

Abstract
Opioid-induced hyperalgesia and tolerance severely impact the clinical efficacy of opiates as pain relievers in animals and humans. The molecular mechanisms underlying both phenomena are not well understood and their elucidation should benefit from the study of animal models and from the design of appropriate experimental protocols.
We describe here a methodological approach for inducing, recording and quantifying morphine-induced hyperalgesia as well as for evidencing analgesic tolerance, using the tail-immersion and tail pressure tests in wild-type mice. As shown in the video, the protocol is divided into five sequential steps. Handling and habituation phases allow a safe determination of the basal nociceptive response of the animals. Chronic morphine administration induces significant hyperalgesia as shown by an increase in both thermal and mechanical sensitivity, whereas the comparison of analgesia time-courses after acute or repeated morphine treatment clearly indicates the development of tolerance manifested by a decline in analgesic response amplitude. This protocol may be similarly adapted to genetically modified mice in order to evaluate the role of individual genes in the modulation of nociception and morphine analgesia. It also provides a model system to investigate the effectiveness of potential therapeutic agents to improve opiate analgesic efficacy.
Keywords: Neuroscience, Issue 89, mice, nociception, tail immersion test, tail pressure test, morphine, analgesia, opioid-induced hyperalgesia, tolerance
Introduction
Opioid-induced hyperalgesia (OIH) and analgesic tolerance limit the clinical efficacy of opiates in animals and humans1-3. The involvement of pro-inflammatory4,5 or of pro-nociceptive (anti-opioid)6,7 systems are currently explored hypotheses. The elucidation of the mechanisms underlying OIH and tolerance necessitates a combination of in vivo and in vitro approaches, using appropriate animal models, experimental protocols and molecular tools.
Behavioral pharmacology is the dominant paradigm to monitor and quantify analgesic and hyperalgesic states in laboratory animals (rats, mice). The application of a noxious stimulus (thermal, mechanical or chemical) to a convenient body part (hindpaw, tail) of the animal leads to a nocifensive withdrawal that can be easily scored.
We propose here a methodological approach for inducing, recording and quantifying OIH and tolerance in wild-type mice, using the tail-immersion and tail pressure tests. The procedure allows an easy, sensitive and reproducible determination of thermal and mechanical nociceptive response values in mice. As demonstrated in the video protocol, C57BL/6 mice experience significant hyperalgesia following chronic morphine administration and maintain this for several days. Both thermal and mechanical nociceptive values are significantly reduced, compared to baseline measurements on naïve animals. Moreover, our experimental set-up allows to monitor, in addition to the development of OIH, the decline of the analgesic response to morphine (tolerance). Presented data support the view that hyperalgesia and tolerance may involve common cellular and molecular mechanisms8,9, although this is disputed in the literature1,10-12. Finally, this protocol may be similarly adapted to genetically modified mice in order to evaluate the role of individual genes in the modulation of pain. It also provides a model system to evaluate the effectiveness of potential therapeutic agents to improve opiate analgesic effects.
Protocol
All experiments were carried out in strict accordance with the European guidelines for the care of laboratory animals (European Communities Council Directive 86/609/ECC) and the ethical guidelines for investigation of experimental pain in conscious animals13. Male C57BL6/N Tac mice (10 weeks, 25 - 30 g) were housed in an institutional animal facility with a care staff in charge of operating the facility in compliance with environmental standards. Animals were housed in groups (maximum five mice per cage) under a 12 hr / 12 hr light / dark cycle at a constant temperature (21 ± 1 °C) with free access to food and water. All experiments were performed at the same period of the day (10:00 am to 4:00 pm) using a cohort of 16 mice. Specific materials and equipment are indicated in the Materials table.
A five-step procedure to monitor morphine induced-hyperalgesia and tolerance
The protocol is divided into five sequential steps (A-E) over a duration of 15 days Figure 1.
1. Mice Handling (Step A; d-7 to d-5)
Handle mice and habituate them to freely enter into the restrainer. This preliminary step reduces stress - thus minimizing any confusion with stress-induced analgesia - and allows the animals to be accustomed to the investigator, handling and manipulation into the mouse restrainer. Each mouse is gently handled for 5 min every day.
2. Basal Nociceptive Response (Step B; d-4 to d-1)
- Measure tail withdrawal latencies using the Tail Immersion Test (TIT):
- Set the thermostat to 48 °C.
- Gently introduce the mouse into the restrainer. Dip the protruding 2/3 end of its tail into the water bath and start the chronometer.
- Stop the chronometer as soon as the mouse withdraws its tail from the hot water and record the latency time (in sec). In the absence of any nociceptive reaction, a 25 sec cut-off is used to prevent tissue damage.
- Replace the mouse in its cage and test the next one until the end of the series.
- Repeat the nociceptive response measurements two more times, taking measurements from the animals in the same order. The nociceptive response latency (sec) for each mouse is determined as the mean value for three successive determinations.
- Measure mechanical responses using the Tail Pressure Test (TPT)
- Gently introduce the mouse into the restrainer and position its tail under the conic tip of the analgesimeter.
- Press the foot-switch to apply uniformly increasing pressure onto the proximal part of the tail until the first nociceptive reaction (struggling, squeaking) occurs. At the moment the animal reacts, record the current force (in grams) that elicits the nociceptive response. In the absence of any reaction, a 600 g cut-off value is used to avoid tissue damage.
- Repeat this measure on the median and distal parts of the tail of the same mouse. An interval of at least 30 sec is observed between measures on a given mouse to avoid adaptation or stress bias. Replace the mouse in its cage and test the next animal until the end of the series (i.e. all mice are tested). The nociceptive value (grams) for each mouse is taken as the mean value for three measurements (i.e. proximal, median and distal parts of the tail of each animal).
Repeat nociceptive testing (all the procedures outlined under step 2) on subsequent days, d-3 to d-1.
3. Measurement of Morphine Analgesia (Step C; d0)
Define the best combination of animals that allows the selection of two groups (n=8 per group) of mice with stable and comparable average nociceptive values, whichever nociceptive modality (TIT or TPT) is considered. This value will be taken as the basal nociceptive response of reference for future ‘Saline’ and ‘Morphine’ groups.
Measure the body weight of each animal.
Prepare a morphine solution (0.5 mg morphine per ml) in sterile physiological saline (NaCl 0.9%) for subcutaneous administration (5 mg morphine per kg of animal’s body weight).
Measure the nociceptive response latency (taken as time point 0) for each mouse of both ‘Saline’ and ‘Morphine’ groups in the TIT (all steps under 2.1 above). Then measure nociception in the TPT (all steps under 2.2 above).
Inject subcutaneously morphine (typically 0.25 ml of a 0.5 mg/ml morphine solution per 25 g mouse weight) and saline (0.25 ml per 25 g mouse weight) to the ‘morphine’ and ‘saline’ groups, respectively.
- Measure nociceptive values on the TIT and TPT (all procedures outlined above under steps 2.1 and 2.2, respectively) over a time-course (at 30 min interval) to assess morphine (5 mg/kg)-induced analgesia:
- After 30 min post-injection, measure the nociceptive response (single determination) for each mouse of the ‘Saline’ and ‘Morphine’ groups, using the TIT then the TPT.
- Then measure nociceptive response values (TIT and TPT) in all mice at time points (in hr): 1-1.5-2-2.5-3 and 3.5 post-injection.
4. Chronic Morphine Treatment - Morphine-induced Hyperalgesia (Step D; d1 to d6)
- On Day: d1
- Measure nociceptive response values on the TIT and TPT as described above (steps 2.1 and 2.2). Carefully annotate withdrawal latencies and pressure limits for each animal.
- Prepare a fresh morphine solution as detailed in step 3.2.
- Inject subcutaneously morphine (5 mg/kg b.w.) to the entire ‘Morphine’ group and physiological saline (0.25 ml per 25 g mouse weight) to the ‘Saline’ group. Let the animals rest until the next day.
On Days: d2, d3, d4, d5 and d6 repeat the operations described under Section 4.1
5. Evidence for Analgesic Tolerance (Step E; d7)
Evaluate morphine-induced analgesia according to the time-course paradigm already detailed in Section 3.
6. Data Acquisition and Statistical Analyses
- Assessment of Basal Nociceptive Response Values (Step B)
- Calculate for each day (over the d-4 to d-1 period) the mean ± SEM values (n=8) for basal nociceptive responses as afforded from TIT and TPT within the ‘Saline’ and ‘Morphine’ groups.
- Plot mean basal nociceptive values as a function of time (day) for both groups Figure 2.
- Analysis of Morphine Analgesia Time-course at Days d0 (Step C) and d7 (Step E)
- Calculate, at each time point after morphine injection, the mean ± SEM values (n=8) for nociceptive responses as afforded from TIT (in sec) and TPT (in g) in each group.
- Plot mean nociceptive response values as a function of time for the ‘Saline’ and ‘Morphine’ groups at day 0 Figure 3 and day 7 Figure 5.
- Development of Morphine-induced Hyperalgesia (Step D)
- Calculate for each day (over the d0-d7 treatment period) the mean ± SEM values (n=8) for basal nociceptive response values as afforded from TIT and TPT within the ‘Saline-treated’ and ‘Morphine-treated’ groups.
- Plot mean basal nociceptive response values as a function of time (day) for the ‘Saline-treated’ and ‘Morphine-treated’ groups Figure 4.
- Evidence for Analgesic Tolerance (Steps C and E)
- Determine from the morphine time-course experiment performed at d0 Figure 3 the time value (or time range) required for morphine to induce a maximal analgesic response.
- Take this value (usually 30 min) as the reference time to estimate at d7 Figure 5 the nociception baseline value (saline-treated group) and the actual analgesic response (morphine-treated group) to acute morphine.
- Nociceptive values taken at time point 30 min from morphine time-course experiments performed at d0 and d7 for the saline-treated and morphine-treated groups, are presented as histograms Figure 6.
- Statistics: Analyze data using one-way repeated measures ANOVA. The factors of variation were treatment (between subjects) and time (within subject). To check differences separately in each group, repeated measures ANOVA was performed. Comparisons between two groups were performed using unpaired t-test or paired t-test when appropriate.
The Level of Significance is Set at P<0.05. All Statistical Analyses are Carried out using the STATview Software.
Representative Results
Assessment of Basal Nociceptive Values of Naïve Mice (Step B)
TIT and TPT were sequentially applied to the whole cohort of mice (n=16), providing mean nociceptive response values. Best combination of animals allowed an a posteriori definition of two groups (n=8) of mice, referred to as Saline and Morphine, which display similar and stable basal nociceptive values Figure 2. The equivalence of both groups is valid whatever the nociceptive test (TIT: Figure 2A; TPT : Figure 2B) that was selected.
Time-course for Morphine Analgesia at Day 0 (Step C)
Morphine analgesia was evaluated following a single injection (s.c.) of morphine (5 mg/kg) in naïve mice using both TIT Figure 3A and TPT Figure 3B. In both tests, statistical analyses with one way repeated measures ANOVA reveal that there is a significant interaction between treatment and time for TIT (F(7, 98) = 72, p<0.001) and TPT (F(7, 98) = 31, p<0.001). TIT and TPT data analyses using repeated measures ANOVA indicate that there is no effect of saline injection (F(7, 49) = 0.49, p>0.05) and F(7, 49) = 1.85, p>0.05 respectively for TIT and TPT tests), whereas morphine injection induces a strong analgesia in mice (F (7, 49) = 92.46, p<0.001) and F(7, 49) =34.37, p<0.001 respectively for TIT and TPT tests). The maximal analgesic effect of morphine was reached after 30 min in TIT and after 60 min in TPT as compared to saline-injected controls (p<0.001, unpaired t-test).
Repeated Morphine Administrations Induces Hyperalgesia in Mice (Step D)
Basal nociceptive values were measured every day before saline or morphine administration (see protocol). As shown in Figure 4, once daily morphine administrations over a 6 days treatment period induced a significant and progressive lowering of thermal (F(7, 56) = 11.6, p<0.001, repeated measures ANOVA; Figure 4A) and mechanical (F(7,56) = 15,55, p<0.001, repeated measures ANOVA; Figure 4B) basal nociceptive values. Hyperalgesia developed rapidly as it started to be significant at day 1 in TIT (p<0.01, unpaired t-test, compared to saline-injected controls) and at day 2 in TPT (p<0.05, unpaired t-test, compared to saline-injected controls).
Time-course for Morphine Analgesia at Day 7, after Chronic Morphine Treatment (Step E)
On day 7, mice that received daily morphine or saline injections over a 7-day period (d0 to d6) were examined in TIT Figure 5A and TPT Figure 5B first for their basal nociceptive values and then for their analgesic response to acute morphine (5 mg/kg, sc.). In agreement with the development of hyperalgesia shown in Figure 4, the basal nociceptive value (time 0) of mice that were chronically treated with morphine was significantly lower than that of saline-injected control mice (p<0.001, unpaired t-test). Following acute morphine, the nociceptive response of the chronic morphine-treated group significantly increased, but only slightly exceeded the basal nociceptive value of saline-injected control mice measured at 30 min in TIT and TPT (p<0.01 and p<0.05, unpaired t-test, respectively) and at 60 min in TIT (p<0.05; unpaired t-test). From 2 hr after morphine treatment until the end of experiment, nociceptive responses returned to lower values than those of control mice (p<0.001, unpaired t-test).
Comparison of Maximum Analgesic Responses of Mice to Morphine before (Day 0) and after Chronic Morphine Treatment (Day 7).
Nociceptive threshold values presented in Figure 6 are from TIT (A) and TPT (B) performed 30 min after saline or morphine injection as illustrated in Figures 3 (day 0) and 5 (day 7). A strong decrease in morphine analgesia was observed in mice after chronic morphine treatment for 7 days in comparison with their initial analgesia response at day 0 in both nociceptive tests (p<0.001 paired t-test). These data demonstrate that tolerance did develop in pain hypersensitive animals.
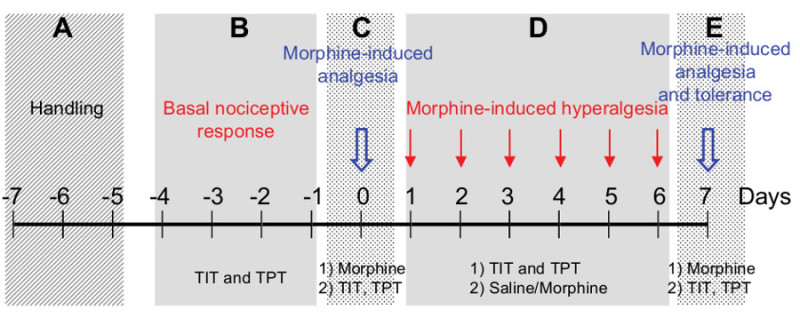 Figure 1.A five-step procedure to monitor morphine induced-hyperalgesia and tolerance. The protocol is divided into five sequential steps (A-E) over a total duration of 15 days.
Figure 1.A five-step procedure to monitor morphine induced-hyperalgesia and tolerance. The protocol is divided into five sequential steps (A-E) over a total duration of 15 days.
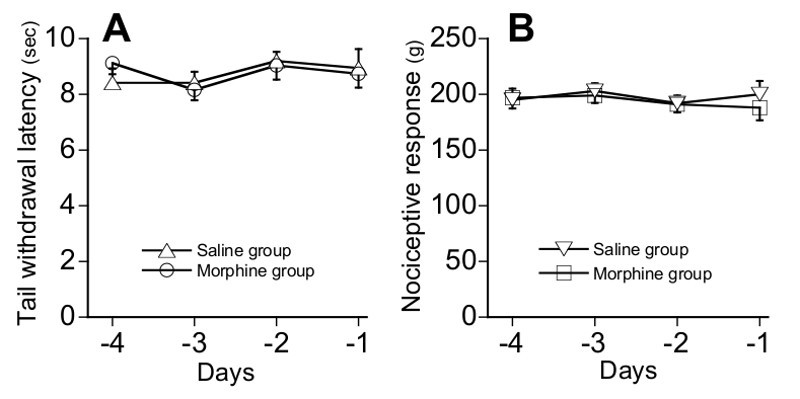 Figure 2.Definition of the basal nociceptive response values (Step B; d-4 to d-1). The tail immersion (TIT) (A) and tail pressure (TPT) (B) tests are applied to the whole series of animals in order to evaluate their basal nociceptive values. Thereafter, two groups of mice (n=8), referred to as ‘Saline’ and ‘Morphine’ groups, are defined so that they exhibit stable and comparable average nociceptive values, whatever the nociceptive modality that is considered.
Figure 2.Definition of the basal nociceptive response values (Step B; d-4 to d-1). The tail immersion (TIT) (A) and tail pressure (TPT) (B) tests are applied to the whole series of animals in order to evaluate their basal nociceptive values. Thereafter, two groups of mice (n=8), referred to as ‘Saline’ and ‘Morphine’ groups, are defined so that they exhibit stable and comparable average nociceptive values, whatever the nociceptive modality that is considered.
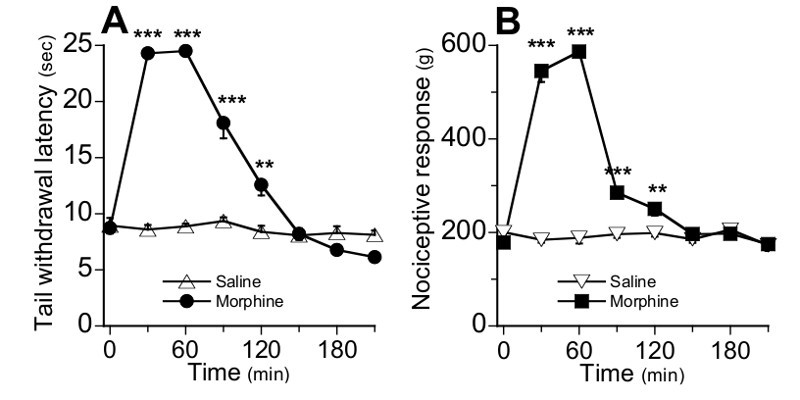 Figure 3. Time-course for morphine analgesia at day 0 (Step C) in TIT (A) and TPT (B). The basal nociceptive response value of mice was determined every 30 min after single morphine (5 mg/kg, sc.) or saline injections. Data are expressed as means ± SEM, n=8 mice per group.*P < 0.05, **P < 0.01, ***P < 0.001, unpaired t-test as compared to control group.
Figure 3. Time-course for morphine analgesia at day 0 (Step C) in TIT (A) and TPT (B). The basal nociceptive response value of mice was determined every 30 min after single morphine (5 mg/kg, sc.) or saline injections. Data are expressed as means ± SEM, n=8 mice per group.*P < 0.05, **P < 0.01, ***P < 0.001, unpaired t-test as compared to control group.
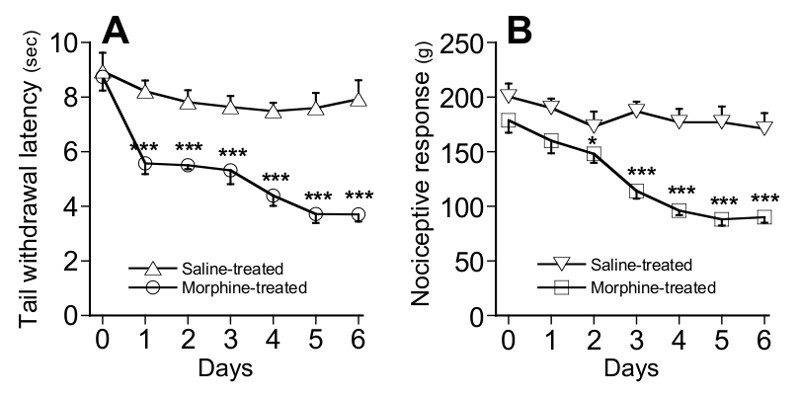 Figure 4.Development of hyperalgesia following repeated morphine administration (Step D; d1 to d6). The basal nociceptive value of mice was determined by TIT(A) and TPT (B) once daily before morphine (5 mg/kg, sc.) or saline administration. Data are expressed as mean values ± SEM, n=8 mice per group. *P < 0.05, **P < 0.01, ***P < 0.001 by unpaired t-test as compared with the saline-treated control group.
Figure 4.Development of hyperalgesia following repeated morphine administration (Step D; d1 to d6). The basal nociceptive value of mice was determined by TIT(A) and TPT (B) once daily before morphine (5 mg/kg, sc.) or saline administration. Data are expressed as mean values ± SEM, n=8 mice per group. *P < 0.05, **P < 0.01, ***P < 0.001 by unpaired t-test as compared with the saline-treated control group.
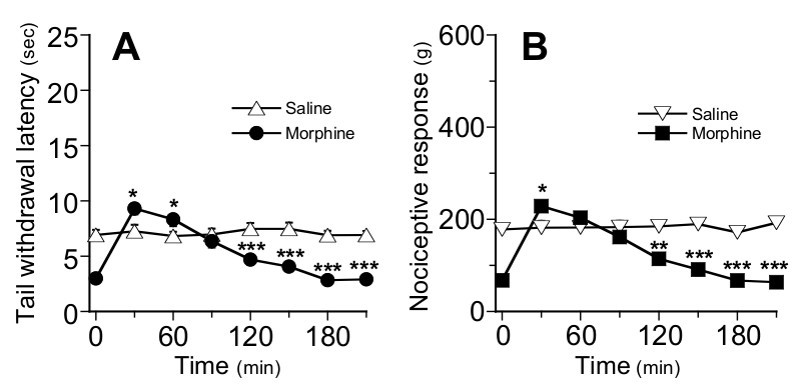 Figure 5.Time-course for morphine analgesia in chronic-morphine treated mice at day 7 (Step E) in TIT (A) and TPT (B). Mice that were chronically treated with morphine (black dots) or saline (white triangles) from day 0 to day 6, received on day 7 a single injection of morphine (5 mg/kg, sc.) or saline, respectively. The nociceptive response of mice was determined every 30 min after morphine or saline injection. Data are expressed as means ± SEM, n=8 mice per group. *P < 0.05, **P < 0.01 by unpaired t-test as compared with the saline-treated control group. Error bars that do not exceed symbols size are hidden.
Figure 5.Time-course for morphine analgesia in chronic-morphine treated mice at day 7 (Step E) in TIT (A) and TPT (B). Mice that were chronically treated with morphine (black dots) or saline (white triangles) from day 0 to day 6, received on day 7 a single injection of morphine (5 mg/kg, sc.) or saline, respectively. The nociceptive response of mice was determined every 30 min after morphine or saline injection. Data are expressed as means ± SEM, n=8 mice per group. *P < 0.05, **P < 0.01 by unpaired t-test as compared with the saline-treated control group. Error bars that do not exceed symbols size are hidden.
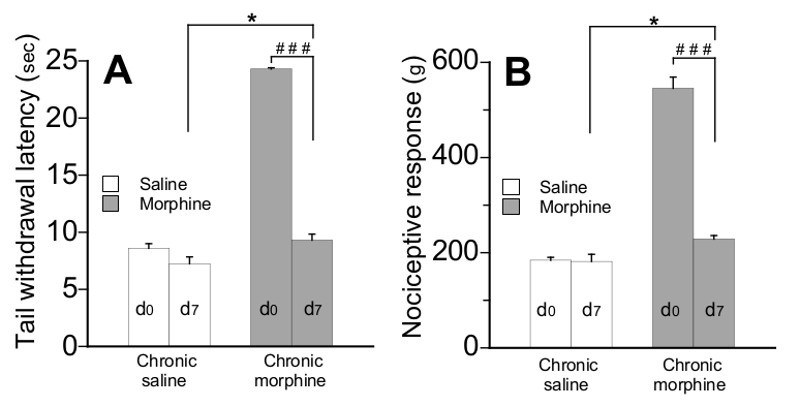 Figure 6.Comparison of maximum analgesic responses of mice to morphine (5 mg/kg, sc.) before (day 0) and after chronic morphine treatment (day 7). Values reported here are from experiments shown in Figure 3 and Figure 5. Nociceptive values were measured using TIT (A) and TPT (B) 30 min after morphine or saline injection. Data are expressed as means ± SEM, n=8 mice per group. ***P < 0.001 by paired t-test.
Figure 6.Comparison of maximum analgesic responses of mice to morphine (5 mg/kg, sc.) before (day 0) and after chronic morphine treatment (day 7). Values reported here are from experiments shown in Figure 3 and Figure 5. Nociceptive values were measured using TIT (A) and TPT (B) 30 min after morphine or saline injection. Data are expressed as means ± SEM, n=8 mice per group. ***P < 0.001 by paired t-test.
Discussion
Critical Steps
Choice of the animal model for nociception measurements
Variability in nociceptive and analgesic sensitivity among mice strains has been examined (reviews 14-16) reviews using various pain models differing in their etiology (nociceptive, inflammatory, neuropathic), modality (thermal, chemical, mechanical), duration (acute, tonic, chronic) and site of administration (cutaneous, subcutaneous, visceral). When compared to other strains, C57BL/6J (“J” for Jackson Laboratory) mice became a popular animal model for pain studies as they exhibit a high basal nociceptive sensitivity17,18 and a moderate analgesic response to opiates14,19. Following chronic morphine treatment, they also develop significant analgesic tolerance20,21, hyperalgesia21,22 and dependence20,23.
Here, experiments were performed on C57BL/6N Tac mice (“N” for National Institute of Health and “Tac’ for Taconics farm) which belong to a separate branch of the B6 lineage. Although C57BL/6 mice have been long considered as interchangeable, recent studies pointed to significant behavioral differences among C57BL/6J and C57BL/6N strains24. In particular, the lower sensitivity of the three C57BL/6N substrains (including the Tac one) to acute thermal pain may be regarded as an advantage for testing this phenotype.
Male mice were selected as the vast majority of pain studies, using mice as the animal model, are performed on juvenile males25. In our hands, they provided robust and reproducible data when examined from the analgesia or hyperalgesia points of view. Occasionally, we noticed a tendency for C57BL/6N females to provide more variable responses, both in the TIT and TPT tests. Although this observation may reflect natural variations linked to the hormonal status of females, overall mechanisms underlying sex differences in pain and analgesia still remain a matter of controversy. Some aspects of this hot debate will be briefly presented in the next ‘Limitations of the technique’ section.
Animal habituation
Mice were first allowed to get accustomed to the animal facility during one week. Similar to any other behavioral study, testing was performed following a 3 day-acclimatization period (Figure 1, step A). As nociceptive tests are sensitive to stress, first measures may give longer latencies than subsequent ones, especially in non-habituated mice26,27. The habituation step allows also the obtention of more stable nociceptive response values within the same day and between days Figures 2 and 4. To reduce circadian effects on nociceptive and analgesic sensitivity28,29, all testings were conducted between 10:00 am and 4:00 pm.
Selection of nociceptive tests
Nociceptive tests use either thermal, mechanical, chemical or electrical stimuli (review 26,27,30. Their choice is critical as different nociceptive modalities may be processed through different nociceptors and fibers18,31,32.
We selected the tail immersion test (TIT)33, a modified version of the classical tail flick test developed by D’amour and Smith34, and the tail pressure test (TPT), adapted from Randall and Selitto35, as examples of thermal and mechanical modalities to study morphine-induced analgesia, hyperalgesia and tolerance in mice. Both tests have been widely used in rats. A cut off time was systematically defined to avoid or limit the risk of tissue damage.
Morphine-induced analgesia, hyperalgesia and tolerance
Morphine, the prototypical mu-opiate agonist, was selected here as it is a potent analgesic and OIH-inducer, both in humans and mice1,2,36. Morphine analgesic potency is known to vary with mice strains, routes of administration and nociceptive modalities. In C57BL/6 mice, reliable analgesia is usually obtained following subcutaneous injections of morphine in the 1-20 mg/kg dose range14,21. Accordingly, we chose to study acute analgesia following a single administration (s.c.) of morphine at 5 mg/kg, close to its ED50 value (7-20 mg/kg) assessed from thermal nociception19,21.
Repeated morphine administration is often accompanied with analgesic tolerance (evidenced either from a rightward shift of the dose-response curve or from a decrease in analgesic response amplitude or duration) and hyperalgesia (an exacerbated sensitivity to painful stimuli evidenced from a decrease in basal nociceptive value). Both adverse phenomena depend on rodent strains, on the nature of the opiate compound which is selected and its dosage, on treatment duration and on nociceptive modalities21. For example, experimental paradigms to study tolerance and hyperalgesia consist in daily administration of high and constant (20 to 40 mg/kg per day)22 or of escalating (up to 50 or even 200 mg/kg)20,21 morphine doses. Accordingly, we promoted the development of hyperalgesia and tolerance in C57BL/6 mice through daily morphine administration (5 mg/kg; s.c.) over a 8-day period. This moderate morphine dose was preferred over higher ones to better mimic clinic usage.
Set up of TIT operational window
A possible pitfall in TIT might be related to the role of the tail in the thermoregulation of rodents26,37. As ambient temperature is a key factor in nociceptive response variations, it should be kept constant (here at 21 °C) throughout experiments38. Heat intensity is usually set up to detect a nociceptive response within 5 to 10 sec27. Indeed, greater latencies may increase the risk for monitoring animal movements unrelated to the nociceptive stimulus, whereas shorter ones may reduce the differential power of the test. We performed TIT measurements at a fixed temperature of 48 °C. Tail withdrawal latencies were close to 9 sec (basal nociceptive value) and varied from 4 sec (hyperalgesia) to 25 sec (maximal analgesia; cut off). In addition to practical reasons, measurements of nociceptive response values at a fixed temperature may a priori involve the same repertoire of nociceptors and circuits, thereby facilitating data interpretation.
Possible Modifications
Optimization of the TIT operational window for analgesia and OIH measurements
When focusing on an analgesic response, low baseline values (higher heat intensity) may favor the detection of a delay in the response. In turn, to address the consequence of a painful stimulus or the development of OIH, higher baseline values (lower heat intensity; here 48 °C) may facilitate the detection of faster responses Figure 4.
Although we found morphine at 5 mg/kg a convenient dose to induce a robust analgesic response Figure 3 and to promote (upon repeated administration) significant hyperalgesia Figure 4, its dosage may be adapted as mentioned before (Critical step : Morphine-induced analgesia, hyperalgesia and tolerance). For example, lower doses may be used to reduce analgesia amplitude (thereby avoiding cut-off limitations) whereas higher doses may be chosen to accelerate hyperalgesia onset and increase its amplitude.
Overall, optimization of the ‘nociceptive window’ should be adapted to the genetic background of mice under study and take into account the possibility for the involvement of distinct arrays of nociceptors and circuits.
Alternative opiate agonists (fentanyl, remifentanyl)
Although most clinically used opiates target the mu-opioid receptor as agonists, they differ considerably with respect to their pharmacological properties both in vitro and in vivo. For example, remifentanyl and fentanyl, in marked contrast with morphine, behave as full agonists and promote internalization of mu-opioid receptors39. Opiate analgesics such as morphine and fentanyl have half-lives in the range of hours40, while remifentanyl has an ultra-short half life of several minutes41. In humans, best evidence for OIH is from patients who received opiates during surgery, including short-acting compounds such as remifentanyl2,42. Thus, fentanyl and remifentanyl may be valuable tools too to study the development of hyperalgesia and tolerance in mice, under TIT and TPT paradigms.
Alternative modes of induction of OIH (chronic vs acute administration)
OIH is seen in humans and animal models as a consequence of opiate administration, whether at very low or extremely high dosages1,2. We report here on OIH development following chronic treatment of mice with moderate doses of morphine. Several days of treatment of C57BL/6N mice were necessary to evidence a clear and reproducible hyperalgesic state Figure 4. Daily morphine injections could be adequately replaced with implanted morphine pellets : upon their removal, both thermal hyperalgesia and mechanical allodynia have been already reported in mice43. Infusion of opiates through a micro-osmotic pump is another possibility44. In rodents, long-lasting hyperalgesia is also achievable following acute administration of fentanyl using a protocol mimicking the use of this mu-opioid agonist in human surgery36,45,46.
Limitations of the Technique
Animal species and models for pain
Comparative studies of numerous mouse strains provided evidence for great variations in nociceptive responses to painful stimuli17,31,47 and in OIH levels following 4-days morphine treatment22. Whether mechanisms underlying pain processing and modulation in animal models (mice and rats) are relevant for chronic pain patients remains a fundamental and opened question. Thus, much caution should be paid to the interpretation of animal data and to their predictive validity for humans16.
Sex differences in pain and analgesia
Most preclinical studies on animal models for pain have been conducted on male rodents16,25,48. Despite this selection bias, the emerging view was to consider males as better responders to opiate analgesics49,50, less prone to develop opioid-induced hyperalgesia 51,52 and more tolerant to morphine analgesia53 than their female counterparts (review 54). However, sex differences with regard to nociception and analgesic drugs efficacy do not resume in such ‘a one size fits all’ paradigm. Indeed, a wealth of data now indicates that numerous variables may influence the magnitude and direction of sex differences such as opioid drug efficacy and selectivity, nociceptive assay, genetic background, age, gonado-hormonal status or social interaction48,54. In humans, clinical pain is more prevalent in women but whether this fact reflects actual sex differences remains a matter of debate48,55,56. For example, global analysis of fifty clinical trials indicated no significant differences in analgesic properties between genders whereas meta-analyses performed on patients-controlled subjects pointed to a significantly greater opioid efficacy in women57.The latter observation, which markedly contrasts with what has been found in rodents, again raises several questions regarding the origin for such divergences16,48,55,57. Altogether, sex differences in analgesia do exist and merit further focus on underlying mechanisms and clinical implications.
About nociceptive tests
The tail withdrawal test is a spinal reflex but it may be subject to supraspinal influences58. TIT is relatively easy to perform on rats but requires more expertise in mice. A potential difficulty is to maintain the mouse in a correct posture without inducing unwanted stress. The proposed protocol may be adjusted according to cohort size. 16 animals (8 control and 8 treated) are easily managed as far as measurement of their basal nociceptive response values (using TIT first, then TPT for the whole series of mice) is under concern. Monitoring analgesia time courses requires the establishment of a precise time schedule and the evaluation of the maximal number of animals that may be tested (TIT first, then TPT) within the imparted time interval (here 30 min). The whole cohort of animals may thus be divided into subgroups to allow the experimenter to respect kinetic limitations.
Significance of the Technique with respect to Existing / Alternative Methods
OIH in rats versus mouse
Rats have been extensively used to study opioid analgesia, hyperalgesia and tolerance, following acute or chronic opiate administration46,59-61. Indeed, for several practical reasons, they may be considered superior to mice as an animal model for pain experiments16,61. However, until recently, the generation of genetically modified rats was not a straightforward procedure. As numerous genetically modified mouse strains are already available, our model offers the opportunity to study the contribution of numerous individual genes in OIH and tolerance development in mice.
TIT and TPT versus other nociceptive tests
TIT is a variant of the tail flick test, the most obvious difference being the area of stimulation. In contrast with radiant heat, immersion of the tail into hot water leads to a quick and uniform increase in its temperature. Compared to other forms of thermal nociception testing (hot-plate or Hargreaves tests), TIT provides fairly reproducible results both across and within subjects.
TPT is a very popular test for the study of mechanical nociception26,27,35 which probably involves distinct nociceptive fibers and molecular transducers then TIT32. It provides quick and reliable measurements59 but requires some expertise from the experimenter and large animal cohorts. As an alternative to the analgesimeter used in the present study, other procedures or apparatus relying on strain gauges do exist (review 27). TPT is best suited for studying mechanical hyperalgesia whereas von Frey filaments are usually taken to evaluate mechanical allodynia (review 27).
Future Applications or Directions after Mastering this Technique
The experimental OIH/tolerance model we present here may be similarly adapted to genetically modified mice in order to evaluate the role of individual genes in the modulation of pain. It also provides a model system to investigate the effectiveness of potential therapeutic agents to relieve chronic pain.
Disclosures
The authors declare that they have no competing financial interests.
Acknowledgments
We thank Dr. J-L. Galzi (UMR7242 CNRS; Illkirch, France) for his support.
This work was supported by the CNRS, INSERM, Université de Strasbourg, Alsace BioValley and by grants from Conectus, Agence National de la Recherche (ANR 08 EBIO 014.02) Conseil Régional d’Alsace (Pharmadol), Communauté Urbaine de Strasbourg (Pharmadol), ICFRC (Pharmadol), OSEO (Pharmadol), Direction Générale des Entreprises (Pharmadol).
References
- Angst MS, Clark JD. Opioid-induced hyperalgesia. A qualitative systematic review. Anesthesiology. 2006;104:570–587. doi: 10.1097/00000542-200603000-00025. [DOI] [PubMed] [Google Scholar]
- Chu LF, Angst MS, Clark D. Opioid-induced hyperalgesia in humans. Molecular mechanisms and clinical considerations. Clin. J. Pain. 2008;24:479–496. doi: 10.1097/AJP.0b013e31816b2f43. [DOI] [PubMed] [Google Scholar]
- Lee M, Silverman S, Hansen H, Patel V, Manchikanti L. A comprehensive review of opioid-induced hyperalgesia. Pain Physician. 2011;14:145–161. [PubMed] [Google Scholar]
- Hutchinson MR, Shavit Y, Grace PM, Rice KC, Maier SF, Watkins LR. Exploring the neuroimmunopharmacology of opioids: An integrative review of mechanisms of central immune signaling and their implications for opioid analgesia. Pharmacol. Rev. 2011;63:772–810. doi: 10.1124/pr.110.004135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang X, et al. Morphine activates neuroinflammation in a manner parallel to endotoxin. Proc. Natl. Acad. Sci. USA. 2012;109:6325–6330. doi: 10.1073/pnas.1200130109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colpaert FC. System theory of pain and of opiate analgesia: No tolerance to opiates. Pharmacol. Rev. 1996;48:355–402. [PubMed] [Google Scholar]
- Simonnet G, Rivat C. Opioid-induced hyperalgesia: abnormal or normal pain. NeuroReport. 2003;14:1–7. doi: 10.1097/00001756-200301200-00001. [DOI] [PubMed] [Google Scholar]
- Mao J, Sung B, Ji RR, Lim G. Chronic morphine induces downregulation of spinal glutamate transporters: implications in morphine tolerance and abnormal pain sensitivity. J. Neurosci. 2002;22:8312–8323. doi: 10.1523/JNEUROSCI.22-18-08312.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- King T, Ossipov MH, Vanderah TW, Porreca F, Lai J. Is paradoxical pain induced by sustained opioid exposure an underlying mechanism of opioid antinociceptive tolerance. Neurosignals. 2005;14:194–205. doi: 10.1159/000087658. [DOI] [PubMed] [Google Scholar]
- DuPen A, Shen D, Ersek M. Mechanisms of opioid-induced tolerance and hyperalgesia. Pain Management Nursing. 2007;8:113–121. doi: 10.1016/j.pmn.2007.02.004. [DOI] [PubMed] [Google Scholar]
- Chu LF, et al. Analgesic tolerance without demonstrable opioid-induced hyperalgesia : A double-blinded, randomized, placebo-controlled trial of sustained-release morphine for treatment of chronic nonradicular low-back. 2012;153:1583–1592. doi: 10.1016/j.pain.2012.02.028. [DOI] [PubMed] [Google Scholar]
- Ferrini F, et al. Morphine hyperalgesia gated through microglia-mediated disruption of neuronal Cl- homeostasis. Nature Neurosci. 2013;16:183–192. doi: 10.1038/nn.3295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zimmermann M. Ethical guidelines for investigation of experimental pain in conscious animals. Pain. 1983;16:109–110. doi: 10.1016/0304-3959(83)90201-4. [DOI] [PubMed] [Google Scholar]
- Mogil JS. The genetic mediation of individual differences in sensitivity to pain and its inhibition. Proc. Natl. Acad. Sci. USA. 1999;96:7744–7751. doi: 10.1073/pnas.96.14.7744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mogil JS, et al. Screening for pain phenotypes : Analysis of three congenic mouse strains on a battery of nine nociceptive tests. Pain. 2006;126:24–34. doi: 10.1016/j.pain.2006.06.004. [DOI] [PubMed] [Google Scholar]
- Mogil JS. Animal models of pain : progress and challenges. Nature Rev. Neurosci. 2009;10:283–294. doi: 10.1038/nrn2606. [DOI] [PubMed] [Google Scholar]
- Mogil JS, et al. Heritability of nociception I : Responses of 11 inbred mouse strains on 12 measures of nociception. Pain. 1999;80:67–82. doi: 10.1016/s0304-3959(98)00197-3. [DOI] [PubMed] [Google Scholar]
- Larivière WR, et al. Heritability of nociception. III. Genetic relationships among commonly used assays of nociception and hypersensitivity. Pain. 2002;97:75–86. doi: 10.1016/s0304-3959(01)00492-4. [DOI] [PubMed] [Google Scholar]
- Elmer GI, Pieper JO, Negus SS, Woods JH. Genetic variance in nociception and its relationship to the potency of morphine-induced analgesia in thermal and chemical tests. Pain. 1998;75:129–140. doi: 10.1016/S0304-3959(97)00215-7. [DOI] [PubMed] [Google Scholar]
- Eidelberg E, Erspamer R, Kreinick CJ, Harris J. Genetically determined differences in the effects of morphine on mice. Eur. J. Pharmacol. 1975;32:329–336. doi: 10.1016/0014-2999(75)90300-3. [DOI] [PubMed] [Google Scholar]
- Kest B, Hopkins E, Palmese CA, Adler M, Mogil JS. Genetic variation in morphine analgesic tolerance: A survey of 11 inbred mouse strains. Pharmacol. Biochem. Behav. 2002;73:821–828. doi: 10.1016/s0091-3057(02)00908-5. [DOI] [PubMed] [Google Scholar]
- Liang D-Y, Liao G, Wang J, Usuka J, Guo Y, Peltz G, Clark JD. A genetic analysis of opioid-induced hyperalgesia in mice. Anesthesiology. 2006;104:1054–1062. doi: 10.1097/00000542-200605000-00023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kest B, Palmese CA, Hopkins E, Adler M, Juni A, Mogil JS. Naloxone-precipitated withdrawal jumping in 11 inbred mouse strains : Evidence for common genetic mechanisms in acute and chronic morphine physical dependence. Neurosci. 2002;115:463–469. doi: 10.1016/s0306-4522(02)00458-x. [DOI] [PubMed] [Google Scholar]
- Bryant CD, et al. Behavioral differences among C57BL/6 substrains: Implications for transgenic and knockout studies. J. Neurogenet. 2008;22:315–331. doi: 10.1080/01677060802357388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mogil JS, Chanda ML. The case for the inclusion of female subjects in basic science studies of pain. Pain. 2005;117:1–5. doi: 10.1016/j.pain.2005.06.020. [DOI] [PubMed] [Google Scholar]
- Le Bars D, Gozariu M, Cadden SW. Animal models of nociception. Pharm. Rev. 2001;53:597–652. [PubMed] [Google Scholar]
- Barrot M. Tests and models of nociception and pain in rodents. Neurosci. 2012;211:39–50. doi: 10.1016/j.neuroscience.2011.12.041. [DOI] [PubMed] [Google Scholar]
- Kavaliers M, Hirst M. Daily rythms of analgesia in mice: effects of age and photoperiod. Brain Res. 1983;279:387–393. doi: 10.1016/0006-8993(83)90216-0. [DOI] [PubMed] [Google Scholar]
- Castellano C, Puglisi-Allegra S, Renzi P, Oliverio A. Genetic differences in daily rhythms of pain sensivity in mice. Pharmacol. Biochem., and Behavior. 1985;23:91–92. doi: 10.1016/0091-3057(85)90135-2. [DOI] [PubMed] [Google Scholar]
- Sandkühler J. Models and mechanisms of hyperalgesia and allodynia. Physiol. Rev. 2009;89:707–758. doi: 10.1152/physrev.00025.2008. [DOI] [PubMed] [Google Scholar]
- Mogil JS, et al. Heritability of nociception II. ‘Types’ of nociception revealed by genetic correlation analysis. Pain. 1999;80:83–93. doi: 10.1016/s0304-3959(98)00196-1. [DOI] [PubMed] [Google Scholar]
- Scherrer G, et al. Dissociation of the opioid receptor mechanisms that control mechanical and heat. 2009;137:1148–1159. doi: 10.1016/j.cell.2009.04.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janssen PAJ, Niemegeers CJE, Dony JGH. The inhibitory effect of fentanyl and other morphine-like analgesics on the warm water induced tail withdrawal reflex. Arzneimittelforsch. 1963;13:502–507. [PubMed] [Google Scholar]
- Amour FE, Smith DL. A method for determining loss of pain sensation. J. Pharmacol. Exp. Ther. 1941;72:74–79. [Google Scholar]
- Randall LO, Selitto JJ. A method for measurement of analgesic activity on inflamed tissue. Arch. Int. Pharmacodyn. Ther. 1957;111:409–419. [PubMed] [Google Scholar]
- Elhabazi K, et al. Involvement of neuropeptides FF receptors in neuroadaptative responses to acute and chronic opiate treatments. Br. J. Pharmacol. 2012;165:424–435. doi: 10.1111/j.1476-5381.2011.01563.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berge O-G, Garcia-Cabrera I, Hole K. Response latencies in the tail-flick test depend on tail skin temperature. Neurosci. Lett. 1988;86:284–288. doi: 10.1016/0304-3940(88)90497-1. [DOI] [PubMed] [Google Scholar]
- Benoist J-M, Pincedé I, Ballantyne K, Plaghi L, Le Bars D. Peripheral and central determinants of a nociceptive reaction: An approach to psychophysics in the rat. PLoS ONE. 2008;3:e3125. doi: 10.1371/journal.pone.0003125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morgan MM, Christie MJ. Analysis of opioid efficacy, tolerance, addiction and dependence from cell culture to human. Br. J. Pharmacol. 2011;164:1322–1334. doi: 10.1111/j.1476-5381.2011.01335.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trescot AM, Datta S, Lee M, Hansen H. Opioid pharmacology. Pain Physician. 2008;11:S133–S153. [PubMed] [Google Scholar]
- Egan TD, et al. The pharmacokinetics of the new short-acting opioid remifentanil (GI87084B) in healthy adult male volunteers. Anesthesiology. 1993;79:881–892. doi: 10.1097/00000542-199311000-00004. [DOI] [PubMed] [Google Scholar]
- Hansen EG, Duedahl TH, Rømsing J, Hilsted KL, Dahl JB. Intra-operative remifentanil might influence pain levels in the immediate post-operative period after major abdominal surgery. Acta Anaesthesiol Scand. 2005;49:1464–1470. doi: 10.1111/j.1399-6576.2005.00861.x. [DOI] [PubMed] [Google Scholar]
- Li X, Angst MS, Clark JD. A murine model of opioid-induced hyperalgesia. Mol. Brain Res. 2001;86:56–62. doi: 10.1016/s0169-328x(00)00260-6. [DOI] [PubMed] [Google Scholar]
- Varnado-Rhodes Y, Gunther J, Terman GW, Chavkin C. Mu opioid analgesia and analgesic tolerance in two mouse strains. C57BL/6 and 129/SvJ. Proc. West Pharmacol. Soc. 2000;43:15–17. [PubMed] [Google Scholar]
- Celerier E, et al. Long-lasting hyperalgesia induced by fentanyl in rats: preventive effect of ketamine. Anesthesiology. 2000;92:465–472. doi: 10.1097/00000542-200002000-00029. [DOI] [PubMed] [Google Scholar]
- Celerier E, Simonnet G, Maldonado R. Prevention of fentanyl-induced delayed pronociceptive effects in mice lacking the protein kinase C gamma gene. Neuropharmacol. 2004;46:264–272. doi: 10.1016/j.neuropharm.2003.08.008. [DOI] [PubMed] [Google Scholar]
- Larivière WR, Chesler EJ, Mogil JS. Transgenic studies of pain and analgesia: Mutation or background phenotype. J. Pharmacol. Exp. Ther. 2001;297:467–473. [PubMed] [Google Scholar]
- Mogil JS. Sex differences in pain and pain inhibition: multiple explanations of a controversial phenomenon. Nature Rev. Neurosci. 2012;13:859–866. doi: 10.1038/nrn3360. [DOI] [PubMed] [Google Scholar]
- Kest B, Wilson SG, Mogil JS. Sex differences in supraspinal morphine analgesia are dependent on genotype. J. Pharmacol. Exp. Ther. 1999;289:1370–1375. [PubMed] [Google Scholar]
- Kest B, Sarton E, Dahan A. Gender differences in opioid-mediated analgesia. Anesthesiology. 2000;93:539–547. doi: 10.1097/00000542-200008000-00034. [DOI] [PubMed] [Google Scholar]
- Holtman JR, Wala EP. Characterization of morphine-induced hyperalgesia in male and female rats. Pain. 2005;114:62–70. doi: 10.1016/j.pain.2004.11.014. [DOI] [PubMed] [Google Scholar]
- Juni A, et al. Sex differences in hyperalgesia during morphine infusion: effect of gonadectomy and estrogen treatment. Neuropharmacol. 2008;54:1264–1270. doi: 10.1016/j.neuropharm.2008.04.004. [DOI] [PubMed] [Google Scholar]
- Craft RM, et al. Sex differences in development of morphine tolerance and dependence in the rat. Psychopharmacol. 1999;143:1–7. doi: 10.1007/s002130050911. [DOI] [PubMed] [Google Scholar]
- Bodnar RJ, Kest B. Sex differences in opioid analgesia, hyperalgesia, tolerance and withdrawal: central mechanisms of action and roles of gonadal hormones. Hormones Behav. 2010;58:72–81. doi: 10.1016/j.yhbeh.2009.09.012. [DOI] [PubMed] [Google Scholar]
- Greenspan JD, et al. Studying sex and gender differences in pain and analgesia: A consensus report. Pain. 2007;132:S26–S45. doi: 10.1016/j.pain.2007.10.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fillingim RB, Ness TJ. Sex-related hormonal influences on pain and analgesic responses. Neurosci. Biobehav. Rev. 2000;24:485–501. doi: 10.1016/s0149-7634(00)00017-8. [DOI] [PubMed] [Google Scholar]
- Niesters M, et al. Do sex differences exist in opioid analgesia? A systematic review and meta-analysis of human experimental and clinical studies. Pain. 2010;151:61–68. doi: 10.1016/j.pain.2010.06.012. [DOI] [PubMed] [Google Scholar]
- Millan MJ. Descending control of pain. Prog. Neurobiol. 2002;66:355–474. doi: 10.1016/s0301-0082(02)00009-6. [DOI] [PubMed] [Google Scholar]
- Celerier E, Laulin J-P, Corcuff J-B, Le Moal M, Simonnet G. Progressive enhancement of delayed hyperalgesia induced by repeated heroin administration : A sensitization process. J. Neurosci. 2001;21:4074–4080. doi: 10.1523/JNEUROSCI.21-11-04074.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simonin F, et al. RF9, a potent and selective neuropeptide FF receptor antagonist, prevents opioid-induced tolerance associated with hyperalgesia. Proc. Natl. Acad. Sci. U.S.A. 2006;103:466–471. doi: 10.1073/pnas.0502090103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson SG, Mogil JS. Measuring pain in the (knockout) mouse: big challenges in a small mammal. Behav. Brain Res. 2001;125:65–73. doi: 10.1016/s0166-4328(01)00281-9. [DOI] [PubMed] [Google Scholar]