

Abstract
AIM: To study the purifying method and characteristics of new gosling viral enteritis virus (NGVEV), the etiological agent of new gosling viral enteritis (NGVE) which was first recognized in China, as well as the pathomorphological development in goslings infected artificially with NGVEV.
METHODS: ① NGVEV virions were purified by the procedure of treatment with chloroform and ammonium sulfate precipitation, dialysis to remove the sulfate radical and ammonium ion and separation by gel filtration chromatography, and SDS-PAGE. ② Forty 2-day-old White Sichuan goslings were orally administered with NGVEV and 24 h later 2 birds were randomly selected and killed at 24 h intervals until death occurred. Specimens (duodenum, ileum, liver, heart, kidney, spleen, lung, proventriculus, pancreas, esophagus, and the intestinal embolus) were taken until all birds in this group died and were sectioned and stained with hemotoxylin and eosin and studied by light microscope.
RESULTS: NGVEV shared the typical characteristics of Adenovirus and which structural proteins consisted of 15 polypeptides. Necrosis and sloughing of the epithelial cells covering the villus tips of the duodenum were first observed in goslings 2 d postinfection artificially with NGVEV. With the progress of infection, this lesion rapidly occurred in the epithelium at the base of the villus and with infiltration of the inflammatory cells, the jejunum tended to be involved. With the intensification of mucosa necrosis and inflammatory exudation of the small intestine, fibrinonecrotic enteritis was further developed and embolus composed of either intestinal contents wrapped by pseudomembrane or of the mixture of fibrous exudate and necrotic intestinal mucosa were observed in the middle-lower part of the small intestine. This structure occluded the intestinal tract and made the intestine dilated in appearance. The intestinal glandular cells underwent degeneration, necrosis and might be found sloughed into the lumen. Hemorrhage and hyperemia could be observed on the lung and kidney. Epithelial cells of the renal tubular underwent degeneration. In some cases, granular degeneration and fatty degeneration could be found in the liver and in some cases at a later stage of this disease the epithelial cells of trachea and proventriculus might be found sloughed. In some cases at an early stage of this disease, cardiac hyperemia and hemorrhage could be observed. Esophagus, pancreas and brain were found normal. Analyses and comparisons between the pathologic lesions of NGVE and Gosling Plague (GP) were available in this paper as well.
CONCLUSION: ① NGVEV is adenovirus. ② Pathological characteristic could be as the data for NGVE diagnosis.
Keywords: enteritis/virology, enteritis/pathology, adenoviridae/isolation & purification, gosling/virology, gosling/new infectious disease
INTRODUCTION
Cheng et al[1] reported that a disease which extremely resembled Gosling Plague (GP)[2-5] in aspects of epizootiology, clinical signs and pathologic lesions was observed in goslings less than 30 d of age in a variety of areas in Sichuan province and that the sausagelike lesion found in birds which died at a later stage of the acute case was almost identical to that observed in the case of GP in terms of gross and histopathological change s. This disease with the name of NGVE was regarded as a new one caused by Adenovirus through preliminary epizootiological investigation, clinical signs, histopathological examination, causal agent isolation and the experiment of artificial infection[1,7-9]. The isolated virus of NGVEV was capable of reproducing the disease identical to natural infection in aspects of clinical signs and histopathological lesions by a variety of routes and the oral route was considered to be the best one. Goslings infected with duck plague virus (DPV) or gosling plague virus (GPV) could be observed enteritis[2,3,5,10-16], but the 36 strains of NGVEV isolated from a variety of areas were antigenically identical and no antigenic relationships with DPV and GPV was demonstrated[1,2,6-9]. The characteristics, purifying method, structural proteins of the representative causal agent of NGVEV-CN as well as the histopathologic developments observed in goslings infected with NGVEV are reported as follows:
MATERIALS AND METHODS
Virus strain
The strain of NGVEV with its minimum lethal dose being 10-6/0.5 mL to 1-day-old goslings by oral administration was isolated from the natural cases of NGVE[1].
Characteristics of NGVEV-CN
According to directions presented in referencer[17], experiments were performed to identify the following properties of NGVEV-CN: hemagglutination, buoyant density, sensitivity to temperature, pH, chloroform and trypsin, type of the nucleic acid and type of the nucleic acid strand.
Virus purification and viral structural polypeptide analysis by SDS-PAGE
Experiments were performed according to directions presented in referencer [18-21].
Virus purification The virus strain of NGVEV-CN inoculated on the primary duck fibroblasts was harvested when 75% of the cells showed cytopathic effect and were centrifugated at 4000rpm for 10 min after treated with chloroform for 5 times. The supernatant was collected and with stir was slowly added the same volume of saturated ammonium sulfate. After placed overnight at 4 °C statically, it was subjected to centrifugation at 10000 rpm for 1 h. The precipitate was dissolved in small volume of sterile distilled water and then underwent dialysis to remove sulfate radical and ammonium ion. Sephadex G200 chromatography was performed to elute the solution with phosphate buffered saline solution (PBS) of 0.15 M and pH7.2 as buffer. Nucleic acid-protein detector was employed to determine the absorbancy at 280 nm of the separately collected eluates. Cellulose acetate electrophoresis and disc polyacrylamide gel electrophoresis (PAGE) were performed to examine the purification of the isolated virus after the virus containing eluates sharing the same absorption peak were mixed. Transmission electronic microscope type H-600 was employed to examine the negatively stained virions.
Viral structural polypeptide analysis by SDS-PAGE Twenty μL of the purified virus was boiled for 5 min after mixed with thesame volume of buffer whose concentration was 2 times of that used for electrophoreses and the virus lysate underwent PAGE, with the discontinuous gradient gel containing 0.4% SDS. The gel was 1.5 mm, 11 cm, 12 cm in depth, width, length respectively and the concentration of the stacking gel and the separating gel was 3% and 12.5% respectively. A solution of Tris-Gly with a pH of 6.8 was used as buffer and electric current was raised from 160 V to 200 V when the sample moved from the stacking gel to the concentrating gel. Electrophoresis was performed at 4 °C for 4 h and six proteins were employed as molecular mass REFERENCES. These proteins and their molecular masses (Daltons) were: chicken albumen lysozyme (14400), trypsin inhibitor (20100), bovine carbonic anhydrase (31000), rabbit actin (43000), bovine serum album in (66200) and rabbit phosphorylase (97400). The gel was then removed and stained with 0.25% Coomassie brilliant blue for 12 h. After that, it was decolored with a solution containing methanol and glacial acetic acid until the proteins were visible as discrete blue bands and their background was hyaline, which took about 48 h. The gel was then photoed by camera and scanned by automatic gel image-forming and analysis system. After analyzed and processed by the software of Gel, molecular masses and relative percentages of each structural polypeptides were obtained.
The pathomorphological development observed in goslings artificially infected with NGVEV
Eighty 2-day-old White Sichuan geese whose mother birds were all vaccinated twice with attenuated GPV vaccine just before egg production were employed and each one was inoculated subcutaneously with 1 mL anti-GPV hyperimmune serum and after observed for one day these birds were divided randomly into 2 groups with each containing 40. Each of the first group was administered orally with 0.5 mL NGVEV which was diluted 1:10 and 24 h later 2 birds were randomly selected and killed at 24 h intervals until death occurred in this group. Specimens were taken until all birds in this group died. Each of the second group was administered orally with 0.5 mL sterile physiological saline solution and was kept as control. One bird was killed each day and specimens were taken until all the birds in the first group died. The specimens included the duodenum, ileum, liver, heart, kidney, spleen, lung, proventriculus, pancreas, esophagus, and the intestinal embolus. These specimens were sectioned and stained with hemotoxylin and eosin and studied by light microscope. Special staining method of Maun methylene blue and eosin was also employed for detecting inclusion bodys.
RESULTS AND DISCUSSION
Characteristics of NGVEV-CN
Hemagglutination Under the temperature of 4 °C, 25 °C, 30 °C, 37 °C, 42 °C, or with the pH of 6.6, 6.8, 7.0, 7.2, 7.4, or with either physiological saline solution or phosphate buffered saline employed, the isolated virus of NGVEV did not agglutinate the newly prepared erythrocytes of chicken, duck, goose, pigeon, yellow cattle, buffalo or pig. This indicated that NGVEV did not have the property of hemagglutination and this agreed with the conclusion that “the majority of fowl adenovirus serotypes (group 1) do not hemagglutinate”[17].
Buoyant density The buoyant density of NGVEV in cesium chloride was 1.32 g/mL.
Sensitivity to temperature The ability of NGVEV to cause primary duck embryo fibroblasts CPE and to cause goslings mortality was unaffected by storage for 36 mo at -15 °C, for 20 mo at 0 °C, or for 45 d at 37 °C. Ability to cause primary duck embryo fibroblasts CPE and infectivity to goslings were not destroyed by heating at 45 °C for 48 h, at 56 °C for 5 h, or at 60 °C for 1 h. Heating for 5 min at 80 °C or for 10 s at 96 °C (boiled water) could make the virus lose infectivity to goslings and make the virus lose the ability to cause primary duck embryo fibroblasts CPE, which was revealed by the phenomenon that no CPE was observed when even seven blind passages were performed. All these indicated that the virus of NGVEV was very resistant to heating.
Sensitivity to pH The pathogenicity and infectivity of NGVEV was stable at pH ranging from 3.0 to 8.0 and the titre of NGVEV dropped to certain extent at pH2.0 or 9.0, which was indicated by the fact that more time was needed for CPE to appear. NGVEV was inactivated at pH3 or at pH10. All these suggested that the isolated virus of NGVEV was adaptable to a comparatively wide range of pH.
Sensitivity to Chloroform Typical CPE could be observed in primary duck embryo fibroblasts inoculated with NGVEV that had been treated with chloroform for either 1 or 2 or 3 times and no differences was observed between the experiment group and the control group. This revealed that NGVEV was not sensitive to chloroform and this corresponded with the fact observed by electronic microscopy that the virions had no envelope.
Sensitivity to trypsin Whether NGVEV was treated with trypsin or not, the same CPE occurred. This suggested that NGVEV was insensitive to chloroform and was able to resist the gastroenteric proteases and gastric acid, therefore could penetrate the intestine easily[1]. NGVEV could result in the intestinal exudative or necrotic inflammation and could cause necrosis and sloughing of the intestinal epithelial cells.
Type of the nucleic acid
Drug inhibition It is well known that 5-iodedeoxyuridine (5-IudR) is similar to thymidine (T) in structure and can supersede T in DNA replication. Therefore DNA with no normal function is synthesized and virus propagation is prevented therefore make DNA virus that is capable of resulting in CPE lose this ability. Five dilutions of NGVEV, 100, 10¯¹, 10-2, 10-3, 10-4, were prepared and 40 bottles of primary duck embryo fibroblasts were randomly divided into 2 groups with each containing 20. Each dilution of NGVEV was inoculated to 4 bottles of primary duck embryo fibroblasts of each group and cultivated at 37 °C for 1 h for attachment of NGVEV to the cells before the virus containing media was removed. Then maintenance media containing 50 μg/mL 5-IudR was added to the experiment group and that containing no 5-IudR was added to the control group and observation was maintained for 14 d. It suggested that NGVEV was a DNA virus by the experimental result that no CPE was observed in the experiment group.
Enzymatic digestion The purified virus containing suspension was adjusted to a final concentration of 0.5% by adding 10% SDS and then underwent extraction by saturated phenol which was dissolved in the solvent of Tris-HCl. The liquid phase was extracted by ether for 4 times and after the ether removed was precipitated by sodium acetate of pH5.2 and dehydrated alcohol. Then it was placed overnight at -20 °C. Then centrifugation at 16500 rpm for 30 min was employed. After dried, the precipitate was dissolved in the buffer of Tris-EDTA with a pH of 7.3 and then was divided into 3 parts. The second part and the third part were added DNase and RNase respectively and the first part was added with no enzyme. λDNA as well as DNA of Egg drop Syndrome Virus (EDSV-DNA) were employed as controls. It proved that the nucleic acid of NGVEV could be degenerated by DNase but could not by Rnase and the result suggested that NGVEV was a DNA virus with a nucleic acid of approximately 32 kb.
Nucleic acid strand type of NGVEV
Egg drop syndrome viruses (EDSV) DNA[20] and λ DNA were used as controls of double-stranded nucleic acid and Gosling Plague Virus (GPV)[17] was used as control of single-stranded nucleic acid.
Acridine orange staining It is known that the mechanism of acridine orange staining is based on the stain concentration and the nucleic acid space configuration, not on the type of nucleic acid (DNA or RNA). Under ultraviolet light the single-stranded nucleic acid of GPV was brilliant flame red and the nucleic acid of NGVEV and EDSV, along with λ DNA, was apple green. This suggested that the NGVEV nucleic acid was double stranded.
Nuclease digestion It is known that nuclease S1 usually degrades the single-stranded nucleic acid, and is capable of, only at a higher concent ration, degrading double-stranded nucleic acid. The result that under the same condition NGVEV-DNA, along with the controls of EDSV-DNA and λDNA, could not but GPV-DNA could be digested by nuclease S1, confirmed the double stranded nucleic acid type of NGVEV[21-25].
Virus purification and viral structural polypeptide analysis by SDS-PAGE
Virus purification It proved, by cellulose acetate electrophoresis and electronic microscopy examination, that purified virions were obtained and could serve as a qualified material for the viral fine structure observation, the nucleic acid extraction and the viral structural protein analysis. Electronic microscopy of the virus revealed NGVEV virion, spherical or slightly elliptical in shape, was an unenveloped icosahedral structure 60-80 nm in diameter, with an average of 70-90 nm. And by morphological analysis, the virus appeared to fulfill the criteria of adenovirus.
Viral structural polypeptide analysis by SDS-PAGE SDS-PAGE Of the disrupted purified virus particles revealed 15 discrete bands (Figure 1). After scanned and analyzed by automatic gel image-forming and analyzing system, the molecular mass and relative percentage of each structural polypeptide were obtained (Table 1). The fact that VP4, VP7, VP8, VP9 and VP14 were the main structural polypeptides and made up about 90.2622% of the gross protein revealed that NGVEV fulfilled the criteria of adenovirus to great extent[17,26-28]. From the relationship between DNA base number and the molecular mass of the encoded protein, it is known that NGVEV structural protein with an overall molecular weight of 71300 D needed DNA of approximately 19.3 kb to code for. But the viral genome of DNA was about 32 kb. This suggested that NGVEV DNA codes not only for the viral structural protein. This property is shared by other adenoviruses as well[17].
Figure 1.
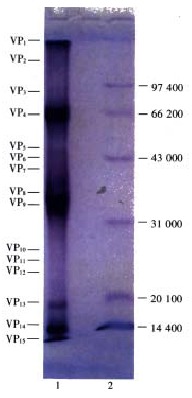
Protein polypetide map of NGVEV-CN virus. 1. Polypetide of NGVEV-CN virus; 2. Low MW. standard protein.
Table 1.
Molecular weights and relative percentages of NGVEV structural polypeptides
| Polyp eptides | VP1 | VP2 | VP3 | VP4 | VP5 | VP6 | VP7 | VP8 | VP9 | VP10 | VP11 | VP12 | VP13 | VP14 | VP15 |
| Molecular weights (Daltons) | 116000 | 104000 | 88000 | 68000 | 60700 | 54000 | 40100 | 36000 | 34500 | 24200 | 22000 | 20000 | 18000 | 14200 | 13400 |
| Relative percentage (%) | 3.0215 | 0.0172 | 0.0014 | 24.5950 | 0.5581 | 0.6225 | 12.6938 | 25.3610 | 18.1807 | 0.1483 | 0.2275 | 0.3826 | 0.9072 | 10.0714 | 3.2118 |
From data above we have got, we know that NGVEV is adenovirus[29-45].
The pathomorphological development observed in goslings artificia lly infected with NGVEV-CN
Signs
The incubation period of the experimental infection mainly varied from 2 to 3 d (36/40 or 85%) and only a few varied from 4 to 5 d; The early signs were: inactivity of the brood, reduced appetite, listlessness, somnolence, loose drooping, ruffled feathers accompanied by drooping of wings. Besides, the voice of the birds was not sonorous as before; At a later stage of this disease, wet vent feathers, soiled vents, inappetence, watery drooping with yellow or whitish yellow mucoid contained could be seen. Brown drooping might be observed in some individuals and the affected birds could not keep balance in walking and standing. Spasmodic prostration and convulsions, kicking spasmodically upwards with both legs might be observed in some affected goslings and opisthotonus might be found in most of the birds which died of this disease. The affected birds usually died of emaciation, extreme weakness, and somnolence. Retarded growth was observed in the affected birds and the body weight might be reduced by one fold compared with the control group. In the experiment group, mortality occurred 4 d postinfection and peaked 10-18 d postinfection and all the birds died at the 25th day postinfection. In the experiment group, altogether 8 birds were killed and 32 died.
Pathological changes
Gross changes
Small intestine In each small intestinal part, no gross lesions was demonstrated one day postinfection and only mild hyperemia could be seen 2 d postinfection; in each part of the small intestine, pronounced hyperemia, mild hemorrhage, obvious mucosa swelling as well as excess mucus production might be observed 3 d postinfection. In addition to pronounced hyperemia as well as excess mucus production, mucosa swelling and serious hemorrhage could be found in each small intestinal part of birds killed 4 d postinfection; severe hemorrhage might be found in each small intestinal part of birds that died of this disease. Besides, swollen intestinal mucosa which appeared bright and contained plenty of mucous secretory products might be observed as well; in addition to the severe hemorrhage which might be demonstrated in each part of the small intestine, a little whitish yellow coagulated fibrous exudate as well as a few pieces of necrotic intestinal epithelia might be found on the intestinal mucosa of birds that died 7-12 d postinfection; coagulative embolus wrapped by yellowish pseudomembrane was first found in birds that died 14 d postinfection and was about 0.2 cm in diameter and over 10 cm in length when first observed. This structure found in birds that died at a later time, which was between 0.5 and 0.7 cm in diameter and over 10 cm in length and whose length might reach 30 cm or more, was bigger than that observed previously and the small intestinal part containing it was 1-2 times distended than the control in appearance (Figure 2). The small intestinal part containing this embolus was much thin in its wall and that without this structure underwent severe hemorrhage with, its mucosa reddened. This structure of embolus was mainly observed in the middle-lower part of the small intestine before the bifurcation of the ceca and most of which was in the form of one section. There were emboluses in the form of two sections, but the number was greatly reduced. The intestinal coagulative embolus observed at necropsy could roughly be classified into 2 types. The first with a diameter of over 0.5 cm and a length of approximately 20 cm was big in size and dense in texture. And this embolus occupying the whole intestinal lumen, whether sectioned transversely or longitudinally, was found to be a two-layed structure. The outer layer, which was a pseudomembrane 0.5 mm-1 mm in depth was dry and brownish in appearance and was composed of the mixture of necrotic tissue and fibrous exudate. The inner layer was the intestinal contents which was dry and dense. The second kind of embolus with the appearance of thin rod and with a diameter of 30 cm or more was obviously thinner but longer than the first and was composed of the coagulative mixture of necrotic intestinal tissue and fibrous exudate. Both kinds of emboluses did not adhere to, and were easily to be separated from the intestinal wall. The wall of the embolus containing part of the intestine was very thin and its transparency was greatly enhanced. The intestinal lesions observed in this disease, especially the coagulative embolus in the middle-lower part of the intestine that occluded the intestinal tract, resembled that observed in the case of gosling plague (GP)[46] to a great extent.
Figure 2.

Particular coagulative embolus was formed in small in testine of death gosling (13 d postinfection), the length is over 40 cm. Haemorrhage was occured in small intestine wall and be dyed red.
Rectum and the cecum Swelling, hyperemia and mild hemorrhage might be observed in birds that died at an early stage of this disease; much mucus was observed and the cloaca was filled with yellowish loose contents in cases that died at a later stage.
Other tissues or organs No gross lesions was found in killed birds. Birds that died at an early stage, usually less than 10 d postinfection, of this disease might show subcutaneous hyperemia or hemorrhage; epcardial mural hyperemia or small punctuate hemorrhages might be observed in a few of the m. The pectoral muscle and the leg muscle underwent hemorrhage and presented themselves dark red in appearance; the liver on which petechial or ecchymotic hemorrhages might be presented underwent venous congestion and was dark red in appearance. The swollen gall bladder which was dilated in appearance and 3-5 times larger than the control was dark greenish red and was full of bile; the kidney underwent hyperemia and mild hemorrhage and presented itself dark red in appearance; no obvious lesions was found in other tissues or organs. In birds that died at a later stage (more than 11 d postinfection), no obvious lesion was found except the liver which appeared dark red and the kidney which underwent mild hyperemia and hemorrhage.
Histopathological lesions
Duodenum In birds killed 1 day postinfection, no lesion was found different from the controls. In birds killed 2-4 d postinfection, some of the epithelial cells covering the villus tips were found sloughed and some of the lamina propria cells underwent coagulative necrosis and presented themselves to be a sheet of red-stained granules among which fibrinoid necrotic interstitial tissue might be detected. In addition, hyperemia of the villi and necrosis of some of the intestinal villus tips were observed (Figure 3). In birds that died 4 d postinfection, epithelium of the mucosa was found denuded and some of the villi might be completely denuded, with the lamina propria exposed. The lamina propria underwent edema. The intestinal glandular cells underwent vacuolar degeneration and were loosely packed. In birds that died 5-10 d postinfection, intestinal epithelial cells were found completely sloughed, parts of the mucosal axletree of lamina propria were found remained, in which large number of erythrocytes were detected. As the infection advanced, a large number of lymphocytes were observed in the swollen lamina propria and in some parts of it much fibrin was found. Most of the intestinal glandular cells underwent vacuolar degeneration, necrosis and were loosely packed; during the course of this disease, the duodenal fibrinonecrotic enteritis was developed and most of the intestinal villi were completely denuded, with the neat separation surface formed (Figure 4). Abundant fibrin, blood cells and bacteria filled the lumen; in birds that died 11 d postinfection, epithelial cells was completely sloughed from the mucosa and the mucosal axletree of lamina propria was exposed. As for the lamina propria, some was infiltrated with abundant lymphocytes and some underwent necrosis and sloughing. Some intestinal glandular cells were contracted and were detached from their surrounding connective tissue, with a space around them formed. The intestinal glandular underwent necrosis, sloughing and a small number of intestinal glandular cells underwent vacuolar degeneration and were loosely packed. The blood vessels in the muscle lays were congested and abundant fibrin as well as sloughed or necrotic cells filled the lumen.
Figure 3.

Many pieces of falled epidermal cells in duodenum cav ity (arrow 1), coagulative necroses was occured in some villus top (arrow 2). (150 × , H.E)
Figure 4.
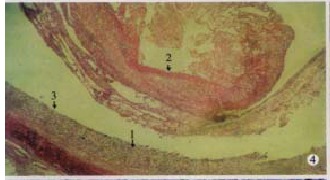
Particular fibrinous and necrosed enteritis of ileum: necrosed mucosae (arrow 1) and fibrinous edudate coagulated into artificial me mbrane and dropped into the cavity (arrow 2), and surface of separation boundry was smooth (arrow 3). (100 × , H.E)
Ileum No differences was found between the birds killed 1-4 d postinfection and the controls; in birds that died 4 d postinfection, epithelial cells covering the villus tips underwent necrosis and sloughing. The glandular cells underwent swelling and vacuolar degeneration and were loosely packed, some of which were represented by their outlines, another part of which might be replaced by abundant proliferated connective tissues and lost their outlines. Some of the intestinal villi were observed completely sloughed from the mucosa and with the prolonged course of this disease, the intestinal villi were observed completely sloughed and the typical fibrinonecrotic enteritis was eventually developed. The pseudomembrane covering the embolus in the intestinal tract was mainly composed of necrotic mucosa, droppings, inflammatory cells, bacterium and the exudate of fibrin (Figure 5).
Figure 5.

Particular fibrinous and necrosed enteritis of ileum: the embo lus consisted of the necrosed mucosal tissue coagulated and dropped materials, it include fibrinous edudate which like thread and inflammatory cells. Fibrin (arrow 1), necrosed cells (arrow 2), inflammatory cells (arrow 3) and bacteria (arrow 4). (400 × , H.E)
Liver No differences was found between the birds killed 1-4 d postinfection and the controls; in some cases that died of this disease, local congestion, mild granular degeneration and fatty degeneration (7/32) (Figure 6) was observed. As for the pathological changes in the case of gosling plague (GP)[46], pronounced inflammatory lesion and focal necrosis of the liver were successively observed 24-48 h postinfection and granular degeneration as well as vacuolar degeneration were observed successively. With the prolonged course of GP, the hepatic cells underwent severe vacuolar degeneration and progressive necrosis, which were most evident in birds that died of this disease. The hepatic cells were swollen and their shape changed from polygonal to round. Many vesicles appeared in the hepatic cytoplasm and thus made it appeared loosely foamy. Cytoplasm dissolution, ballooning degeneration, together with the marked vacuolation of the hepatocytes were observed in more serious cases and the Sudan III staining for fat was negative. With the further development of this disease, hepatocytes underwent rupture, necrosis and dissolution and the necrosis-dissolution foci were developed, in which were light red-stained oedematous fluid as well as a small number of monocytes and lymphocytes. Dehydration, condensation and enhanced cytoplasmic acidophilia occurred in a few hepatocyte cytoplasm, in which round eosinophilic droplets were formed and shared great similarity to that structure observed in the case of human viral hepatitis. The hepatic cords were disorganized in structure and some of the hepatic sinusoid, with the necrosis and dissolution of the hepatocyte, were disrupted and erythrocytes were released. The hepatic interstitial blood vessels and the hepatic sinusoids were congested and loose and lost their normal structure and underwent fibrinoid necrosis. The interlobular monocytes and lymphocytes underwent diffuse proliferation and showed diffuse infiltration. Nodular proliferative foci were formed in some parts of the interlobular tissue.
Figure 6.

Fatty degeneration were occured in liver cells. (500 × , H.E)
Kidney In birds killed 1-4 d postinfection, the main lesions were: the renal tubular tract was not clear in structure, mild grandular degeneration could be seen in the epithelial cells of the renal tubular; in birds that died of this disease 4 d postinfection, hemorrhage focus and granular degeneration of the epithelium of the renal tubular might be found in some cases (19/32); vacuolar degeneration (Figure 7) and abundant sloughed epithelial cells in the ureter tract might be observed in some cases (9/32), Some cases (5/32) were found normal.
Figure 7.
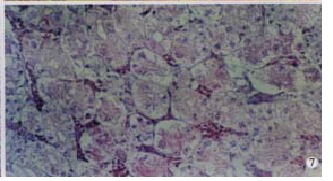
Kidney: Hyperaemia. Granular degeneration was occured seriously in kidney small vessles (arrow 1), or even vacuolar degeneration (arrow 2). (500 × , H.E)
Spleen Mild hyperemia might be observed in some (4/8) of the birds killed 1-4 d postinfection; Mild hyperemia, hemorrhage might be observed in a few (5/32) of the cases that died of this disease and the lymphoid follide of these cases were found equivocal in structure. As for the case of gosling plague[46], no obvious lesions was found during the early stage of this disease and during the later stage, splenic sinusoid might be found congested and contained a large number of monocytes and a small number of heterophiles. Besides, the splenic corpuscles underwent atrophy and was equivocal in structure; lymphocytes underwent necrosis and karyorrhexis; small necrotic foci might be found in the lymphoid nodule; proliferation of monocytes or reticular cells might be observed in the parenchyma of the spleen.
Lung In cases 1 day postinfection, the blood vessels were congested; in cases 2 d postinfection, tertiary bronchuses underwent hyperemia and hemorrhage; in cases 3-4 d postinfection, hyperemia, hemorrhage was found and the vein was congested. The tertiary bronchus and the atrium contained a lot of erythrocytes and the secondary bronchus contained quantities of blood; most of the cases that died 4 d postinfection showed pulmonary congestion and hemorrhage, caseation necrosis foci could only be observed in a few cases (2/32).
Proventriculus No lesions were observed in birds killed 1-4 d postinfection or birds that died less than 5 d postinfection; sloughing of the epithelium from the mucosa was observed in most (30/32) of the birds that died more than 5 d postinfection and there were abundant sloughed cells in the cavity. Besides, sloughed glandular cells might be found in parts of the glandlobular cavities. As for the cases of GP[46], 48 h until death, degeneration, necrosis and sloughing of the epithelial cells of the mucosa might be observed. The lamina propria underwent hyperemia and was infiltrated with inflammatory cells. Compound tubular glandular cells might be observed sloughed; many inflammatory cells and sloughed glandular cells filled the collecting sinusoid.
Heart Most (27/32) of the killed birds were found to be normal and only a few (5/32) that died at an early stage of this disease showed mild hyperemia and hemorrhage in their hearts. But in the case of gosling plague[46], the following might occur 12-36 d postinfection: granular degeneration of the myocardium, Cross striations of the myocardium got dim or disappeared; the myocardium underwent karyorrhexis and karyolysis; small necrotic foci and inflammatory cell infiltration might be observed; Focal pericarditis might developed.
Pancreas, brain and esophagus These organs were found normal. But in the case of GP[46] 72 h postinfection, the following was observed: Interstitial blood vessels were congested, the glandular cells underwent degeneration and was separated from the basal membrane. In birds that died of GP, the acinuses were found disorganized in structure and the glandular cells were necrosed, sloughed, dissolved and finally the necrosis-dissolution focus was formed; the interstitial tissues were infiltrated with inflammatory cells. In the case of GP[46], basically the same lesion, which was not obvious at an early stage, was shown in the cerebrum and the cerebellum; at a later stage, the following might occur: nenix underwent swelling and hyperemia, blood vessels in the cerebral parenchyma were dilated and congested with the perivascular space enlarged, foci formed by the perivacular proliferation of a small number of lymphocyte and monocytes could be seen, matrix of the cerebrum was swollen, the neuron underwent degeneration and vesicles might be observed in its cytoplasm, pykosis was observed and the nucleus was equivocal in structure, the neurogliocytes underwent diffuse proliferation and glial nodules might be formed in some areas within it.
About the structure of inclusion body According to the related reports[17,18,46], intranuclear or intracytoplasmic inclusions might be observed in myocardium, hepatocytes, and epithelial cells of the intestine in the case of GP. Wang[46] reported that eosinophilic granules were found in the hepatocytes. In many cases of poultry infected with adenovirus, inclusion body could be observed[47-52]. But none of these mentioned above was observed in the case of NGVE.
Histopathological and ultrastructure analysis is the key method to understand mechanism of human ang animal disease[53-64]. The histopathological change of Goslings be infected with NGVEV can help us to understand mechanism of NGVE, Pathological characteristic could be provide helpful materials for the NGVE diagnosis and differential diagnosis from GP.
ACKNOWLEDGEMENTS
We would like to thanks associated professor Fang Jing for making, reading the pathological slides and taking the pathological photos. We also grateful to technician Zhou Yi for his work about transmission electronic microscope.
Footnotes
Edited by Wang WQ
Supported by the National Nature Scientific Foundation of China, No.39970561 and the Funds for the Leaders in Academia of Sichuan Province, No.9900009240018
References
- 1.Cheng AC. Research on a new infectious disease of goslings. Zhongguo Shouyi Keji. 1998;28:3–6. [Google Scholar]
- 2.Kardi V, Szegletes E. Use of ELISA procedures for the detection of Derzsy's disease virus of geese and of antibodies produced against it. Avian Pathol. 1996;25:25–34. doi: 10.1080/03079459608419117. [DOI] [PubMed] [Google Scholar]
- 3.Zsak L, Kisary J. Characterisation of adenoviruses isolated from geese. Avian Pathol. 1984;13:253–264. doi: 10.1080/03079458408418529. [DOI] [PubMed] [Google Scholar]
- 4.Gough RE. Application of the agar gel precipitin and virus neutralisation tests to the serological study of goose parvovirus. Avian Pathol. 1984;13:501–509. doi: 10.1080/03079458408418551. [DOI] [PubMed] [Google Scholar]
- 5.Gough RE. Persistence of parvovirus antibody in geese that have survived Derzsy's disease. Avian Pathol. 1987;16:327–330. doi: 10.1080/03079458708436379. [DOI] [PubMed] [Google Scholar]
- 6.Roszkowski J, Gaździński P, Kozaczyński W, Bartoszcze M. Application of the immunoperoxidase technique FDR the detection of Derzsy's disease virus antigen in cell culture and goslings. Avian Pathol. 1982;11:571–578. doi: 10.1080/03079458208436132. [DOI] [PubMed] [Google Scholar]
- 7.Cheng AC. Isolation, identification and properties of goslings new type viral enteritis virus. Xumu Shouyi Xuebao. 2000;31:548–556. [Google Scholar]
- 8.Cheng AC. Studies on Dot Immunogold Staining (Dot-IGS) for detecting the ant ibodies against new gosling viral enteritis (NGVE) Zhongguo Shouyi Keji. 1999;29:3–6. [Google Scholar]
- 9.Cheng AC. Study on the agar gel precipitin test to detect antigen and anti body of the gosling new type viral enteritis. Zhongguo Shouyi Zazhi. 1999;25:3–6. [Google Scholar]
- 10.Gough RE, Borland ED, Keymer IF, Stuart JC. An outbreak of duck virus enteritis in commercial ducks and geese in East Anglia. Vet Rec. 1987;121:85. doi: 10.1136/vr.121.4.85. [DOI] [PubMed] [Google Scholar]
- 11.Kisary J, Zsak L. Comparative studies on duck viral enteritis (DVE) virus strains in geese. Avian Pathol. 1983;12:395–408. doi: 10.1080/03079458308436185. [DOI] [PubMed] [Google Scholar]
- 12.Gough RE. Laboratory confirmed outbreaks of duck virus enteritis (duck plague) in the United Kingdom from 1977 to 1982. Vet Rec. 1984;114:262–265. doi: 10.1136/vr.114.11.262. [DOI] [PubMed] [Google Scholar]
- 13.Gough RE, Alexander DJ. Duck virus enteritis in waterfowl. Vet Rec. 1987;121:72. doi: 10.1136/vr.121.3.72-b. [DOI] [PubMed] [Google Scholar]
- 14.Leibovitz L. Gross and histopathologic changes of duck plague (duck virus enteritis) Am J Vet Res. 1971;32:275–290. [PubMed] [Google Scholar]
- 15.Gough RE, Spackman D, Collins MS. Isolation and characterisation of a parvovirus from goslings. Vet Rec. 1981;108:399–400. doi: 10.1136/vr.108.18.399. [DOI] [PubMed] [Google Scholar]
- 16.Mengeling WL, Paul PS, Bunn TO, Ridpath JF. Antigenic relationships among autonomous parvoviruses. J Gen Virol. 1986;67(Pt 12):2839–2844. doi: 10.1099/0022-1317-67-12-2839. [DOI] [PubMed] [Google Scholar]
- 17.Yin Z, Liu JH. Animal Virology. 2nd editor. Beijing: Science Press; 1997. pp. 67–82, 204-437, 1145-1174. [Google Scholar]
- 18.Encyclopedia of Virology, Edited by Robert G. Webster and Allan Grano ff. Academic Press. Beijing: Science Press; 1994. pp. 1–23, 1052-1067. [Google Scholar]
- 19.Zhang LX, Zhang TF, Li LA. Experimental method and technology for bio chemical. Beijing: Higher Education Press; 1984. pp. 94–119. [Google Scholar]
- 20.Luo YS, Li TX, Feng F, Zhao L. Study on egg drop syndrome 1976 virus II: Protein characteristics of GC2 strain. Xumu Shouyi Xuebao. 1995;26:534–536. [Google Scholar]
- 21.Adam SA, Dreyfuss G. Adenovirus proteins associated with mRNA and hnRNA in infected HeLa cells. J Virol. 1987;61:3276–3283. doi: 10.1128/jvi.61.10.3276-3283.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Stewart PL, Burnett RM, Cyrklaff M, Fuller SD. Image reconstruction reveals the complex molecular organization of adenovirus. Cell. 1991;67:145–154. doi: 10.1016/0092-8674(91)90578-m. [DOI] [PubMed] [Google Scholar]
- 23.Akopian TA, Kruglyak VA, Rivkina MB, Naroditsky BS, Tikhonenko TI. Sequence of an avian adenovirus (CELO) DNA fragment (0-11.2%) Nucleic Acids Res. 1990;18:2825. doi: 10.1093/nar/18.9.2825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wold WS, Gooding LR. Region E3 of adenovirus: a cassette of genes involved in host immunosurveillance and virus-cell interactions. Virology. 1991;184:1–8. doi: 10.1016/0042-6822(91)90815-s. [DOI] [PubMed] [Google Scholar]
- 25.Sheppard M, Trist H. Characterization of the avian adenovirus penton base. Virology. 1992;188:881–886. doi: 10.1016/0042-6822(92)90546-2. [DOI] [PubMed] [Google Scholar]
- 26.Li P, Bellett AJ, Parish CR. The structural proteins of chick embryo lethal orphan virus (fowl adenovirus type 1) J Gen Virol. 1984;65(Pt 10):1803–1815. doi: 10.1099/0022-1317-65-10-1803. [DOI] [PubMed] [Google Scholar]
- 27.Hasson TB, Soloway PD, Ornelles DA, Doerfler W, Shenk T. Adenovirus L1 52- and 55-kilodalton proteins are required for assembly of virions. J Virol. 1989;63:3612–3621. doi: 10.1128/jvi.63.9.3612-3621.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Cepko CL, Sharp PA. Assembly of adenovirus major capsid protein is mediated by a nonvirion protein. Cell. 1982;31:407–415. doi: 10.1016/0092-8674(82)90134-9. [DOI] [PubMed] [Google Scholar]
- 29.Bandara LR, La Thangue NB. Adenovirus E1a prevents the retinoblastoma gene product from complexing with a cellular transcription factor. Nature. 1991;351:494–497. doi: 10.1038/351494a0. [DOI] [PubMed] [Google Scholar]
- 30.Fatemie-Nainie S, Marusyk R. Biophysical and serologic comparison of four equine adenovirus isolates. Am J Vet Res. 1979;40:521–528. [PubMed] [Google Scholar]
- 31.Swain P, Kataria JM, Verma KC. Biological characterisation of an Indian isolate of egg drop syndrome-76 virus. Res Vet Sci. 1993;55:396–397. doi: 10.1016/0034-5288(93)90118-y. [DOI] [PubMed] [Google Scholar]
- 32.Perdue ML, Cohen JC, Kemp MC, Randall CC, O'Callaghan DJ. Characterization of three species of nucleocapsids of equine herpesvirus type-1 (EHV-1) Virology. 1975;64:187–204. doi: 10.1016/0042-6822(75)90091-4. [DOI] [PubMed] [Google Scholar]
- 33.Ishiyama T, Shinagawa M, Sato G, Fujinaga K, Padmanabhan R. Generation of packaging-defective DNA molecules of equine adenovirus. Virology. 1986;151:66–76. doi: 10.1016/0042-6822(86)90104-2. [DOI] [PubMed] [Google Scholar]
- 34.Sheppard M, Trist H. Characterization of the avian adenovirus penton base. Virology. 1992;188:881–886. doi: 10.1016/0042-6822(92)90546-2. [DOI] [PubMed] [Google Scholar]
- 35.Anderson CW, Young ME, Flint SJ. Characterization of the adenovirus 2 virion protein, mu. Virology. 1989;172:506–512. doi: 10.1016/0042-6822(89)90193-1. [DOI] [PubMed] [Google Scholar]
- 36.Sheppard M, Trist H. The identification of genes for the major core proteins of fowl adenovirus serotype 10. Arch Virol. 1993;132:443–449. doi: 10.1007/BF01309553. [DOI] [PubMed] [Google Scholar]
- 37.Das BB, Pradhan HK. Outbreaks of egg drop syndrome due to EDS-76 virus in quail (Coturnix coturnix japonica) Vet Rec. 1992;131:264–265. doi: 10.1136/vr.131.12.264. [DOI] [PubMed] [Google Scholar]
- 38.Dren CN, Nemeth I, Sari I, Ratz F, Glavits R, Somogyi P. Isolation of a reticuloendotheliosis-like virus from naturally occurring lymphoreticular tumours of domestic goose. Avian Pathol. 1988;17:259–277. doi: 10.1080/03079458808436446. [DOI] [PubMed] [Google Scholar]
- 39.Stewart PL, Burnett RM, Cyrklaff M, Fuller SD. Image reconstruction reveals the complex molecular organization of adenovirus. Cell. 1991;67:145–154. doi: 10.1016/0092-8674(91)90578-m. [DOI] [PubMed] [Google Scholar]
- 40.Cepko CL, Sharp PA. Assembly of adenovirus major capsid protein is mediated by a nonvirion protein. Cell. 1982;31:407–415. doi: 10.1016/0092-8674(82)90134-9. [DOI] [PubMed] [Google Scholar]
- 41.Ishiyama T, Shinagawa M, Sato G, Fujinaga K, Padmanabhan R. Generation of packaging-defective DNA molecules of equine adenovirus. Virology. 1986;151:66–76. doi: 10.1016/0042-6822(86)90104-2. [DOI] [PubMed] [Google Scholar]
- 42.Anderson CW, Young ME, Flint SJ. Characterization of the adenovirus 2 virion protein, mu. Virology. 1989;172:506–512. doi: 10.1016/0042-6822(89)90193-1. [DOI] [PubMed] [Google Scholar]
- 43.Sheppard M, Werner W. Expression of fowl adenovirus type 10 antigens in Escherichia coli. Vet Microbiol. 1990;24:105–112. doi: 10.1016/0378-1135(90)90057-3. [DOI] [PubMed] [Google Scholar]
- 44.Nazerian K, Lee LF, Payne WS. Structural polypeptides of type II avian adenoviruses analyzed by monoclonal and polyclonal antibodies. Avian Dis. 1991;35:572–578. [PubMed] [Google Scholar]
- 45.Belin MT, Boulanger P. Processing of vimentin occurs during the early stages of adenovirus infection. J Virol. 1987;61:2559–2566. doi: 10.1128/jvi.61.8.2559-2566.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Wang KY. Pathomorphological study on experimental gosling plague. Xumu Shouyi Xuebao. 1998;29:377–384. [Google Scholar]
- 47.Anderson CW, Young ME, Flint SJ. Characterization of the adenovirus 2 virion protein, mu. Virology. 1989;172:506–512. doi: 10.1016/0042-6822(89)90193-1. [DOI] [PubMed] [Google Scholar]
- 48.Riddell C. Viral hepatitis in domestic geese in Saskatchewan. Avian Dis. 1984;28:774–782. [PubMed] [Google Scholar]
- 49.Meteyer CU, Mohammed HO, Chin RP, Bickford AA, Trampel DW, Klein PN. Relationship between age of flock seroconversion to hemorrhagic enteritis virus and appearance of adenoviral inclusions in the spleen and renal tubule epithelia of turkeys. Avian Dis. 1992;36:88–96. [PubMed] [Google Scholar]
- 50.Trampel DW, Meteyer CU, Bickford AA. Hemorrhagic enteritis virus inclusions in turkey renal tubular epithelium. Avian Dis. 1992;36:1086–1091. [PubMed] [Google Scholar]
- 51.Goodwin MA. Adenovirus inclusion body ventriculitis in chickens and captive bobwhite quail (Colinus virginianus) Avian Dis. 1993;37:568–571. [PubMed] [Google Scholar]
- 52.Lammert F, Südfeld S, Busch N, Matern S. Cholesterol crystal binding of biliary immunoglobulin A: visualization by fluorescence light microscopy. World J Gastroenterol. 2001;7:198–202. doi: 10.3748/wjg.v7.i2.198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Ji XL, Gong GH, Wang MW. Histopathological study of colon mucosa in the elderly. Xin Xiaohuabingxue Zazhi. 1995;3:154–155. [Google Scholar]
- 54.Liu YQ, Li DX, Cui GL, Tang FA, Li ZF. Immunohistochemical study of gastrin-immunoreactive cells in benign and malignant biospy tissues of stomach. Xin Xiaohuabingxue Zazhi. 1995;3:72–73. [Google Scholar]
- 55.Cai JL, Zhao DC, Li BQ. Study on the ultrastructure of rectal exfoliative cells. Xin Xiaohuabingxue Zazhi. 1995;3:142–144. [Google Scholar]
- 56.Cao B, Ge ZJ, Meng XY, Bi SZ. Ultrastructural study of colonal mucosa in patients with irritable bowel syndrome. Xin Xiaohuabingxue Zazhi. 1994;2:236–237. [Google Scholar]
- 57.Li QM, Wu YF, Wang LY, Huang QH, Deng YF, Wu JL, Chen W. Ultrastructu re of gastric mucosa and pathogenesis of spleen-energy deficiency. Xin Xiaohuabingxue Zazhi. 1994;2:156–157. [Google Scholar]
- 58.Hu SX, Tang RJ, Zuo DY, Chen PH. Histopathological study of pancreatic duct in acute fulminant pancreatitis. China Natl J New Gastroenterol. 1996;2:243–245. [Google Scholar]
- 59.Tang FC, Zhang YF, Xu YD, Zhong SQ, Wang XP, Wang YX. Scanning electron microscopic studies of lymphatic corrosion casts in the rabbit appendix. China Natl J New Gastroenterol. 1996;2:238–240. [Google Scholar]
- 60.Yang SM, Lin BZ, Fang Y, Zheng Y. Ultrastructural observation on relation of H. pylori to gastric epithelia in chronic gastritis and peptic ulcer. China Natl J New Gastroenterol. 1996;2:152–154. [Google Scholar]
- 61.Yu JY, D′Adda T. Quantitative ultrastructure analysis of neuroendocr ine cells of gastric mucosa in normal and pathological conditions. China Natl J New Gastroenterol. 1996;2:155–157. [Google Scholar]
- 62.Ji XL, Liu YX, Wang YH, Zhao H. Histopathological study of hepatocellular carcinoma after transcatheter hepatic arterial embolization. China Natl J New Gastroenterol. 1996;2:79–81. [Google Scholar]
- 63.Bu JK, Zhang ZL, Zhao JX. Stereological and ultrastructural research on the rats with chronic gastritis treated based on the differential diagnosis of traditional Chinese medicine (TCM) China Natl J New Gastroenterol. 1996;2:76–78. [Google Scholar]
- 64.Li YQ, Bing HB. Light and electron microscopic observation of malignant gastric leiomyoblastoma. China Natl J New Gastroenterol. 1996;2:61–62. [Google Scholar]