

Abstract
Recently, RASAL2 has been found to function as a tumor and metastasis suppressor. RASAL2 inhibited tumor growth, progression, and metastasis in several cancers. However, the regulatory effect of RASAL2 in NPC still remains unclear. The purpose of this paper was to identify the role of RASAL2 in the metastasis of NPC. The expression of RASAL2 in human NPC was obviously down-regulated by real-time PCR analysis. Furthermore, knock-down of RASAL2 with siRNA treatment resulted in a promotion of cell proliferation and migration in vitro. Additionally, transwell analysis results revealed that the number of invasion cells was increased of cells treated with RASAL2 siRNA. We further explored the mechanism and demonstrated that the down-regulated RASAL2 promoted migration and invasion via EMT induction with E-cadherin decreased expression and vimentin, N-cadherin, and Snail increased expression in NPC cells.In conclusion, RASAL2 inhibited the proliferation and metastasis capability of NPC cells.
Keywords: RASAL2, tumor suppressor, proliferation, metastasis, EMT, NPC
Introduction
Nasopharyngeal carcinoma (NPC) is a head and neck cancer and a common cancer in southern China and Southeast Asia [1-3]. Despite recent achievements in radio therapy and chemotherapy, there has not been any improvement in the local recurrence and distant metastases. NPC is characterized by lack of typical symptoms in early stage and often invades adjacent regions and metastasizes to regional lymph nodes and distant organs [4]. Therefore, it is important to identify new potential diagnostic markers for NPC.
Recently, it has been reported that the RasGAP gene, RASAL2, functions as a tumor and metastasis suppressor. RASAL2 is mutated or suppressed in human breast cancer, and RASAL2 ablation promotes tumor growth, progression, and metastasis in mouse models [5]. Epithelial-mesenchymal transition (EMT) is a recently identified novel concept implicated in tumor metastasis. These invasive cancer cells acquire mesenchymal, fibroblast-like morphology and show reduced intercellular adhesion and increased motility, and the processes collectively termed epithelial-to-mesenchymal transitions (EMT) [6-9]. Moreover, Studies have confirmed that several molecules/signaling associated with NPC EMT occurrence [10-13]. Weeks A et al. identified RASAL2 as an ECT2-interacting protein that regulated RHO activity and RASAL2 knockdown leaded to a conversion to an amoeboid phenotype in astrocytoma cells [14]. Huang Y et al. found that RASAL2 which knockdown activated the Ras-ERK pathway as an EMT regulator and tumor suppressor in ovarian cancer, and down-regulation of RASAL2 promoted ovarian cancer progression [15]. Li N et al. demonstrated that inactivation of RASAL2 promoted lung cancer cell migration through the induction of epithelial mesenchymal transition (EMT) and promoted lung metastasis in nude mice [16]. Here, we described that loss of RASAL2 promotes cancer progression and metastasis in the nasopharyngeal carcinoma. Hence it could serve as a potential target for the development of NPC therapies.
Materials and methods
Patient samples
Study data were obtained from 30 patients at The Northern Jiangsu People’s Hospital (Yangzhou, Jiangsu, China). The study was approved by our Institutional Ethics Committee. All research was performed in compliance with government policies and the Helsinki Declaration. Experiments were undertaken with the understanding and written consent of each subject.
Cell culture and transfection
The nasopharyngeal carcinoma cell lines were obtained from Cell bank of Sun Yat-sen University (Guangzhou, China) and maintained in DMEM medium (Life Technologies, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (Life Technologies, Carlsbad, CA, USA), 100 units/ml penicillin, and 100 μg/ml streptomycin (Invitrogen, Carlsbad, CA, USA) in a humidified 5% CO2 incubator at 37°C. Short interfering RNA specifically targeting RASAL2 (sc-62924, Santa Cruz Biotechnology, Santa Cruz, USA) and the corresponding scrambled siRNA control (Santa Cruz Biotechnology, Santa Cruz, USA) were transfected into cells in 6-well plates using Lipofectamine 2000 reagent (Invitrogen, Carlsbad, USA) according to the manufacturer’s instructions.
Real-time PCR
Real-time PCR was performed to determine the expression levels of RASAL2 and GAPDH mRNAs. Total RNA was obtained from tissues using TRIzol reagent as described by the manufacturer (Invitrogen Life Technologies Co, CA, USA). RNAs were reverse transcribed using the reverse transcription kit (Takara, Tokyo, Japan) according to the manufacture. GAPDH was used as an internal control. Primer sequences were as follows: RASAL2 5’-AAAAGAGTCACGTTCCCATGAAT-3’, reverse primer: 5’-CCAAGGATGCTACTATGAAGTGG-3’; GAPDH, forward primer: 5’-CTGGGCTACACTGAGCACC-3’, and reverse primer: 5’-AAGTGGTCGTTGAGGGCAATG-3’. Real-time PCR was performed using ABI StepOne plus (Applied Biosystems, CA, USA) with Quantifast SYBR Green PCR Kit (TAKARA, Tokyo, Japan) at 95°C for 5 min, followed by 40 cycles of 95°C for 10 s and 60°C for 30 s. Data analyses for the gene expression were performed using the 2-ΔΔCt method.
Western blot
Proteins were extracted from tissues or cultured cells using RIPA lysis buffer supplemented with protease inhibitor cocktail (Roche, Mannheim, Germany). Equal amount of protein loading in each lane was confirmed using GAPDH antibody. The extracted proteins were separated in a 10% SDS-PAGE and transferred to a PVDF membrane. The membranes were first blocked with 5% (w/v) nonfat dry milk in TBST and then with the indicated primary antibodies at 4°C overnight. Antibodies against RASAL2, E-cadherin, and N-cadherin were purchased from Cell Signaling Technology (CST, Beverly, MA, USA). Antibodies against Vimentin, snail, and GAPDH were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). After washing the membranes three times, the membranes were incubated with the appropriate secondary antibodies for 1 hour. The signals were revealed using Pierce ECL kit (Thermo Scientific Pierce, Rockford, USA). The integrated density of the band was quantified by Image J software.
Cell proliferation assay
Cell proliferation was measured by using the Cell Counting Kit-8 assay (CCK-8, Dojindo, Japan). Briefly, cells were plated into 96-well plates at a density of 104 cells/well with 100 μL of culture medium. After adhesion for 24 hours, the cells were transfected with siRNA or scramble and incubated for 24, 48 and 72 hours. At the end of each culture period, 10 μL CCK8 reagent was added to each well and incubated for another 4 hours, then the absorbance was measured at 450 nm wave length.
Wound healing assay
Cells were plated in 6-well plate and allowed to grow to confluence. Medium was removed and wounds were introduced by scraping the confluent cell cultures with a 200 μL pipette tip. Floating cells were carefully removed before complete medium was added. The cells were incubated at 37°C. The wound healing process was monitored under an inverted light microscope (Nikon, Tokyo, Japan).
Transwell invasion assay
Cells with serum-free culture medium were seeded into each well of the upper transwell chamber with Bio Coat Matrigel (BD Biosciences, San Jose, CA). In the lower chamber, 0.7 ml DMEM with 10% FBS was added. After incubating for 24 hours, the cells were stained with crystal violet. The number of cells was counted under a microscope in ten random visual fields acquired using NIS Elements image analysis software (Nikon, Tokyo, Japan).
Statistical analysis
χ2 tests was used to analyze the relationship of expression level of RASAL2 and clinical characteristics in tissues. Data were expressed as mean ± SEM. Differences between two independent groups were tested with the student’s test or one-way analysis of variance for three or more conditions. All statistical analyses were carried out using SPSS version 13.0 and presented with Graph pad prism 5.0 software. In all cases, P<0.05 was considered significant.
Results
Expression of RASAL2 was down-regulated in the nasopharyngeal carcinoma
To explore the expression of RASAL2 the nasopharyngeal carcinoma, we performed the real-time PCR assay in the 8 normal tissues and 22 nasopharyngeal carcinoma tissues. The results indicated that the levels of RASAL2 in human nasopharyngeal carcinoma tissues were obviously down-regulated compared with the normal tissues (Figure 1A).
Figure 1.
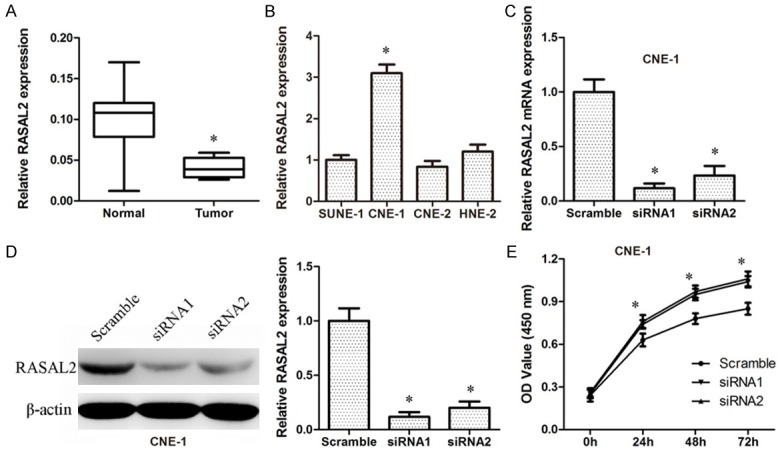
RASAL2 was down-regulated and inhibited proliferation in nasopharyngeal carcinoma. A. The relatively decreased level of RASAL2 was detected via real-time PCR in 22 nasopharyngeal carcinoma tissues compared with 8 normal tissues. B. The mRNA expression level of RASAL2 in CNE-1 cells was highest compared with SUNE-1, CNE-2, and HNE-2 cells. C. The mRNA expression level of RASAL2 in CNE-1 cells with siRNA treated by real-time PCR assay. The mRNA expression level of GAPDH was served as control. D. Western blotting assays showed decreased expression of RASAL2 in CNE-1 cells with siRNA treatment compared with scramble sequence treatment. The quantification of experiments was shown in right panel. E. The CCK-8 assay showed that a decreased level of RASAL2 promoted the growth of CNE-1 cells. Absorbance at 450 nm was presented as the mean ± SEM. All experiments were performed in triplicate and presented as the mean ± SEM. *, indicates a significant difference compared with the control group (P<0.05).
Down-regulated RASAL2 promoted proliferation in nasopharyngeal carcinoma cells
To detect whether RASAL2 affected cell phenotypes, we first detected RASAL2 expression in multiple nasopharyngeal carcinoma cell lines. Finally, we selected CNE-1 since it revealed RASAL2 high expression (Figure 1B). Both mRNA level and protein level of RASAL2 expression was knocked down by siRNA sequences (Figure 1C, 1D). We used the CCK-8 assay to determine the role of RASAL2 in cell growth. Knock-down of RASAL2 enhanced proliferation of CNE-1 cells after 24 h compared with scramble sequence transfected cells (Figure 1E). Based on the results above, we proposed that RASAL2 inhibited cell proliferation in nasopharyngeal carcinoma cells.
Down-regulated RASAL2 promoted migration and invasion via EMT induction in nasopharyngeal carcinoma cells
The clinicopathologic relevance analysis revealed that RASAL2 might play a part in nasopharyngeal carcinoma metastasis. Then, we performed the wound healing assay to discover the role of RASAL2 in nasopharyngeal carcinoma migration. The relative rate of cell migration was increased in CNE-1 treated with RASAL2 siRNA (Figure 2A). To detect the effects of RASAL2 in cell invasion ability, we conducted the transwell assay. The numbers of invasion cells was increased in CNE-1 treated with RASAL2 siRNA compare with scramble sequence transfected cells (Figure 2B). Western blot revealed that the E-cadherin expression was decreased and the expression of vimentin, N-cadherin, and snail was increased in CNE-1 treated with RASAL2 siRNA compare with scramble sequence transfected cells (Figure 2C). These results indicated that down-regulated RASAL2 promoted migration and invasion via EMT induction in nasopharyngeal carcinoma cells.
Figure 2.
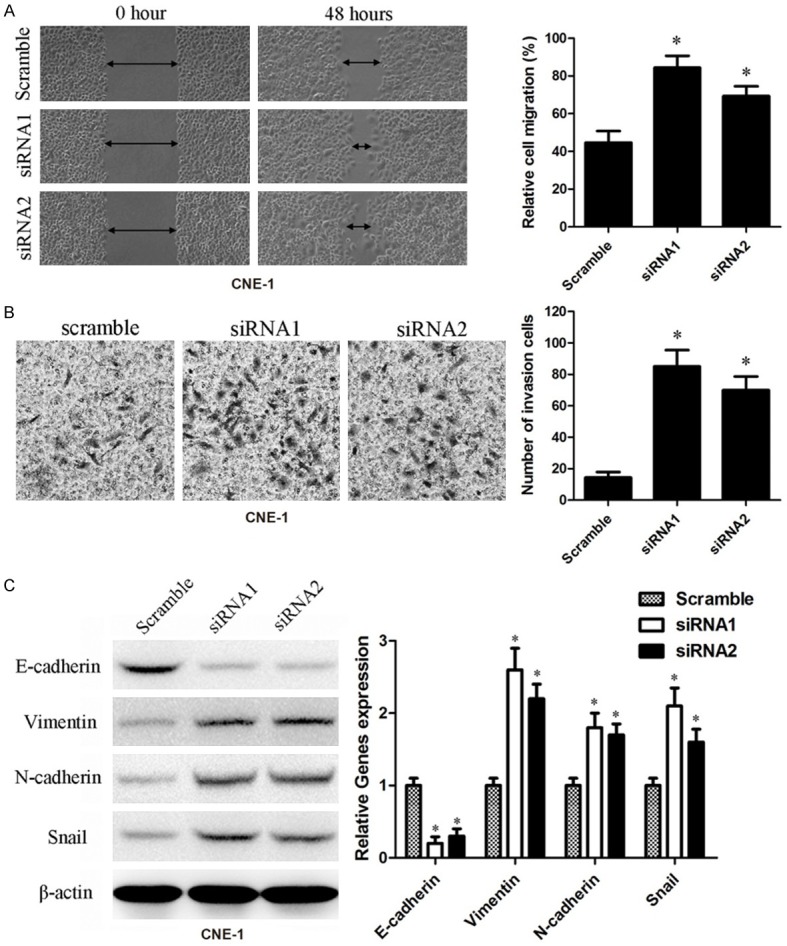
Down-regulated RASAL2 promoted migration and invasion via EMT induction in nasopharyngeal carcinoma. A. RASAL2 reduced the migration capability in CNE-1 cells with siRNA treatment compared with scramble sequence treatment by wound healing assay. The rate of cell migration was shown in the right panel. Data are presented as the mean ± SEM based on at least three independent experiments. *, indicated P<0.05. B. RASAL2 decreased the cell invasion by transwell invasion assay. Cell morphology graph of invasive cells in CNE-1 cells after transfection of siRNA or scramble. The number of cells invasion was shown in the right panel and data are presented as the mean ± SEM based on at least three independent experiments. *, indicated P<0.05. C. Western blot assay suggested that down-regulated RASAL2 decreased levels of E-cadherin and increased levels of vimentin, N-cadherin, and Snail in CNE-1 cells. The quantification of experiments was shown in right panel and data are presented as the mean ± SEM based on at least three independent experiments. *, indicated P<0.05.
Discussion
The Ras pathway is one of the most commonly deregulated pathways in human cancer and is also frequently activated as a consequence of alterations in upstream regulators and downstream effectors. Ras is regulated by Ras GTPase-activating proteins (RasGAPs), which catalyze the hydrolysis of Ras-GTP to Ras-GDP [17]. Jiang et al. demonstrated that synergism of BARF1 with Ras induces malignant transformation in human nasopharyngeal epithelial cells [18]. Zhang et al. revealed that inactivation of RASSF2A by promoter methylation correlates with lymph node metastasis in nasopharyngeal carcinoma [19]. Mo et al. found that promoter hypermethylation of Ras-related GTPase gene RRAD inactivates a tumor suppressor function in nasopharyngeal carcinoma. Ectopic RRAD expression in NPC cell lines inhibited the cell growth, colony formation, and cell migration. RRAD might act as a functional tumor suppressor in NPC development [20]. Consequently, the expression of RASAL2 decreased in NPC was confirmed in our study. Meanwhile, we found that cell proliferation capability was reduced in RASAL2 knock-down cells.
So we explored whether RASAL2 is a novel index of nasopharyngeal carcinoma metastasis. Our study provided evidences for the crucial role of RASAL2 in the pathogenesis of nasopharyngeal carcinoma migration and invasion. Horikawa et al. found that expression of Twist and LMP1 is directly correlated and expression of Twist is associated with metastasis via inducing EMT occurrence in human NPC [21]. Yang et al. demonstrated that co-expression of HIF-1alpha, Twist and Snail in primary tumors of patients with head and neck cancers correlated with metastasis and the worst prognosis. However, repression of Twist in HIF-1alpha-overexpressing or hypoxic cells reversed EMT and metastatic phenotypes [22]. Kong et al. discovered that LMP2A induces EMT and stem-like cell self-renewal in NPC, suggesting a novel mechanism by which Epstein-Barr virus induces the initiation, metastasis and recurrence of NPC [23]. Horikawa et al. revealed that overexpression of Snail positively correlated with expression of LMP1, metastasis and independently correlated inversely with expression of E-cadherin is associated with in carcinomatous cells in NPC tissues [24]. This study strengthens the association of EMT with the metastatic behavior of NPC [24]. Yadav et al. demonstrated that IL-6 promotes head and neck tumor metastasis by inducing epithelial-mesenchymal transition via the JAK-STAT3-SNAIL signaling pathway [25]. Li et al. found that IL-8 promotes metastasis of nasopharyngeal carcinoma through induction of epithelial-mesenchymal transition and activation of Akt signaling [26]. Luo et al. confirmed that EMT may play an important role in the biological progression of NPC, and nuclear vimentin and cytoplasmic E-cadherin might have independent prognostic value in NPC patients and serve as novel targets for prognostic therapeutics [27]. Zong et al. newly suggested that ZNF488 acted as an oncogene, promoting invasion and tumorigenesis by activating the Wnt/beta-catenin pathway to induce EMT in NPC [28]. To further explore the embedded mechanism, we investigated the EMT occurrence in RASAL2 knock-down cells. Taken together, the combination results confirmed that the down-regulated RASAL2 promoted migration and invasion via EMT induction in nasopharyngeal carcinoma.
In conclusion, we identified a novel tumor suppresser RASAL2 in human nasopharyngeal carcinoma which was significantly correlated with metastasis. Down-regulated expression of RASAL2 increased proliferation, migration and invasion capability via EMT induction in nasopharyngeal carcinoma cells. We proposed that RASAL2 could be applied as a potential target for the prevention of nasopharyngeal carcinoma progression.
Acknowledgements
This work was supported by grants from the Jiangsu Natural Science Foundation BK20131233.
Disclosure of conflict of interest
None.
References
- 1.Le QT, Tate D, Koong A, Gibbs IC, Chang SD, Adler JR, Pinto HA, Terris DJ, Fee WE, Goffinet DR. Improved local control with stereotactic radiosurgical boost in patients with nasopharyngeal carcinoma. Int J Radiat Oncol Biol Phys. 2003;56:1046–1054. doi: 10.1016/s0360-3016(03)00117-2. [DOI] [PubMed] [Google Scholar]
- 2.Chang ET, Adami HO. The enigmatic epidemiology of nasopharyngeal carcinoma. Cancer Epidemiol Biomarkers Prev. 2006;15:1765–1777. doi: 10.1158/1055-9965.EPI-06-0353. [DOI] [PubMed] [Google Scholar]
- 3.Ong YK, Heng DM, Chung B, Leong SS, Wee J, Fong KW, Tan T, Tan EH. Design of a prognostic index score for metastatic nasopharyngeal carcinoma. Eur J Cancer. 2003;39:1535–1541. doi: 10.1016/s0959-8049(03)00310-1. [DOI] [PubMed] [Google Scholar]
- 4.Lee AW, Poon YF, Foo W, Law SC, Cheung FK, Chan DK, Tung SY, Thaw M, Ho JH. Retrospective analysis of 5037 patients with nasopharyngeal carcinoma treated during 1976-1985: overall survival and patterns of failure. Int J Radiat Oncol Biol Phys. 1992;23:261–270. doi: 10.1016/0360-3016(92)90740-9. [DOI] [PubMed] [Google Scholar]
- 5.McLaughlin SK, Olsen SN, Dake B, De Raedt T, Lim E, Bronson RT, Beroukhim R, Polyak K, Brown M, Kuperwasser C, Cichowski K. The RasGAP gene, RASAL2, is a tumor and metastasis suppressor. Cancer Cell. 2013;24:365–378. doi: 10.1016/j.ccr.2013.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Larue L, Bellacosa A. Epithelial-mesenchymal transition in development and cancer: role of phosphatidylinositol 3’ kinase/AKT pathways. Oncogene. 2005;24:7443–7454. doi: 10.1038/sj.onc.1209091. [DOI] [PubMed] [Google Scholar]
- 7.Kalluri R. EMT: when epithelial cells decide to become mesenchymal-like cells. J Clin Invest. 2009;119:1417–1419. doi: 10.1172/JCI39675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Lopez-Novoa JM, Nieto MA. Inflammation and EMT: an alliance towards organ fibrosis and cancer progression. EMBO Mol Med. 2009;1:303–314. doi: 10.1002/emmm.200900043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Zhou C, Liu J, Tang Y, Liang X. Inflammation linking EMT and cancer stem cells. Oral Oncol. 2012;48:1068–75. doi: 10.1016/j.oraloncology.2012.06.005. [DOI] [PubMed] [Google Scholar]
- 10.Song LB, Li J, Liao WT, Feng Y, Yu CP, Hu LJ, Kong QL, Xu LH, Zhang X, Liu WL, Li MZ, Zhang L, Kang TB, Fu LW, Huang WL, Xia YF, Tsao SW, Li M, Band V, Band H, Shi QH, Zeng YX, Zeng MS. The polycomb group protein Bmi-1 represses the tumor suppressor PTEN and induces epithelial-mesenchymal transition in human nasopharyngeal epithelial cells. J Clin Invest. 2009;119:3626–3636. doi: 10.1172/JCI39374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kondo S, Wakisaka N, Muramatsu M, Zen Y, Endo K, Murono S, Sugimoto H, Yamaoka S, Pagano JS, Yoshizaki T. Epstein-Barr virus latent membrane protein 1 induces cancer stem/progenitor-like cells in nasopharyngeal epithelial cell lines. J Virol. 2011;85:11255–11264. doi: 10.1128/JVI.00188-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Aga M, Bentz GL, Raffa S, Torrisi MR, Kondo S, Wakisaka N, Yoshizaki T, Pagano JS, Shackelford J. Exosomal HIF1alpha supports invasive potential of nasopharyngeal carcinoma-associated LMP1-positive exosomes. Oncogene. 2014;33:4613–4622. doi: 10.1038/onc.2014.66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cai LM, Lyu XM, Luo WR, Cui XF, Ye YF, Yuan CC, Peng QX, Wu DH, Liu TF, Wang E, Marincola FM, Yao KT, Fang WY, Cai HB, Li X. EBV-miR-BART7-3p promotes the EMT and metastasis of nasopharyngeal carcinoma cells by suppressing the tumor suppressor PTEN. Oncogene. 2015;34:2156–2166. doi: 10.1038/onc.2014.341. [DOI] [PubMed] [Google Scholar]
- 14.Weeks A, Okolowsky N, Golbourn B, Ivanchuk S, Smith C, Rutka JT. ECT2 and RASAL2 mediate mesenchymal-amoeboid transition in human astrocytoma cells. Am J Pathol. 2012;181:662–674. doi: 10.1016/j.ajpath.2012.04.011. [DOI] [PubMed] [Google Scholar]
- 15.Huang Y, Zhao M, Xu H, Wang K, Fu Z, Jiang Y, Yao Z. RASAL2 down-regulation in ovarian cancer promotes epithelial-mesenchymal transition and metastasis. Oncotarget. 2014;5:6734–6745. doi: 10.18632/oncotarget.2244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Li N, Li S. RASAL2 promotes lung cancer metastasis through epithelial-mesenchymal transition. Biochem Biophys Res Commun. 2014;455:358–362. doi: 10.1016/j.bbrc.2014.11.020. [DOI] [PubMed] [Google Scholar]
- 17.Downward J. Targeting RAS signalling pathways in cancer therapy. Nat Rev Cancer. 2003;3:11–22. doi: 10.1038/nrc969. [DOI] [PubMed] [Google Scholar]
- 18.Jiang R, Cabras G, Sheng W, Zeng Y, Ooka T. Synergism of BARF1 with Ras induces malignant transformation in primary primate epithelial cells and human nasopharyngeal epithelial cells. Neoplasia. 2009;11:964–973. doi: 10.1593/neo.09706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zhang Z, Sun D, Van do N, Tang A, Hu L, Huang G. Inactivation of RASSF2A by promoter methylation correlates with lymph node metastasis in nasopharyngeal carcinoma. Int J Cancer. 2007;120:32–38. doi: 10.1002/ijc.22185. [DOI] [PubMed] [Google Scholar]
- 20.Mo Y, Midorikawa K, Zhang Z, Zhou X, Ma N, Huang G, Hiraku Y, Oikawa S, Murata M. Promoter hypermethylation of Ras-related GTPase gene RRAD inactivates a tumor suppressor function in nasopharyngeal carcinoma. Cancer Lett. 2012;323:147–154. doi: 10.1016/j.canlet.2012.03.042. [DOI] [PubMed] [Google Scholar]
- 21.Horikawa T, Yang J, Kondo S, Yoshizaki T, Joab I, Furukawa M, Pagano JS. Twist and epithelial-mesenchymal transition are induced by the EBV oncoprotein latent membrane protein 1 and are associated with metastatic nasopharyngeal carcinoma. Cancer Res. 2007;67:1970–1978. doi: 10.1158/0008-5472.CAN-06-3933. [DOI] [PubMed] [Google Scholar]
- 22.Yang MH, Wu MZ, Chiou SH, Chen PM, Chang SY, Liu CJ, Teng SC, Wu KJ. Direct regulation of TWIST by HIF-1alpha promotes metastasis. Nat Cell Biol. 2008;10:295–305. doi: 10.1038/ncb1691. [DOI] [PubMed] [Google Scholar]
- 23.Kong QL, Hu LJ, Cao JY, Huang YJ, Xu LH, Liang Y, Xiong D, Guan S, Guo BH, Mai HQ, Chen QY, Zhang X, Li MZ, Shao JY, Qian CN, Xia YF, Song LB, Zeng YX, Zeng MS. Epstein-Barr virus-encoded LMP2A induces an epithelial-mesenchymal transition and increases the number of side population stem-like cancer cells in nasopharyngeal carcinoma. PLoS Pathog. 2010;6:e1000940. doi: 10.1371/journal.ppat.1000940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Horikawa T, Yoshizaki T, Kondo S, Furukawa M, Kaizaki Y, Pagano JS. Epstein-Barr Virus latent membrane protein 1 induces Snail and epithelial-mesenchymal transition in metastatic nasopharyngeal carcinoma. Br J Cancer. 2011;104:1160–1167. doi: 10.1038/bjc.2011.38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Yadav A, Kumar B, Datta J, Teknos TN, Kumar P. IL-6 promotes head and neck tumor metastasis by inducing epithelial-mesenchymal transition via the JAK-STAT3-SNAIL signaling pathway. Mol Cancer Res. 2011;9:1658–1667. doi: 10.1158/1541-7786.MCR-11-0271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Li XJ, Peng LX, Shao JY, Lu WH, Zhang JX, Chen S, Chen ZY, Xiang YQ, Bao YN, Zheng FJ, Zeng MS, Kang TB, Zeng YX, Teh BT, Qian CN. As an independent unfavorable prognostic factor, IL-8 promotes metastasis of nasopharyngeal carcinoma through induction of epithelial-mesenchymal transition and activation of AKT signaling. Carcinogenesis. 2012;33:1302–1309. doi: 10.1093/carcin/bgs181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Luo W, Fang W, Li S, Yao K. Aberrant expression of nuclear vimentin and related epithelial-mesenchymal transition markers in nasopharyngeal carcinoma. Int J Cancer. 2012;131:1863–1873. doi: 10.1002/ijc.27467. [DOI] [PubMed] [Google Scholar]
- 28.Zong D, Yin L, Zhong Q, Guo WJ, Xu JH, Jiang N, Lin ZR, Li MZ, Han P, Xu L, He X, Zeng MS. ZNF 488 Enhances the Invasion and Tumorigenesis in Nasopharyngeal Carcinoma via the Wnt Signaling Pathway Involving Epithelial Mesenchymal Transition. Cancer Res Treat. 2015 doi: 10.4143/crt.2014.311. [Epub ahead of print] [DOI] [PMC free article] [PubMed] [Google Scholar]