2015;32:109–115. doi: 10.1016/j.trim.2014.12.004. [DOI] [PubMed] [Google Scholar]
- 99.Forte P, Baumann BC, Weiss EH, et al. HLA-E expression on porcine cells: protection from human NK cytotoxicity depends on peptide loading. Am J Transplant. 2005;5:2085–2093. doi: 10.1111/j.1600-6143.2005.00987.x. [DOI] [PubMed] [Google Scholar]
- 100.Soin B, Smith KG, Zaidi A, et al. Physiological aspects of pig-to-primate renal xenotransplantation. Kidney Int. 2001;60:1592–1597. doi: 10.1046/j.1523-1755.2001.00973.x. [DOI] [PubMed] [Google Scholar]
- 101.Ibrahim Z, Busch J, Awwad M, et al. Selected physiologic compatibilities and incompatibilities between human and porcine organ systems. Xenotransplantation. 2006;13:488–499. doi: 10.1111/j.1399-3089.2006.00346.x. [DOI] [PubMed] [Google Scholar]
- 102.van der Windt DJ, Bottino R, Casu A, et al. Long-term controlled normoglycemia in diabetic non-human primates after transplantation with hCD46 transgenic porcine islets. Am J Transplant. 2009;9:2716–2726. doi: 10.1111/j.1600-6143.2009.02850.x. [DOI] [PubMed] [Google Scholar]
- 103.Ekser B, Long C, Echeverri GJ, et al. Impact of thrombocytopenia on survival of baboons with genetically modified pig liver transplants: clinical relevance. Am J Transplant. 2010;10:273–285. doi: 10.1111/j.1600-6143.2009.02945.x. [DOI] [PubMed] [Google Scholar]
- 104.Burlak C, Paris LL, Chihara RK, et al. The fate of human platelets perfused through the pig liver: implications for xenotransplantation. Xenotransplantation. 2010;17:350–361. doi: 10.1111/j.1399-3089.2010.00605.x. [DOI] [PubMed] [Google Scholar]
- 105.Ezzelarab M, Ekser B, Gridelli B, et al. Thrombocytopenia after pig-to-baboon liver xenotransplantation: where do platelets go? Xenotransplantation. 2011;18:320–327. doi: 10.1111/j.1399-3089.2011.00679.x. [DOI] [PubMed] [Google Scholar]
- 106.Burlak C, Paris LL, Lutz AJ, et al. Reduced binding of human antibodies to cells from GGTA1/CMAH KO pigs. Am J Transplant. 2014;14:1895–1900. doi: 10.1111/ajt.12744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Paris LL, Estrada JL, Li P, et al. Reduced human platelet uptake by pig livers deficient in the asialoglycoprotein receptor 1 protein. Xenotransplantation. 2015;22:203–210. doi: 10.1111/xen.12164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Ekser B, Echeverri GJ, Hassett AC, et al. Hepatic function after genetically engineered pig liver transplantation in baboons. Transplantation. 2010;90:483–493. doi: 10.1097/TP.0b013e3181e98d51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Onions D, Cooper DK, Alexander TJ, et al. An approach to the control of disease transmission in pig-to-human xenotransplantation. Xenotransplantation. 2000;7:143–155. doi: 10.1034/j.1399-3089.2000.00047.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Fishman JA, Patience C. Xenotransplantation: infectious risk revisited. Am J Transplant. 2004;4:1383–1390. doi: 10.1111/j.1600-6143.2004.00542.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Denner J, Tonjes RR. Infection barriers to successful xenotransplantation focusing on porcine endogenous retroviruses. Clin Microbiol Rev. 2012;25:318–343. doi: 10.1128/CMR.05011-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Smith DM. Endogenous retroviruses in xenografts. N Engl J Med. 1993;328:142–143. doi: 10.1056/NEJM199301143280218. [DOI] [PubMed] [Google Scholar]
- 113.Dieckhoff B, Karlas A, Hofmann A, et al. Inhibition of porcine endogenous retroviruses (PERVs) in primary porcine cells by RNA interference using lentiviral vectors. Arch Virol. 2007;152:629–634. doi: 10.1007/s00705-006-0868-y. [DOI] [PubMed] [Google Scholar]
- 114.Dieckhoff B, Petersen B, Kues WA, et al. Knockdown of porcine endogenous retrovirus (PERV) expression by PERV-specific shRNA in transgenic pigs. Xenotransplantation. 2008;15:36–45. doi: 10.1111/j.1399-3089.2008.00442.x. [DOI] [PubMed] [Google Scholar]
- 115.Ramsoondar J, Vaught T, Ball S, et al. Production of transgenic pigs that express porcine endogenous retrovirus small interfering RNAs. Xenotransplantation. 2009;16:164–180. doi: 10.1111/j.1399-3089.2009.00525.x. [DOI] [PubMed] [Google Scholar]
- 116.Hsu PD, Lander ES, Zhang F. Development and applications of CRISPR-Cas9 for genome engineering. Cell. 2014;157:1262–1278. doi: 10.1016/j.cell.2014.05.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Kim H, Kim JS. A guide to genome engineering with programmable nucleases. Nat Rev Genet. 2014;15:321–334. doi: 10.1038/nrg3686. [DOI] [PubMed] [Google Scholar]
- 118.Bibikova M, Golic M, Golic KG, et al. Targeted chromosomal cleavage and mutagenesis in Drosophila using zinc-finger nucleases. Genetics. 2002;161:1169–1175. doi: 10.1093/genetics/161.3.1169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Urnov FD, Rebar EJ, Holmes MC, et al. Genome editing with engineered zinc finger nucleases. Nat Rev Genet. 2010;11:636–646. doi: 10.1038/nrg2842. [DOI] [PubMed] [Google Scholar]
- 120.Hauschild J, Petersen B, Santiago Y, et al. Efficient generation of a biallelic knockout in pigs using zinc-finger nucleases. Proc Natl Acad Sci U S A. 2011;108:12013–12017. doi: 10.1073/pnas.1106422108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Tesson L, Usal C, Menoret S, et al. Knockout rats generated by embryo microinjection of TALENs. Nat Biotechnol. 2011;29:695–696. doi: 10.1038/nbt.1940. [DOI] [PubMed] [Google Scholar]
- 122.Galli C, Perota A, Brunetti D, et al. Genetic engineering including superseding microinjection: new ways to make GM pigs. Xenotransplantation. 2010;17:397–410. doi: 10.1111/j.1399-3089.2010.00590.x. [DOI] [PubMed] [Google Scholar]
- 123.Seruggia D, Montoliu L. The new CRISPR-Cas system: RNA-guided genome engineering to efficiently produce any desired genetic alteration in animals. Transgenic Res. 2014;23:707–716. doi: 10.1007/s11248-014-9823-y. [DOI] [PubMed] [Google Scholar]
- 124.Doudna JA, Charpentier E. Genome editing. The new frontier of genome engineering with CRISPR-Cas9. Science. 2014;346:1258096. doi: 10.1126/science.1258096. [DOI] [PubMed] [Google Scholar]
- 125.Ni W, Qiao J, Hu S, et al. Efficient gene knockout in goats using CRISPR/Cas9 system. PLoS One. 2014;9:e106718. doi: 10.1371/journal.pone.0106718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Mali P, Esvelt KM, Church GM. Cas9 as a versatile tool for engineering biology. Nat Methods. 2013;10:957–963. doi: 10.1038/nmeth.2649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Cooper DK, Ekser B, Burlak C, et al. Clinical lung xenotransplantation--what donor genetic modifications may be necessary? Xenotransplantation. 2012;19:144–158. doi: 10.1111/j.1399-3089.2012.00708.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Wynyard S, Nathu D, Garkavenko O, et al. Microbiological safety of the first clinical pig islet xenotransplantation trial in New Zealand. Xenotransplantation. 2014 May 7; doi: 10.1111/xen.12102. Epub ahead of print. [DOI] [PubMed] [Google Scholar]
- 129.Zhang MC, Liu X, Jin Y, et al. Lamellar keratoplasty treatment of fungal corneal ulcers with acellular porcine corneal stroma. Am J Transplant. 2015;15:1068–1075. doi: 10.1111/ajt.13096. [DOI] [PubMed] [Google Scholar]
- 130.Hara H, Ezzelarab M, Rood PP, et al. Allosensitized humans are at no greater risk of humoral rejection of GT-KO pig organs than other humans. Xenotransplantation. 2006;13:357–365. doi: 10.1111/j.1399-3089.2006.00319.x. [DOI] [PubMed] [Google Scholar]
- 131.Wong BS, Yamada K, Okumi M, et al. Allosensitization does not increase the risk of xenoreactivity to alpha1,3-galactosyltransferase gene-knockout miniature swine in patients on transplantation waiting lists. Transplantation. 2006;82:314–319. doi: 10.1097/01.tp.0000228907.12073.0b. [DOI] [PubMed] [Google Scholar]
- 132.Cooper DK, Tseng YL, Saidman SL. Alloantibody and xenoantibody cross- reactivity in transplantation. Transplantation. 2004;77:1–5. doi: 10.1097/01.TP.0000105116.74032.63. [DOI] [PubMed] [Google Scholar]
- 133.Ye Y, Luo Y, Kobayashi T, et al. Secondary organ allografting after a primary “bridging” xenotransplant. Transplantation. 1995;60:19–22. doi: 10.1097/00007890-199507150-00004. [DOI] [PubMed] [Google Scholar]
- 134.Baertschiger RM, Dor FJ, Prabharasuth D, et al. Absence of humoral and cellular alloreactivity in baboons sensitized to pig antigens. Xenotransplantation. 2004;11:27–32. doi: 10.1111/j.1399-3089.2004.00075.x. [DOI] [PubMed] [Google Scholar]
- 135.Cooper DKC, Bottino R. Recent advances in understanding xenotransplantation: implications for the clinic. Expert Rev Clin Immunol. 2015 doi: 10.1586/1744666X.2015.1083861. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]