

Abstract
Background
Natural food products have been used for combating human diseases for thousands of years. Naturally occurring flavonoids including flavones, flavonols, flavanones, flavonols, isoflavones and anthocyanidins have been proposed as effective supplements for management and prevention of diabetes and its long-term complications based on in vitro and animal models.
Aim
To summarize the roles of dietary flavonoids in diabetes management and their molecular mechanisms.
Findings
Tremendous studies have found that flavonoids originated from foods could improve glucose metabolism, lipid profile, regulating the hormones and enzymes in human body, further protecting human being from diseases like obesity, diabetes and their complications.
Conclusion
In the current review, we summarize recent progress in understanding the biological action, mechanism and therapeutic potential of the dietary flavonoids and its subsequent clinical outcomes in the field of drug discovery in management of diabetes mellitus.
Keywords: Dietary flavonoids, Diabetes mellitus, Glycemic control, Insulin secretion, Hyperglycemia
Background
Type 2 diabetes mellitus (DM) is characterized by impaired insulin secretion, and increased insulin resistance (or resistance to insulin mediated glucose disposal). DM is possibly one of the world’s fast growing diseases [1] and disabling micro and macrovascular complications [2]. It has been estimated that nearly 592 million adults become diabetic patients by the year 2035 due to aging, high population growth size, increased urbanization, high prevalence of obesity, rise in living standards and the spread of calorie rich, fatty and fast foods [3]. Although many drugs are commercially available for treating the diseases, many of them are out of reach for a significant proportion of the population and are beset with some adverse effects [4]. The use of medicinal plants and their phytochemicals for treating diabetes is not just a search for safer alternatives to pharmaceuticals, which transiently lower the blood glucose, preventing heart disease and high blood pressure, and also enhancing the antioxidant system, insulin action and secretion [5]. Phyto-constituents have always guided the search for clinical trials, drug discovery and development. Hence, there is search for dietary constituents as well as natural antioxidants that can regulate blood glucose levels.
In the present review the connection between flavonoids and DM is focused on the basis of recent studies. The anti-diabetic activities of the flavonoids found in dietary plants, fruits, and summarize the underlying molecular signaling of the dietary flavonoids using in vitro and in vivo models to clarify their anti-diabetic effects.
Molecular mechanisms of insulin resistance
The molecular levels, the mechanisms underlying insulin resistance are being explored. Several mechanisms including abnormal insulin production, mutations in insulin receptor (IR) and its substrates, and insulin antagonists have been proposed, but it is now clearly known that defects in post-receptor signaling are the major cause of insulin resistance in target tissues [6]. Reduced expression, diminished tyrosine phosphorylation, or increased degradation of early insulin signaling molecules have been implicated.
Serine/threonine phosphorylation of IRS proteins can inhibit their activity and block downstream signaling. Various kinases including stress - activated protein kinase, c-Jun N-terminal kinase (JNK), inhibitor of nuclear factor kappa B (NF-κB) kinase (IκB), and protein kinase C (PKC) can phosphorylate IRS-1 and 2 at specific serine and threonine residues, leading to inhibition of insulin signaling [7]. Another underlying mechanism is the induction of inhibitory factors such as suppressors of cytokine signaling (SOCS-1 and 3). SOCS proteins block insulin signaling via competition with IRS-1 for association with the IR and by augmentation of proteosomal degradation of IRS-1 [8].
Increased activity of phosphatases which dephosphorylate intermediate signaling molecules can also inhibit the insulin signal pathway [9]. Several phosphatases have been implicated as inhibitors of insulin action. Protein tyrosine phosphatases (eg PTP1B) have negative effects on insulin signaling and are suggested to be major regulators of insulin signaling [10]. Other phosphatases such as phosphatase and tensin homologue (PTEN) which inactivates PI3-K and SH2-containing inositol 5′ phosphatase-2 [SHIP 2] have been shown to have a negative role on insulin signaling [11, 12].
Regulation of lipogenesis in adipocytes
Adipocyte transcription factors such as peroxisome proliferator-activated receptor gamma (PPARγ), sterol regulatory element-binding protein (SREBP-1c), adipokines like resistin, play crucial roles in adipocyte differentiation, adipogenesis, and accumulation of cellular lipid droplets.
PPARγ, a nuclear hormone receptor, is mainly expressed in adipose tissue and to a lesser extent in colon, immune cells and retina [13]. It plays a critical role in peripheral glucose homeostasis and energy metabolism and it also has been implicated in modulating adipogenesis and insulin sensitivity in vivo. PPARγ, induces glucose and fatty acid uptake by directly or indirectly enhancing the transcription of genes encoding proteins such as, fatty acid binding proteins (aP2), GLUT4 [14], fatty acid transport proteins, and acyl-CoA synthetase. Recent studies indicate that partial PPARγ antagonism by various flavonoids may be beneficial in improving insulin sensitivity and may also inhibit adipocyte differentiation and lipid accumulation [15].
Adiponectin is supposed to improve primarily glucose and lipid metabolism. Adiponectin also inhibits the expression of several pro-inflammatory cytokines [16], including tumor necrosis factor. Among other factors released within adipose tissue, tumor necrosis factor alpha (TNFα) promotes lipolysis and increases (free fatty acids) FFAs; both TNFα and interleukin 6 (IL-6) are related to mitochondrial dysfunction. Increased expression of TNFα and relatively low levels of adiponectin promote lipolysis and FFA release.
Leptin is an adipocyte-secreted hormone whose absence leads to dramatic metabolic derangements. Leptin regulates food intake at the level of hypothalamus and stimulates FFA oxidation in peripheral tissues to avoid lipid deposition. Insulin resistance in animal results in hyperleptinemia induces leptin resistance and causes lipotoxicity [17]. Leptin resistance causes a rise in FFA release to circulation followed by triglycerides (TG) synthesis (fatty liver) and decreased FFA oxidation in liver [18].
Flavonoids and their effects on diabetes
Flavonoids represent a large class of at least 6000 phenolic compounds found in fruits, vegetables, nuts, grain seeds, cocoa, chocolate, tea, soy, red wine, herbs and beverage products. Structurally, flavonoids consist of two aromatic rings (A and B rings) linked by a 3-carbon chain that forms an oxygenated heterocyclic ring (C ring). There are six subclasses of flavonoids including flavones, flavonols, flavanones, flavonols, isoflavones and anthocyanidins based on differences in generic structure of the C ring, functional groups on the rings and the position at which the B ring is attached to the C ring. Within each subclass, individual compounds are characterized by specific hydroxylation and conjugation patterns. Flavonoids that have the ability to scavenge free radicals and chelate metals [19]. Given the hypothesized relation between diabetes and inflammation [20, 21] and the potential for flavonoids to protect the body against free radicals and other pro-oxidative compounds [22, 23], it is biologically plausible that consumption of flavonoids or flavonoid-rich foods may reduce the risk of diabetes [24, 25]. New concepts have appeared with this trend, such as nutraceuticals, nutritional therapy, phytonutrients and phytotherapy. This functional foods and phytomedicines play positive roles in maintaining blood glucose levels, glucose uptake and insulin secretion and modulating immune function to prevent specific DM [26, 27]. In current years various approaches have been made to utilize the flavonoids in vitro and in vivo models by incorporating few novel methods to improve its antidiabetic activity. They are categorized in Table 1.
Table 1.
Important anti-diabetic potential and the underlying mechanism of dietary flavonoids
| Structure of flavonoid (s) | Plants/Dietary source | Specific mechanism of action | Model | References |
|---|---|---|---|---|
Diosmin 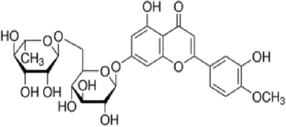
|
Scrophularia nodosa L., and citrus fruits | Stimulating the insulin production from the existing β-cells of pancreas. | STZ-nicotinamide-induced diabetic rats. | Srinivasan and Pari 2010, 2012, 2013 [30, 31, 33] |
| ↓ Lipids profile, improving lipid metabolizing enzymes, antioxidant and ↓ lipid peroxidation. | ||||
| ↓ Blood glucose, lipid peroxides, NO and ↑ body weight. | STZ-induced diabetic neuropathy rats | Jain 2014 [32] | ||
| ↓ Glycation. | STZ-diabetic rats | Vertommen 1994 [34] | ||
Fisetin 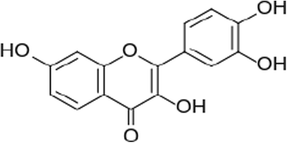
|
Strawberries, onion and persimmon | Improved glucose homeostasis. | STZ-diabetic rats | Prasath 2014 [38] |
| ↑ Glycolysis, ↓ gluconeogenesis. | STZ-diabetic rats | Prasath and Subramanian 2011, 2013 [39, 45] | ||
| ↓ Blood glucose, HbA1c, NF-κB p65, IL-1β and nitric oxide. | ||||
| Suppress HATs, particularly p300, leading to deacetylation of the p65 subunit of NF-κB. | THP-1 cell line | Kim 2012 [40] | ||
| Reduced cataract formation. | STZ-diabetic cataract in mice | Kan 2015 [41] | ||
| ↑ HDL, ↓ LDL and VLDL. | STZ-diabetic rats | Prasath and Subramanian 2014 [42] | ||
| Stimulate the NF-κB pathway, downregulated of adhesion molecules, inhibition of monocyte-endothelial adhesion. | Human umbilical vein endothelial cells and C57BL/6 mice | Kwak 2014 [43] | ||
| Improved glycemic and antioxidant status. | STZ-diabetic rats. | Prasath and Subramanian 2013 [44] | ||
| ↑ Mitochondrial function. | ||||
| ↓ Level methylglyoxal-dependent protein glycation. | C57BL/6-Ins2 Akita mice | Maher 2011 [46] | ||
Morin 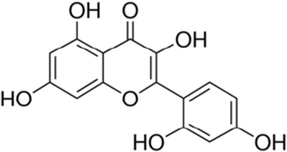
|
Prunus dulcis (Mill.) D.A. Webb., Chlorophora tinctoria (L.) Gaud., Psidium guajava L., fruits and wine | ↑ Insulin sensitivity and ↓ oxidative stress. | HFD-STZ-induced diabetic rats | Sendrayaperumal 2014 [49] |
| Inhibition of PTP1B, which behaves as an activator and sensitizer of the insulin receptor stimulating the metabolic pathways. | HepG2 cell line | Paoli 2013 [52] | ||
| Preventing the destruction of β-cells of the islets of Langerhans. | STZ induced diabetic rats | Vanitha 2014 [53] | ||
| Inhibition in ROS generation, translocation of apoptotic proteins, up-regulation of antioxidantgenes and Bcl-2 gene expression. | Hepatocytes cell line | Kapoor 2012 [54] | ||
Eriodictyol 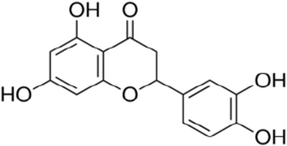
|
Eriodictyon californicum (Hook. & Arn.) Torr, Millettia duchesnei De Wild., Eupatorium arnottianum Griseb and lemon | ↑ Glucose uptake and improve insulin resistance | HepG2 cell line | Zhang 2012 [56] |
| ↓ TNFα, ICAM-1, VEGF, and eNOS. | STZ-induced diabetic rats | Bucolo 2012 [57] | ||
Hesperidin 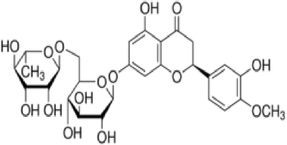
|
Orange citrus aurantium | Down-regulates generation of free radical, release of cytokines (TNF- α and IL-1β). | Neuropathy rats | Visnagri 2014 [60] |
| Anti-angiogenic, anti-inflammatory effects. | STZ-induced diabetic rats. | Shi 2012 [62] | ||
| ↓ Blood glucose by altering the activity of glucose-regulating enzymes. | STZ-induced type 1 diabetic rats | Akiyama 2010 [63] | ||
| Reduced oxidative stress, apoptosis and improving cardiac function via the PPAR-γ pathway. | STZ-isoproternol induced diabetic rats | Yo 2014 [64] | ||
| ↓ Inflammatory cytokies | HFD-STZ-induced type 2 diabetic rats. | Mahmoud 2012 [65] | ||
| ↓ HbA1c, glucose, CES LDL, TC, TG levels, systolic and diastolic blood pressure. | STZ-nicotinamide induced myocardial infarction in diabetes in rats | Kakadiya 2010 [66] | ||
| Regulation of glucose and lipid metabolism. | Goto-Kakizaki type 2 diabetes rats | Akiyama 2009 [67] | ||
| Regulation of glycolysis, gluconeogenesis, hepatic glycogen stores. | C57BL/KsJ-db/db mice | Jung 2014 [68] | ||
| ↓ Lipid peroxidation, ↑ GSH, GR and GST | STZ-induced diabetes rats | Ashafaq 2014 [69] | ||
Naringenin 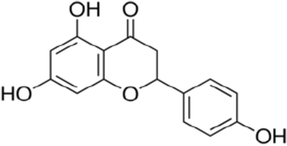
|
Cochlospermum vitifolium (Willd.) Spreng., grapefruits, oranges and tomatoes | Inhibition of intestinal α-glucosidase activity. | High HFD-STZ induced diabetic rats. | Priscilla 2014 [73] |
| Reduced oxidative damage | STZ-induced diabetes rats | Fallahi 2012 [75] | ||
| ↓ Cholesterol and cholesterol ester synthesis. | High-fat induced diabetic mice | Mulvihill 2009 [80] | ||
| Improved overall insulin sensitivity and glucose tolerance. | ||||
| Suppressed monocyte chemoattractant protein-1 and inhibition of c-Jun NH2-terminal kinase pathway. | HFD-induced obesity | Yoshida 2014 [82] | ||
| ↓ Hyperglycemia and ↑ antioxidant enzyme (SOD). | STZ-induced hyperalgesia and allodynia in rats | Hasanein 2014 [83] | ||
| Stimulated insulin secretion | INS-1E cells | Bhattacharya 2014 [84] | ||
| Decreased fasting glucose and inflammatory cytokines. | HFD-fed mice | Yoshida 2013 [85] | ||
| ↓ Oxidative stress. | STZ-induced diabetes rats | Rahigude 2012 [86] | ||
Apigenin 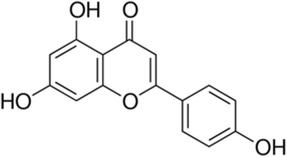
|
Hypericum perforatum L., Matricaria chamomilla L., parsley, onions, oranges, tea, chamomile and wheat sprouts. | ↓ Glucose and G-6-Pase activity and ↑ antioxidant enzymes. | Alloxan-induced diabetic mice | Panda and Kar 2007 [88] |
| ↓ Apoptosis, ↑ antioxidant and mitochondrial protection. | HIT-T15 pancreatic β-cells | Suh 2012 [90] | ||
| Inhibition TNF-α and IL-1β-induced activation of NF-κB. | Human THP-1 monotypic cells | Zhang 2014 [92] | ||
| Inhibition the expression of VCAM1, IKKα and IKKepsilon/IKKi. | Human endothelial cells | Yamagata 2010 [93] | ||
| Insulin-secretagogue. | Male Wistar rats | Cazarolli 2009 [94] | ||
Baicalein 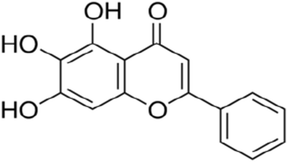
|
Scutellaria baicalensis Georgi and Scutellaria lateriflora L. | Improved glucose tolerance, and islet β-cell survival and mass. | HFD-induced obese mice. | Fu 2014 [97] |
| Suppressed the activation of NF-κB, ↓ iNOS, TGF-β1, ALP, SGOT and SGPT. | HFD-STZ-induced type 2 diabetic Wistar rats | Ahad 2014 [99] | ||
| Reduced AGEs and TNF-α level, decreased NF-κB activation. | STZ-induced diabetic rats | El-Bassossy 2014 [100] | ||
| Improvement of insulin resistance, protective by phosphorylating AMPKα AND INS-1. | HFD-induced mice | Pu 2012 [101] | ||
| Restored the impairment of PI3K/Akt pathway and ↓ GSK3β. | STZ-induced diabetic Wistar rats | Qi 2015 [102] | ||
Chrysin 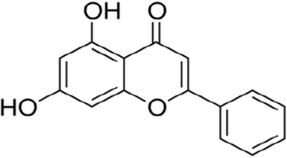
|
Honey, Passiflora caerulea (L.), Pelargonium peltatum (L.), Tilia tomentosa Moench, Pelargonium quercifolium (L.f.) L’Hér. and Pelargonium crispum (Berg.) L’Her | Inhibition of TNF-α pathway, leads to the decreased secretion of pro-inflammatory cytokines. | HFD-STZ-induced type 2 diabetic Wistar albino rats | Ahad 2014 [107] |
| Downregulated the increased expression of TGF-β, fibronectin and collagen-IV proteins. | ||||
| ↓ Blood glucose, oxidative stress, improved learning and memory function. | STZ-induced diabetic rats | Li 2014 [108] | ||
Luteolin 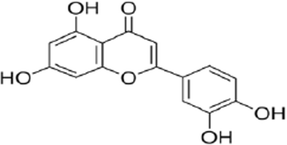
|
Celery, parsley, broccoli, onion leaves, carrots, peppers, cabbages and apple skins. | inhibition of the NF-κB pathway. | HFD-induced in obesity mice | Liu 2014 [116] |
| Increased HO-1 expression and elevated antioxidants. | STZ-Induced Diabetic Rats. | Wang 2011 [117] | ||
| Decreased activity of NF-κB was implicated in inhibition by luteolin of increased iNOS. | Min6 insulin secreting cell line | Ding 2014 [119] | ||
| Reduced CREB-binding protein/p300 gene expression. | Human monocytic (THP-1) cell line | Kim 2014 [120] | ||
| Suppression of hepatic lipogenesis and increased in uptake of FFAs. | HFD-induced C57BL/6 J mice | Kwon 2015 [121] | ||
| Up-regulated the myocardial eNOS pathway and downstream effects include the enhancement of MnSOD and inhibition of mPTP. | STZ/L-NAME-induced diabetes rats | Yang 2015 [122] | ||
| Reduced mast cell and macrophage infiltrations and inflammatory cytokine levels. | Diet-induced obesity | Xu 2014 [123] | ||
Tangeretin 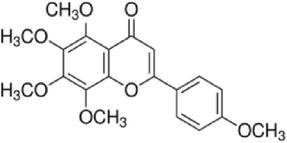
|
Citrus fruit rinds, mandarin orange and Poncirus trifoliate (L.) Raf. | Stimulated AMPK activation may be associated with anti-inflammatory. | HFD-induced obese mice | Kim 2012 [124] |
| ↑ Insulin, glycogen. | STZ-induced diabetic rats | Sundaram 2014 [125] | ||
Wogonin 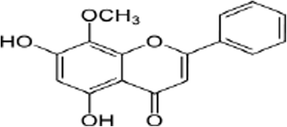
|
Scutellaria baicalensis Gerogi | Inhibition of p38 MAPK by its specific inhibitor SB203580 increasing PPARα activity and decreasing OPN expression. | STZ induced type 1 diabetes | Zhang 2015 [130] |
| Anti-adipogenic effect by acting as a PPARα agonist, which could prevent weight gain. | C57BLKS/J-Leprdb/Leprdb mice and 3 T3-L1 cells | Bak 2014 [128] | ||
Isorhamnetin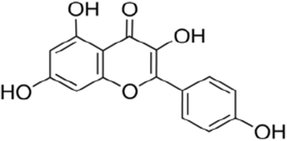
|
Hippophae rhamnoides L., Oenanthe javanica (Blume) DC, Ginkgo biloba L., and Opuntia ficus-indica (L.) Mill. | Insulin secretion, associated with increased GLUT2 and PPARγ. | HFD-induced C57BL/6 mice | Rodríguez-Rodríguez 2015 [133] |
| Inhibition adipogenesis through downregulation of PPARγ and C/EBPα. | 3 T3-L1 cells | Lee 2009 [134] | ||
Kaempferol 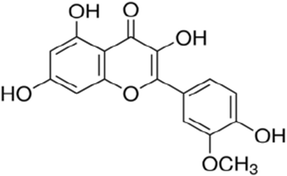
|
Tea, cruciferous vegetables, grapefruit, Gingko biloba L., and some edible berrie. | Inhibited cellular apoptosis, and reduced caspase-3 activity in beta-cells. | INS-1E β-cells | Zhang 2011 [139] |
| ↑ Antioxidant and ↓ decreased of lipid peroxidation markers. | STZ-induced diabetic rats | Al-Numair 2015 [141] | ||
| ↓ PPAR-γ and SREBP-1c expression. | HFD-obese mice | Zang 2015 [143] | ||
| Restore deranged activity of membrane-bound ATPases. | STZ-induced diabetes | Al-Numair 2015 [144] | ||
| Enhancing β-cell survival, improved cAMP signaling. | INS-1E cells. | Zhang 2013 [145] | ||
| ↑ GLUT 4, AMPK | HFD-induced diabetic mice | Alkhalidy 2015 [146] | ||
Rutin 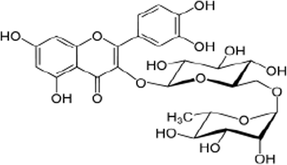
|
Buckwheat, oranges, grapes, lemons, limes, peaches and berries | Inhibited inflammatory cytokines, improving antioxidant and lipid profiles. | HFD-STZ-induced type 2 diabetic model | Niture 2014 [154] |
| ↓ Glucose, TBARS, caspase-3 and ↑ insulin, Bcl-2 protein. | STZ-induced diabetic rat retina | Ola 2015 [156] | ||
| Protected pancreatic beta-cell by decreasing oxidative stress. | STZ induced diabetic rats | Kamalakkannan and Prince 2006 [157] | ||
| ↓ MDA levels and ↑ SOD and CAT. | STZ-induced type 1 diabetic rats | Butchi 2011 [158] | ||
Quercetin 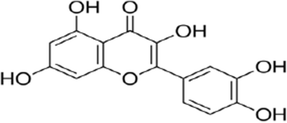
|
Chokeberries, black currants, apples and cherries | Increased the activity of glycogen synthase, the rate-limiting enzyme of glycogen synthesis. | Murine H4IIE and human HepG2 cells. | Eid 2015 [164] |
| Inhibition of the two transcriptional factors and the activation of mTORC1/p70S6K. | HK-2 and NRK-52E cells | Lu 2015 [167] | ||
| Inhibitory effects on NF-kB and caspase-3 expression. | STZ-induced diabetic rats | Kumar 2014 [168] | ||
| Ameliorated hyperglycemia and oxidative stress. | Alloxan induced type 2 diabetic mice. | Alam 2014 [169] | ||
| Prevented β-cell death via the mitochondrial pathway and NF-κB signaling. | RINm5F β-cells. | Dai 2013 [170] | ||
| Reduced expression of inducible iNOS and inhibited translocation of NF-κB. | ||||
| Reduced TBARS levels, TC and elevated activities of SOD, CAT, and GSH-Px and HDL-cholesterol. | Diet-C57BL/KsJ-db/db mice | Jeong 2012 [171] | ||
| Improved renal function in rats with diabetic nephropathy by inhibiting the overexpressions of TGF-β1 and CTGF. | STZ-induced diabetic rats | Lai 2012 [172] | ||
| ↓ Glucose and blood HbA1c. | STZ-induced diabetic rats | Kim 2011 [173] | ||
Genistein 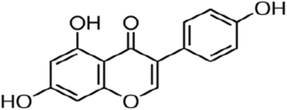
|
Fava bean, soybeans and kudzu. | ↑ cAMP signalling ↑ PKA activation. | HG-induced diabetic mice | Babu 2012 [176] |
| ↑ Insulin-positive β-cell. | HFD-induced C57BL/6 mice | Fu 2012 [179] | ||
| Activation of ERα seems to stimulate muscular GLUT4 functionality, activation of ERβ. | Zucker diabetic fatty rats | Weigt 2015 [182] | ||
| ↓ Glucose, HbA1c, C-reactive protein, TNFα and TGFβ1 protein expressions. | STZ-induced diabetes rats | Gupta 2015 [183] | ||
| ↓ Inflammatory markers and improved oxidative stress. | Alloxan-induced diabetic mice | Kim and Lim 2013 [184] | ||
| Improved wound angiogenesis. | STZ-induced type 1 diabetic mice | Tie 2013 [185] | ||
| Reduced hyperglycemia via minimization of islet cell loss. | Alloxan-induced Sprague–Dawley rats | Yang 2011 [186] | ||
| Reduced glucose tolerance and improved insulin levels. | STZ-induced diabetic mice | Fu 2010 [187] | ||
| Inhibition the secretion of ECM components and the expression of TGF-beta. | HG-cultured rat mesangial cells | Yuan 2012 [188] | ||
| Suppressed the expression of CCAAT/enhancer binding protein alpha (C/EBPalpha). | 3 T3-L1 cells | Zhang 2009 [189] | ||
| ↓ TGF-β2, αB-crystallin, and fibronectin. | Human lens epithelial (HLE-B3) cells | Kim 2008 [190] | ||
| ↓ G6Pase, PEPCK and ↑ lipogenic enzymes activities. | Non-obese diabetic mice | Choi 2008 [191] | ||
Daidzein 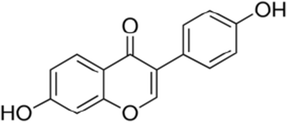
|
Soy milk, soybeans and nuts | Potent α-glucosidase inhibitor and suppress the postprandial hyperglycemia. | STZ-induced diabetic mice | Park 2013 [194] |
| ↓ Blood glucose and urinary glucose excretion. | HFD-induced type 2 diabetes | Cheong 2014 [196] | ||
| Improved the endothelial dysfunction. | STZ-induced diabetic rats. | Roghani 2013 [197] | ||
| ↑ IRS-1, GLUT4 and enhanced insulin stimulated glucose uptake. | 3 T3-L1and C3H10T1/2 cells | Cho 2010 [198] |
Diosmin
Diosmin was first isolated in 1925 from Scrophularia nodosa L. Diosmin is a naturally occurring flavonoid glycoside that can be isolated from various plant sources or derived by dehydrogenation of the corresponding flavanone glycoside hesperidin that is abundant in the pericarp of various citrus fruits [28]. Diosmin has been shown to improve factors associated with diabetic complications. Blood parameters of glycation and oxidative stress was measured in type I diabetic patients before and after intervention with a diosmin. A decrease in glycated hemoglobin (HbA1c) was accompanied by an increase in glutathione peroxidase (GPx) [29]. In studies with rats, orally treatment of diosmin for 45 days significantly lowered plasma glucose level, increased the activities of hepatic key enzymes such as hexokinase and glucose-6- phosphate dehydrogenase (G6PD) in addition to decreasing glucose-6-phosphatase (G6Pase) and fructose-1,6-bisphosphatase (FDPase) in streptozotocin (STZ)-nicotinamide treated rats exhibiting its anti-hypeglycemic activities [30]. Studies from the same author claimed that, diosmin lowered plasma glucose and increased plasma insulin levels in diabetic rats by ameliorating the oxidative stress induced by STZ-nicotinamide and the activities of antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT), GPx, and reduced glutathione (GST), vitamin C, vitamin E and reduced glutathione were increased while lipid peroxidation was reduced in liver and kidney of diabetic rats upon treatment with diosmin [31]. In recent study, treatment with diosmin at doses of 50 and 100 mg/kg bw for 1 month ameliorated hyperglycemia and oxidative stress [32].
Fisetin
Fisetin is a flavonoid, also a dietary ingredient found strawberry, apple, persimmon, grape, onion, cucumber and Cotinus coggygria Scop. [35, 36]. The results of Constantin et al. [37] showed that the fisetin inhibition of pyruvate transport into the mitochondria and the reduction of the cytosolic NADH-NAD (+) potential redox could be the causes of the gluconeogenesis inhibition. Fisetin could also prevent hyperglycemia by decreasing glycogen breakdown or blocking the glycogenolytic action of hormones. Fisetin was also reported that at a dose of 10 mg/kg bw to diabetic rats for 30 days, displayed reductions in blood glucose, HbA1c levels, increased plasma insulin and decreased mRNA and protein expression levels of gluconeogenicgenes, such as phosphoenol pyruvate carboxykinase and G6PD in liver [38]. There is evidence based in vivo animal studies showed that fisetin treatment significantly decreased the levels of blood glucose, HbA1c, NF-κB p65 unit and interleukin-1 beta (IL-1β), serum nitric oxide (NO) with an improved in plasma insulin antioxidant status [39]. Moreover, fisetin inhibits high glucose (HG)-induced cytokine production in monocytes, through epigenetic changes involving NF-κB. Thus, fisetin supplementation could be considered for diabetes prevention [40].
Morin
Morin, a natural flavonoid, and a major component of traditional medicinal herbs were Prunus dulcis (Mill.) D.A. Webb, Chlorophora tinctoria (L.) Gaud., Psidium guajava L., fruits and wine [47, 48]. In animal models, oral administration of morin for 30 days significantly improved hyperglycemia, glucose intolerance, and insulin resistance. The elevated levels of lipid peroxides were declined and the antioxidant competence was found to be improved in diabetic rats treated with the morin. The status of the lipid and lipoprotein profile in the serum was normalized upon treatment. Levels of TNFα decreased upon treatment with morin [49]. A study carried out by Abuohashish et al., [50] demonstrated that morin (30 mg/kg bw) was effective in reducing the elevated inflammatory cytokines IL-1β, IL-6 and TNF-α in diabetic animals, which support its anti-inflammatory property and also its possible beneficial effects in diseases where inflammation. Morin was found to ameliorate high fructose-induced hepatic SphK1/S1P signaling pathway impairment, resulting in the reduction of hepatic NF-κB activation with IL-1b, IL-6 and TNF levels in rat liver and BRL3A cells. Subsequently, morin recovered hepatic insulin and leptin sensitivity, then reduced hyperlipidemia and liver lipid accumulation in animal and cell line models [51]. Paoli et al. [52] reported that dietary morin inhibitor of PTP1B, which behaves as an activator and sensitizer of the insulin receptor stimulating the metabolic pathways. However, in this context Vanitha et al. [53] showed that morin treatment significantly reduced the blood glucose, G6Pase and FDPase and increased the insulin levels, hexokinase and G6PD activities. Thus, morin, through these metabolic effects, may exert several beneficial effects in the prevention of diabetes.
Eriodictyol
Eriodictyol is present in lemon fruit. It has been demonstrated [55] that supplementation of lemon flavonoids, such as eriocitrin and hesperidin, significantly suppressed the oxidative stress in diabetic rats. Eriodictyol treatment was associated with up-regulated the mRNA expression of PPARγ2 and adipocyte-specific fatty acid-binding protein as well as the protein levels of PPARγ2 in differentiated 3 T3-L1 adipocytes. Furthermore, it reactivated Akt in HepG2 cells with HG-induced insulin resistance [56]. Eriodictyol has been reported to significantly lower retinal TNFα, intercellular adhesion molecule 1 (ICAM-1), vascular endothelial growth factor (VEGF), and endothelial NOS (eNOS). Further, treatment with eriodictyol significantly suppressed diabetes-related lipid peroxidation [57].
Hesperidin
Hesperidin is an abundant and inexpensive by product of Citrus cultivation and isolated from the ordinary orange Citrus aurantium L., and other species of the genus Citrus (family: Rutaceae) contain large amounts of hesperidin [58, 59]. Hesperidin not only attenuated the diabetic condition but also reversed neuropathic pain via control over hyperglycemia as well as hyperlipidemia to down-regulate generation of free radical, release of pro-inflammatory cytokines [60]. Reduced oxidative stress by hesperidin was also noted in this studies [61, 62]. Treatment of hesperidin (10 g/kg diet) decreased blood glucose by altering the activity of glucose regulating enzymes, and normalized the lipids and adiponectin levels [63]. In a recent study, Yo et al. [64] found that hesperidin treatment significantly improved mean arterial pressure, reduced left ventricular end-diastolic pressure, and improved both inotropic and lusitropic function of the heart. Furthermore, hesperidin treatment significantly decreased the level of thiobarbituric acid reactive substances (TBARS) and increased the activity of lactate dehydrogenase (LDH).
Naringenin
Naringenin is abundantly found in citrus fruits such as grapefruits, oranges and tomatoes that has been reported to have antioxidant potential [70]. In vitro studies have shown that naringenin had an insulin-mimic effect to decrease poliprotein B secretion in hepatocytes [71]. In vivo studies of Cochlospermum vitifolium (Willd.) Spreng which contains naringenin decreased blood glucose levels in healthy male Wistar rats [72]. Oral treatment of naringenin (25 mg/kg bw) exerts significant inhibition of intestinal α-glucosidase activity in vivo thereby delaying the absorption of carbohydrates in diabetic rats, thus resulting in significant lowering of postprandial blood glucose levels. Both in vitro and in vivo results were compared to the commercially available α-glucosidase inhibitor acarbose [73]. Naringenin was found to inhibit the glucose uptake in everted rat intestinal sleeves by the inhibition of intestinal sodium-glucose co-transporters [74]. Naringenin was also found to prevent the functional changes in vascular reactivity in diabetic rats through nitric oxide and not prostagland in dependent pathway [75]. Naringin is effective in protecting against the development of metabolic syndrome through changing the expression of hepatic genes involved in lipid metabolism and gluconeogenesis via upregulation of both PPAR and 5' adenosine monophosphate-activated protein kinase (AMPK), involving the activation of multiple types of intracellular signaling in mice exposed to a HFD [76]. The research findings of Zygmunt et al. [77] provide support for the stimulation of muscle glucose uptake by naringenin in a dose-dependent manner and independent of insulin. The in vivo anti-diabetic effects of naringenin may be AMPK-mediated. Activation of AMPK increases glucose tolerance, insulin sensitivity. Furthermore, naringenin administration decreased plasma glucose levels in STZ-induced diabetic rats [78], improved insulin sensitivity in fructose-fed insulin resistant rats [79], and reduced insulin resistance in HFD mice [80]. A recent study stated that naringenin (25 mg/kg bw) treatment to diabetic rats for 45 days markedly reduced hyperglycemia and hyperinsulinemia, restored lipid profile changes, decreased membrane lipid peroxidation; enhanced the activities of antioxidants and improved hepatic function markers [81].
Apigenin
Apigenin is a member of the flavone family and is found in many fruits, vegetables, nuts, onion, orange and tea [87]. Alloxan-induced elevation in serum cholesterol, hepatic lipid peroxidation and a decrease in the activity of cellular antioxidants, such as CAT, SOD and GSH content were observed, administration of apigenin to diabetic mice ameliorated hyperglycemic and improved antioxidants [88]. Apigenin reduced parathyroid hormone related protein stimulated increases in the human pancreatic stellate cells messenger RNA expression levels of extracellular matrix proteins collagen 1A1 and fibronectin, proliferating cell nuclear antigen, TGF-β, and IL-6 [89]. Suh et al. [90] suggest that apigenin attenuates dRib induced cell damage in pancreatic β-cells via oxidative stress-related signaling. Apigenin preserves the cellular architecture of vital tissues towards normal in STZ-induced diabetic rats. Furthermore, enhanced GLUT4 translocation upon apigenin treatment suggests more glucose lowering as well as β-cell preserving efficacy [91].
Baicalein
Baicalein, a flavonoid originally isolated from the roots of Scutellaria baicalensis Georgi and fruits of Oroxylum indicum (L.) Benth has been shown to exhibit strong free radical scavenging [95, 96]. Fu et al. [97] induced diabetes mice by HFD feeding and low doses of STZ and they the administered HF diet containing 0.25 or 0.5 g baicalein/kg diet. They observed that diabetic mice treated with baicalein displayed significantly improved hyperglycemia, glucose tolerance, and insulin levels. They provide the rationale for screening and preclinical studies of hydroxyflavones, especially those with an improved pharmacological profile, as potential therapeutics for diabetic ant its complications [98]. Baicalein treatment significantly lowered food intake, body weight and levels of fasting blood glucose, HbA1c in diabetic rats. Baicalein also suppressed the activation of NF-κB, decreased expression of iNOS and TGF-β1, and ameliorated the structural changes in renal tissues [99]. Previous findings also reported that treatment with baicalein reduced the advanced glycation end-product (AGEs) and TNF level, decreased NF-κB activation and inhibited histopathological changes [100]. Mechanism of its action was up-regulation of AMPK and its related signal pathway. AMPK is not only a major cellular energy sensor, but also a master regulator of metabolic homeostasis involving inflammation and oxidative stress. Activated AMPK could abolish inflammation through the MAPKs signaling pathway; activated AMPK could attenuate insulin resistance by phosphorylating IRS-1, AKT and dephosphorylate ERK, JNK and NF-κB; it also suppresses fatty acid synthesis, gluconeogenesis and increases mitochondrial β-oxidation [101].
Chrysin
Chrysin, found in honey, bee pollen, propolis, fruits, vegetables, beverages and medicinal plants such as Passiflora caerulea (L.), Pelargonium peltatum (L.), Tilia tomentosa Moench, Pelargonium quercifolium (L.f.) L’Hér and Pelargonium crispum (Berg.) L’Her [103–106]. Chrysin is the main component of Oroxylum indicum (L.) Benth. ex Kurz, which is one of the most common herbal medicines used in China and other East Asian countries. Ahad et al. [107] in a recent study mentioned that chrysin treatment improved renal pathology and suppressed TGF-β, fibronectin and collagen-IV protein expressions in renal tissues. Chrysin also significantly reduced the serum levels of pro-inflammatory cytokines, IL-1β and IL-6. Therefore, chrysin prevents the development of diabetic neuropathy (DN) in HFD/STZ-induced diabetic rats through anti-inflammatory effects in the kidney by specifically targeting the TNF-α pathway. Li et al., [108] investigation revealed that chrysin significantly and dose-dependently inhibited the oxidative stress, together with improved cognition in diabetic rats. In addition, treatment chrysin was shown to reduce glucose and lipid peroxidation level and improve insulin levels in diabetic rats [109], suggesting that this chrysin may exert anti-hypertensive and vascular complication associated with anti-diabetic effects [110].
Luteolin
Vegetables and fruits such as celery, parsley, broccoli, onion leaves, carrots, peppers, cabbages, apple skins, and chrysanthemum flowers are rich in luteolin [111–114]. That luteolin potentiates insulin action, increases expression and transcriptional activation of PPARγ and expression of the PPARγ target genes adiponectin, leptin and GLUT4 in 3T3-L1 adipocytes, as well as in primary mouse adipose cells, and that PPARγ antagonist inhibits this induction [115]. It has also been reported that luteolin up-regulated the expression of the synaptic proteins, and alleviated the HFD-induced cognitive deficits. It is possible that the decrease in circulating levels of inflammatory molecules MCP-1, resistin and the elevation of adiponectin levels in obese mice by luteolin may, in turn, mediate beneficial effects on metabolic pathways implicated in insulin resistance and DM pathophysiology [116]. The mechanism of the renoprotective effect of luteolin may be related to increasing HO-1 expression and elevating antioxidant in diabetic nephropathy [117]. It has been reported that luteolin ameliorated inflammation related endothelial insulin resistance in an IKKb/IRS-1/Akt/eNOS-dependent pathway [118]. Luteolin, also improved insulin secretion in uric acid damaged pancreatic β-cells by suppressing the decrease of MafA mainly through the NF-κB, iNOS-NO signaling pathway [119]. In addition, it has been reported that luteolin was significantly reduced cAMP-response element binding protein (CREB)-binding protein/p300 gene expression, as well as the levels of acetylation and histone acetyl transferase activity of the CBP/p300 protein, which is a known NF-κB coactivator [120].
Tangeretin
Tangeretin, abundant in the citrus fruit rinds, including mandarin orange, Poncirus trifoliate Raf. (Rutaceae), and Yuja, found in Korea. Administration of HFD plus 200 mg/kg bw of tangeretin exhibited reduction in body weight, total cholesterol (TG), blood glucose and decreased adipocytokinese such as adiponectin, leptin, resistin, IL-6, and MCP-1 [124]. Oral administration of tangeretin (100 mg/kg bw) to diabetic rats for 30 days resulted in a significant reduction in the levels of plasma glucose, HbA1c and increased in the levels of insulin and hemoglobin. Tangeretin enhances the glycolytic enzymes and controls the glucose metabolism in the hepatic tissues of diabetic rats by stimulating insulin production from existing β-cells of pancreas by its antioxidant potential [125]. Tangeretin 3T3-L1 preadipocytes into adipocytes possessing less intracellular triglyceride as compared to vehicle-treated differentiated 3T3-L1 adipocytes. Tangeretin increased the secretion of an insulin-sensitizing factor, adiponectin, but concomitantly decreased the secretion of an insulin-resistance factor, monocyte chemotactic protein-1 (MCP-1), in 3T3-L1 adipocytes [126].
Wogonin
Wogonin extracted from the root of Scutellaria baicalensis Gerogi (Scutellariae radix) has long been used as a traditional medicine in East Asian countries [127]. Wogonin has beneficial effects on blood glucose level, insulin sensitivity, and lipid metabolism via selective PPARα and AMPK activation without the adverse side-effects of weight gain and fatty liver [128]. High glucose (HG)-induced markedly increased vascular permeability, monocyte adhesion, expressions of cell adhesion molecules, formation of ROS and activation of NF-κB. Remarkably, all of the above mentioned vascular inflammatory effects of HG were attenuated by pretreatment with wogonin [129].
Isorhamnetin
Isorhamnetin is a bioactive compound found in medicinal plants, such as Hippophae rhamnoides L., Oenanthe javanica (Blume) DC., and Ginkgo biloba L. In a STZ-induced model of diabetes, oral administration of isorhamnetin (10, 20 mg/kg BW) for 10 days ameliorated hyperglycemia and oxidative stress [131]. In another study, administered orally at 25 mg/kg in STZ-induced diabetic rats caused not only a significant inhibition of serum glucose concentration but also sorbitol accumulation in the lenses, red blood cells, and sciatic nerves [132]. In recent year, there is experimental evidence suggesting that isorhamnetin glycosides may possess the antidiabetic effect and their influence on lipid content, endoplasmic reticulum stress markers and the expression of enzymes regulating lipid metabolism [133].
Kaempferol
Kaempferol is a flavonol that is relatively abundant in Ginkgo biloba L., cruciferous vegetables, grapefruit, tea and some edible berries [135–137]. Also, kaempferol is isolated from Bauhinia forficata leaves, is able to diminish the increased serum glucose level and increase glucose uptake in the rat soleus muscle as efficiently as insulin [138]. The in vitro results demonstrated that kaempferol treatment (10 μM) promoted viability, inhibited cellular apoptosis, and reduced caspase-3 activity in β-cells and human islets chronically exposed to hyperglycemic condition. These protective effects are associated with improved cAMP signaling, anti-apoptotic Akt and Bcl-2 protein expression, and insulin secretion and synthesis in β-cells [139]. The author suggests that kaempferol stimulates glucose uptake in the rat soleus muscle via the PI3K and PKC pathways and, at least in part, independently of MEK pathways and the synthesis of new glucose transporters [140]. Administration of kaempferol to diabetic rats was shown back to near normal levels in plasma glucose, insulin, lipid peroxidation products, enzymatic, and non-enzymatic antioxidants [141]. Published data showed that kaempferol reduced IL-1β, TNF-α, lipid peroxidation and nitrite, concomitant with the improvement of antioxidant defense and body weight gain [142]. Orally administrated kaempferol was significantly decreased fasting blood glucose, serum HbA1c levels and improved insulin resistance. Gene expression analysis of the liver showed that kaempferol decreased PPAR-γ and SREBP-1c expression. Anti-obese and anti-diabetic effects of kaempferol are mediated by SREBP-1c and PPAR-γ regulation through AMPK activation [143].
Rutin
Rutin can be broadly extracted from natural sources such as buckwheat, oranges, grapes, lemons, limes, peaches and berries [147, 148]. Specifically, diabetic mice fed rutin at 100 mg/kg diet displayed significant lower of plasma glucose and increase in insulin levels were observed along with the restoration of glycogen content and the activities of carbohydrate metabolic enzymes [149]. Fernandes et al. [150] showed that rutin could improve the metabolic status of rats with experimentally-induced diabetes. Rutin is involved in the activation of liver enzymes associated with gluconeogenic, and lipid metabolic processes [151] and also decreased the levels of fasting blood glucose, creatinine, blood urea nitrogen, urine protein, the intensity of oxidative stress and p-Smad 7 significantly. The expression of AGEs, collagen IV and laminin, TGF-β1, p-Smad 2/3 and connective tissue growth factor was inhibited by rutin significantly [152]. Rutin have been shown to stimulate glucose uptake in the rat soleus muscle via the PI3K, a typical protein kinase C and mitogen-activated protein kinase pathways [153].
Rutin has been reported to significantly improved body weight, reduced plasma glucose and HbA1c, pro-inflammatory cytokines (IL-6 and TNF-alpha), and restored the depleted liver antioxidant status and serum lipid profile in HFD/STZ induced diabetic rats [154]. Notably, rutin was shown to protect and improve myocardial dysfunction, oxidative stress, apoptosis and inflammation in the hearts of the diabetic rats [155].
A recent study showed that rutin supplementation enhanced the reduced levels of brain-derived neurotrophic factor, nerve growth factor, and GSH, and reduced the level of TBARS. In addition, rutin treatment showed anti-apoptotic activity by decreasing the level of caspase-3 and increasing the level of Bcl-2 in the diabetic retina [156].
Quercetin
Numerous studies have focused on quercetin to develop it as antidiabetic drug to prevent and manage DM. Quercetin is one of the most widely used flavonols in human dietary nutrition [159]. It is widely distributed in different types of fruits, tea, lovage, pepper, coriander, fennel, radish, dill, berries, onions, apples and wine [160]. Several studies have reported quercetin mechanism of action in diabetes, such as decreases in lipid peroxidation, increases in antioxidant enzymes (like SOD, GPX, and CAT) activities, inhibition of insulin-dependent activation of PI3K, and reduction in intestinal glucose absorption by inhibiting GLUT2 [161, 162]. The study performed by Kwon et al. [163] evaluated the effect of quercetin on Caco-2E intestinal cells, the study documented that the transport of fructose and glucose by GLUT2 was strongly inhibited by quercetin. Blockage of tyrosine kinase is another mechanism by which quercetin is reported to have effects against diabetes. Eid et al. [164] reported that quercetin stimulates GLUT4 translocation and expression in skeletal muscle, by mechanisms associated with the activation of AMPK rather than insulin-dependent pathways such as Akt. The study performed in rats indicated that quercetin ameliorated the expression of markers of oxidative stress and inflammation, such as Nrf2, heme oxygenase-1, and NF-κB, suggesting that quercetin anti-inflammatory effects on adipose tissue can be associated with the reduction of body weight [165]. Likewise, Kobori et al. [166] reported that quercetin in the diet led to the recovery of cell proliferation in diabetic mice.
Isoflavones
Isoflavonoids are another subclass of the phenolic phytonutrients. Soybeans are an unusually concentrated source of isoflavones, including genistein and daidzein, and soy is the major source of dietary isoflavones, is reported to have numerous health benefits attributed to multiple biological functions. Over the past 10 years, numerous studies have demonstrated that isolavones has anti-diabetic effects, in particular, direct effects on β-cell proliferation, glucose-stimulated insulin secretion.
Genistein
Genistein, a naturally occurring soy isoflavone, is a flavonoid presented in legumes and Chinese plants Genista tinctoria Linn and Sophora subprostrala Chun et T Chen [174]. Molecular mechanisms underlying the modulatory effect of genistein on diabetes, that specifically focus on neutrophils, are needed to understand contributions of estrogenic and enzyme inhibitory activities (tyrosine kinase inhibition) to dysregulat glucose homeostasis [175]. Genistein has also been reported to improve hyperglycemia caused human vascular endothelial inflammation ex vivo, which is at least partially mediated through promoting the cAMP/PKA signaling pathway [176]. Palanisamy et al. [177], focused on the protective role of genistein on renal malfunction in rats fed a fructose rich diet, through the modulation of insulin resistance induced pathological pathways. Another study has shown that genistein injections (10 mg/kg) reduced urinary TBARs excretion and renal gp91phox expression, as well as decreased production of inflammatory markers, including p-ERK, ICAM-1, and MCP-1, in DN mice [178]. In this study, dietary intake of genistein (250 mg/kg bw) improved hyperglycemia, glucose tolerance, and blood insulin level in obese diabetic mice, whereas it did not affect body weight gain, food intake, fat deposit, plasma lipid profile, and peripheral insulin sensitivity [179]. Furthermore, genistein has been shown to protect against oxidative stress and inflammation, neuropathic pain, and neurotrophic and vasculature deficits in the diabetic mouse model [180]. Indeed, recent findings indicated that genistein administration significantly decreased β-cells loss and improved glucose and insulin levels [181].
Daidzein belongs to the isoflavone subclass of flavonoids and is found in fruits, nuts, soybeans, and soy-based products [192]. An earlier study suggested that daidzein exerts anti-diabetic effects by improves glucose and lipid metabolism [193, 194]. Cederroth et al. [195] demonstrated that the dietary soy including genistein improve insulin sensitivity by increasing glucose uptake in skeletal muscle in mice. Treatment of daidzein proved to be effective in reducing blood glucose; TC levels and improved the AMPK phosphorylation in gastrocnemius muscle [196].
Anthocyanins
Anthocyanins are flavonoids in flowers and fruits (red, blue and purple tints in apples, berries, red grapes, eggplant, red cabbage and radishes). Dietary consumption of anthocyanins is high compared to other flavonoids, owing to their wide distribution in plant materials. Based upon many cell-line studies and animal models, it has been suggested that anthocyanins possess anti-diabetic activities (Table 2).
Table 2.
The in vitro and in vivo effect on anti-diabetic and underlying mechanism of anthocyanins
| Strcture of anthocyanins | Plants/dietary source | Specific mechanism of action | Model | References |
|---|---|---|---|---|
Cyanidin 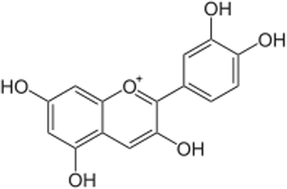
|
Grapes, bilberry, blackberry, blueberry, cherry, cranberry, elderberry, hawthorn, logan berry, acai berry and raspberry. | ↑ pAMPK, pACC signaling and improve insulin signaling (pAkt, pFOXO-1). | HFD-induced obesity rats | Park 2015 [206] |
| ↑ PGC-1α, SIRT1 and UCP-3 genes. | 3 T3-Ll cells | Matsukawa 2015 [207] | ||
| Lowered fasting glucose and improved insulin sensitivity. | C57BL/6 J obese mice | Guo 2012 [205] | ||
| Decreased c-Jun N-terminal kinase activation and FoXO1. | ||||
| Upregulated the GLUT4 and down-regulation of the inflammatory adipocytokines. | HFD-KK-A(y) mice | Sasaki 2007 [208] | ||
| Suppressed the mRNA levels of enzymes involved in FA and TG synthesis and lowered the SREBP-1 level. | High fat-induced diabetic mice | Tsuda 2003 [209] | ||
| ↓ Glucose, mitochondrial (ROS) | INS-1 cells and STZ-induced diabetic mice | Sun 2012 [210] | ||
Delphinidin 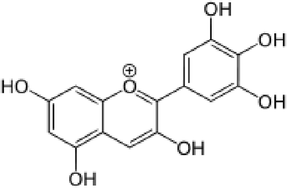
|
Berries, dark grapes and vegetables such as eggplant, tomato, carrot, purple sweet potato, red cabbage and red onion | ↓ Albumin and HbA1c glycation. | Diabetic rats | Gharib 2013 [212] |
| Cyclooxygenase inhibitor restored the relaxant responses to Ach and SNP. | Diabetic microangiopathy. | Bertuglia 1995 [211] | ||
Pelargonidin 
|
Ficus bengalensis Linn and billberry | ↓ Glucose, TBARS and ↑ SOD | STZ-injected diabetic rats | Mirshekar 2010 [215] |
| Improved retention and recall capability. | STZ-diabetic rats | Mirshekar 2011 [217] |
Cyanidin
Cyanidin and its glycosides belong to anthocyanins, and are widely distributed in various human diets through crops, vegetables, fruits, and red wine suggesting that we daily ingest significant amounts of these compounds from plant-based diets. Cyanidin has been demonstrated to inhibit intestinal α-glucosidase and pancreatic α-amylase, which is one of the therapeutic approaches for treatment of DM [199]. Anthocyanin was also found to reverse degenerative changes in β-cells in STZ-induced diabetic rats by activating insulin receptor phosphorylation and preventing pancreatic apoptosis [200]. Nasri et al. [201] performed in vivo chronic treatment of diabetic rats with cyanidin-3-glucoside (C3G) could prevent the functional changes in vascular reactivity observed in diabetic rats through endothelium dependent pathways and via attenuation of aortic lipid peroxidation. C3G, one of the most prevalent anthocyanins existing in our diet, can protect hepatocytes against HG-induced damage by improving antioxidant status and inhibiting the mitochondria-mediated apoptotic pathway through activation of Akt and inactivation of JNK [202, 203] and another author reported that anthocyanins exert a hepatoprotective effect against hyperglycemia-accelerated steatohepatitis in NAFLD [204]. Moreover Guo et al. [205] showed that C3G significantly reduced macrophage infiltration and the mRNA levels of MCP-1, TNF-α and IL-6 in adipose tissue and phosphorylation of FoxO1 via the Akt-dependent pathway, and the extent of phosphorylation represents the FoxO1 transcriptional activity in liver and adipose tissues of HFD and db/db mice.
Delphinidin
Delphinidin present in pigmented fruits such as pomegranate, berries, dark grapes and vegetables such as eggplant, tomato, carrot, purple sweet potato, red cabbage and red onion and it possesses strong antioxidant activities. Delphinidin was observed in vivo at the microcirculatory level prevent the injury to endothelial cell function associated with diabetes and/or oxidative stress [211]. Moreover, administration of 100 mg/kg delphinidin chloride-loaded liposomes to diabetic mice at 8 weeks could decrease the rate of albumin and HbA1c glycation [212].
Pelargonidin
Pelargonidin can be found in berries such as ripe raspberries, blueberries, blackberries, cranberries and saskatoon berries [213] Pelargonidin treatment counteracts hyperglycemia and relieves the oxidative stress including hemoglobin (Hb) induced iron mediated oxidative reactions by lowering the glycation level and free iron of Hb [214]. Pelargonidin was also demonstrated to reduce TBARS formation and non-significantly reversed elevation of nitrite level and reduction of antioxidant defensive enzyme superoxide dismutase in diabetic rats [215]. Pelargonidin-3-galactoside and its aglycone stimulate insulin secretion in rodent pancreatic β-cells in vitro in presence of glucose [216].
Conclusions
The actual antidiabetic prospective associated with flavonoids are usually large as a result of their modulatory effects on blood sugar transporter by enhancing insulin secretion, reducing apoptosis and promoting proliferation of pancreatic β-cells, reducing insulin resistance, inflammation and oxidative stress in muscle and promoting translocation of GLUT4 via PI3K/AKT and AMPK pathways (Fig. 1). The molecular mechanisms underlying the glucose and lipid metabolism in diabetes would provide new insights in the field of drug development, continue to fuel excitement in this area of research and buoys the hope that future discoveries may one day yield therapeutic benefits. With the rapidly increasing incidence of diabetes worldwide, there is a greater need for safe and effective functional biomaterials with antidiabetic activity. Hence, meticulously intended human studies are needed to further measure the likely connected with a number of nutritional flavonoids to treat diabetes and its complication.
Fig. 1.
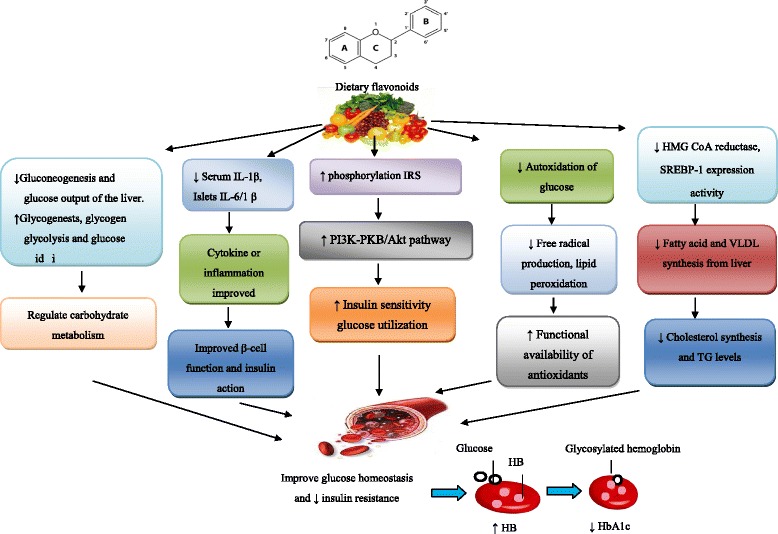
Schematic of the proposed role of flavonoids on management of blood glucose in diabetes. AKT; v-akt murine thymoma viral oncogene homolog, IRS; Insulin receptor substrate, HB; hemoglobin, HbA1c; Glycated hemoglobin, HMG-CoA :3-hydroxy-3-methylglutaryl-coenzyme A, IL-1β; Interleukin-1 beta, PI3K; Phosphatidylinositol-3-kinase, SREBP-1c; Sterol regulatory element-binding protein, TG; Triglycerides, VLDL; Very low density lipoprotein, (↑ Increase, ↓ Decrease)
Acknowledgments
This review work was financially supported by two research grants (R201327 and R201402) from Beijing Normal University-Hong Kong Baptist University United International College, China.
Abbreviations
- AGEs
advanced glycation end products
- AKT
v-akt murine thymoma viral oncogene homolog
- AMPK
5′AMP-activated protein kinase
- CAT
catalase
- DM
diabetes mellitus
- eNOS
endothelial nitric oxide synthase
- FFA
free fatty acid
- TG
triglycerides
- GPx
glutathione peroxidase
- GR
glutathione reductase
- GST
glutathione S-transferase
- HB
hemoglobin
- HbA1c
glycated hemoglobin
- HDL
high density lipoprotein
- HFD
high-fat diet
- HG
high glucose
- HMG-CoA
3-hydroxy-3-methylglutaryl-coenzyme A
- ICAM-1
intercellular adhesion molecule 1
- IL-1β
interleukin-1 beta
- iNOS
inducible nitric oxide synthase
- IR
insulin resistance
- IRS-1
insulin receptor substrate-1
- IRS-2
insulin receptor substrate-2
- LDL
low density lipoprotein
- MAPK
mitogen activated protein kinase
- NF-κB
nuclear factor kappa B
- NO
nitric oxide
- PI3K
phosphatidylinositol-3-kinase
- PPARγ
peroxisome proliferator-activated receptor gamma
- ROS
reactive oxygen species
- SOD
superoxide dismutase
- SREBP-1c
sterol regulatory element-binding protein
- STZ
streptozotocin
- TNFα
tumor necrosis factor α
- VEGF
vascular endothelial growth factor
- VLDL
very low density lipoprotein
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
VR wrote the manuscript and BX has been involved in designing, drafting the manuscript and revising it critically. VR and BX have given final approval of the version to be manuscript.
References
- 1.Devendra D, Liu E, Eisenbarth GS. Type 1diabetes: Recent developments. BMJ. 2004;328:750–54. doi: 10.1136/bmj.328.7442.750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Fowler MJ. Microvascular and macrovascular complications of diabetes. Clin Diabetes. 2008;26:2. [Google Scholar]
- 3.International Diabetes Federation, Sixth edition. 2014.
- 4.Kumar R, Laloo D, Hemalatha S. Diabetes mellitus: An overview on its pharmacological aspects and reported medicinal plants having antidiabetic activity. Asian Pac J Trop Biomed. 2012;2:411–20. doi: 10.1016/S2221-1691(12)60067-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Surveswaran S, Zhong Cai Y, Corke H, Sun M. Systematic evaluation of natural phenolic antioxidants from 133 Indian medicinal plants. Food Chem. 2007;102:938–53. doi: 10.1016/j.foodchem.2006.06.033. [DOI] [Google Scholar]
- 6.Burgering BM, Coffer PJ. Protein kinase B (c-Akt) in phosphatidylinositol-3-OH kinase signal transduction. Nature. 1995;376:599–602. doi: 10.1038/376599a0. [DOI] [PubMed] [Google Scholar]
- 7.Weickert MO, Piffer AFH. Signalling mechanisms linking hepatic glucose and lipid metabolism. Diabetologia. 2006;49:1732–41. doi: 10.1007/s00125-006-0295-3. [DOI] [PubMed] [Google Scholar]
- 8.Kile BT, Schulman BA, Alexander WS, Nicola NA, Martin HM, Hilton DJ. The SOCS box: A tale of destruction and degradation. Trends Biochem Sci. 2002;27:235–41. doi: 10.1016/S0968-0004(02)02085-6. [DOI] [PubMed] [Google Scholar]
- 9.Schinner S, Scherbaum WA, Bornstein SR, Barthel A. Molecular mechanisms of insulin resistance. Diabet Med. 2005;22:674–82. doi: 10.1111/j.1464-5491.2005.01566.x. [DOI] [PubMed] [Google Scholar]
- 10.Kaszubska W, Falls HD, Schaefer VG, Haasch D, Frost L, Hessler P, et al. Protein tyrosine phosphatase 1B negatively regulates leptin signalling in a hypothalamic cell line. Mol Cell Endocrinol. 2002;195:109–18. doi: 10.1016/S0303-7207(02)00178-8. [DOI] [PubMed] [Google Scholar]
- 11.Clement S, Krause U, Desmedt F, Tanti JF, Behrends J, Pesesse X, et al. The lipid phosphatase SHIP2 controls insulin sensitivity. Nature. 2001;409:92–7. doi: 10.1038/35051094. [DOI] [PubMed] [Google Scholar]
- 12.Butler M, McKay RA, Popoff IJ, Gaarde WA, Witchell D, Murray SF, et al. Specific inhibition of PTEN expression reverses hyperglycemia in diabetic mice. Diabetes. 2002;51:1028–34. doi: 10.2337/diabetes.51.4.1028. [DOI] [PubMed] [Google Scholar]
- 13.Tyagi S, Gupta P, Saini AS, Kaushal C, Sharma S. The peroxisome proliferator activated receptor: A family of nuclear receptors role in various diseases. J Adv Pharm Technol Res. 2011;2:236–40. doi: 10.4103/2231-4040.90879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Furuhashi M, Hotamisligil GS. Fatty acid-binding proteins: Role in metabolic diseases and potential as drug targets. Nat Rev Drug Discov. 2008;7:489–503. doi: 10.1038/nrd2589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Richard AJ, Amini-Vaughan Z, Ribnicky DM, Stephens JM. Naringenin inhibits adipogenesis and reduces insulin sensitivity and adiponectin expression in adipocytes. Evid Based Complement Alternat Med. 2013;2013:549750. doi: 10.1155/2013/549750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Makki K, Froguel P, Wolowczuk I. Adipose tissue in obesity-related inflammation and insulin resistance: Cells, cytokines, and chemokines. ISRN Inflamm. 2013;2013:139239. doi: 10.1155/2013/139239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Moon HS, Dalamaga M, Kim SY, Polyzos SA, Hamnvik OP, Magkos F, et al. Leptin’s role in lipodystrophic and nonlipodystrophic insulin-resistant and diabetic individuals. Endocr Rev. 2013;34:377–412. doi: 10.1210/er.2012-1053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Jung UJ, Choi MS. Obesity and its metabolic complications: The role of adipokines and the relationship between obesity, inflammation, insulin resistance, dyslipidemia and nonalcoholic fatty liver disease. Int J Mol Sci. 2014;15:6184–223. doi: 10.3390/ijms15046184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Arts IC, Hollman PC. Polyphenols and disease risk in epidemiologic studies. Am J Clin Nutr. 2005;81:317S–25S. doi: 10.1093/ajcn/81.1.317S. [DOI] [PubMed] [Google Scholar]
- 20.Duncan BB, Schmidt MI, Pankow JS, Ballantyne CM, Couper D, Vigo A, et al. Low-grade systemic inflammation and the development of type 2 diabetes: The atherosclerosis risk in communities study. Diabetes. 2003;52:1799–805. doi: 10.2337/diabetes.52.7.1799. [DOI] [PubMed] [Google Scholar]
- 21.Pickup JC. Inflammation and activated innate immunity in the pathogenesis of type 2 diabetes. Diabetes Care. 2004;27:813–23. doi: 10.2337/diacare.27.3.813. [DOI] [PubMed] [Google Scholar]
- 22.Rice-Evans CA, Miller NJ, Paganga G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic Biol Med. 1996;20:933–56. doi: 10.1016/0891-5849(95)02227-9. [DOI] [PubMed] [Google Scholar]
- 23.Kandaswami C, Middleton E., Jr Free radical scavenging and antioxidant activity of plant flavonoids. Adv Exp Med Biol. 1994;366:351–76. doi: 10.1007/978-1-4615-1833-4_25. [DOI] [PubMed] [Google Scholar]
- 24.Pandey KB, Rizvi SI. Plant polyphenols as dietary antioxidants in human health and disease. Oxid Med Cell Longev. 2009;2:270–8. doi: 10.4161/oxim.2.5.9498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bahadoran Z, Mirmiran P, Azizi F. Dietary polyphenols as potential nutraceuticals in management of diabetes: A review. J Diabetes Metab Disord. 2013;12:43. doi: 10.1186/2251-6581-12-43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hanhineva K, Törrönen R, Bondia-Pons I, Pekkinen J, Kolehmainen M, Mykkänen H, et al. Impact of dietary polyphenols on carbohydrate metabolism. Int J Mol Sci. 2010;11:1365–402. doi: 10.3390/ijms11041365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hajiaghaalipour F, Khalilpourfarshbafi M, Arya A. Modulation of glucose transporter protein by dietary flavonoids in type 2 diabetes mellitus. Int J Biol Sci. 2015;11:508–24. doi: 10.7150/ijbs.11241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Campanero MA, Escolar M, Perez G, Garcia-Quetglas E, Sadaba B, Azanza JR. Simultaneous determination of diosmin and diosmetin in human plasma by ion trap liquid chromatography-atmospheric pressure chemical ionization tandem mass spectrometry: application to a clinical pharmacokinetic study. J Pharm Biomed Anal. 2010;51:875–881. doi: 10.1016/j.jpba.2009.09.012. [DOI] [PubMed] [Google Scholar]
- 29.Manuel Y, Keenoy B, Vertommen J, De Leeuw I. The effect of flavonoid treatment on the glycation and antioxidant status in type I diabetic patients. Diabetes Nutr Metab. 1999;12:256–63. [PubMed] [Google Scholar]
- 30.Pari L, Srinivasan S. Antihyperglycemic effect of diosmin on hepatic key enzymes of carbohydrate metabolism in streptozotocin-nicotinamide-induced diabetic rats. Biomed Pharmacother. 2010;64:477–81. doi: 10.1016/j.biopha.2010.02.001. [DOI] [PubMed] [Google Scholar]
- 31.Srinivasan S, Pari L. Antihyperlipidemic effect of diosmin: A citrus flavonoid on lipid metabolism in experimental diabetic rats. J Funct Foods. 2012;5:484–92. doi: 10.1016/j.jff.2012.12.004. [DOI] [Google Scholar]
- 32.Jain D, Bansal MK, Dalvi R, Upganlawar A, Somani R. Protective effect of diosmin against diabetic neuropathy in experimental rats. J Integr Med. 2014;12:35–41. doi: 10.1016/S2095-4964(14)60001-7. [DOI] [PubMed] [Google Scholar]
- 33.Srinivasan S, Pari L. Ameliorative effect of diosmin, a citrus flavonoid against streptozotocin-nicotinamide generated oxidative stress induced diabetic rats. Chem Biol Interact. 2012;195:43–51. doi: 10.1016/j.cbi.2011.10.003. [DOI] [PubMed] [Google Scholar]
- 34.Vertommen J, van den Enden M, Simoens L, de Leeuw I. Flavonoid treatment reduces glycation and lipid peroxidation in experimental diabetic rats. Phytother Res. 1994;8:430–32. doi: 10.1002/ptr.2650080711. [DOI] [Google Scholar]
- 35.Du HQ. Isolation and identification of fisetin in Cotinus coggygria. Zhong Yao Tong Bao. 1983;8:29–30. [PubMed] [Google Scholar]
- 36.Arai Y, Watanabe S, Kimira M, Shimoi K, Mochizuki R, Kinae N. Dietary intakes of flavonols, flavones and isoflavones by Japanese women and the inverse correlation between quercetin intake and plasma LDL cholesterol concentration. J Nutr. 2000;130:2243–50. doi: 10.1093/jn/130.9.2243. [DOI] [PubMed] [Google Scholar]
- 37.Constantin RP, Constantin J, Pagadigorria CL, Ishii-Iwamoto EL, Bracht A, Ono Mde K, et al. The actions of fisetin on glucose metabolism in the rat liver. Cell Biochem Funct. 2010;28:149–58. doi: 10.1002/cbf.1635. [DOI] [PubMed] [Google Scholar]
- 38.Prasath GS, Pillai S, Subramanian SP. Fisetin improves glucose homeostasis through the inhibition of gluconeogenic enzymes in hepatic tissues of streptozotocin induced diabetic rats. Eur J Pharmacol. 2014;740:248–54. doi: 10.1016/j.ejphar.2014.06.065. [DOI] [PubMed] [Google Scholar]
- 39.Prasath GS, Sundaram CS, Subramanian SP. Fisetin averts oxidative stress in pancreatic tissues of streptozotocin-induced diabetic rats. Endocrine. 2013;44:359–68. doi: 10.1007/s12020-012-9866-x. [DOI] [PubMed] [Google Scholar]
- 40.Kim HJ, Kim SH, Yun JM. Fisetin inhibits hyperglycemia-induced proinflammatory cytokine production by epigenetic mechanisms. Evid Based Complement Alternat Med. 2012;2012:639469. doi: 10.1155/2012/639469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kan E, Kiliçkan E, Ayar A, Colak R. Effects of two antioxidants; α-lipoic acid and fisetin against diabetic cataract in mice. Int Ophthalmol. 2015;35:115–20. doi: 10.1007/s10792-014-0029-3. [DOI] [PubMed] [Google Scholar]
- 42.Prasath GS, Subramanian SP. Antihyperlipidemic effect of fisetin, a bioflavonoid of strawberries, studied in streptozotocin-induced diabetic rats. J Biochem Mol Toxicol. 2014;28:442–9. doi: 10.1002/jbt.21583. [DOI] [PubMed] [Google Scholar]
- 43.Kwak S, Ku SK, Bae JS. Fisetin inhibits high-glucose-induced vascular inflammation in vitro and in vivo. Inflamm Res. 2014;63:779–87. doi: 10.1007/s00011-014-0750-4. [DOI] [PubMed] [Google Scholar]
- 44.Prasath GS, Subramanian SP. Fisetin, a tetra hydroxy flavone recuperates antioxidant status and protects hepatocellular ultrastructure from hyperglycemia mediated oxidative stress in streptozotocin induced experimental diabetes in rats. Food Chem Toxicol. 2013;59:249–55. doi: 10.1016/j.fct.2013.05.062. [DOI] [PubMed] [Google Scholar]
- 45.Prasath GS, Subramanian SP. Modulatory effects of fisetin, a bioflavonoid, on hyperglycemia by attenuating the key enzymes of carbohydrate metabolism in hepatic and renal tissues in streptozotocin-induced diabetic rats. Eur J Pharmacol. 2011;668:492–6. doi: 10.1016/j.ejphar.2011.07.021. [DOI] [PubMed] [Google Scholar]
- 46.Maher P, Dargusch R, Ehren JL, Okada S, Sharma K, Schubert D. Fisetin lowers methylglyoxal dependent protein glycation and limits the complications of diabetes. PLoS One. 2011;6:e21226. doi: 10.1371/journal.pone.0021226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Ricardo KFS, Toledo de Oliveira T, Nagem TJ, Pinto AS, Oliveira MGA, Soares JF. Effect of flavonoids morin; quercetin and nicotinic acid on lipid metabolism of rats experimentally fed with triton. Braz Arch Biol Technol. 2001;44:263–67. doi: 10.1590/S1516-89132001000300007. [DOI] [Google Scholar]
- 48.Sreedharan V, Venkatachalam KK, Namasivayam N. Effect of morin on tissue lipid peroxidation and antioxidant status in 1, 2-dimethylhydrazine induced experimental colon carcinogenesis. Invest New Drugs. 2009;27:21–30. doi: 10.1007/s10637-008-9136-1. [DOI] [PubMed] [Google Scholar]
- 49.Sendrayaperumal V, Iyyam Pillai S, Subramanian S. Design, synthesis and characterization of zinc-morin, a metal flavonol complex and evaluation of its antidiabetic potential in HFD-STZ induced type 2 diabetes in rats. Chem Biol Interact. 2014;219:9–17. doi: 10.1016/j.cbi.2014.05.003. [DOI] [PubMed] [Google Scholar]
- 50.Abuohashish HM, Al-Rejaie SS, Al-Hosaini KA, Parmar MY, Ahmed MM. Alleviating effects of morin against experimentally-induced diabetic osteopenia. Diabetol Metab Syndr. 2013;5:5. doi: 10.1186/1758-5996-5-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Wang X, Zhang DM, Gu TT, Ding XQ, Fan CY, Zhu Q, et al. Morin reduces hepatic inflammation-associated lipid accumulation in high fructose-fed rats via inhibiting sphingosine kinase 1/sphingosine 1-phosphate signaling pathway. Biochem Pharmacol. 2013;86:1791–804. doi: 10.1016/j.bcp.2013.10.005. [DOI] [PubMed] [Google Scholar]
- 52.Paoli P, Cirri P, Caselli A, Ranaldi F, Bruschi G, Santi A, et al. The insulin-mimetic effect of morin: A promising molecule in diabetes treatment. Biochim Biophys Acta. 2013;30:3102–11. doi: 10.1016/j.bbagen.2013.01.017. [DOI] [PubMed] [Google Scholar]
- 53.Vanitha P, Uma C, Suganya N, Bhakkiyalakshmi E, Suriyanarayanan S, Gunasekaran P, et al. Modulatory effects of morin on hyperglycemia by attenuating the hepatic key enzymes of carbohydrate metabolism and β-cell function in streptozotocin-induced diabetic rats. Environ Toxicol Pharmacol. 2014;37:326–35. doi: 10.1016/j.etap.2013.11.017. [DOI] [PubMed] [Google Scholar]
- 54.Kapoor R, Kakkar P. Protective role of morin, a flavonoid, against high glucose induced oxidative stress mediated apoptosis in primary rat hepatocytes. PLoS One. 2012;7:e41663. doi: 10.1371/journal.pone.0041663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Miyake Y, Yamamoto K, Tsujihara N, Osawa T. Protective effects of lemon flavonoids on oxidative stress in diabetic rats. Lipids. 1998;33:689–95. doi: 10.1007/s11745-998-0258-y. [DOI] [PubMed] [Google Scholar]
- 56.Zhang WY, Lee JJ, Kim Y, Kim IS, Han JH, Lee SG, et al. Effect of eriodictyol on glucose uptake and insulin resistance in vitro. J Agric Food Chem. 2012;60:7652–8. doi: 10.1021/jf300601z. [DOI] [PubMed] [Google Scholar]
- 57.Bucolo C, Leggio GM, Drago F, Salomone S. Eriodictyol prevents early retinal and plasma abnormalities in streptozotocin-induced diabetic rats. Biochem Pharmacol. 2012;84:88–92. doi: 10.1016/j.bcp.2012.03.019. [DOI] [PubMed] [Google Scholar]
- 58.Emim JA, Oliveira AB, Lapa AJ. Pharmacological evaluation of the anti-inflammatory activity of a citrus bioflavonoid, hesperidin, and the isoflavonoids, duartin and claussequinone, in rats and mice. J Pharm Pharmacol. 1994;46:118–22. doi: 10.1111/j.2042-7158.1994.tb03753.x. [DOI] [PubMed] [Google Scholar]
- 59.Kawaguchi K, Mizuno T, Aida K, Uchino K. Hesperidin as an inhibitor of lipases from porcine pancreas and Pseudomonas. Biosci Biotechnol Biochem. 1997;61:102–4. doi: 10.1271/bbb.61.102. [DOI] [PubMed] [Google Scholar]
- 60.Visnagri A, Kandhare AD, Chakravarty S, Ghosh P, Bodhankar SL. Hesperidin, a flavanoglycone attenuates experimental diabetic neuropathy via modulation of cellular and biochemical marker to improve nerve functions. Pharm Biol. 2014;52:814–28. doi: 10.3109/13880209.2013.870584. [DOI] [PubMed] [Google Scholar]
- 61.Gumieniczek A. Effect of the new thiazolidinedione-pioglitazone on the development of oxidative stress in liver and kidney of diabetic rabbits. Life Sci. 2003;74:553–62. doi: 10.1016/j.lfs.2003.03.004. [DOI] [PubMed] [Google Scholar]
- 62.Shi X, Liao S, Mi H, Guo C, Qi D, Li F, et al. Hesperidin prevents retinal and plasma abnormalities in streptozotocin-induced diabetic rats. Molecules. 2012;17:12868–81. doi: 10.3390/molecules171112868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Akiyama S, Katsumata S, Suzuki K, Ishimi Y, Wu J, Uehara M. Dietary hesperidin exerts hypoglycemic and hypolipidemic effects in streptozotocin-induced marginal type 1 diabetic rats. J Clin Biochem Nutr. 2010;46:87–92. doi: 10.3164/jcbn.09-82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Yo A, Sharma PK, Shrivastava B, Ojha S, Upadhya HM, Arya DS, et al. Hesperidin produces cardioprotective activity via PPAR-γ pathway in ischemic heart disease model in diabetic rats. PLoS One. 2014;9:e111212. doi: 10.1371/journal.pone.0111212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Mahmoud AM, Ashour MB, Abdel-Moneim A, Ahmed OM. Hesperidin and naringin attenuate hyperglycemia-mediated oxidative stress and proinflammatory cytokine production in high fat fed/streptozotocin-induced type 2 diabetic rats. J. Diabetes Complicat. 2012;26:483–90. doi: 10.1016/j.jdiacomp.2012.06.001. [DOI] [PubMed] [Google Scholar]
- 66.Kakadiya J, Mulani H, Shah N. Protective effect of hesperidin on cardiovascular complication in experimentally induced myocardial infarction in diabetes in rats. J Basic Clin Pharm. 2010;1:85–91. [PMC free article] [PubMed] [Google Scholar]
- 67.Akiyama S, Katsumata S, Suzuki K, Nakaya Y, Ishimi Y, Uehara M. Hypoglycemic and hypolipidemic effects of hesperidin and cyclodextrin-clathrated hesperetin in Goto-Kakizaki rats with type 2 diabetes. Biosci Biotechnol Biochem. 2009;73:2779–82. doi: 10.1271/bbb.90576. [DOI] [PubMed] [Google Scholar]
- 68.Jung UJ, Lee MK, Jeong KS, Choi MS. The hypoglycemic effects of hesperidin and naringin are partly mediated by hepatic glucose-regulating enzymes in C57BL/KsJ-db/db mice. J Nutr. 2004;134:2499–503. doi: 10.1093/jn/134.10.2499. [DOI] [PubMed] [Google Scholar]
- 69.Ashafaq M, Varshney L, Khan MH, Salman M, Naseem M, Wajid S, et al. Neuromodulatory effects of hesperidin in mitigating oxidative stress in streptozotocin induced diabetes. Biomed Res Int. 2014;249031:9. doi: 10.1155/2014/249031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Wilcox LJ, Borradaile NM, Huff MW. Antiatherogenic properties of naringenin, a citrus flavonoid. Cardiovasc Drug Rev. 1999;17:160–78. doi: 10.1111/j.1527-3466.1999.tb00011.x. [DOI] [Google Scholar]
- 71.Van Acker FA, Schouten O, Haenen GR, van der Vijgh WJ, Bast A. Flavonoids can replace alpha-tocopherol as an antioxidant. FEBS Lett. 2000;473:145–8. doi: 10.1016/S0014-5793(00)01517-9. [DOI] [PubMed] [Google Scholar]
- 72.Sanchez-Salgado JC, Ortiz-Andrade RR, Aguirre-Crespo F, Vergara-Galicia J, Leon-Rivera I, Montes S, et al. Hypoglycemic, vasorelaxant and hepatoprotective effects of Cochlospermum vitifolium (willd.) sprengel: A potential agent for the treatment of metabolic syndrome. J Ethnopharmacol. 2007;109:400–5. doi: 10.1016/j.jep.2006.08.008. [DOI] [PubMed] [Google Scholar]
- 73.Priscilla DH, Roy D, Suresh A, Kumar V, Thirumurugan K. Naringenin inhibits α-glucosidase activity: A promising strategy for the regulation of postprandial hyperglycemia in high fat diet fed streptozotocin induced diabetic rats. Chem Biol Interact. 2014;210:77–85. doi: 10.1016/j.cbi.2013.12.014. [DOI] [PubMed] [Google Scholar]
- 74.Li JM, Che CT, Lau CBS, Leung PS, Cheng CHK. Inhibition of intestinal andrenal Na+-glucose co-transporter by naringenin. Int J Biochem Cell Biol. 2006;38:985–95. doi: 10.1016/j.biocel.2005.10.002. [DOI] [PubMed] [Google Scholar]
- 75.Fallahi F, Roghani M, Moghadami S. Citrus flavonoid naringenin improves aortic reactivity in streptozotocin-diabetic rats. Indian J Pharmacol. 2012;44:382–96. doi: 10.4103/0253-7613.96350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Pu P, Gao DM, Mohamed S, Chen J, Zhang J, Zhou XY, et al. Naringin ameliorates metabolic syndrome by activating AMP-activated protein kinase in mice fed a high-fat diet. Arch Biochem Biophys. 2012;518:61–70. doi: 10.1016/j.abb.2011.11.026. [DOI] [PubMed] [Google Scholar]
- 77.Zygmunt K, Faubert B, MacNeil J, Tsiani E. Naringenin, a citrus flavonoid, increases muscle cell glucose uptake via AMPK. Biochem Biophys Res Commun. 2010;398:178–83. doi: 10.1016/j.bbrc.2010.06.048. [DOI] [PubMed] [Google Scholar]
- 78.Choi JS, Yokozawa T, Oura H. Improvement of hyperglycemia, hyperlipemiain streptozotocin-diabetic rats by a methanolic extract of Prunus davidiana stems, its main component. Prunin Planta Med. 1991;57:208–11. doi: 10.1055/s-2006-960075. [DOI] [PubMed] [Google Scholar]
- 79.Kannappan S, Anuradha CV. Naringenin enhances insulin-stimulated tyrosine phosphorylation and improves the cellular actions of insulin in a dietary model of metabolic syndrome. Eur J Nutr. 2010;49:101–9. doi: 10.1007/s00394-009-0054-6. [DOI] [PubMed] [Google Scholar]
- 80.Mulvihill EE, Allister EM, Sutherland BG, Telford DE, Sawyez CG, Edwards JY, et al. Naringenin prevents dyslipidemia, apolipoprotein B overproduction, and hyperinsulinemia in LDL receptor-null mice with diet-induced insulin resistance. Diabetes. 2009;58:2198–10. doi: 10.2337/db09-0634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Priscilla DH, Jayakumar M, Thirumurugan K. Flavanone naringenin: An effective antihyperglycemic and antihyperlipidemic nutraceutical agent on high fat diet fed streptozotocin induced type 2 diabetic rats. J Funct Foods. 2015;14:363–73. doi: 10.1016/j.jff.2015.02.005. [DOI] [Google Scholar]
- 82.Yoshida H, Watanabe H, Ishida A, Watanabe W, Narumi K, Atsumi T, et al. Naringenin suppresses macrophage infiltration into adipose tissue in an early phase of high-fat diet-induced obesity. Biochem Biophys Res Commun. 2014;454:95–101. doi: 10.1016/j.bbrc.2014.10.038. [DOI] [PubMed] [Google Scholar]
- 83.Hasanein P, Fazeli F. Role of naringenin in protection against diabetic hyperalgesia and tactile allodynia in male Wistar rats. J Physiol Biochem. 2014;70:997–1006. doi: 10.1007/s13105-014-0369-5. [DOI] [PubMed] [Google Scholar]
- 84.Bhattacharya S, Oksbjerg N, Young JF, Jeppesen PB. Caffeic acid, naringenin and quercetin enhance glucose-stimulated insulin secretion and glucose sensitivity in INS-1E cells. Diabetes Obes Metab. 2014;16:602–12. doi: 10.1111/dom.12236. [DOI] [PubMed] [Google Scholar]
- 85.Yoshida H, Watanabe W, Oomagari H, Tsuruta E, Shida M, Kurokawa M. Citrus flavonoid naringenin inhibits TLR2 expression in adipocytes. J Nutr Biochem. 2013;24:1276–84. doi: 10.1016/j.jnutbio.2012.10.003. [DOI] [PubMed] [Google Scholar]
- 86.Rahigude A, Bhutada P, Kaulaskar S, Aswar M, Otari K. Participation of antioxidant and cholinergic system in protective effect of naringenin against type-2 diabetes-induced memory dysfunction in rats. Neuroscience. 2012;226:62–72. doi: 10.1016/j.neuroscience.2012.09.026. [DOI] [PubMed] [Google Scholar]
- 87.Ross JA, Kasum CM. Dietary flavonoids: Bioavailability, metabolic effects, and safety. Annu Rev Nutr. 2002;22:19–34. doi: 10.1146/annurev.nutr.22.111401.144957. [DOI] [PubMed] [Google Scholar]
- 88.Panda S, Kar A. Apigenin (4',5,7-trihydroxyflavone) regulates hyperglycaemia, thyroid dysfunction and lipid peroxidation in alloxan-induced diabetic mice. J Pharm Pharmacol. 2007;59:1543–8. doi: 10.1211/jpp.59.11.0012. [DOI] [PubMed] [Google Scholar]
- 89.Mrazek AA, Porro LJ, Bhatia V, Falzon M, Spratt H, Zhou J, et al. Apigenin inhibits pancreatic stellate cell activity in pancreatitis. J Surg Res. 2015;196:8–16. doi: 10.1016/j.jss.2015.02.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Suh KS, Oh S, Woo JT, Kim SW, Kim JW, Kim YS, et al. Apigenin attenuates 2-deoxy-D-ribose-induced oxidative cell damage in HIT-T15 pancreatic β-cells. Biol Pharm Bull. 2012;35:121–6. doi: 10.1248/bpb.35.121. [DOI] [PubMed] [Google Scholar]
- 91.Hossain CM, Ghosh MK, Satapathy BS, Dey NS, Mukherjee B. Apigenin causes biochemical modulation, GLUT4 and Cd38 alterations to improve diabetes and to protect damages of some vital organs in experimental diabetes. Am J Pharmacol Toxicol. 2014;9:39–52. doi: 10.3844/ajptsp.2014.39.52. [DOI] [Google Scholar]
- 92.Zhang X, Wang G, Gurley EC, Zhou H. Flavonoid apigenin inhibits lipopolysaccharide-induced inflammatory response through multiple mechanisms in macrophages. PLoS One. 2014;9:e107072. doi: 10.1371/journal.pone.0107072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Yamagata K, Miyashita A, Matsufuji H, Chino M. Dietary flavonoid apigenin inhibits high glucose and tumor necrosis factor alpha-induced adhesion molecule expression in human endothelial cells. J Nutr Biochem. 2010;21:116–24. doi: 10.1016/j.jnutbio.2008.11.003. [DOI] [PubMed] [Google Scholar]
- 94.Cazarolli LH, Folador P, Moresco HH, Brighente IM, Pizzolatti MG, Silva FR. Mechanism of action of the stimulatory effect of apigenin-6-C-(2''-O-alpha-l-rhamnopyranosyl)-beta-L-fucopyranoside on 14C-glucose uptake. Chem Biol Interact. 2009;179:407–12. doi: 10.1016/j.cbi.2008.11.012. [DOI] [PubMed] [Google Scholar]
- 95.Kim YO, Leem K, Park J, Lee P, Ahn DK, Lee BC, et al. Cytoprotective effect of Scutellaria baicalensis in CA1 hippocampal neurons of rats after global cerebral ischemia. J Ethnopharmacol. 2001;77:183–88. doi: 10.1016/S0378-8741(01)00283-5. [DOI] [PubMed] [Google Scholar]
- 96.Lapchak PA, Maher P, Schubert D, Zivin JA. Baicalein, an antioxidant12/15-lipoxygenase inhibitor improves clinical rating scores following multiple infarct embolic strokes. Neuroscience. 2007;150:585–91. doi: 10.1016/j.neuroscience.2007.09.033. [DOI] [PubMed] [Google Scholar]
- 97.Fu Y, Luo J, Jia Z, Zhen W, Zhou K, Gilbert E, et al. Baicalein protects against type 2 diabetes via promoting islet β-cell function in obese diabetic mice. Int J Endocrinol. 2014;2014:846742. doi: 10.1155/2014/846742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Stavniichuk R, Drel VR, Shevalye H, Maksimchyk Y, Kuchmerovska TM, Nadler JL, et al. Baicalein alleviates diabetic peripheral neuropathy through inhibition of oxidative-nitrosative stress and p38 MAPK activation. Exp Neurol. 2011;230:106–13. doi: 10.1016/j.expneurol.2011.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Ahad A, Mujeeb M, Ahsan H, Siddiqui WA. Prophylactic effect of baicalein against renal dysfunction in type 2 diabetic rats. Biochimie. 2014;106:101–10. doi: 10.1016/j.biochi.2014.08.006. [DOI] [PubMed] [Google Scholar]
- 100.El-Bassossy HM, Hassan NA, Mahmoud MF, Fahmy A. Baicalein protects against hypertension associated with diabetes: Effect on vascular reactivity and stiffness. Phytomedicine. 2014;21:1742–45. doi: 10.1016/j.phymed.2014.08.012. [DOI] [PubMed] [Google Scholar]
- 101.Pu P, Wang XA, Salim M, Zhu LH, Wang L, Chen KJ, et al. Baicalein, a natural product, selectively activating AMPKα(2) and ameliorates metabolic disorder in diet-induced mice. Mol Cell Endocrinol. 2012;362:128–38. doi: 10.1016/j.mce.2012.06.002. [DOI] [PubMed] [Google Scholar]
- 102.Qi Z, Xu Y, Liang Z, Li S, Wang J, Wei Y, et al. Baicalein alters PI3K/Akt/GSK3β signaling pathway in rats with diabetes-associated cognitive deficits. Int J Clin Exp Med. 2015;8:1993–2000. [PMC free article] [PubMed] [Google Scholar]
- 103.Siess MH, LeBon AM, Canivenc-Laver MC, Amiot MJ, Sabatier S, Aubert SY, et al. Flavonoids of honey and propolis: Characterization and effects on hepatic drug-metabolising enzymes and benzo[a]pyrene-DNA binding in rats. J Agric Food Chem. 1996;44:2379–83. doi: 10.1021/jf9504733. [DOI] [Google Scholar]
- 104.Dhawan K, Kumar S, Sharma A. Beneficial effects of chrysin and benzoflavone on virility in 2-year-old male rats. J Med Food. 2002;5:43–8. doi: 10.1089/109662002753723214. [DOI] [PubMed] [Google Scholar]
- 105.Sandborn WJ, Faubion WA. Clinical pharmacology of inflammatory bowel disease therapies. Curr Gastroenterol Rep. 2000;2:440–5. doi: 10.1007/s11894-000-0005-0. [DOI] [PubMed] [Google Scholar]
- 106.Williams CA, Harborne JB, Newman M, Greenham J, Eagles J. Chrysin and other leaf exudate flavonoids in the genus Pelargonium. Phytochemistry. 1997;46:1349–53. doi: 10.1016/S0031-9422(97)00514-1. [DOI] [PubMed] [Google Scholar]
- 107.Ahad A, Ganai AA, Mujeeb M, Siddiqui WA. Chrysin, an anti-inflammatory molecule, abrogates renal dysfunction in type 2 diabetic rats. Toxicol Appl Pharmacol. 2014;279:1–7. doi: 10.1016/j.taap.2014.05.007. [DOI] [PubMed] [Google Scholar]
- 108.Li R, Zang A, Zhang L, Zhang H, Zhao L, Qi Z, et al. Chrysin ameliorates diabetes-associated cognitive deficits in Wistar rats. Neurol Sci. 2014;35:1527–32. doi: 10.1007/s10072-014-1784-7. [DOI] [PubMed] [Google Scholar]
- 109.Sirovina D, Orsolić N, Koncić MZ, Kovacević G, Benković V, Gregorović G. Quercetin vs chrysin: Effect on liver histopathology in diabetic mice. Hum Exp Toxicol. 2013;32:1058–66. doi: 10.1177/0960327112472993. [DOI] [PubMed] [Google Scholar]
- 110.El-Bassossy HM, Abo-Warda SM, Fahmy A. Chrysin and luteolin attenuate diabetes-induced impairment in endothelial-dependent relaxation: Effect on lipid profile, AGEs and NO generation. Phytother Res. 2013;27:1678–84. doi: 10.1002/ptr.4917. [DOI] [PubMed] [Google Scholar]
- 111.Neuhouser ML. Dietary flavonoids and cancer risk: Evidence from human population studies. Nutr Cancer. 2004;50:1–7. doi: 10.1207/s15327914nc5001_1. [DOI] [PubMed] [Google Scholar]
- 112.Miean KH, Mohamed S. Flavonoid (myricetin, quercetin, kaempferol, luteolin, and apigenin) content of edible tropical plants. J Agric Food Chem. 2001;49:3106–12. doi: 10.1021/jf000892m. [DOI] [PubMed] [Google Scholar]
- 113.Gates MA, Tworoger SS, Hecht JL, De Vivo I, Rosner B, Hankinson SE, et al. A prospective study of dietary flavonoid intake and incidence of epithelial ovarian cancer. Int J Cancer. 2007;121:2225–32. doi: 10.1002/ijc.22790. [DOI] [PubMed] [Google Scholar]
- 114.Sun T, Xu Z, Wu CT, Janes M, Prinyawiwatkul W, No HK. Antioxidant activities of different colored sweet bell peppers (Capsicum annuum L) J Food Sci. 2007;72:98–102. doi: 10.1111/j.1750-3841.2006.00245.x. [DOI] [PubMed] [Google Scholar]
- 115.Ding L, Jin D, Chen X. Luteolin enhances insulin sensitivity via activation of PPARγ transcriptional activity in adipocytes. J Nutr Biochem. 2010;21:941–47. doi: 10.1016/j.jnutbio.2009.07.009. [DOI] [PubMed] [Google Scholar]
- 116.Liu Y, Fu X, Lan N, Li S, Zhang J, Wang S, et al. Luteolin protects against high fat diet-induced cognitive deficits in obesity mice. Behav Brain Res. 2014;267:178–188. doi: 10.1016/j.bbr.2014.02.040. [DOI] [PubMed] [Google Scholar]
- 117.Wang GG, Lu XH, Li W, Zhao X, Zhang C. Protective effects of luteolin on diabetic nephropathy in STZ-induced diabetic rats. Evid Based Complement Alternat Med. 2011;2011:323171. doi: 10.1155/2011/323171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Deqiu Z, Kang L, Jiali Y, Baolin L, Gaolin L. Luteolin inhibits inflammatory response and improves insulin sensitivity in the endothelium. Biochimie. 2011;93:506–12. doi: 10.1016/j.biochi.2010.11.002. [DOI] [PubMed] [Google Scholar]
- 119.Ding Y, Shi X, Shuai X, Xu Y, Liu Y, Liang X, et al. Luteolin prevents uric acid-induced pancreatic β-cell dysfunction. J Biomed Res. 2014;28:292–8. doi: 10.7555/JBR.28.20130170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Kim HJ, Lee W, Yun JM. Luteolin inhibits hyperglycemia-induced proinflammatory cytokine production and its epigenetic mechanism in human monocytes. Phytother Res. 2014;28:1383–91. doi: 10.1002/ptr.5141. [DOI] [PubMed] [Google Scholar]
- 121.Kwon EY, Jung UJ, Park T, Yun JW, Choi MS. Luteolin attenuates hepatic steatosis and insulin resistance through the interplay between the liver and adipose tissue in mice with diet-induced obesity. Diabetes. 2015;64:1658–69. doi: 10.2337/db14-0631. [DOI] [PubMed] [Google Scholar]
- 122.Yang JT, Qian LB, Zhang FJ, Wang J, Ai H, Tang LH, et al. Cardioprotective effects of luteolin on ischemia/reperfusion injury in diabetic rats are modulated by eNOS and the mitochondrial permeability transition pathway. J Cardiovasc Pharmacol. 2015;65:349–56. doi: 10.1097/FJC.0000000000000202. [DOI] [PubMed] [Google Scholar]
- 123.Xu N, Zhang L, Dong J, Zhang X, Chen YG, Bao B, et al. Low-dose diet supplement of a natural flavonoid, luteolin, ameliorates diet-induced obesity and insulin resistance in mice. Mol Nutr Food Res. 2014;58:1258–68. doi: 10.1002/mnfr.201300830. [DOI] [PubMed] [Google Scholar]
- 124.Kim MS, Hur HJ, Kwon DY, Hwang JT. Tangeretin stimulates glucose uptake via regulation of AMPK signaling pathways in C2C12 myotubes and improves glucose tolerance in high-fat diet-induced obese mice. Mol Cell Endocrinol. 2012;358:127–34. doi: 10.1016/j.mce.2012.03.013. [DOI] [PubMed] [Google Scholar]
- 125.Sundaram R, Shanthi P, Sachdanandam P. Effect of tangeretin, a polymethoxylated flavone on glucose metabolism in streptozotocin-induced diabetic rats. Phytomedicine. 2014;21:793–9. doi: 10.1016/j.phymed.2014.01.007. [DOI] [PubMed] [Google Scholar]
- 126.Miyata Y, Tanaka H, Shimada A, Sato T, Ito A, Yamanouchi T, et al. Regulation of adipocytokine secretion and adipocyte hypertrophy by polymethoxy flavonoids, nobiletin and tangeretin. Life Sci. 2011;88:613–18. doi: 10.1016/j.lfs.2011.01.024. [DOI] [PubMed] [Google Scholar]
- 127.Tai MC, Tsang SY, Chang LY, Xue H. Therapeutic potential of wogonin: A naturally occurring flavonoid. CNS Drug Rev. 2005;11:141e50. doi: 10.1111/j.1527-3458.2005.tb00266.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Bak EJ, Kim J, Choi YH, Kim JH, Lee DE, Woo GH, et al. Wogonin ameliorates hyperglycemia and dyslipidemia via PPARα activation in db/db mice. Clin Nutr. 2014;33:156–63. doi: 10.1016/j.clnu.2013.03.013. [DOI] [PubMed] [Google Scholar]
- 129.Ku SK, Bae JS. Baicalin, baicalein and wogonin inhibits high glucose-induced vascular inflammation in vitro and in vivo. BMB Rep. 2015;48:519–24. doi: 10.5483/BMBRep.2015.48.9.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Zhang YM, Li MX, Tang Z, Wang CH. Wogonin suppresses osteopontin expression in adipocytes by activating PPARα. Acta Pharmacol Sin. 2015;36:987–97. doi: 10.1038/aps.2015.37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Yokozawa T, Kim HY, Cho EJ, Choi JS, Chung HY. Antioxidant effects of isorhamnetin 3,7-di-O-beta-D-glucopyranoside isolated from mustard leaf (Brassica juncea) in rats with streptozotocin-induced diabetes. J Agric Food Chem. 2002;50:5490–95. doi: 10.1021/jf0202133. [DOI] [PubMed] [Google Scholar]
- 132.Lee YS, Lee S, Lee HS, Kim BK, Ohuchi K, Shin KH. Inhibitory effects of isorhamnetin-3-O-beta-D-glucoside from Salicornia herbacea on rat lens aldose reductase and sorbitol accumulation in streptozotocin-induced diabetic rat tissues. Biol Pharm Bull. 2005;28:916–8. doi: 10.1248/bpb.28.916. [DOI] [PubMed] [Google Scholar]
- 133.Rodríguez-Rodríguez C, Torres N, Gutiérrez-Uribe JA, Noriega LG, Torre-Villalvazo I, Leal-Díaz AM, et al. The effect of isorhamnetin glycosides extracted from Opuntia ficus-indica in a mouse model of diet induced obesity. Food Funct. 2015;6:805–15. doi: 10.1039/C4FO01092B. [DOI] [PubMed] [Google Scholar]
- 134.Lee J, Jung E, Lee J, Kim S, Huh S, Kim Y, et al. Isorhamnetin represses adipogenesis in 3 T3-L1 cells. Obesity (Silver Spring) 2009;17:226–32. doi: 10.1038/oby.2008.472. [DOI] [PubMed] [Google Scholar]
- 135.An G, Gallegos J, Morris ME. The bioflavonoid kaempferol is an abcg2 substrateand inhibits abcg2-mediated quercetin efflux. Drug Metab Dispos. 2011;39:426–32. doi: 10.1124/dmd.110.035212. [DOI] [PubMed] [Google Scholar]
- 136.Hakkinen SH, Karenlampi SO, Heinonen IM, Mykkanen HM, Torronen AR. Content of the flavonolsquercetin, myricetin, and kaempferolin 25 edible berries. J Agric Food Chem. 1999;47:2274–79. doi: 10.1021/jf9811065. [DOI] [PubMed] [Google Scholar]
- 137.Nirmala P, Ramanathan M. Effect of kaempferol on lipid peroxidation and antioxidant status in 1,2-dimethyl hydrazine induced colorectal carcinoma in rats. Eur J Pharmacol. 2011;654:75–9. doi: 10.1016/j.ejphar.2010.11.034. [DOI] [PubMed] [Google Scholar]
- 138.Jorge AP, Horst H, de Sousa E, Pizzolatti MG, Silva FR. Insulinomimetic effects of kaempferitrin on glycaemia and on 14C-glucose uptake in rat soleus muscle. Chem Biol Interact. 2004;149:89–96. doi: 10.1016/j.cbi.2004.07.001. [DOI] [PubMed] [Google Scholar]
- 139.Zhang Y, Liu D. Flavonol kaempferol improves chronic hyperglycemia-impaired pancreatic beta-cell viability and insulin secretory function. Eur J Pharmacol. 2011;670:325–32. doi: 10.1016/j.ejphar.2011.08.011. [DOI] [PubMed] [Google Scholar]
- 140.Zanatta L, Rosso A, Folador P, Figueiredo MS, Pizzolatti MG, Leite LD, et al. Insulinomimetic effect of kaempferol 3-neohesperidoside on the rat soleus muscle. J Nat Prod. 2008;71:532–5. doi: 10.1021/np070358+. [DOI] [PubMed] [Google Scholar]
- 141.Al-Numair KS, Chandramohan G, Veeramani C, Alsaif MA. Ameliorative effect of kaempferol, a flavonoid, on oxidative stress in streptozotocin-induced diabetic rats. Redox Rep. 2015;20:198–209. doi: 10.1179/1351000214Y.0000000117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Abo-Salem OM. Kaempferol attenuates the development of diabetic neuropathic pain in mice: Possible anti-inflammatory and anti-oxidant mechanisms. Macedonian J Med Sci. 2014;7:424–30. doi: 10.3889/oamjms.2014.073. [DOI] [Google Scholar]
- 143.Zang Y, Zhang L, Igarashi K, Yu C. The anti-obesity and anti-diabetic effects of kaempferol glycosides from unripe soybean leaves in high-fat-diet mice. Food Funct. 2015;6:834–41. doi: 10.1039/C4FO00844H. [DOI] [PubMed] [Google Scholar]
- 144.Al-Numair KS, Veeramani C, Alsaif MA, Chandramohan G. Influence of kaempferol, a flavonoid compound, on membrane-bound ATPases in streptozotocin-induced diabetic rats. Pharm Biol. 2015;8:1–7. doi: 10.3109/13880209.2014.982301. [DOI] [PubMed] [Google Scholar]
- 145.Zhang Y, Zhen W, Maechler P, Liu D. Small molecule kaempferol modulates PDX-1 protein expression and subsequently promotes pancreatic β-cell survival and function via CREB. J Nutr Biochem. 2013;24:638–46. doi: 10.1016/j.jnutbio.2012.03.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Alkhalidy H, Moore W, Zhang Y, McMillan R, Wang A, Ali M, et al. Small molecule kaempferol promotes insulin sensitivity and preserved pancreatic β-cell mass in middle-aged obese diabetic mice. J Diabetes Res. 2015;2015:532984. doi: 10.1155/2015/532984. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 147.Kreft S, Knapp M, Kreft I. Extraction of rutin from buckwheat (Fagopyrum esculentum Moench) seeds and determination by capillary electrophoresis. J Agric Food Chem. 1999;47:4649–52. doi: 10.1021/jf990186p. [DOI] [PubMed] [Google Scholar]
- 148.Huang WY, Zhang HC, Liu WX, Li CY. Survey of antioxidant capacity and phenolic composition of blueberry, blackberry, and strawberry in Nanjing. J Zhejiang Univ Sci B. 2012;13:94–102. doi: 10.1631/jzus.B1100137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Prince PSM, Kamalakkannan N. Rutin improves glucose homeostasis in streptozotocin diabetic tissues by altering glycolytic and gluconeogenic enzymes. J Biochem Mol Toxicol. 2006;20:96–102. doi: 10.1002/jbt.20117. [DOI] [PubMed] [Google Scholar]
- 150.Fernandes AA, Novelli EL, Okoshi K, Okoshi MP, Di Muzio BP, Guimarães JF, et al. Influence of rutin treatment on biochemical alterations in experimental diabetes. Biomed Pharmacother. 2010;64:214–19. doi: 10.1016/j.biopha.2009.08.007. [DOI] [PubMed] [Google Scholar]
- 151.Prince PSM, Kamalakkannan N. Protective effect of rutin on lipids, lipoproteins, lipid metabolizing enzymes and glycoproteins in streptozotocin-induced diabetic rats. J Pharm Pharmacol. 2006;58:1373–83. doi: 10.1211/jpp.58.10.0011. [DOI] [PubMed] [Google Scholar]
- 152.Hao HH, Shao ZM, Tang DQ, Lu Q, Chen X, Yin XX, et al. Preventive effects of rutin on the development of experimental diabetic nephropathy in rats. Life Sci. 2012;91:959–67. doi: 10.1016/j.lfs.2012.09.003. [DOI] [PubMed] [Google Scholar]
- 153.Kappel VD, Cazarolli LH, Pereira DF, Postal BG, Zamoner A, Reginatto FH, et al. Involvement of GLUT-4 in the stimulatory effect of rutin on glucose uptake in rat soleus muscle. J Pharm Pharmacol. 2013;65:1179–86. doi: 10.1111/jphp.12066. [DOI] [PubMed] [Google Scholar]
- 154.Niture NT, Ansari AA, Naik SR. Anti-hyperglycemic activity of rutin in streptozotocin-induced diabetic rats: an effect mediated through cytokines, antioxidants and lipid biomarkers. Indian J Exp Biol. 2014;52:720–7. [PubMed] [Google Scholar]
- 155.Wang YB, Ge ZM, Kang WQ, Lian ZX, Yao J, Zhou CY. Rutin alleviates diabetic cardiomyopathy in a rat model of type 2 diabetes. Exp Ther Med. 2015;9:451–55. doi: 10.3892/etm.2014.2090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Ola MS, Ahmed MM, Ahmad R, Abuohashish HM, Al-Rejaie SS, Alhomida AS. Neuroprotective effects of rutin in streptozotocin-induced diabetic rat retina. J Mol Neurosci. 2015;56:440–8. doi: 10.1007/s12031-015-0561-2. [DOI] [PubMed] [Google Scholar]
- 157.Kamalakkannan N, Prince PS. Antihyperglycaemic and antioxidant effect of rutin, a polyphenolic flavonoid, in streptozotocin-induced diabetic wistar rats. Basic Clin Pharmacol Toxicol. 2006;98:97–103. doi: 10.1111/j.1742-7843.2006.pto_241.x. [DOI] [PubMed] [Google Scholar]
- 158.Butchi Akondi R, Kumar P, Annapurna A, Pujari M. Protective effect of rutin and naringin on sperm quality in streptozotocin induced type 1 diabetic rats. Iran J Pharm Res. 2011;10:585–96. [PMC free article] [PubMed] [Google Scholar]
- 159.Hollman PCH, de Vries JHM, van Leeuwen SD, Mengelers MJB, Katan MB. Absorption of dietary quercetin glycosides and quercetin in healthy ileostomy volunteers. Am J Clin Nutr. 1995;62:1276–82. doi: 10.1093/ajcn/62.6.1276. [DOI] [PubMed] [Google Scholar]
- 160.Alinezhad H, Azimi A, Zare M, Ebrahimzadeh MA, Eslami S, Nabavi SF, et al. Antioxidant and antihemolytic activities of ethanolic extract of flowers, leaves, and stems of Hyssopus officinalis L. var. angustifolius. Int J Food Prop. 2013;16:1169–78. doi: 10.1080/10942912.2011.578319. [DOI] [Google Scholar]
- 161.Coskun O, Kanter M, Korkmaz A, Oter S. Quercetin, a flavonoid antioxidant, prevents and protects streptozotocin-induced oxidative stress and beta-cell damage in rat pancreas. Pharmacol Res. 2005;51:117–23. doi: 10.1016/j.phrs.2004.06.002. [DOI] [PubMed] [Google Scholar]
- 162.Stewart LK, Wang Z, Ribnicky D, Soileau JL, Cefalu WT, Gettys TW. Failure of dietary quercetin to alter the temporal progression of insulin resistance among tissues of C57BL/6 J mice during the development of diet-induced obesity. Diabetologia. 2009;52:514–23. doi: 10.1007/s00125-008-1252-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Kwon O, Eck P, Chen S, Corpe CP, Lee JH, Kruhlak M, et al. Inhibition of the intestinal glucose transporter GLUT2 by flavonoids. FASEB J. 2007;21:366–77. doi: 10.1096/fj.06-6620com. [DOI] [PubMed] [Google Scholar]
- 164.Eid HM, Nachar A, Thong F, Sweeney G, Haddad PS. The molecular basis of the antidiabetic action of quercetin in cultured skeletal muscle cells and hepatocytes. Pharmacogn Mag. 2015;11:74–81. doi: 10.4103/0973-1296.149708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Panchal SK, Poudyal H, Brown L. Quercetin ameliorates cardiovascular, hepatic, and metabolic changes in diet-induced metabolic syndrome in rats. J Nutr. 2012;142:1026–32. doi: 10.3945/jn.111.157263. [DOI] [PubMed] [Google Scholar]
- 166.Kobori M, Masumoto S, Akimoto Y, Takahashi Y. Dietary quercetin alleviates diabetic symptoms and reduces streptozotocin-induced disturbance of hepatic gene expression in mice. Mol Nutr Food Res. 2009;53:859–68. doi: 10.1002/mnfr.200800310. [DOI] [PubMed] [Google Scholar]
- 167.Lu Q, Ji XJ, Zhou YX, Yao XQ, Liu YQ, Zhang F, et al. Quercetin inhibits the mTORC1/p70S6K signaling-mediated renal tubular epithelial-mesenchymal transition and renal fibrosis in diabetic nephropathy. Pharmacol Res. 2015;99:237–47. doi: 10.1016/j.phrs.2015.06.006. [DOI] [PubMed] [Google Scholar]
- 168.Kumar B, Gupta SK, Nag TC, Srivastava S, Saxena R, Jha KA, et al. Retinal neuroprotective effects of quercetin in streptozotocin-induced diabetic rats. Exp Eye Res. 2014;125:193–202. doi: 10.1016/j.exer.2014.06.009. [DOI] [PubMed] [Google Scholar]
- 169.Alam MM, Meerza D, Naseem I. Protective effect of quercetin on hyperglycemia, oxidative stress and DNA damage in alloxan induced type 2 diabetic mice. Life Sci. 2014;109:8–14. doi: 10.1016/j.lfs.2014.06.005. [DOI] [PubMed] [Google Scholar]
- 170.Dai X, Ding Y, Zhang Z, Cai X, Li Y. Quercetin and quercitrin protect against cytokine-induced injuries in RINm5F β-cells via the mitochondrial pathway and NF-κB signaling. Int J Mol Med. 2013;31:265–71. doi: 10.3892/ijmm.2012.1177. [DOI] [PubMed] [Google Scholar]
- 171.Jeong SM, Kang MJ, Choi HN, Kim JH, Kim JI. Quercetin ameliorates hyperglycemia and dyslipidemia and improves antioxidant status in type 2 diabetic db/db mice. Nutr Res Pract. 2012;6:201–7. doi: 10.4162/nrp.2012.6.3.201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Lai PB, Zhang L, Yang LY. Quercetin ameliorates diabetic nephropathy by reducing the expressions of transforming growth factor-β1 and connective tissue growth factor in streptozotocin-induced diabetic rats. Ren Fail. 2012;34:83–7. doi: 10.3109/0886022X.2011.623564. [DOI] [PubMed] [Google Scholar]
- 173.Kim JH, Kang MJ, Choi HN, Jeong SM, Lee YM, Kim JI. Quercetin attenuates fasting and postprandial hyperglycemia in animal models of diabetes mellitus. Nutr Res Pract. 2011;5:107–11. doi: 10.4162/nrp.2011.5.2.107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Patisaul HB, Jefferson W. The pros and cons of phytoestrogens. Front Neuroendocrinol. 2010;31:400–19. doi: 10.1016/j.yfrne.2010.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Guo TL, Wang Y, Xiong T, Ling X, Zheng J. Genistein modulation of streptozotocin diabetes in male B6C3F1 mice can be induced by diet. Toxicol Appl Pharmacol. 2014;280:455–66. doi: 10.1016/j.taap.2014.08.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Babu PV, Si H, Fu Z, Zhen W, Liu D. Genistein prevents hyperglycemia-induced monocyte adhesion to human aortic endothelial cells through preservation of the cAMP signaling pathway and ameliorates vascular inflammation in obese diabetic mice. J Nutr. 2012;142:724–30. doi: 10.3945/jn.111.152322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Palanisamy N, Viswanathan P, Anuradha CV. Effect of genistein, a soy isoflavone, on whole body insulin sensitivity and renal damage induced by a high-fructose diet. Ren Fail. 2008;30:645–54. doi: 10.1080/08860220802134532. [DOI] [PubMed] [Google Scholar]
- 178.Elmarakby AA, Ibrahim AS, Faulkner J, Mozaffari MS, Liou GI, Abdelsayed R. Tyrosine kinase inhibitor, genistein, reduces renal inflammation and injury in streptozotocininduced diabetic mice. Vascular Pharma. 2011;55:149–56. doi: 10.1016/j.vph.2011.07.007. [DOI] [PubMed] [Google Scholar]
- 179.Fu Z, Gilbert ER, Pfeiffer L, Zhang Y, Fu Y, Liu D. Genistein ameliorates hyperglycemia in a mouse model of nongenetic type 2 diabetes. Appl Physiol Nutr Metab. 2012;37:480–8. doi: 10.1139/h2012-005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Valsecchi AE, Franchi S, Panerai AE, Rossi A, Sacerdote P, Colleoni M. The soy isoflavone genistein reverses oxidative and inflammatory state, neuropathic pain, neurotrophic and vasculature deficits in diabetes mouse model. Eur J Pharmacol. 2011;650:694–702. doi: 10.1016/j.ejphar.2010.10.060. [DOI] [PubMed] [Google Scholar]
- 181.El-Kordy EA, Alshahrani AM. Effect of genistein, a natural soy isoflavone, on pancreatic-cells of streptozotocin-induced diabetic rats: Histological and immunohistochemical study. J Microsc Ultrastruct. 2015;3:108–119. doi: 10.1016/j.jmau.2015.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Weigt C, Hertrampf T, Flenker U, Hülsemann F, Kurnaz P, Fritzemeier KH, et al. Effects of estradiol, estrogen receptor subtype-selective agonists and genistein on glucose metabolism in leptin resistant female Zucker diabetic fatty (ZDF) rats. J Steroid Biochem Mol Biol. 2015;154:12–22. doi: 10.1016/j.jsbmb.2015.06.002. [DOI] [PubMed] [Google Scholar]
- 183.Gupta SK, Dongare S, Mathur R, Mohanty IR, Srivastava S, Mathur S, et al. Genistein ameliorates cardiac inflammation and oxidative stress in streptozotocin-induced diabetic cardiomyopathy in rats. Mol Cell Biochem. 2015;408:63–72. doi: 10.1007/s11010-015-2483-2. [DOI] [PubMed] [Google Scholar]
- 184.Kim MJ, Lim Y. Protective effect of short-term genistein supplementation on the early stage in diabetes-induced renal damage. Mediators Inflamm. 2013;510212:14. doi: 10.1155/2013/510212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Tie L, An Y, Han J, Xiao Y, Xiaokaiti Y, Fan S, et al. Genistein accelerates refractory wound healing by suppressing superoxide and FoxO1/iNOS pathway in type 1 diabetes. J Nutr Biochem. 2013;24:88–96. doi: 10.1016/j.jnutbio.2012.02.011. [DOI] [PubMed] [Google Scholar]
- 186.Yang W, Wang S, Li L, Liang Z, Wang L. Genistein reduces hyperglycemia and islet cell loss in a high-dosage manner in rats with alloxan-induced pancreatic damage. Pancreas. 2011;40:396–402. doi: 10.1097/MPA.0b013e318204e74d. [DOI] [PubMed] [Google Scholar]
- 187.Fu Z, Zhang W, Zhen W, Lum H, Nadler J, Bassaganya-Riera J, et al. Genistein induces pancreatic beta-cell proliferation through activation of multiple signaling pathways and prevents insulin-deficient diabetes in mice. Endocrinology. 2010;151:3026–37. doi: 10.1210/en.2009-1294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Yuan WJ, Jia FY, Meng JZ. Effects of genistein on secretion of extracellular matrix components and transforming growth factor beta in high-glucose-cultured rat mesangial cells. J Artif Organs. 2009;12:242–6. doi: 10.1007/s10047-009-0479-y. [DOI] [PubMed] [Google Scholar]
- 189.Zhang M, Ikeda K, Xu JW, Yamori Y, Gao XM, Zhang BL. Genistein suppresses adipogenesis of 3 T3-L1 cells via multiple signal pathways. Phytother Res. 2009;23:713–8. doi: 10.1002/ptr.2724. [DOI] [PubMed] [Google Scholar]
- 190.Kim YS, Kim NH, Jung DH, Jang DS, Lee YM, Kim JM, et al. Genistein inhibits aldose reductase activity and high glucose-induced TGF-beta2 expression in human lens epithelial cells. Eur J Pharmacol. 2008;594:18–25. doi: 10.1016/j.ejphar.2008.07.033. [DOI] [PubMed] [Google Scholar]
- 191.Choi MS, Jung UJ, Yeo J, Kim MJ, Lee MK. Genistein and daidzein prevent diabetes onset by elevating insulin level and altering hepatic gluconeogenic and lipogenic enzyme activities in non-obese diabetic (NOD) mice. Diabetes Metab Res Rev. 2008;24:74–81. doi: 10.1002/dmrr.780. [DOI] [PubMed] [Google Scholar]
- 192.Liggins J, Bluck LJ, Runswick S, Atkinson C, Coward WA, Bingham SA. Daidzein and genistein content of fruits and nuts. J Nutr Biochem. 2000;11:326–31. doi: 10.1016/S0955-2863(00)00085-1. [DOI] [PubMed] [Google Scholar]
- 193.Kapiotis S, Jilma B, Szalay Y. Evidence against an effect of endothelin-1 on blood coagulation, fibrinolysis, and endothelial cell integrity in healthy men. Arterioscler Thromb Vasc Biol. 1997;17:2861–67. doi: 10.1161/01.ATV.17.11.2861. [DOI] [PubMed] [Google Scholar]
- 194.Park SA, Choi MS, Cho SY, Seo JS, Jung UJ, Kim MJ, et al. Genistein and daidzein modulate hepatic glucose and lipid regulating enzyme activities in C57BL/Ks J-db/db mice. Life Sci. 2006;79:1207–13. doi: 10.1016/j.lfs.2006.03.022. [DOI] [PubMed] [Google Scholar]
- 195.Cederroth CR, Vinciguerra M, Gjinovci A, Kuhne F, Klein M, Cederroth M, et al. Dietary phytoestrogens activate AMP-activated protein kinase with improvement in lipid and glucose metabolism. Diabetes. 2008;57:1176–85. doi: 10.2337/db07-0630. [DOI] [PubMed] [Google Scholar]
- 196.Cheong SH, Furuhashi K, Ito K, Nagaoka M, Yonezawa T, Miura Y, et al. Daidzein promotes glucose uptake through glucose transporter 4 translocation to plasma membrane in L6 myocytes and improves glucose homeostasis in type II diabetic model mice. J Nutr Biochem. 2014;25:136–43. doi: 10.1016/j.jnutbio.2013.09.012. [DOI] [PubMed] [Google Scholar]
- 197.Roghani M, Vaez Mahdavi MR, Jalali-Nadoushan MR, Baluchnejadmojarad T, Naderi G, Roghani-Dehkordi F, et al. Chronic administration of daidzein, a soybean isoflavone, improves endothelial dysfunction and attenuates oxidative stress in streptozotocin-induced diabetic rats. Phytother Res. 2013;27:112–7. doi: 10.1002/ptr.4699. [DOI] [PubMed] [Google Scholar]
- 198.Cho KW, Lee OH, Banz WJ, Moustaid-Moussa N, Shay NF, Kim YC. Daidzein and the daidzein metabolite, equol, enhance adipocyte differentiation and PPAR γ transcriptional activity. J Nutr Biochem. 2010;21:841–7. doi: 10.1016/j.jnutbio.2009.06.012. [DOI] [PubMed] [Google Scholar]
- 199.Akkarachiyasit S, Charoenlertkul P, Yibchok-Anun S, Adisakwattana S. Inhibitory activities of cyanidin and its glycosides and synergistic effect with acarbose against intestinal α-glucosidase and pancreatic α-amylase. Int J Mol Sci. 2010;11:3387–96. doi: 10.3390/ijms11093387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Nizamutdinova IT, Jin YC, Chung JI, Shin SC, Lee SJ, Seo HG, et al. The anti-diabetic effect of anthocyanins in streptozotocin-induced diabetic rats through glucose transporter 4 regulation and prevention of insulin resistance and pancreatic apoptosis. Mol Nutr Food Res. 2009;53:1419–29. doi: 10.1002/mnfr.200800526. [DOI] [PubMed] [Google Scholar]
- 201.Nasri S, Roghani M, Baluchnejadmojarad T, Rabani T, Balvardi M. Vascular mechanisms of cyanidin-3-glucoside response in streptozotocin-diabetic rats. Pathophysiology. 2011;18:273–8. doi: 10.1016/j.pathophys.2011.03.001. [DOI] [PubMed] [Google Scholar]
- 202.Zhu W, Jia Q, Wang Y, Zhang Y, Xia M. The anthocyanin cyanidin-3-O-betaglucoside, a flavonoid, increases hepatic glutathione synthesis and protects hepatocytes against reactive oxygen species during hyperglycemia: Involvement of a cAMP-PKA-dependent signaling pathway. Free RadicBiol Med. 2012;52:314–27. doi: 10.1016/j.freeradbiomed.2011.10.483. [DOI] [PubMed] [Google Scholar]
- 203.Kurimoto Y, Shibayama Y, Inoue S, Soga M, Takikawa M, Ito C, et al. Black soybean seed coat extract ameliorates hyperglycemia and insulin sensitivityvia the activation of AMP-activated protein kinase in diabetic mice. J Agric Food Chem. 2013;61:5558–64. doi: 10.1021/jf401190y. [DOI] [PubMed] [Google Scholar]
- 204.Jiang X, Tang X, Zhang P, Liu G, Guo H. Cyanidin-3-O-β-glucoside protects primary mouse hepatocytes against high glucose-induced apoptosis by modulating mitochondrial dysfunction and the PI3K/Akt pathway. Biochem Pharmacol. 2014;90:135–44. doi: 10.1016/j.bcp.2014.04.018. [DOI] [PubMed] [Google Scholar]
- 205.Guo H, Xia M, Zou T, Ling W, Zhong R, Zhang W. Cyanidin 3-glucoside attenuates obesity-associated insulin resistance and hepatic steatosis in high-fat diet-fed and db/db mice via the transcription factor FoxO1. J Nutr Biochem. 2012;23:349–60. doi: 10.1016/j.jnutbio.2010.12.013. [DOI] [PubMed] [Google Scholar]
- 206.Park S, Kang S, Jeong DY, Jeong SY, Park JJ, Yun HS. Cyanidin and malvidin in aqueous extracts of black carrots fermented with Aspergillus oryzae prevent the impairment of energy, lipid and glucose metabolism in estrogen-deficient rats by AMPK activation. Genes Nutr. 2015;10:455. doi: 10.1007/s12263-015-0455-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Matsukawa T, Inaguma T, Han J, Villareal MO, Isoda H. Cyanidin-3-glucoside derived from black soybeans ameliorate type 2 diabetes through the induction of differentiation of preadipocytes into smaller and insulin-sensitive adipocytes. J Nutr Biochem. 2015;26:860–7. doi: 10.1016/j.jnutbio.2015.03.006. [DOI] [PubMed] [Google Scholar]
- 208.Sasaki R, Nishimura N, Hoshino H, Isa Y, Kadowaki M, Ichi T, et al. Cyanidin 3-glucoside ameliorates hyperglycemia and insulin sensitivity due to down-regulation of retinol binding protein 4 expression in diabetic mice. Biochem Pharmacol. 2007;74:1619–27. doi: 10.1016/j.bcp.2007.08.008. [DOI] [PubMed] [Google Scholar]
- 209.Tsao TS, Burcelin R, Charron MJ. Regulation of hexokinase II gene expression by glucose flux in skeletal muscle. J Biol Chem. 1996;271:14959–63. doi: 10.1074/jbc.271.9.4993. [DOI] [PubMed] [Google Scholar]
- 210.Sun CD, Zhang B, Zhang JK, Xu CJ, Wu YL, Li X, et al. Cyanidin-3-glucoside-rich extract from Chinese bayberry fruit protects pancreatic β-cells and ameliorates hyperglycemia in streptozotocin-induced diabetic mice. J Med Food. 2012;15:288–98. doi: 10.1089/jmf.2011.1806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Bertuglia S, Malandrino S, Colantuoni A. Effects of the natural flavonoid delphinidin on diabetic microangiopathy. Arzneimittelforschung. 1995;45:481–85. [PubMed] [Google Scholar]
- 212.Gharib A, Faezizadeh Z, Godarzee M. Treatment of diabetes in the mouse model by delphinidin and cyanidin hydrochloride in free and liposomal forms. Planta Med. 2013;79:1599–604. doi: 10.1055/s-0033-1350908. [DOI] [PubMed] [Google Scholar]
- 213.Mazza G. Compositional and functional properties of saskatoon berry and blueberry. Int J Fruit Sci. 2005;5:101. doi: 10.1300/J492v05n03_10. [DOI] [Google Scholar]
- 214.Roy M, Sen S, Chakraborti AS. Action of pelargonidin on hyperglycemia and oxidative damage in diabetic rats: Implication for glycation-induced hemoglobin modification. Life Sci. 2008;82:1102–10. doi: 10.1016/j.lfs.2008.03.011. [DOI] [PubMed] [Google Scholar]
- 215.Mirshekar M, Roghani M, Khalili M, Baluchnejadmojarad T, Arab MS. Chronic oral pelargonidin alleviates streptozotocin-induced diabetic neuropathic hyperalgesia in rat: Involvement of oxidative stress. Iran Biomed J. 2010;14:33–9. [PMC free article] [PubMed] [Google Scholar]
- 216.Jayaprakasam B, Vareed SK, Olson LK, Nair MG. Insulin secretion by anthocyanins and anthocyanidins. J Agric Food Chem. 2005;53:2519–23. doi: 10.1021/jf049018+. [DOI] [PubMed] [Google Scholar]
- 217.Mirshekar M, Roghani M, Khalili M, Baluchnejadmojarad T. Chronic oral pelargonidin alleviates learning and memory disturbances in streptozotocin diabetic rats. Iran J Pharm Res. 2011;10:569–75. [PMC free article] [PubMed] [Google Scholar]