

Abstract
The historic suggestion that Mycobacterium avium subsp. paratuberculosis (Map) might be a zoonotic pathogen was based on the apparent similarity of lesions in the intestine of patients with Crohn’s disease (CD) with those present in cattle infected with Map, the etiological agent of Johne’s disease. Reluctance to fully explore this possibility has been attributed to the difficulty in demonstrating the presence of Map in tissues from patients with CD. Advances in technology have resolved this problem and revealed the presence of Map in a significant proportion of patients with CD and other diseases. The seminal finding from recent investigations, however, is the detection of Map in healthy individuals with no clinical signs of disease. The latter observation indicates all humans are susceptible to infection with Map and lends support to the thesis that Map is zoonotic, with a latent stage of infection similar to tuberculosis, where infection leads to the development of an immune response that controls but does not eliminate the pathogen. This clarifies one of the reasons why it has been so difficult to document that Map is zoonotic and associated with the pathogenesis of CD and other diseases. As discussed in the present review, a better understanding of the immune response to Map is needed to determine how infection is usually kept under immune control during the latent stage of infection and elucidate the triggering events that lead to disease progression in the natural host and pathogenesis of CD and immune related diseases in humans.
Keywords: Crohn’s disease, Johne’s disease, Mycobacterium avium subsp. paratuberculosis, Animal model, Monoclonal antibodies, Flow cytometry, Cytokines
Core tip: The seminal finding from recent investigations is the detection of Mycobacterium avium subsp. paratuberculosis (Map) in healthy individuals with no clinical signs of disease. The latter observation indicates all humans are susceptible to infection with Mycobacterium avium subsp. paratuberculosis (Map) and lends support to the thesis that Map is zoonotic, with a latent stage of infection similar to M. tuberculosis, where infection leads to the development of an immune response that controls but does not eliminate the pathogen. This clarifies one of the reasons why it has been so difficult to document that Map is zoonotic and associated with the pathogenesis of Crohn’s disease and other diseases.
INTRODUCTION
A controversy has persisted since the initial suggestion that Map may be a zoonotic pathogen and the causative agent of Crohn’s disease (CD). Map is a mycobacterial pathogen with a broad host range that can infect many species[1]. The primary hosts are ruminants. It was initially diagnosed as the causative agent of Johne’s disease (JD), a chronic enteritis in cattle[2]. The similarities in the clinical manifestations of JD with CD led Dalziel et al[3] and later Crohn et al[4] to suggest Map may be the causative agent of CD[1]. As reviewed comprehensively by Naser et al[5] and Gitlin et al[6], the debate continues. The reluctance to fully explore the issue of whether Map is associated with the pathogenesis of CD has been the difficulty in isolating Map from all patients with CD. Chiodini et al[7-9] were the first investigators to successfully isolate Map from 3 patients. Initial attempts by others, however, proved unsuccessful raising questions as to the role of Map in the pathogenesis of CD. Even with improvements in technology, success in detection of Map has varied with some investigators more successful than others in isolating Map from patients with CD. Sufficient evidence, however, has been obtained over recent years to clearly show that many patients with CD are infected with Map. Investigations have revealed the presence of Map or Map DNA in blood or lesions from adults and children with CD[10-16]. Of special interest, Map has also been found in patients with other diseases as well as healthy subjects using several methods of detection[17-20]. A study long overdue, has now been conducted by Singh et al[21,22]. The study was conducted in India. It was designed to examine the level of infection in humans where Map is endemic in livestock and the general environment. They obtained specimens of blood, serum and stool from 42400 patients, submitted to pathology laboratories and hospitals in different areas of India, where Map is endemic, for evaluation over a span of two plus years. The samples were from patients presenting with a spectrum of infectious and non-infectious diseases including typhoid, TB, malaria, skin disorders, type 1 diabetes, anemia, thyroid, ion imbalance, abdominal disorders, lipid profile disorders, and intestinal inflammatory illness of different origins. They also collected samples from healthy individuals. Map was detected in all categories of patients at different frequencies, including healthy individuals, reflecting the consequence of large scale exposure of the resident populations to Map in the environment, infected animals and the food supply. The predominant biotype of Map detected in the samples was the bison type prevalent in livestock. The results clearly show, humans like other species, are equally susceptible to infection with Map regardless of health status. It can be predicted that any further large scale screening will provide similar results with the frequency of infected individuals dependent on the prevalence of Map in the environment and presence in food supplies[23]. The question remaining and reason for the unending debate is, “is Map a zoonotic pathogen and if so, what role does it play in the pathogenesis of CD and other diseases?” The answer is yes for at least a subset of patients presenting with the clinical signs of CD[5,24]. Koch’s postulates were actually fulfilled in subsequent studies by Chiodini el al[25] where he and his associates demonstrated that passage of one of the first human isolates of Map in kid goats led to the development of enteritis similar to the enteritis in CD and JD. A later investigation of an enteritis in a colony of stumptail macaque (Macaca arctoides) revealed Map was the causative agent, the first report showing the susceptibility of non-human primates to infection with Map[26]. The study was the first to show that Map could be isolated from feces as an acid fast bacillus, extending what was known about the host range of species susceptible to infection with Map. The ensuing studies have shown Map isolated from patients with CD may be present in affected tissues with or without the mycobacterial cell wall[9], a characteristic of other mycobacterial pathogens[27]. Studies of healthy individuals and patients with clinical signs of CD and other diseases have shown Map, with an intact cell wall, can be detected in blood, tissues, and feces[22]. Cell wall deficient forms have also been found in tissues from animals with JD[28].
ROLE OF Map IN CROHN’S AND OTHER DISEASES
The difficulty in elucidating the role of Map in CD and other diseases has been the lack of understanding of the mechanisms of pathogenesis of JD. Map, like other pathogenic mycobacteria doesn’t cause epidemic disease, where all infected animals develop clinical disease. This is similar to tuberculosis in humans where infection leads to the development of an immune response that controls but does not clear infection. Only an estimated 10% of individuals infected with Mtb develop clinical disease. Disease progression is associated with a breakdown in protective immunity. Intensive ongoing investigations have not yet identified the triggering events associated with disruption of protective immunity during the latent stage of infection. No comprehensive studies have been conducted on the immune response to Map in humans. However, the presence of an immune response to Map in patients with clinical manifestations of CD and the presence of antibody to Map in healthy individuals and patients with various diseases show exposure leads to development of an immune response to Map[22,29,30]. The detection of Map in blood and affected tissues shows that infection persists providing a constant stimulus to the immune system.
Map as a model pathogen to study the mechanism of immunopathogenesis of mycobacterial pathogens
There are two ways Map could contribute to the pathogenesis of diseases in humans. The first is as the etiologic agent of CD, where there is a breakdown of immunity present during the latent stage of infection, similar to the breakdown that occurs with tuberculosis[5,24,31]. The second is through mimicry where mimics of proteins present in humans are produced by Map that interfere with immune function and or elicit development of cross-reactive antibodies associated with autoimmune diseases[32,33].
For ethical reasons much of the information needed to elucidate the role of Map in pathogenesis of CD and other diseases can’t be obtained through investigations conducted with human subjects. The alternative is the use of cattle, the natural host of Map, as a model species to study the role of Map in the immunopathogenesis of CD and other diseases. The added advantage is that it affords an opportunity address the broader problem, the need to elucidate the mechanisms of immunopathogenesis of mycobacterial pathogens in general. The similarities in latency suggest the pathogens use the same signaling pathways to dysregulate the immune response and establish a persistent infection that eventually, over time, leads to a complete breakdown of protective immunity and development of clinical disease. The unique advantage of using Map to explore this possibility is that it can be used to study the interaction of the pathogen with the host at all stages of infection. Our studies have been based on the premise that Map is a zoonotic pathogen that is the etiologic agent of a subset of patients with CD[5,24,31] and that it plays a role in pathogenesis of other disease through mimicry[22,32].
To provide some background and a review of the current status of knowledge on the mechanisms of immunopathogenesis of JD in cattle, we became interested in the potential role of Map in CD when limited information was available on the immune response to Map in cattle. Progress was limited by the lack of immune reagents needed to characterize the immune system in cattle and study the cell mediated immune response to Map. Studies in dairy herds had shown the disease is characterized by a long latency period, where infected animals were difficult to detect by available diagnostic techniques during the initial stages of infection. Some animals could be detected by development of cell mediated immunity using the tuberculin skin test (reviewed in[1]). At later stages of infection, infected animals could be detected by intermittent shedding of Map in feces. Persistent shedding was associated with development of clinical disease where the absorptive mucosa of the intestine is destroyed, leading to death by starvation. The long latency suggested that Map could be passed through the gastrointestinal system without infecting all animals and constant exposure was needed for infection to occur. The latency also suggested that there was an age related difference in susceptibility to infection, with older animals more resistant to infection than young animals. Development of immune reagents to characterize the immune system of cattle and a flow cytometric assay provided an opportunity to begin analyzing the immune response to Map ex vivo and determine whether there was an age difference in susceptibility to infection and whether it was possible for “pass through” of Map without infection[34-36]. Studies in neonatal calves revealed exposure through application of Map to tonsillar crypts[36] or by ingestion of Map in milk supplement, leads to uniform infection in all animals and development of a lymphoproliferative response ex vivo to Map purified protein derivative (PPD) and soluble Map antigens (Ag) dominated by memory CD45R0+ CD4 T cells 5 mo post infection (PI)[34]. Peripheral colonization of tissue was detected in some tissues at necropsy, with some bacteria detectable in feces in the initial study[36]. In the second study, no gross or microscopic lesions were detectable in intestinal tissue at necropsy one year PI, with Map detectable in some tissues by PCR[34]. A surgical intervention model used to directly inoculate the ileum showed Map is taken up rapidly and disseminated to other tissues within an hour PI. A proliferative response was detected by 9 mo but no humoral response[37]. Development of a cannulated ileum model allowed us to continuously monitor the early stages of infection in experimentally infected calves for up to 18 mo PI (Figure 1)[35]. Use of the model showed bacteria directly introduced into the ileum, once or multiple times over several days, were rapidly cleared with detection of bacteria in feces only during the first weeks PI. No gross or microscopic lesions were found in biopsies during the 18 mo of the studies. However, Map was detected by PCR with IS900 and culture of some tissues. An immune response was detected using a flow cytometric assay at 3 mo[35]. A subsequent study, with the cannulation model, was focused on determining whether passage of Map in humans altered their capacity to infect the natural host and establish a persistent infection. Two of the original isolates obtained from humans by Chiodini et al[7] (Linda and Ben) were used to compare the capacity of isolates from humans and cattle to infect calves. The comparison revealed no difference in infectivity or the immune response to the bovine or human isolates of Map[38]. Sequence analysis of the human isolate obtained from tissue had identical single sequence repeat as the original isolate, 7g-4ggt, a sequence common in isolates from cattle[39]. This observation indicates it will not be necessary to use human isolates of Map to model mechanisms of persistence and pathogenesis in cattle.
Figure 1.
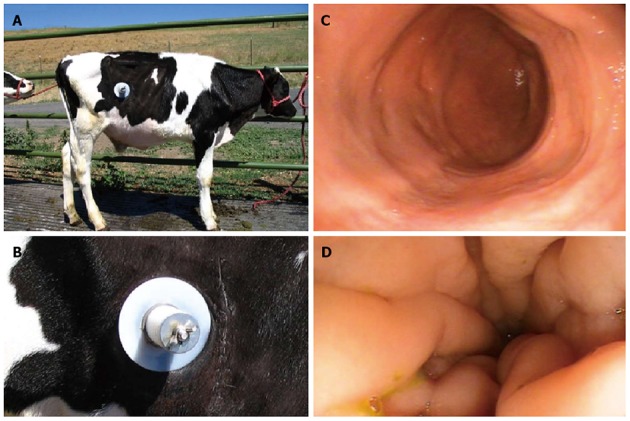
Early stages of infection in experimentally infected calves for up to 18 mo post infection. Pictures showing the cannula used in the studies (A and B); Endoscopic field showing the condition of ileal mucosa 8 mo following infection with Map (C); Endoscopic field taken before necropsy of a naturally infected animal showing the characteristic swelling and corrugation of the ileal mucosa that occurs at the clinical stage of infection (D). Modified from[35,38]. Map: Mycobacterium avium subsp. paratuberculosis.
MECHANISMS OF IMMUNE EVASION AND IMMUNOPATHOGENESIS
The early events of infection by pathogenic mycobacteria that lead to the development of an immune response that controls but fails to eliminate the pathogen remain to be elucidated. Also, the triggering events that disrupt protective immunity have not been identified. Efforts to breakdown the immune response to Map in cattle and accelerate progression to clinical disease have revealed the immune response to Map is resilient. Exposure of calves to massive doses of Map by different routes or administration of dexamethasone to alter the immune response to Map did not modulate the immune response to Map[40]. In addition, transient depletion of CD4 T cells in calves with a monoclonal antibody to CD4 did not impair the development of an immune response to Map or accelerate progression to clinical disease[41]. Under all conditions tested, including exposure at birth, an immune response develops that controls but does not clear infection. These observations suggest a similar resilient response also occurs in the majority of humans exposed to Map that do not develop clinical disease. The challenge that remains is identifying the factors that trigger a breakdown in protective immunity.
Thus far, analysis of the immune response to Map has shown the cellular response, under experimental conditions, is characterized by development of an immune response dominated by CD45R0+ memory CD4 T cells using live Map, Map PPD, and soluble Map Ags[34,35,38]. Although a CD8 T cell response also develops, the proliferative response to Map ex vivo is less than the proliferative response of CD4 T cells. The response increases over time, however. In a comparative study, the CD8 T cell proliferative response was equivalent to the response observed with naturally infected animals at the clinical stage of infection[38]. qRT-PCR showed interferon (IFN)-γ, interleukin (IL)-17, and IL-22 gene expression was up-regulated to equivalent extent in experimentally and naturally infected animals[38].
Ongoing efforts to develop a vaccine have provided additional information on the immune response to Map and an approach for elucidating how Map and mycobacterial pathogens dysregulate the immune response to establish a persistent infection. We used site directed mutation to develop 3 mutants to genes associated with virulence (relA, lsr2, and pknG)[42,43] as part of a multi-institutional endeavor to identify deletion mutants as vaccine candidates[44]. Mutants, made by other investigators, selected for challenge studies in mice and goats based on reduced capacity to survive in macrophages were unsuccessful. All the mutants were able to establish an infection[44]. In contrast, one of the mutants we selected for a challenge study in goats and calves, ΔMap-relA, was unable to establish a persistent infection in goats and calves[45]. Further analysis in calves showed no clear difference in the proliferative response to Map antigens. The increases in expression IFN-γ, IL-17, and IL22 were comparable indicating more in depth studies are needed to determine how deletion of relA abrogates the capacity of Map to establish a persistent infection.
The last observation of interest is on the effect of therapeutic vaccination with BCG on the immune response to Mtb in guinea pigs and humans at the clinical stage of infection made by Koch[46]. Vaccination led to a reversal of pathology, showing the triggering events associated with a breakdown in protective immunity, could be reversed by vaccination. Although subsequent studies revealed vaccination only led to a transient reversal, the observation is of interest because studies of vaccination at the clinical stage of infection could reveal which components of the immune response are restored that account for a reversal of pathology. Similar observations have been made more recently by Singh et al[47,48] with Map in goats and cattle. Although the results looked very promising, reversal of pathology was transient with no improvement even with repeated vaccination (personal communication). The important observation made by Koch et al[46] and Singh et al[47,48] is that reversal of pathology was associated with an increase in the inflammatory response following vaccination that resolved lesions containing infected cells. Map affords an opportunity to explore the cellular and molecular basis for this phenomenon. A preliminary test with a cow at a late clinical stage of infection, with an anergic response to a skin test with Map Ag and a diminished proliferative response to Ag ex vivo, showed vaccination with ΔMaprelA restored the proliferative response to Map. With the cannulation model it will be possible to isolate affected tissue before and after vaccination for analysis and characterize the effector cells restored by vaccination and characterize their effector activity.
CONCLUSION
Cumulative studies clearly show humans, like other species, are equally susceptible to infection with Map[22]. This places the whole populations at risk of infection, depending on the prevalence of Map in the environment and food supply[22,23]. Ongoing studies have shown CD may contribute to the pathogenesis of disease in humans as the etiologic agent of a subset of patients with CD[5] and through antigenic mimicry[32]. Use of the natural host as a model, to understand the immunopathogenesis of CD mediated by Map, has provided opportunity to gain insight into the immune response to Map in humans during the early, latent, and clinical stages of infection that cannot be obtained through study of Map in humans, except at the clinical stage of infection[29]. Analysis has shown the immune response during the latent stage of infection is resilient and not readily disrupted by methods of immunosuppression[40,41]. The components of the immune response to Map that control infection during latency remain to be elucidated. The development of a mutant that cannot establish a persistent infection offers 2 opportunities to identify these components, one through analysis of the early stages of infection and the second through analysis of the immune response following restoration by vaccination at the clinical stage of infection.
Footnotes
Supported by Some of the studies cited here were funded in part with funds from the National Institute of Allergy and Infectious Diseases, National Institutes of Health, Department of Health and Human Services, under Contract No. N01-AI-30055, the JDIP program USDA-CSREES-NRI-CAP Award No. 2007-01019 awarded to Davis WC, USDA-NRI grant No. 2005-35204-16106 and the Washington State University Monoclonal Antibody Center.
Conflict-of-interest statement: The author declares no conflict of interest.
Open-Access: This article is an open-access article which was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Peer-review started: August 11, 2015
First decision: October 14, 2015
Article in press: November 19, 2015
P- Reviewer: Gazouli M, Sakuraba A S- Editor: Qi Y L- Editor: A E- Editor: Zhang DN
References
- 1.Rideout BA, Brown ST, Davis WC, Gay JM, Giannella RA, Hines ME, Hueston WD, Hutchinson LJ. Diagnosis and Control of Johne’s Disease. Washington, DC: The National Academy Press; 2003. pp. 1–229. [Google Scholar]
- 2.Johne HA, Frothingham L. Ein eigenthumlicher fall von tuberculosis beim rind [A peculiar case of tuberculosis in a cow] Deutsche Zeitschr Tierm Path. 1895;21:438–454. [Google Scholar]
- 3.Dalziel TK. Thomas Kennedy Dalziel 1861-1924. Chronic interstitial enteritis. Dis Colon Rectum. 1989;32:1076–1078. doi: 10.1007/BF02553886. [DOI] [PubMed] [Google Scholar]
- 4.Crohn BB, Ginzburg L, Oppenheimer GD. Regional ileitis; a pathologic and clinical entity. Am J Med. 1952;13:583–590. doi: 10.1016/0002-9343(52)90025-9. [DOI] [PubMed] [Google Scholar]
- 5.Naser SA, Sagramsingh SR, Naser AS, Thanigachalam S. Mycobacterium avium subspecies paratuberculosis causes Crohn’s disease in some inflammatory bowel disease patients. World J Gastroenterol. 2014;20:7403–7415. doi: 10.3748/wjg.v20.i23.7403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gitlin L, Borody TJ, Chamberlin W, Campbell J. Mycobacterium avium ss paratuberculosis-associated diseases: piecing the Crohn’s puzzle together. J Clin Gastroenterol. 2012;46:649–655. doi: 10.1097/MCG.0b013e31825f2bce. [DOI] [PubMed] [Google Scholar]
- 7.Chiodini RJ, Van Kruiningen HJ, Merkal RS, Thayer WR, Coutu JA. Characteristics of an unclassified Mycobacterium species isolated from patients with Crohn’s disease. J Clin Microbiol. 1984;20:966–971. doi: 10.1128/jcm.20.5.966-971.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Chiodini RJ, Van Kruiningen HJ, Thayer WR, Merkal RS, Coutu JA. Possible role of mycobacteria in inflammatory bowel disease. I. An unclassified Mycobacterium species isolated from patients with Crohn’s disease. Dig Dis Sci. 1984;29:1073–1079. doi: 10.1007/BF01317078. [DOI] [PubMed] [Google Scholar]
- 9.Chiodini RJ, Van Kruiningen HJ, Thayer WR, Coutu JA. Spheroplastic phase of mycobacteria isolated from patients with Crohn’s disease. J Clin Microbiol. 1986;24:357–363. doi: 10.1128/jcm.24.3.357-363.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Naser SA, Collins MT, Crawford JT, Valentine JF. Culture of Mycobacterium avium subspecies paratuberculosis (MAP) from the blood of patients with Crohn’s disease: a follow-up blind multi center investigation. The Open Inflamm J. 2009;2:22–23. [Google Scholar]
- 11.Autschbach F, Eisold S, Hinz U, Zinser S, Linnebacher M, Giese T, Löffler T, Büchler MW, Schmidt J. High prevalence of Mycobacterium avium subspecies paratuberculosis IS900 DNA in gut tissues from individuals with Crohn’s disease. Gut. 2005;54:944–949. doi: 10.1136/gut.2004.045526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bull TJ, McMinn EJ, Sidi-Boumedine K, Skull A, Durkin D, Neild P, Rhodes G, Pickup R, Hermon-Taylor J. Detection and verification of Mycobacterium avium subsp. paratuberculosis in fresh ileocolonic mucosal biopsy specimens from individuals with and without Crohn’s disease. J Clin Microbiol. 2003;41:2915–2923. doi: 10.1128/JCM.41.7.2915-2923.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kirkwood CD, Wagner J, Boniface K, Vaughan J, Michalski WP, Catto-Smith AG, Cameron DJ, Bishop RF. Mycobacterium avium subspecies paratuberculosis in children with early-onset Crohn’s disease. Inflamm Bowel Dis. 2009;15:1643–1655. doi: 10.1002/ibd.20967. [DOI] [PubMed] [Google Scholar]
- 14.Lee A, Griffiths TA, Parab RS, King RK, Dubinsky MC, Urbanski SJ, Wrobel I, Rioux KP. Association of Mycobacterium avium subspecies paratuberculosis with Crohn Disease in pediatric patients. J Pediatr Gastroenterol Nutr. 2011;52:170–174. doi: 10.1097/MPG.0b013e3181ef37ba. [DOI] [PubMed] [Google Scholar]
- 15.Mendoza JL, San-Pedro A, Culebras E, Cíes R, Taxonera C, Lana R, Urcelay E, de la Torre F, Picazo JJ, Díaz-Rubio M. High prevalence of viable Mycobacterium avium subspecies paratuberculosis in Crohn’s disease. World J Gastroenterol. 2010;16:4558–4563. doi: 10.3748/wjg.v16.i36.4558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chiappini E, de Martino M, Mangiantini F, Lionetti P. Crohn disease and mycobacterial infection in children: an intriguing relationship. J Pediatr Gastroenterol Nutr. 2009;49:550–558. doi: 10.1097/MPG.0b013e3181b0f908. [DOI] [PubMed] [Google Scholar]
- 17.Juste RA, Elguezabal N, Pavón A, Garrido JM, Geijo M, Sevilla I, Cabriada JL, Tejada A, García-Campos F, Casado R, et al. Association between Mycobacterium avium subsp. paratuberculosis DNA in blood and cellular and humoral immune response in inflammatory bowel disease patients and controls. Int J Infect Dis. 2009;13:247–254. doi: 10.1016/j.ijid.2008.06.034. [DOI] [PubMed] [Google Scholar]
- 18.Tuci A, Tonon F, Castellani L, Sartini A, Roda G, Marocchi M, Caponi A, Munarini A, Rosati G, Ugolini G, et al. Fecal detection of Mycobacterium avium paratuberculosis using the IS900 DNA sequence in Crohn’s disease and ulcerative colitis patients and healthy subjects. Dig Dis Sci. 2011;56:2957–2962. doi: 10.1007/s10620-011-1699-6. [DOI] [PubMed] [Google Scholar]
- 19.Singh AV, Singh SV, Makharia GK, Singh PK, Sohal JS. Presence and characterization of Mycobacterium avium subspecies paratuberculosis from clinical and suspected cases of Crohn’s disease and in the healthy human population in India. Int J Infect Dis. 2008;12:190–197. doi: 10.1016/j.ijid.2007.06.008. [DOI] [PubMed] [Google Scholar]
- 20.Singh AV, Singh SV, Singh PK, Sohal JS, Singh MK. High prevalence of Mycobacterium avium subspecies paratuberculosis (‘Indian bison type’) in animal attendants suffering from gastrointestinal complaints who work with goat herds endemic for Johne’s disease in India. Int J Infect Dis. 2011;15:e677–e683. doi: 10.1016/j.ijid.2011.04.013. [DOI] [PubMed] [Google Scholar]
- 21.Singh SV, Singh PK, Singh AV, Sohal JS, Kumar N, Chaubey KK, Gupta S, Rawat KD, Kumar A, Bhatia AK, et al. ‘Bio-load’ and bio-type profiles of Mycobacterium avium subspecies paratuberculosis infection in the domestic livestock population endemic for Johne’s disease: a survey of 28 years (1985-2013) in India. Transbound Emerg Dis. 2014;61 Suppl 1:43–55. doi: 10.1111/tbed.12216. [DOI] [PubMed] [Google Scholar]
- 22.Singh SV, Kumar N, Sohal JS, Singh AV, Singh PK, Agrawal ND, Gupta S, Chaubey KK, Deb R, Dhama R, et al. First mass screening of the human population to estimate the bio-load of Mycobacterium avium sub-species paratuberculosis in North India. JPHE. 2014;6:20–29. [Google Scholar]
- 23.Rhodes G, Richardson H, Hermon-Taylor J, Weightman A, Higham A, Pickup R. Mycobacterium avium Subspecies paratuberculosis: Human Exposure through Environmental and Domestic Aerosols. Pathogens. 2014;3:577–595. doi: 10.3390/pathogens3030577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Chiodini RJ, Dowd SE, Davis B, Galandiuk S, Chamberlin WM, Kuenstner JT, McCallum RW, Zhang J. Crohn’s disease may be differentiated into 2 distinct biotypes based on the detection of bacterial genomic sequences and virulence genes within submucosal tissues. J Clin Gastroenterol. 2013;47:612–620. doi: 10.1097/MCG.0b013e31827b4f94. [DOI] [PubMed] [Google Scholar]
- 25.Van Kruiningen HJ, Chiodini RJ, Thayer WR, Coutu JA, Merkal RS, Runnels PL. Experimental disease in infant goats induced by a Mycobacterium isolated from a patient with Crohn’s disease. A preliminary report. Dig Dis Sci. 1986;31:1351–1360. doi: 10.1007/BF01299814. [DOI] [PubMed] [Google Scholar]
- 26.McClure HM, Chiodini RJ, Anderson DC, Swenson RB, Thayer WR, Coutu JA. Mycobacterium paratuberculosis infection in a colony of stumptail macaques (Macaca arctoides) J Infect Dis. 1987;155:1011–1019. doi: 10.1093/infdis/155.5.1011. [DOI] [PubMed] [Google Scholar]
- 27.Beran V, Havelkova M, Kaustova J, Dvorska L, Pavlik I. Celll wall defficient forms of mycobacteria: a review. Vet Med. 2006;51:365–389. [Google Scholar]
- 28.Hulten K, Karttunen TJ, El-Zimaity HM, Naser SA, Collins MT, Graham DY, El-Zaatari FA. Identification of cell wall deficient forms of M. avium subsp. paratuberculosis in paraffin embedded tissues from animals with Johne’s disease by in situ hybridization. J Microbiol Methods. 2000;42:185–195. doi: 10.1016/s0167-7012(00)00185-8. [DOI] [PubMed] [Google Scholar]
- 29.Olsen I, Tollefsen S, Aagaard C, Reitan LJ, Bannantine JP, Andersen P, Sollid LM, Lundin KE. Isolation of Mycobacterium avium subspecies paratuberculosis reactive CD4 T cells from intestinal biopsies of Crohn’s disease patients. PLoS One. 2009;4:e5641. doi: 10.1371/journal.pone.0005641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Olsen I, Wiker HG, Johnson E, Langeggen H, Reitan LJ. Elevated antibody responses in patients with Crohn’s disease against a 14-kDa secreted protein purified from Mycobacterium avium subsp. paratuberculosis. Scand J Immunol. 2001;53:198–203. doi: 10.1046/j.1365-3083.2001.00857.x. [DOI] [PubMed] [Google Scholar]
- 31.Davis WC, Madsen-Bouterse SA. Crohn’s disease and Mycobacterium avium subsp. paratuberculosis: the need for a study is long overdue. Vet Immunol Immunopathol. 2012;145:1–6. doi: 10.1016/j.vetimm.2011.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Sechi LA, Dow CT. Mycobacterium avium ss. paratuberculosis Zoonosis - The Hundred Year War - Beyond Crohn’s Disease. Front Immunol. 2015;6:96. doi: 10.3389/fimmu.2015.00096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Sechi LA, Gazouli M, Ikonomopoulos J, Lukas JC, Scanu AM, Ahmed N, Fadda G, Zanetti S. Mycobacterium avium subsp. paratuberculosis, genetic susceptibility to Crohn’s disease, and Sardinians: the way ahead. J Clin Microbiol. 2005;43:5275–5277. doi: 10.1128/JCM.43.10.5275-5277.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Koo HC, Park YH, Hamilton MJ, Barrington GM, Davies CJ, Kim JB, Dahl JL, Waters WR, Davis WC. Analysis of the immune response to Mycobacterium avium subsp. paratuberculosis in experimentally infected calves. Infect Immun. 2004;72:6870–6883. doi: 10.1128/IAI.72.12.6870-6883.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Allen AJ, Park KT, Barrington GM, Lahmers KK, Hamilton MJ, Davis WC. Development of a bovine ileal cannulation model to study the immune response and mechanisms of pathogenesis of paratuberculosis. Clin Vaccine Immunol. 2009;16:453–463. doi: 10.1128/CVI.00347-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Waters WR, Miller JM, Palmer MV, Stabel JR, Jones DE, Koistinen KA, Steadham EM, Hamilton MJ, Davis WC, Bannantine JP. Early induction of humoral and cellular immune responses during experimental Mycobacterium avium subsp. paratuberculosis infection of calves. Infect Immun. 2003;71:5130–5138. doi: 10.1128/IAI.71.9.5130-5138.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wu CW, Livesey M, Schmoller SK, Manning EJ, Steinberg H, Davis WC, Hamilton MJ, Talaat AM. Invasion and persistence of Mycobacterium avium subsp. paratuberculosis during early stages of Johne’s disease in calves. Infect Immun. 2007;75:2110–2119. doi: 10.1128/IAI.01739-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Allen AJ, Park KT, Barrington GM, Lahmers KK, Abdellrazeq GS, Rihan HM, Sreevatsan S, Davies C, Hamilton MJ, Davis WC. Experimental infection of a bovine model with human isolates of Mycobacterium avium subsp. paratuberculosis. Vet Immunol Immunopathol. 2011;141:258–266. doi: 10.1016/j.vetimm.2011.03.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Amonsin A, Li LL, Zhang Q, Bannantine JP, Motiwala AS, Sreevatsan S, Kapur V. Multilocus short sequence repeat sequencing approach for differentiating among Mycobacterium avium subsp. paratuberculosis strains. J Clin Microbiol. 2004;42:1694–1702. doi: 10.1128/JCM.42.4.1694-1702.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Stabel JR, Palmer MV, Harris B, Plattner B, Hostetter J, Robbe-Austerman S. Pathogenesis of Mycobacterium avium subsp. paratuberculosis in neonatal calves after oral or intraperitoneal experimental infection. Vet Microbiol. 2009;136:306–313. doi: 10.1016/j.vetmic.2008.11.025. [DOI] [PubMed] [Google Scholar]
- 41.Allen AJ, Stabel JR, Robbe-Austerman S, Park KT, Palmer MV, Barrington GM, Lahmers KK, Hamilton MJ, Davis WC. Depletion of CD4 T lymphocytes at the time of infection with M. avium subsp. paratuberculosis does not accelerate disease progression. Vet Immunol Immunopathol. 2012;149:286–291. doi: 10.1016/j.vetimm.2012.07.010. [DOI] [PubMed] [Google Scholar]
- 42.Park KT, Dahl JL, Bannantine JP, Barletta RG, Ahn J, Allen AJ, Hamilton MJ, Davis WC. Demonstration of allelic exchange in the slow-growing bacterium Mycobacterium avium subsp. paratuberculosis, and generation of mutants with deletions at the pknG, relA, and lsr2 loci. Appl Environ Microbiol. 2008;74:1687–1695. doi: 10.1128/AEM.01208-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Park KT, Allen AJ, Bannantine JP, Seo KS, Hamilton MJ, Abdellrazeq GS, Rihan HM, Grimm A, Davis WC. Evaluation of two mutants of Mycobacterium avium subsp. paratuberculosis as candidates for a live attenuated vaccine for Johne’s disease. Vaccine. 2011;29:4709–4719. doi: 10.1016/j.vaccine.2011.04.090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Bannantine JP, Hines ME, Bermudez LE, Talaat AM, Sreevatsan S, Stabel JR, Chang YF, Coussens PM, Barletta RG, Davis WC, et al. A rational framework for evaluating the next generation of vaccines against Mycobacterium avium subspecies paratuberculosis. Front Cell Infect Microbiol. 2014;4:126. doi: 10.3389/fcimb.2014.00126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Park KT, Allen AJ, Barrington GM, Davis WC. Deletion of relA abrogates the capacity of Mycobacterium avium paratuberculosis to establish an infection in calves. Front Cell Infect Microbiol. 2014;4:64. doi: 10.3389/fcimb.2014.00064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Koch R. Die Aetiologie der tuberkulose. Mittheilungen ausdem Kaiserlichen Gesundheitsamte. 1884;2:1–88. [Google Scholar]
- 47.Singh K, Chandel BS, Chauhan HC, Dadawala A, Singh SV, Singh PK. Efficacy of ‘indigenous vaccine’ using native ‘Indian bison type’ genotype of Mycobacterium avium subspecies paratuberculosis for the control of clinical Johne’s disease in an organized goat herd. Vet Res Commun. 2013;37:109–114. doi: 10.1007/s11259-013-9551-4. [DOI] [PubMed] [Google Scholar]
- 48.Singh SV, Singh PK, Singh AV, Sohal JS, Sharma MC. Therapeutic Effects of a New “Indigenous Vaccine” Developed Using Novel Native “Indian Bison Type” Genotype of Mycobacterium avium Subspecies paratuberculosis for the Control of Clinical Johne’s Disease in Naturally Infected Goatherds in India. Vet Med Int. 2010;2010:351846. doi: 10.4061/2010/351846. [DOI] [PMC free article] [PubMed] [Google Scholar]