

Abstract
Development of the vertebrate craniofacial structures requires precise coordination of cell migration, proliferation, adhesion and differentiation. Patterning of the Meckel's cartilage, a first pharyngeal arch derivative, involves the migration of cranial neural crest (CNC) cells and the progressive partitioning, proliferation and organization of differentiated chondrocytes. Several studies have described CNC migration during lower jaw morphogenesis, but the details of how the chondrocytes achieve organization in the growth and extension of Meckel’s cartilage remains unclear. The sox10 restricted and chemically induced Cre recombinase-mediated recombination generates permutations of distinct fluorescent proteins (RFP, YFP and CFP), thereby creating a multi-spectral labeling of progenitor cells and their progeny, reflecting distinct clonal populations. Using confocal time-lapse photography, it is possible to observe the chondrocytes behavior during the development of the zebrafish Meckel’s cartilage.
Multispectral cell labeling enables scientists to demonstrate extension of the Meckel’s chondrocytes. During extension phase of the Meckel’s cartilage, which prefigures the mandible, chondrocytes intercalate to effect extension as they stack in an organized single-cell layered row. Failure of this organized intercalating process to mediate cell extension provides the cellular mechanistic explanation for hypoplastic mandible that we observe in mandibular malformations.
Keywords: Developmental Biology, Issue 104, Craniofacial development, cranial neural crest, confocal microscopy, fate mapping, multi-spectral clonal analysis, sox10, Zebrabow, zebrafish, lower jaw, Meckel’s cartilage
Introduction
Craniofacial development requires complex molecular, cellular and tissue interactions to drive cell proliferation, migration and differentiation1,2,3. This tightly regulated and complex process is subject to genetic and environmental perturbations, such that craniofacial deformities are amongst the most common birth malformations1-9. While surgical interventions remain the mainstay of treatment for craniofacial anomalies, understanding the development basis is essential to innovate future therapies. Therefore, studying the morphogenesis and the mechanisms in the convergence and extension and cell integration provides novel insights into the formation of the craniofacial skeleton1.
Cranial neural crest migrate and populate the first pharyngeal arch, then form paired mandibular processes that extend to form the Meckel’s cartilage, which prefigures the mandible. Morphogenesis of the Meckel’s cartilage requires chondrocyte organization via directional proliferation, cell polarization and differentiation1,10. However, the intricacy of chondrocyte organization in the growth and extension of the Meckel's cartilage remains unclear. Understanding dynamic cell behavior is critical to understanding congenital malformations affecting mandibular size, such as hypoplastic mandible phenotypes11.
Zebrafish embryos offer many developmental and genetic advantages for detailed study of Meckel’s cartilage morphogenesis. Their genetic tractability, transparency, ex vivo and rapid development are powerful advantages lending it well for observation of cell movement and organization by live imaging6. Using lineage-tracing tools, such as sox10:kaede transgenic line, we and others have delineated the neural crest origins of the embryonic craniofacial skeleton1,5. Using the sox10:ERT2-Cre with the ubi:Zebrabow-M transgenic line, it is now possible to explore details of cellular movements during craniofacial development. The Zebrabow-M, is a transgenic line engineered with the ubiquitin promoter driving the expression of different fluorophores, each flanked by Lox sites8. The Zebrabow-M default fluorophore is Red, expressing RFP. After induction of Cre expression, the Zebrabow-M construct recombines and cells express a combination of different fluorophores (RFP, CFP and YFP) creating multi-spectral expression in the embryo. All the daughter cells that divide from the labeled cells after the recombination event are then clonally labeled, so that cell populations that derive from different juxtaposed progenitors are clonally labeled. By this cloning cell labeling, cells proliferation and migration with clonal resolution can be followed (Figure 1 and 2).
Protocol
Massachusetts General Hospital Institutional Animal Care and Use Committee (IACUC) approved all procedures under protocol number #2010N000106. This is in compliance with the Association for Assessment and Accreditation of Laboratory Animal Care International (AAALAC) guidelines.
1. Reagents and Materials Preparation
Prepare 1 L of 50X E3 embryo medium (See Table 1) and prepare 1 L of 1X E3 embryo medium (See Table 2).
Prepare E3 embryo medium with N-Phenylthiourea (PTU) by adding 30 mg of PTU to 1 L of 1X E3. Stir O/N to ensure that the PTU has completely dissolved. Store at RT.
To prepare pronase solution, dilute 150 µl of pronase (0.5 mg/ml) with 15 ml of 1X E3. This amount is sufficient for 1 petri dish. Use immediately.
- Prepare Methylcellulose solution.
- Prepare E3-tricaine solution by mixing 75 ml of 0.2% tricaine with 25 ml of 1X E3. Boil 3 g of methylcellulose in the 75 ml of E3-tricaine solution.
- Make up the final volume to 100 ml with E3-tricaine solution. The final methylcellulose solution is viscous and opaque with a yellowish tint. Storage: RT protected from the light.
- Tamoxifen stock and induction solution
- Prepare 5 mM tamoxifen stock solution by dissolving 4-hydroxytamoxifen in 100% ethanol. Store stock at -80 °C.
- Prepare fresh dilution of tamoxifen at the desire concentration in 1X E3 to obtain the induction solution. CAUTION! Please consult MSDS before use.
Prepare 1 L of 0.2% tricaine as per Table 3.
Obtain confocal slides. Glue four coverglass (18 x 18) together to form a block then place 2 blocks on a 24 x 60 coverglass 2 cm apart to allow mounting of the embryo in the middle.
2. Embryo Preparation
Transgenic lines. NOTE: Zebrabow-M was a generous gift from Alex Schier’s laboratory.
- sox10: ERT2-Cre NOTE: Generate the targeting vector pDestTol2-sox10:ERT2-Cre using standard molecular cloning methods and commercially obtained recombination cloning technology.
- Combine 5’ entry clone (pENTR5’-sox10p) containing 7.2Kb of the sox10 promoter, a Gateway middle clone (pME-ERT2-Cre), and a 3’ entry clone (pENTR3’-polyA) in an LR reaction with lens destination vector pDestTol2-α-crystallin:YFP.
- Purify construct pDestTol2-sox10:ERT2-Cre. Co-inject the purified construct (100 ng/µl) with Tol2 transposase mRNA (50 ng/µl) into WT embryos (F0) at the one cell stage.
- Screen F0 adult founders for germ-line transmission based on strong yellow fluorescence in F1 embryos.
- sox10: ERT2-Cre;Zebrabow-M
- Outcross the Zebrabow-M line (2.1.1) with the sox10:ERT2-Cre transgenic line.
- Select embryos exhibiting a strong ubiquitous RFP expression and YFP lens marker and grow them till adults.
3. Embryo Collection
Set up several pairs of sox10:ERT2-Cre;Zebrabow-M transgenic zebrafish between 5 pm and 6 pm on Day 1.
The following day, pull the dividers in the morning and collect the eggs around noon.
Transfer them to petri dishes and add 15 ml of the pronase solution to dechorionate the embryos.
Incubate the embryos for 1 to 5 min until approximately 60% of the embryos have been dechorionated. Stop the reaction by decanting the pronase solution and wash the embryos 3-4 times with fresh E3 medium.
Incubate the dechorionated embryos in E3 at RT O/N and let them develop to reach the 10 somites stage. Incubate 50-60 embryos per clutch to avoid different developmental growth.
4. Treatment
Check the embryos’ developmental stage under a light field microscope to ensure they are at the 10 somites stage.
Using a fluorescent microscope with a red fluorescent protein (RFP) filter, isolate the bright red embryos.
Proceed to the induction of the Cre-recombination by adding 10 µM of hydroxytamoxifen solution to the petri dish and incubating the isolated embryos at 28.5 °C for 3 hr.
Decant the tamoxifen solution and wash the embryos several times with E3. CAUTION! discard the tamoxifen solution in a special biological waste container.
Incubate the embryos at 28.5 °C O/N.
When the embryos are approximately the 28-29 somites stage (around 24 hr post fertilization), incubate the embryos at 28.5 °C in E3-PTU solution hereafter. NOTE: PTU is here used to inhibit formation of melanophores in the developing embryos. This treatment is necessary to avoid the signal distortion of fluorescence by auto-fluorescent melanophores.
Incubate the embryos at 28.5 °C
5. Embryo Selection
To visualize the fully developed craniofacial structures, select the embryos at 4 days post fertilization for the intensity of the Red signal and Cre expression marker positivity (yellow in the retina).
Anesthetize the embryos with E3/0.015% tricaine solution. Anesthesia is confirmed when no active movements or fin activity is observed when gently prodding the embryo.
6. Mounting the Embryo in Methycellulose
Transfer the selected embryo for imaging into a petri dish containing a drop of methylcellulose and gently embalm the embryo with methylcellulose.
On a clean confocal slide, place a drop of fresh methylcellulose and mount your embryo within the drop using a microloader tip. Position embryo dorsally taking caution not to touch the embryo directly.
Cover the mounted embryo with a 25 x 25 coverslip and adjust the position of the embryo if necessary by manipulating the coverslip.
The embryo is now ready to image.
7. Analysis by Fluorescence Microscopy
Use the 20x dry lens objective (numerical aperture 0.75; pinhole size 2.0 µm) for focusing the embryo.
Press the L100 button on the microscope and select the channels in the software.
Select the confocal setup button and click ‘auto’ and select the following channels before closing the window: Ch1: CFP, Ch2: GFP, Ch3: RFP or mCherry
Turn on Ch2 and Ch3 before clicking the ‘remove interlock’ button. Click ‘play’ to begin scanning.
Focus on structure of interest, which is best done by starting with ½ frame/sec and high voltage (HV) 150 and then adjusting the settings accordingly until the desired image quality is achieved. The RFP would be brighter than other colors, therefore adjust the HV. Change the Z position to find both YFP and RFP positive cells.
Once set with the settings, capture the image of your embryo. Save your picture in the confocal imaging software format and TIFF format for image processing in each split channel.
Next, for CFP imaging, turn off Ch2 and Ch3 and turn on Ch1. Re-adjust the settings as in step 5 without changing the focus area. NOTE: CFP can be really hard to detect due to background signal in blue channel that can be circumvented by adjusting the pinhole size. A larger pinhole size permits better detection of the fluorescence by reducing signal/background noise ratio. However, a larger pinhole will reduce confocal effect, compromising image quality. Therefore, careful adjustment of the settings is required to achieve the desired image quality and intensity.
Capture the image and save it in the both format (confocal software and TIFF) for image processing. Images can then be processed using common image processing software to merge layers and improve color settings as described by Pan and colleagues 8.
Representative Results
Traditional cartilage visualization by whole mount Alcian blue stains has been invaluable in observing the developing Meckel’s cartilage and commonly used to visualize final cellular organization12 (Figure 1A). To further analyze the developing chondrocytes overtime, lineage tracing using sox10:Kaede transgenic lines has enabled us to study cell migration, convergence and extension in live embryos2,12(Figure 1B). However, the organization of chondrocytes in the craniofacial skeletal structure remains poorly understood. By live imaging, the Zebrabow-M transgenic line is able to reveal the process of chondrocyte intercalation and elongation that mediates the extension and growth of the Meckel’s cartilage (Figure 1C-G).
In this protocol, ubi:Zebrabow-M;sox10:ERT2-Cre embryos were treated with tamoxifen at 10 somites and chondrocytes in the lower jaw were visualized at different time points during development (from 3 dpf to 5 dpf). The lower jaw is best pictured ventrally to allow clear views of the Meckel's cartilage without the interference of the ethmoid plate and neurocranium dorsally. The stably maintained and inherited colors allow easy lineage tracing and CNCC cells fate mapping as indicated by the black arrows in Figure 2A-2D. The single blue cell appears to migrate and intercalate with its neighboring cells. It then elongates along the anterior-posterior axis to form a uniformly shaped chondrocyte thus maintaining the global shape of the Meckel’s cartilage as it extends anteriorly.
This technique allows the study of lower jaw growth malformations at the cellular and organ level in conditions such as Robin sequence where micrognathia is one of the prominent features described. Failure in proliferation, extension or intercalation can be effectively visualized in the growing chondrocytes over time.
| NaCl | 14.61 g |
| KCl | 0.65 g |
| CaCl2 | 1.83 g |
| MgSO4 | 1.99 g |
| Deionized H2O | 1,000 ml |
| *Storage: RT | |
| **Add 1 drop of blue methylene as an anti-fungal |
Table 1. 50X E3 embryonic medium recipe
| 50X E3 | 20 ml |
| Deionized H2O | 980 ml |
| *Storage: RT |
Table 2. 1X E3 embryonic medium recipe
| 1M Tris-Cl (pH 9.5) | 9 ml |
| tricaine | 2 g |
| Deionized H2O | Make up to 1 L |
| *Storage: RT |
Table 3. 0.2% Tricaine recipe
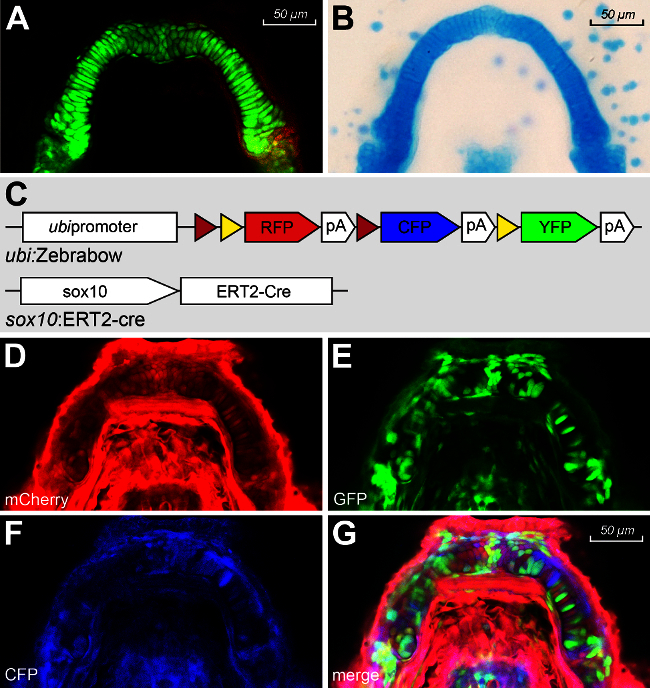 Figure 1. (A) Fluorescence Meckel’s cartilage of sox10:GFP. (B) Dissected Meckel’s cartilage of a 4 dpf Tübingen embryo stained with Alcian Blue. (C) Transgenic zebrafish lines ubi:Zebrabow-M cross with sox10:ERT2-cre. (D-G) ubi:Zebrabow-M ;sox10:ERT2-Cre image in RFP (D), YFP (E), CFP (F) channels and merge (G). Please click here to view a larger version of this figure.
Figure 1. (A) Fluorescence Meckel’s cartilage of sox10:GFP. (B) Dissected Meckel’s cartilage of a 4 dpf Tübingen embryo stained with Alcian Blue. (C) Transgenic zebrafish lines ubi:Zebrabow-M cross with sox10:ERT2-cre. (D-G) ubi:Zebrabow-M ;sox10:ERT2-Cre image in RFP (D), YFP (E), CFP (F) channels and merge (G). Please click here to view a larger version of this figure.
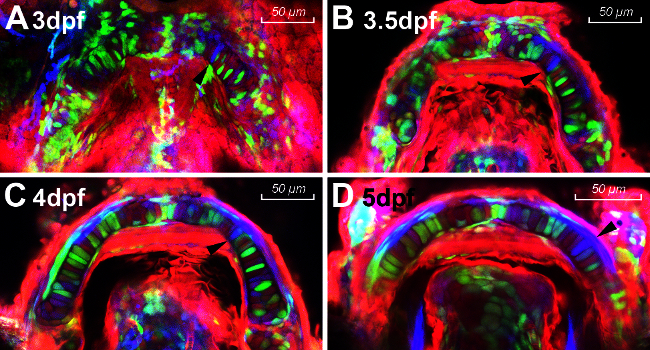 Figure 2. (A-D) Timelapse photography of Zebrabow-M :sox10 :ERT2-Cre at 3dpf (A), 3.5dpf (B), 4dpf (C) and 5dpf (D). Black arrows point to the same single clone. Please click here to view a larger version of this figure.
Figure 2. (A-D) Timelapse photography of Zebrabow-M :sox10 :ERT2-Cre at 3dpf (A), 3.5dpf (B), 4dpf (C) and 5dpf (D). Black arrows point to the same single clone. Please click here to view a larger version of this figure.
Discussion
Alcian blue and photoconvertible transgenic lines as described above complements each other to define the intricate process of cartilage and bone development. However, live cellular migration and organization during organogenesis has long been hypothesized and indirectly demonstrated but never visualized. Zebrabow-M transgenic line coupled with a cartilage specific Cre permits simultaneous live observation of all these distinct events involved in bone and cartilage formation. This technique allows the study of lower jaw growth defects at the cellular and organ level in conditions such as Robin sequence where micrognathia is one of the prominent features described. Failure in proliferation, extension or intercalation can be effectively visualized in the growing chondrocytes over time.
Color diversity optimization
Optimizing Cre levels by titrating the concentration of 4-hydroxytamoxifen can control the extent of recombination and color diversity without resulting in embryo lethality. Estrogen is critical for normal retinal development and therefore, higher doses of tamoxifen (>25µM) can induce retina degeneration, high mortality rates (~60 to 80%), craniofacial malformations and developmental delays. In addition, 4-hydroxytamoxifen is diluted in ethanol, which in itself is toxic during embryonic development13. Therefore, it is important to determine the best concentration and duration of exposition to obtain a good recombination rate but limit the toxicity for embryos.
The desired concentration of 4-hydroxytamoxifen has been optimized in this protocol. Different concentrations (5-25 µM), induction time points (10 somites, 24, 36 & 48 hpf) and duration (30 min to 6 hr) were tested (data not shown). A low dose of tamoxifen (5 µM) produced a limited recombination while higher doses (from 10 µM to 20 µM) produced similar recombination and color diversity. Doses greater than 20 µM compromised normal development of embryos and therefore not suitable for studying craniofacial malformations. Induction durations less than 2 hr were not sufficient to create adequate recombination while over 6 hr of induction induced toxicity and developmental anomalies in the embryos. Induction for 3 or 4 hr did not differ in the amount of recombination and final results.
Induction after the 24 hpf embryonic stage did not produced desired recombination and fluorescent expression. Therefore, the best compromise between normal development and suitable amount of colors without resulting in significantly high numbers of embryonic mortality was determined to be 3 hr of induction at 10 somites with a 10µM concentration of tamoxifen.
Image quality
Obtaining consistent and high quality images can be challenging. If a single field of view is unable to visualize the whole Meckel’s cartilage or craniofacial structure of interest, the whole structure can be mapped by taking Z-stacks. However, obtaining Z-stacks can be challenging in live embryo imaging due to movement artifacts. Therefore it is important to have a compromise between the quality of the Z-stack image, the amount of information required during the imaging session and prevention of laser-induced photo-bleaching during longer term laser exposure.
Another limitation of this technique is that fluorescent proteins do not show the same stability during imaging. For example RFP is a relatively stable protein but YFP can be unstable. Therefore, it is important to determine the best imaging settings empirically to detect the maximum number of colors in a given embryo with suitable intensity.
In addition to color stability, the confocal microscope is another limitation because it is unable to read the CFP and YFP channels together. This is due to the fact that the emission from CFP is detected in both 470nm and 535nm channels. In addition, the emission spectrum of CFP (470-535nm) and the excitation spectrum of YFP (530nm) overlap, creating a FRET (fluorescence resonance energy transfer) artifact. In other words, when the CFP fluorescence is detected, it excites YFP fluorescence thus creating a non-specific signal. For this reason, this protocol outlines scanning all fluorescence channels were separately and merging the individual images separately using a common imaging processing software. To circumvent the limitations of the confocal microscope, a two-photons microscope can be used as described in Pan et al’s paper where all three fluorescence expressions can be read simultaneously allowing for more color diversity8. Pan et al has described a large spectrum of color diversity in their Zebrabow-M lines (up to 30 colors). Following the conditions described in this protocol, different intensities of fluorescence and recombinations can be obtained as shown in Figure 2, although some color permutations could have been lost during the post-imaging processing.
Color inheritance and clonal analyses
Pan et al has described the stably maintained color inheritance in Zebrabow-M from progenitor to progeny that allows for fate mapping8. This protocol focused on neural CNCC cells to characterize the formation of the Meckel’s cartilage. The use of CreERT2 in zebrafish allows for efficient tamoxifen induced loxP cassette recombination and the ability to control the onset of CNCC lineage labeling during craniofacial development. This method can easily be adapted to visualize all neural crest derivatives besides the Meckel’s cartilage. In addition, using a different transgenic reporter line for clonal analysis in various different organ systems will highlight different modes of cell growth during organogenesis.
Disclosures
The authors declare that they have no competing financial interests.
Acknowledgments
The authors thank Alex Schier for kindly sharing the Zebrabow-M transgenic line, Geoffrey Burns for the pDEST vector and Renee Ethier-Daigle for excellent care of the fish facility and lines.
FUNDING:
We are grateful for generous funding support from NIDCR RO3DE024490 and Shriners Hospitals for Children (E.C.L.) and post-doctoral training fellowships from Shriners Hospitals for Children (L.R. and Y.K).
References
- Dougherty M, et al. Distinct requirements for wnt9a and irf6 in extension and integration mechanisms during zebrafish palate morphogenesis. Development. 2013;140:76–81. doi: 10.1242/dev.080473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dougherty M, et al. Embryonic fate map of first pharyngeal arch structures in the sox10: kaede zebrafish transgenic model. The Journal of craniofacial surgery. 2012;23:1333–1337. doi: 10.1097/SCS.0b013e318260f20b. [DOI] [PubMed] [Google Scholar]
- Eberhart JK, Swartz ME, Crump JG, Kimmel CB. Early Hedgehog signaling from neural to oral epithelium organizes anterior craniofacial development. Development. 2006;133:1069–1077. doi: 10.1242/dev.02281. [DOI] [PubMed] [Google Scholar]
- Dixon MJ, Marazita ML, Beaty TH, Murray JC. Cleft lip and palate: understanding genetic and environmental influences. Nature reviews Genetics. 2011;12:167–178. doi: 10.1038/nrg2933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gfrerer L, Dougherty M, Liao EC. Visualization of craniofacial development in the sox10: kaede transgenic zebrafish line using time-lapse confocal microscopy. Journal of visualized experiments : JoVE. 2013. p. e50525. [DOI] [PMC free article] [PubMed]
- McCollum CW, Ducharme NA, Bondesson M, Gustafsson JA. Developmental toxicity screening in zebrafish. Birth defects research. Part C, Embryo today : reviews. 2011;93:67–114. doi: 10.1002/bdrc.20210. [DOI] [PubMed] [Google Scholar]
- Mossey P. Dental education and CPD: are you being served. British dental journal. 2002;Suppl:3–4. [PubMed] [Google Scholar]
- Pan YA, et al. Zebrabow: multispectral cell labeling for cell tracing and lineage analysis in zebrafish. Development. 2013;140:2835–2846. doi: 10.1242/dev.094631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vieira AR. Journal of dental research. 2008;87:119–125. doi: 10.1177/154405910808700202. [DOI] [PubMed] [Google Scholar]
- Le Pabic P, Ng C, Schilling TF. Fat-Dachsous Signaling Coordinates Cartilage Differentiation and Polarity during Craniofacial Development. PLoS genetics. 2014;10:e1004726. doi: 10.1371/journal.pgen.1004726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ricks JE, Ryder VM, Bridgewater LC, Schaalje B, Seegmiller RE. Altered mandibular development precedes the time of palate closure in mice homozygous for disproportionate micromelia: an oral clefting model supporting the Pierre-Robin sequence. Teratology. 2002;65:116–120. doi: 10.1002/tera.10022. [DOI] [PubMed] [Google Scholar]
- Javidan Y, Schilling TF. Development of cartilage and bone. Methods in cell biology. 2004;76:415–436. doi: 10.1016/s0091-679x(04)76018-5. [DOI] [PubMed] [Google Scholar]
- McCarthy N, et al. Pdgfra protects against ethanol-induced craniofacial defects in a zebrafish model of FASD. Development. 2013;140:3254–3265. doi: 10.1242/dev.094938. [DOI] [PMC free article] [PubMed] [Google Scholar]