

Abstract
To enter its human host, herpes simplex virus type 1 (HSV-1) must overcome the barrier of mucosal surfaces, skin, or cornea. HSV-1 targets keratinocytes during initial entry and establishes a primary infection in the epithelium, which is followed by latent infection of neurons. After reactivation, viruses can become evident at mucocutaneous sites that appear as skin vesicles or mucosal ulcers. How HSV-1 invades skin or mucosa and reaches its receptors is poorly understood. To investigate the invasion route of HSV-1 into epidermal tissue at the cellular level, we established an ex vivo infection model of murine epidermis, which represents the site of primary and recurrent infection in skin. The assay includes the preparation of murine skin. The epidermis is separated from the dermis by dispase II treatment. After floating the epidermal sheets on virus-containing medium, the tissue is fixed and infection can be visualized at various times postinfection by staining infected cells with an antibody against the HSV-1 immediate early protein ICP0. ICP0-expressing cells can be observed in the basal keratinocyte layer already at 1.5 hr postinfection. With longer infection times, infected cells are detected in suprabasal layers, indicating that infection is not restricted to the basal keratinocytes, but the virus spreads to other layers in the tissue. Using epidermal sheets of various mouse models, the infection protocol allows determining the involvement of cellular components that contribute to HSV-1 invasion into tissue. In addition, the assay is suitable to test inhibitors in tissue that interfere with the initial entry steps, cell-to-cell spread and virus production. Here, we describe the ex vivo infection protocol in detail and present our results using nectin-1- or HVEM-deficient mice.
Keywords: Immunology, Issue 102, Virology, herpes simplex virus type 1, ex vivo infection, murine epidermis, viral entry, viral spread
Introduction
Herpes simplex virus (HSV) can cause a range of diseases in humans from mild uncomplicated mucocutaneous lesions to life-threatening infections. HSV type 1 (HSV-1) is dominantly associated with orofacial infections and encephalitis, whereas HSV type 2 (HSV-2) more likely causes genital infections1. While there is significant progress in understanding how HSV enters cells in culture, initiates infection and produces viral progeny, we know little about the viral invasion pathway(s) into tissue at the cellular level2. For studies of HSV skin or mucosal infections, mice, rabbits and guinea pigs have been used as animal models. Skin infection was established by intradermal injection or by scratching the skin in the presence of virus, and disease development was correlated with virus production. These methods helped to understand various aspects of disease pathogenesis, and are used to evaluate antiviral drugs. To study HSV infection at the tissue level, organotypic human skin models have been applied. As the rate of infection is restricted in these raft cultures, only a limited number of studies investigating infection, viral spread and the effects of antiviral components have been published3-6.
In order to characterize cellular determinants that play a role during HSV-1 infection in the intact epithelium, we established a protocol for ex vivo infection studies of murine epidermis7. Skin was prepared from newborns or from the tails of adult mice. Since HSV-1 could not infect complete skin samples, which were submerged in virus-containing medium, we separated the epidermis from the dermis by dispase II treatment. After floating of the epidermal sheets on virus-containing medium, infected cells can be visualized in the epidermal basal layer at various times postinfection (p.i.)7. To visualize the initiation of infection in individual cells prior to viral replication and virus production, we stained with an antibody against the infected-cell protein 0 (ICP0), which is one of the first proteins expressed during HSV-1 infection. The cellular localization of ICP0 passes through distinct phases during early infection. While ICP0 is present in nuclear foci during an early stage of viral gene expression, relocalization of ICP0 to the cytoplasm indicates a subsequent phase of infection8.
We used the ex vivo infection assay of epidermal sheets from different mouse models to test the potential role of various cellular factors during infection. To address the impact of Rac1 as a key regulator of actin dynamics, we infected the epidermis of mice with a keratinocyte-specific deletion of the rac1 gene9. This model allowed us to study the consequences of deficient Rac1 on the efficiency of HSV-1 infection in epidermal keratinocyte layers. The comparison to infected epidermis of control littermates revealed no significant difference, indicating that the absence of Rac1 had no effect on the initiation of infection in the basal layer of the epidermis7. The use of further mouse models allowed us to address which cellular receptors mediate entry into the epidermis. Infecting epidermal sheets from either nectin-1- or HVEM-deficient mice with HSV-1 revealed that the initial viral entry into tissue strongly depends on the presence of nectin-110. Furthermore, our results demonstrate that HVEM can also serve as receptor in murine epidermis, although less efficiently than nectin-110.
To address the spatial distribution of infected cells in the epidermal layers, we visualize ICP0 expression in tissue sections and epidermal whole mounts (Figure 1). In cryosections of complete skin, no ICP0-expressing cells are detected (Figure 1). In contrast, cryosections of epidermal sheets demonstrate cytoplasmic ICP0 expression in the basal layer already at 3 hours p.i. (Figure 1). At later times, viral spreading to suprabasal layers can be visualized. The spatial distribution of infected cells in the basal layer can be easily followed in epidermal whole mounts (Figure 1). Upon infection with HSV-1 at 100 PFU/cell, approximately 50% of the basal keratinocytes in the interfollicular epidermis show ICP0 expression at 1.5 hr p.i. At this time point most infected cells express nuclear ICP0. The relocalization of ICP0 to the cytoplasm indicating a later stage of early gene expression is present in nearly all cells at 3 hr p.i. (Figure 1). These modes of visualizing infected cells upon ex vivo HSV-1 infection provide a powerful assay to study the effect of inhibitors or of deleted/mutated cellular components on viral entry and spread in tissue.
Protocol
Ethics statement.
The preparation of epidermal sheets from sacrificed animals is carried out in strict accordance with the recommendations of the Guide of Landesamt für Natur, Umwelt and Verbraucherschutz, Northrhine-Westphalia (Germany). The study was approved by LANUV NRW (Number 8.84-02.05.20.13.018).
1. Preparation of Instruments and Culture Media
Cultivate epidermal sheets in Dulbecco’s modified Eagle’s medium (DMEM)/high glucose containing glutamine dipeptide, 10% FCS, 100 IU/ml penicillin, and 100 µg/ml streptomycin (hereinafter referred to as “DMEM”).
For the preparation of epidermal sheets, carefully dissolve dispase II powder (5 mg/ml) in heated PBS (up to 37 °C). Filter the solution through a 0.22 µm filter and use it directly for treatment.
For the preparation of epidermal whole mounts13, prepare blocking buffer with 0.5% milk powder, 0.25% gelatin from cold water fish skin and 0.5% Triton X-100 in TBS. Allow substances to dissolve by incubating the mixture for at least 2 hr at RT on a rotating mixer. Also prepare 0.2% Tween 20 in PBS as washing buffer. For fixation, prepare 3.4% and 4% formaldehyde in PBS.
For the immunostaining of cryosections, prepare blocking buffer with 5% normal goat serum and 0.2% Tween 20 in PBS. For fixation, prepare 0.5% formaldehyde in PBS.
2. Preparation of Epidermal Sheets from Murine Skin
- Preparation of epidermis from newborn back skin for infection studies
- Place a decapitated mouse (1 to 3 days after birth) into a dissection dish and cut off extremities and the tail close to the torso to allow efficient removal of the skin from the complete torso. During removal of the extremities, try to keep the cuts as small as possible in diameter.
- Fix the torso with curved fine forceps and gently move blunt tip scissors underneath the skin from cranial to caudal along the back to separate the skin from the torso. Slit the skin along the back.
- For FACS analysis, isolation of keratinocytes or preparation of RNA, peel off the complete skin from the torso using forceps to fix the tissue and the blunt tip scissors to push off the torso. For the preparation of cryosections or whole mounts take only pieces of the back skin.
- Chop the back skin with a scalpel into 1-2 pieces of approximately 10 x 10 mm. Put the pieces in one well of a 6-well plate with ~1 ml sterile PBS. Make sure that the dermal side faces the bottom.
- Replace PBS by 1.5 ml dispase II (5 mg/ml PBS) and incubate for either 30 min at 37 °C (for whole mounts) or O/N at 4 °C. Wash 3 times with PBS. Gently remove the epidermis as an intact sheet using one forceps to fix the underlying dermis and the other forceps to lift the epidermis. Use a binocular to make this step easy. Transfer the epidermal sheet into DMEM with its basal side towards the bottom. Use the sheets immediately for infection experiments. NOTE: Use the preparation of epidermis from newborn skin mainly for FACS analysis, isolation of keratinocytes or preparation of RNA. Because of its fragility, the preparation of infected sheets for cryosections or whole mounts is less suitable.
- Preparation of epidermis from adult tail skin for infection studies
- Euthanize the mice at the age of 1 to 3 months by cervical dislocation. Take tail instead of back skin since the long embedded hair follicles on the back hamper separation of intact epidermal sheets. Cut off the tail from sacrificed mice and slit it lengthwise with a scalpel.
- Peel off the skin from the bone and chop it with a scalpel into pieces of about 5 x 5 mm. Put up to 4 pieces in one well and proceed with the incubation in dispase II as described under 2.1.5 NOTE: Use adult tail skin to prepare cryosections or whole mounts. Alternatively to the immediate use, tails can be stored or transported for approximately 24 hr at 4 °C without losing significant infection efficiency. If tails are kept up to 48 hr, epidermal sheets could be hardly infected with HSV-1.
- Dissociation of epidermis for FACS analysis NOTE: Detection of surface protein expression by FACS analysis requires appropriate cell dissociation techniques that do not harm the cell surface and the protein of interest.
- Incubate newborn epidermal sheets in a 6-well plate with the basal side on 1.5 ml/well recombinant trypsin-based cell dissociation solution for 15 min at RT and for 15 min at 37 °C.
- Stop the incubation by adding 3 ml DMEM. Dissociate keratinocytes by using a curved fine forceps to gently move the epidermis in the 6-well plate so that the keratinocytes get dissolved. Collect the cell suspension and repeat this step 3 times always using fresh DMEM. NOTE: This dissociation procedure is appropriate to detect nectin-1 on the surface of keratinocytes.
- Alternatively, incubate the epidermal sheets on enzyme-free cell dissociation solution (CDS) for 15 min at RT and 15 min at 37 °C. Incubate another 15 min at RT to gently flush out the keratinocytes by pipetting CDS up and down.
- Collect the cell suspension and repeat the flushing step 3 times using fresh DMEM. NOTE: This dissociation is suitable to detect surface expression of HVEM. The disadvantage of the CDS solution is that fewer cells are dissociated than with recombinant trypsin-based cell dissociation solution. In addition to surface expression, nuclear or cytoplasmic expression such as ICP0 expression can be analyzed by FACS after dissociation of the infected epidermal sheets with recombinant trypsin-based cell dissociation solution.
- Dissociation of epidermis to isolate keratinocytes or extract RNA
- Put newborn epidermal sheets in a 6-well plate with the basal side towards the bottom of the dish which contains 1.5 ml/well recombinant trypsin-based cell dissociation solution. Incubate for 30 min at RT. Add 3 ml DMEM and dissociate keratinocytes by using a curved fine forceps to gently move the epidermis in the dish.
- Collect the cell suspension and repeat this step 3 times using fresh DMEM. Use this cell suspension to extract RNA or to culture primary murine keratinocytes.
3. Infection of Epidermal Sheets with HSV-1
Prepare a solution of HSV-1 wt strain 1711 in not less than 500 µl DMEM and replace the medium by the virus solution on which the epidermal sheets are floating. This time point is defined as time 012. Perform infection of an epidermal sheet in one well of a 24-well plate at 37 °C. The calculation of the virus titer is based on the estimated number of cells in the basal layer per epidermal sheet (approximately 2 x 105 cells per 5 x 5 mm sheet).
Infect with HSV-1 at approximately 100 plaque forming units (PFU)/cell and replace the virus-containing medium by DMEM at 1 hr p.i. NOTE: In case of epidermal sheets from newborn skin, keep in mind that sheets start to dissociate during incubation.
4. Visualization of Infected Cells
- Epidermal Whole Mounts13
- Fix epidermal sheets with 3.4% formaldehyde either for 2 hr at RT or O/N at 4 °C. Wash two times with PBS and block with 0.5% milk powder, 0.25% gelatin from cold water fish skin and 0.5% Triton X-100 in TBS for 1 hr at RT14.
- Incubate the sheets O/N with mouse anti-ICP0 (monoclonal antibody 11060)15 diluted 1:60 in blocking buffer at RT. To visualize the intermediate filament network incubate simultaneously with rabbit polyclonal anti-mouse keratin 14 (100 ng/ml).
- Wash 4 to 5 times with PBS-0.2% Tween 20 during 4 hr at RT. Incubate O/N with AF488-conjugated anti-mouse IgG (1 µg/ml), AF555-conjugated anti-rabbit IgG (1 µg/ml), and DAPI (4’, 6-diamidino-2-phenylindole) (100 ng/ml) in blocking buffer at RT.
- Wash with PBS-0.2% Tween 20 for 4 hr at RT. Mount epidermal sheets with their basal side on top of a specimen slide, embed in fluorescent mounting medium and cover with coverslips.
- Epidermal Cryosections
- Adult epidermal sheets were fixed in 4% formaldehyde in PBS for 20 min at RT and washed 2 times with PBS. Embed fixed sheets in tissue freezing medium and freeze in liquid nitrogen. Cut 8-10 µm sections with a cryomicrotome.
- Fix the sections again with 0.5% formaldehyde in PBS for 10 min at RT, wash 2 times with PBS, block with 5% normal goat serum and 0.2% Tween 20 in PBS for 30 min at RT, and then stain for 60 min with mouse anti-ICP0 (monoclonal antibody 11060)15 diluted 1:60 in blocking buffer, followed by 3 times washing with the blocking buffer.
- Then incubate with AF488-conjugated anti-mouse IgG (1 µg/ml) and DAPI (100 ng/ml) in blocking buffer for 45 min at RT and wash 3 times. Embed sections in fluorescent mounting medium and cover with coverslips.
Representative Results
The challenge of the method is to prepare epidermal sheets into which HSV-1 can penetrate from the basal layer. The critical step is the separation of the epidermis from the dermis by dispase II treatment, which, depending on the mouse strain, needs to be adapted. The concentration of dispase II can range from 2.5 to 5 mg/ml, and the time of incubation from 20 to 45 min. The staining of the intermediate filament protein keratin 14 readily allows predicting whether the basal epidermal layer can be infected or whether it will show only a very limited number of infected cells (Figure 2). Ideally, the keratin 14 staining of epidermal whole mounts should indicate an intact layer of basal cells (Figure 2A). In the worst case, dispase II treatment disrupted the basal layer and there are no cells stained in the interfollicular epidermis indicating that the keratin 14-expressing basal keratinocytes are dissociated (Figure 2B). While epidermal sheets from C57BL/6 mice were smoothly separated by treatment with 5 mg/ml dispase II for 30 min at 37 °C, this same procedure disrupts the basal layer of the epidermis from B6.A2G-Mx1 mice16 (Figure 2B). Only after reduction of dispase II to 2.5 mg/ml for 30 min, an intact basal layer was obtained as visualized by keratin 14 staining (Figure 2C).
Because of the dense hair follicles as appendages of the epidermis, the separation of the epidermis from the dermis is most convenient when skin is taken from the tails of adult mice. Alternatively, back skin of newborn mice with developing hair follicles can be used, although the fragility of the sheets after incubation limits the size of analyzable areas. So far, no difference in the efficiency of HSV-1 infection was noticed depending on whether epidermis was taken from newborn or adult mice (Figure 3). Whole mounts of newborn epidermis visualize efficient infection of the interfollicular epidermis as well as of the densely packed cells lining the developing hair follicles (Figure 3). While the interfollicular epidermis of adult epidermis is efficiently infected, hair germs and only parts of the outer root sheaths of hair follicles are infected (Figure 3). The sebaceous glands underneath the hair shaft are always stained, which is, however, due to non-specific staining of the secondary antibody as shown in uninfected controls (Figure 3).
Using inhibitor studies we investigated, whether cholesterol plays a role for HSV-1 entry into epidermis. Epidermal sheets of adult mice were pretreated with methyl-β-cyclodextrin (MβCD), which depletes cholesterol from the plasma membrane17-19. Prior to infection, MβCD was removed by several washing steps to avoid any depleting effect on cholesterol in the viral envelope. Upon pre-treatment with 20 mM MβCD almost no infected cells were seen in either the interfollicular epidermis or hair germs, but only the non-specific staining of the sebaceous glands was visible (Figure 4A). As a control that the tissue was not damaged by inhibitor treatment, we added 500 µg/ml cholesterol to replenish the MβCD-treated epidermal sheets (Figure 4A). This treatment completely restored infectivity demonstrating that the pretreatment with the inhibitor did not interfere with viability of the tissue.
Using mouse models we addressed the involvement of nectin-1 and herpesvirus entry mediator (HVEM) as cellular receptors for HSV-1 in epidermis10. Both proteins are known to act as efficient receptors for HSV in various cell lines20,21, and they are sufficient for disease development in mice22. We determined the epidermis-specific roles of nectin-1 and HVEM as HSV-1 receptors10. After floating epidermal sheets from wildtype, nectin-1- or HVEM-deficient mice on virus-containing medium, whole mounts were prepared at 3 hr p.i. to visualize the number of infected cells. The comparison of the interfollicular epidermises indicates that only nectin-1 deficiency dramatically reduced the number of ICP0-expressing cells (Figure 4B). In addition, viral spread was rather limited in the absence of nectin-1 as shown in cryosections at 18 hr p.i.(Figure 4C). For the quantification of the infected cells, it is important to monitor whether the number of infected cells is equally distributed across the interfollicular epidermis or whether clusters of infected cells are observed. In nectin-1-deficient epidermis for example, the infection efficiency varied with approximately 10% infection in some areas and approximately 30% in other areas of the interfollicular epidermis10. Thus, we decided to give no numbers of infected cells to demonstrate the decrease of infection, but present whole mounts as qualitative data.
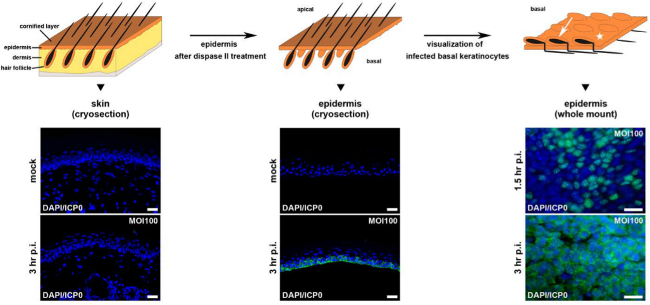 Figure 1: Visualization of infected cells in epidermal sections or whole mounts.
Please click here to view a larger version of this figure.
Figure 1: Visualization of infected cells in epidermal sections or whole mounts.
Please click here to view a larger version of this figure.
At the top is a scheme illustrating complete adult skin samples, epidermal sections after separation from dermis, and epidermal whole mounts with the visible surface of the basal keratinocyte layer. The arrow points to a hair follicle and the star indicates the interfollicular epidermis. Skin samples and epidermal sheets prepared from 3 week old mice were infected with HSV-1 at approximately 100 PFU/cell and fixed at 1.5 or 3 hr p.i. Samples were stained with anti-ICP0 (green) after mock-infection or at 1.5 or 3 hr p.i. to visualize infected cells, and counterstaining of the nuclei with DAPI (blue) demonstrates all cells in the various layers. As a control, mock-infected complete skin samples are shown in comparison to infected samples at 3 hr p.i. demonstrating no ICP0 staining (left part). Mock-infected or infected epidermal sheets prepared as cryosections are shown in the middle part. Epidermal whole mounts infected for 1.5 or 3 hr are shown on the right part. Bar, 25 µm (cryosections), and 40 µm (whole mounts).
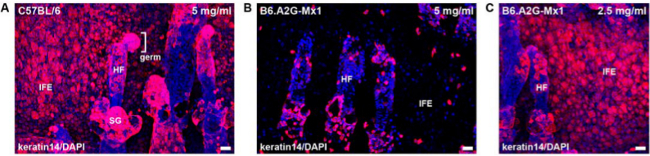 Figure 2: Keratin 14 staining of epidermal whole mounts after incubation with different concentrations of dispase II. Epidermis from C57BL/6 or B6.A2G-Mx1 mice was separated from dermis with dispase II. Epidermal whole mounts were stained with anti-keratin 14 (red) and with DAPI (blue). Epidermal sheets were incubated with 5 mg/ml (A, B) or 2.5 mg/ml (C) dispase II for 30 min at 37 °C. Bar, 25 µm. Abbreviations: hair follicle (HF), interfollicular epidermis (IFE), sebaceous gland (SG). Please click here to view a larger version of this figure.
Figure 2: Keratin 14 staining of epidermal whole mounts after incubation with different concentrations of dispase II. Epidermis from C57BL/6 or B6.A2G-Mx1 mice was separated from dermis with dispase II. Epidermal whole mounts were stained with anti-keratin 14 (red) and with DAPI (blue). Epidermal sheets were incubated with 5 mg/ml (A, B) or 2.5 mg/ml (C) dispase II for 30 min at 37 °C. Bar, 25 µm. Abbreviations: hair follicle (HF), interfollicular epidermis (IFE), sebaceous gland (SG). Please click here to view a larger version of this figure.
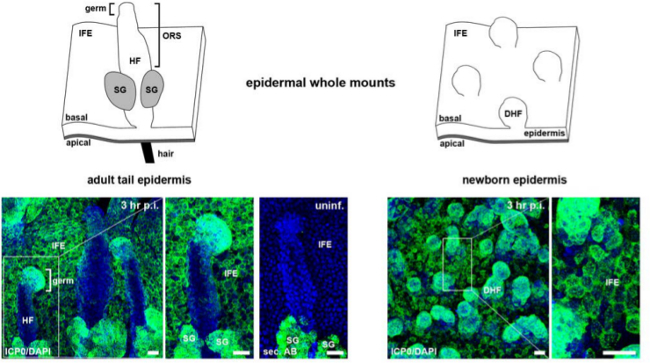 Figure 3: Visualization of ICP0 expression in whole mounts of newborn or adult epidermis. Schemes illustrating epidermal whole mounts with hair follicles or developing hair follicles, showing the visible surface of the basal layer from adult and newborn epidermises, respectively. Epidermal sheets were infected with HSV-1 at approximately 100 PFU/cell, and whole mounts were stained with anti-ICP0 (green) and with DAPI (blue) at 3 hr p.i. As a control, uninfected epidermal whole mounts from adult tail skin were stained only with secondary antibody demonstrating the non-specific staining of the sebaceous glands. Bar, 25 µm. Abbreviations: developing hair follicle (DHF), hair follicle (HF), interfollicular epidermis (IFE), sebaceous gland (SG).
Please click here to view a larger version of this figure.
Figure 3: Visualization of ICP0 expression in whole mounts of newborn or adult epidermis. Schemes illustrating epidermal whole mounts with hair follicles or developing hair follicles, showing the visible surface of the basal layer from adult and newborn epidermises, respectively. Epidermal sheets were infected with HSV-1 at approximately 100 PFU/cell, and whole mounts were stained with anti-ICP0 (green) and with DAPI (blue) at 3 hr p.i. As a control, uninfected epidermal whole mounts from adult tail skin were stained only with secondary antibody demonstrating the non-specific staining of the sebaceous glands. Bar, 25 µm. Abbreviations: developing hair follicle (DHF), hair follicle (HF), interfollicular epidermis (IFE), sebaceous gland (SG).
Please click here to view a larger version of this figure.
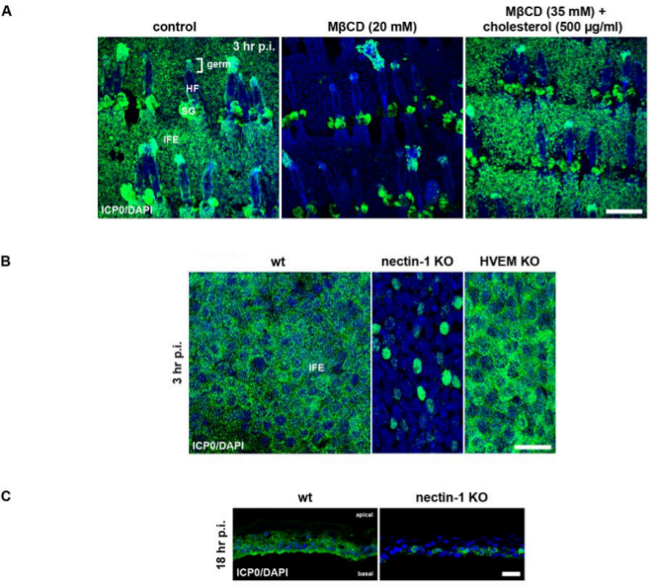 Figure 4: Assessing the impact of cellular factors by using either the inhibitor MβCD or nectin-1- or HVEM-deficient epidermises. (A) Epidermal sheets from adult tail skin of 3 week old wt mice (C57BL/6) were pretreated with MβCD (20 mM) and infected with HSV-1 at approximately 100 PFU/cell. As a control, untreated epidermal sheets were infected, and epidermal sheets pretreated with 35 mM MβCD were treated with 500 µg/ml cholesterol prior to infection. Epidermal whole mounts were stained with anti-ICP0 (green) and with DAPI (blue) at 3 hr p.i. to demonstrate the number of infected cells after depletion of cholesterol by MβCD treatment. As shown in Figure 3, staining of sebaceous glands (SG) is non-specific. Abbreviations: HF (hair follicle), IFE (interfollicular epidermis), SG (sebaceous gland). Bar, 150 µm. (B) Adult tail epidermises of wt (C57BL/6), nectin-1-23, or HVEM-deficient mice24 (1 month-old) were infected at approximately 100 PFU/cell. Epidermal whole mounts were stained with anti-ICP0 (green) and with DAPI (blue) at 3 hr p.i. (C) Cryosections of epidermal sheets from wt (C57BL/6) or nectin-1-deficient mice23 were stained at 18 hr p.i. Results shown in (B) and (C) are modified from reference 10. In A, B and C confocal projections and merged images are shown. Bar, 150 µm (A), and 25 µm (B, C). Please click here to view a larger version of this figure.
Figure 4: Assessing the impact of cellular factors by using either the inhibitor MβCD or nectin-1- or HVEM-deficient epidermises. (A) Epidermal sheets from adult tail skin of 3 week old wt mice (C57BL/6) were pretreated with MβCD (20 mM) and infected with HSV-1 at approximately 100 PFU/cell. As a control, untreated epidermal sheets were infected, and epidermal sheets pretreated with 35 mM MβCD were treated with 500 µg/ml cholesterol prior to infection. Epidermal whole mounts were stained with anti-ICP0 (green) and with DAPI (blue) at 3 hr p.i. to demonstrate the number of infected cells after depletion of cholesterol by MβCD treatment. As shown in Figure 3, staining of sebaceous glands (SG) is non-specific. Abbreviations: HF (hair follicle), IFE (interfollicular epidermis), SG (sebaceous gland). Bar, 150 µm. (B) Adult tail epidermises of wt (C57BL/6), nectin-1-23, or HVEM-deficient mice24 (1 month-old) were infected at approximately 100 PFU/cell. Epidermal whole mounts were stained with anti-ICP0 (green) and with DAPI (blue) at 3 hr p.i. (C) Cryosections of epidermal sheets from wt (C57BL/6) or nectin-1-deficient mice23 were stained at 18 hr p.i. Results shown in (B) and (C) are modified from reference 10. In A, B and C confocal projections and merged images are shown. Bar, 150 µm (A), and 25 µm (B, C). Please click here to view a larger version of this figure.
Discussion
When epidermal sheets of adult skin from C57BL/6 are infected with HSV-1 at approximately 100 PFU/cell, we observe infection in nearly all cells of the basal layer in the interfollicular epidermis while lower virus doses correlate with less infected cells and a slower progress of infection. In general, hair follicles show a variable number of infected cells; while most of the rather small keratinocytes lining the developing hair follicles are infected, only the hair germ of the adult hair follicle is completely infected. Depending on how gentle the separation of the epidermis by dispase II treatment proceeds, parts of the hair follicles get lost. In most cases, the keratinocyte layers between the sebaceous glands and the interfollicular epidermis are missing and the layer lining the outer root sheaths may be disrupted. In any case, the procedure of separating the epidermis from the dermis and infection efficiency can vary from one mouse strain to another (Figure 2).
Taken together, the challenge of the ex vivo infection assay is the preparation and maintenance of the epidermal sheets. The fragility of the tissue requires very careful handling and controls how well the tissue is preserved after separation from the dermis. A further issue is the limited viability of the sheets in culture. Since cell viability decreases over time, infection experiments should be performed immediately or at least 3 hr after preparation of the sheets. In addition to the murine skin, epidermal sheets can be also prepared from human skin or mucosa samples. Depending on the nature and the thickness of the epidermis, the dispase treatment has to be adapted.
To follow the course of infection, it is most suitable to visualize infected cells in cryosections. One has to keep in mind that with high PFU/cell and longer times of infection, the epidermal sheets start some kind of disintegration. Based on the cytopathic effect, cells round up and cell-cell contacts get disrupted. By 24 hr p.i. with 100 PFU/cell, some infected basal cells could get lost, which may also depend on the dispase II treatment. With lower PFU/cell, infection times can be extended without leading to severe tissue damage. Alternatively, the extent of damage could be used as marker of how pathogenic various virus strains are.
We prefer to visualize infected cells by staining ICP0 expression. The advantage is a quite early detection of infected cells, and, based on the ICP0 staining pattern, cells at an early stage during infection can be distinguished from those at an advanced stage. Alternatively, infected cells can be visualized by staining expression of other early genes such as the viral capsid protein VP5 and the viral envelope protein gD.
We also tried to visualize incoming virus particles prior to viral gene expression either by infection with GFP-tagged HSV-1 or by staining capsids in fixed samples. According to our experience, the pattern of GFP-fluorescence or the staining pattern in tissue sections are too complex to clearly demonstrate the presence of internalized virus particles. Alternatively, virus particles can be visualized by electron microscopy, when high virus titers of ~1,000 - 1,500 PFU/cell are used. In case of lower virus dose, virus particles can be hardly detected.
Instead of visualizing infected cells in intact epidermal sheets, the epidermis can be dissociated to analyze RNA levels or quantify the number of cells with surface, cytoplasmic or nuclear expression of genes of interest by FACS analysis.
We assume that this ex vivo infection protocol can be adapted at least to those viruses that are known to infect basal keratinocytes.
Disclosures
The authors declare that they have no competing financial interests.
Acknowledgments
We thank Peter Staeheli for providing B6.A2G-Mx1 mice and Semra Özcelik for technical advice.
This work was supported by the German Research Foundation through SFB829 and KN536/16, and the Köln Fortune Program/Faculty of Medicine, University of Cologne.
References
- Koelle DM, Corey L. Herpes simplex: insights on pathogenesis and possible vaccines. Annu. Rev. Med. 2013;59:381–395. doi: 10.1146/annurev.med.59.061606.095540. [DOI] [PubMed] [Google Scholar]
- Roizman B, Knipe DM, Whitley RJ. Herpes simplex Viruses. Fields Virology. 6th edition. 2013. pp. 1823–1897.
- Syrjänen S, Mikola H, Nykänen M, Hukkanen V. In vitro establishment of lytic and nonproductive infection by herpes simplex virus type 1 in three-dimensional keratinocyte culture. J. Virol. 1996;70:6524–6528. doi: 10.1128/jvi.70.9.6524-6528.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Visalli RJ, Courtney RJ, Meyers C. Infection and replication of herpes simplex virus type 1 in an organotypic epithelial culture system. Virology. 1997;230:236–243. doi: 10.1006/viro.1997.8484. [DOI] [PubMed] [Google Scholar]
- Hukkanen V, Mikola H, Nykänen M, Syrjänen S. Herpes simplex virus type 1 infection has two separate modes of spread in three-dimensional keratinocyte culture. J. Gen. Virol. 1999;80:2149–2155. doi: 10.1099/0022-1317-80-8-2149. [DOI] [PubMed] [Google Scholar]
- Gescher K, et al. Inhibition of viral adsorption and penetration by an aqueous extract from Rhododendron ferrugineum L. as antiviral principle against herpes simplex virus type-1. Antiviral Res. 2010;82:408–413. doi: 10.1016/j.fitote.2010.11.022. [DOI] [PubMed] [Google Scholar]
- Petermann P, Haase I, Knebel-Mörsdorf D. Impact of Rac1 and Cdc42 signaling during early herpes simplex virus type 1 infection of keratinocytes. J. Virol. 2009;83:9759–9772. doi: 10.1128/JVI.00835-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Everett RD, Maul GG. HSV-1 IE protein Vmw110 causes redistribution of PML. EMBO J. 1994;13:5062–5069. doi: 10.1002/j.1460-2075.1994.tb06835.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chrostek A, et al. Rac1 is crucial for hair follicle integrity but is not essential for maintenance of the epidermis. Mol. Cell. Biol. 2006;26:6957–6970. doi: 10.1128/MCB.00075-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petermann P, et al. Entry mechanisms of Herpes Simplex Virus Type 1 into murine epidermis: Involvement of nectin-1 and HVEM as cellular receptors. J. Virol. 2015;89:262–274. doi: 10.1128/JVI.02917-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGeoch DJ, et al. The complete DNA sequence of the long unique region in the genome of herpes simplex virus type 1. J. Gen. Virol. 1988;69:1531–1574. doi: 10.1099/0022-1317-69-7-1531. [DOI] [PubMed] [Google Scholar]
- Schelhaas M, Jansen M, Haase I, Knebel-Mörsdorf D. Herpes simplex virus type 1 exhibits a tropism for basal entry in polarized epithelial cells. J. Gen. Virol. 2003;84:2473–2484. doi: 10.1099/vir.0.19226-0. [DOI] [PubMed] [Google Scholar]
- Braun KM, Niemann C, Jensen UB, Sundberg JP, Silva-Vargas V, Watt FM. Manipulation of stem cell proliferation and lineage commitment: visualisation of label-retaining cells in wholemounts of mouse epidermis. Development. 2003;130:5241–5255. doi: 10.1242/dev.00703. [DOI] [PubMed] [Google Scholar]
- Rahn E, Petermann P, Hsu MJ, Rixon FJ, Knebel-Mörsdorf D. Entry pathways of herpes simplex virus type 1 into human keratinocytes are dynamin- and cholesterol-dependent. PLoS One. 2011;6:e25464. doi: 10.1371/journal.pone.0025464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Everett RD, Cross A, Orr A. A truncated form of herpes simplex virus type 1 immediate-early protein Vmw110 is expressed in a cell type dependent manner. Virology. 1993;197:751–756. doi: 10.1006/viro.1993.1651. [DOI] [PubMed] [Google Scholar]
- Horisberger MA, Staeheli P, Haller O. Interferon induces a unique protein in mouse cells bearing a gene for resistance to influenza virus. Proc. Natl. Acad. Sci. USA. 1983;80:1910–1914. doi: 10.1073/pnas.80.7.1910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christian AE, Haynes MP, Phillips MC. Use of cyclodextrins for manipulating cellular cholesterol content. J. Lipid Res. 1997;38:2264–2272. [PubMed] [Google Scholar]
- Ilangumaran S, Hoessli DC. Effects of cholesterol depletion by cyclodextrin on the sphingolipid microdomains of the plasma membrane. Biochem. J. 1998;335:433–440. doi: 10.1042/bj3350433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yancey PG, et al. Cellular cholesterol efflux mediated by cyclodextrins: Demonstration of kinetic pools and mechanism of efflux. J. Biol. Chem. 1996;271:16026–16034. doi: 10.1074/jbc.271.27.16026. [DOI] [PubMed] [Google Scholar]
- Montgomery RI, Warner MS, Lum BJ, Spear PG. Herpes simplex virus-1 entry into cells mediated by a novel member of the TNF/NGF receptor family. Cell. 1996;87:427–436. doi: 10.1016/s0092-8674(00)81363-x. [DOI] [PubMed] [Google Scholar]
- Geraghty RJ, Krummenacher C, Cohen GH, Eisenberg RJ, Spear PG. Entry of alphaherpesviruses mediated by poliovirus receptor-related protein 1 and poliovirus receptor. Science. 1998;280:1618–1620. doi: 10.1126/science.280.5369.1618. [DOI] [PubMed] [Google Scholar]
- Taylor JM, Lin E, Susmarski N, Yoon M, Zago A, Ware CF, Pfeffer K, Miyoshi J, Takai Y, Spear PG. Alternative entry receptors for herpes simplex virus and their roles in disease. Cell Host Microbe. 2007;2:19–28. doi: 10.1016/j.chom.2007.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barron MJ, Brookes SJ, Draper CE, Garrod D, Kirkham J, Shore RC. The cell adhesion molecule nectin-1 is critical for normal enamel formation in mice. Hum. Mol. Genet. 2008;17:3509–3520. doi: 10.1093/hmg/ddn243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Y, Subudhi SK, Anders RA, Lo J, Sun Y, Blink S, Wang Y, Liu X, Mink K, Degrandi D, Pfeffer K, Fu YX. The role of herpesvirus entry mediator as a negative regulator of T cell-mediated responses. J. Clin. Invest. 2005;115:711–717. doi: 10.1172/JCI200522982. [DOI] [PMC free article] [PubMed] [Google Scholar]