

INTRODUCTION
Plasminogen activator inhibitor type 1 (PAI-1), an approximately Mr50000 glycoprotein, is the major physiological inhibitor of plasminogen activators. It is not only the priming factor for atherosclerosis and coronary thrombosis[1-3], but also participates in the genesis of chronic hepatitis and liver fibrosis[4-11]. However, there has been no available report yet about the research of hepatic PAI-1 gene expression in hyperlipidemia and fatty liver. The present study AIMed to explore the change of hepatic PAI-1 mRNA and its plasma activity by means of animal model.
MATERIALS AND METHODS
Animal model
Seventeen male New Zealand white rabbits weighing 2200 g-2500 g were randomly divided into two groups. The seven rabbits of control group were fed with normal rabbit diet. The ten rabbits of model group were fed with a fat-rich diet, which was prepared by addition of 10 g•kg¯¹ cholesterol and 100 g•kg¯¹ lard oil to the normal diet. Vein blood samples of rabbits were collected before the beginning of study and at the 6th week and 12th week of the study respectively. Then the plasma samples were obtained and maintained at -70 °C until assayed. When the rabbits were sacrificed at the 12th week, their body mass and wet liver mass were measured. And two small samples of liver tissue were obtained from the right hepatic lobe. One sample was fixed in formalin, embedded in paraffin and stained with H&E. The other sample was stored in liquid nitrogen for detection of PAI-1 mRNA.
Measurement of plasma biochemistry
Plasma levels of alanine aminotransferase (ALT), aspartate aminotransferase (AST), triglycerides (TG) and total cholesterol (TCH) were determined by automatic biochemical analysis instrument (Olympus AU 1000, made in Japan). Plasma PAI-1 activity was determined by a chromogenic activity kit purchased from Biotechnology Co., Ltd. of the former Shanghai Medical University.
Detection of hepatic PAI-1 mRNA
The primer for rabbit PAI-1 was designed based on its complement DNA (cDNA) sequence published previously[6,12,13] and synthesized by DNA synthesis instrument. The sequence is 5’-ATGGAATTCCCGTGGAACAAGAATGAGATCAG-3’and 5’-TGAGCCATCATGGGCACAGAG. The primer for β-actin was given by Shanghai Institute of Cell Biology of the Chinese Academy of Sciences, and the sequence is 5’-ATCTGGCACCACACCTTCTACAATGAGCTGC-3’ and 5’-CGTCATACTCCTGCTTGCTGATCCACATCTGC-3’. The samples of liver tissues were cut into pieces quickly and the total RNAs were extracted with Trizol Reagent (Gibco BRL Co., Ltd.). Reverse transcription-polymerase chain reaction (RT-PCR) was carried out to amplificate the mRNA of PAI-1 and β-actin, and followed by electrophoresis of the products, photography and computer image scan. And the results were validated by the OD value of β-actin because of its equal expression in various tissues.
Statistical analysis
All results were expressed as mean ± SD. Statistical differences between means were determined by Student t test. P < 0.05 was selected to reflect significance.
RESULTS
General condition
During the experiment, no animal death occurred in both groups. The body mass of rabbits in the model group increased more rapidly than in the control group. The model group had significantly greater body mass and liver index (wet liver mass per body mass) than the control group (P < 0.05 respectively, Table 1). Compared with the control group, the plasma TCh and TG levels in the model group already increased significantly at the 6th week (P < 0.05 vs controls) and increased more significantly at the 12th week (P < 0.01 vs controls). However, the plasma levels of ALT and AST in the model group only had a tendency to increase as compared with those of control group (Table 2).
Table 1.
The changes of body mass and liver index
| Groups | n |
m (body)/(kg) |
Liver index /(g•kg¯¹) End | |
| Start | End | |||
| Control | 7 | 2.4 ± 0.3 | 3.1 ± 0.2 | 26 ± 3 |
| Model | 10 | 2.3 ± 0.6 | 3.4 ± 0.1a | 31 ± 2a |
P < 0.05 vs control.
Table 2.
Plasma biochemical test at the 12th week
| Groups | n | c (TG)/(mmol•L¯¹) | c (TCh)/(mmol•L¯¹) | b (ALT)/(nkat•L¯¹) | b (AST)/(nkat•L¯¹) |
| Control | 7 | 0.34 ± 0.11 | 0.76 ± 0.17 | 372 ± 130 | 215 ± 45 |
| Model | 10 | 0.54 ± 0.17b | 26.40 ± 3.89b | 505 ± 215 | 263 ± 127 |
P < 0.01 vs control.
Changes of plasma PAI-1 activity
In the model group, the plasma PAI-1 activity of rabbits already increased significantly at the 6th week (P < 0.05) and increased more significantly at the 12th week (P < 0.01, Table 3) compared with those at the beginning of the experiment. Moreover, the difference in plasma PAI-1 activity between the two groups at the same period also had highly statistical significance (P < 0.05 and P < 0.01, respectively).
Table 3.
The changes of plasma PAI-1 activity (× 103 AU•L¯¹)
| Groups | n | 0 wk | 6 wk | 12 wk |
| Control | 7 | 5.6 ± 2.6 | 6.8 ± 1.4 | 4.8 ± 2.2 |
| Model | 10 | 4.9 ± 3.1 | 7.7 ± 1.1a | 14.0 ± 2.5b |
P < 0.05,
P < 0.01 vs control.
Changes of liver pathology
In the gross, the livers of the model group enlarged markedly as compared with those of control group. Under the light microscope, all the liver tissue sections stained with H&E of the model group showed diffuse and severe foamy steatosis of hepatocyte with mild necrosis and inflammatory cell infiltration, among which monocytes were dominant. And these lesions were located mainly in portal areas.
Expression of hepatic PAI-1 mRNA
The mRNA fragments of PAI-1 and β-actin were 360 bp and 828 bp respectively (Figure 1, Figure 2). The lanes (No.3, 4, 5) of hepatic PAI-1 mRNA of the model group were markedly more intense than those of control group (No.1, 2). The ratio of hepatic PAI-1 mRNA to β-actin mRNA of model group was significantly greater than that of the control group (3.474 ± 0.051 vs 1.210 ± 0.031, P < 0.01).
Figure 1.
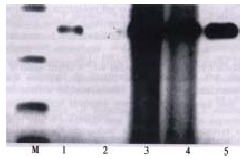
The expression of hepatic PAI-1 mRNA.1, 2: control group; 3, 4, 5: model group; M: marker
Figure 2.
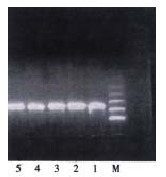
The expression of hepatic β-action mRNA.1, 2: control group; 3, 4, 5: model group; M: marker
DISCUSSION
Fibrinolysis system has an inactive zymogen that is called plasminogen. It can be activated by plasminogen activators and be converted to plasmin[11]. Plasmin can not only degrade the fibrin and remove it from the blood circulation, but also degrade extracellular matrix (ECM) directly and protect the tissues from fibrosis[1-11]. Among all fibrinolysis components, PAI-1 plays a central role in the pathophysiology of cardioangiological diseases. It is the major physiological inhibitor of urokinase plasminogen activator (uPA) and tissue plasminogen activator (tPA). The plasma PAI-1 regulates the plasmin cascade by its interaction with the tPA[1,2]. The increased activity of plasma PAI-1 could inactive the tPA in circulation, and the PAI-1 depositing in ECM could accelerate the clearance of tPA in tissues. PAI-1 exists in all of human tissues. Analysis of human tissues for PAI-1 antigen and activity has shown that the greatest quantity and highest specific activity were found in the liver. In plasma and ECM, the free PAI-1 produced by hepatocytes, endothelial cells and smooth muscle cells binds vitronectin (VN) to maintain the stable activity. The binding PAI-1 which increases the anti-fibrinolysis activity of blood vessels and accelerates the deposition of ECM can in duce the genesis of atherosclerosis, coronary heart disease, pulmonary fibrosis and kidney fibrosis[1-3]. Recent studies have shown that the activity of PAI-1 is also associated with the occurrence and progression of chronic viral hepatitis and liver fibrosis. With the progression of chronic liver disease, the plasma PAI-1 antigen level and expression of PAI-1 in the liver increased gradually and culminated in the stage of liver cirrhosis, then decreased with the deterioration of liver function (Child-Pugh classification A to C)[4-8]. In vitro, hepatic stellate cells (HSC), also known as hepatic lipocytes, fat-storing cells, or Ito cells, could express PAI-1 when activated[12-15]. And previous studies have already demonstrated that HSC played a critical role in the genesis and development of liver fibrosis[16,17]. However, there has been no report yet about the effect of PAI-1 on the pathogenesis of fatty liver.
The present study established a rabbit model with overweight, hyperlipidemia and fatty liver via a fat-and-cholesterol-rich diet. The rabbits of this model had significantly greater body weight and higher plasma lipid levels than the normal control rabbits and their serum levels of ALT and AST had a tendency to increase. In hepatic histopathology, this model showed severe hepatocyte steatosis with mild hepatocyte necrosis and inflammatory cell infiltration. These characteristics resemble the presentation of human fatty liver associated with obesity and hyperlipidemia[18]. So this model can be used to explore the pathogenesis of nonalcoholic fatty liver. In the present study, while hyperlipidemia occurred in the model rabbits after 6 wk of the fatty- rich diets, the plasma PAI-1 activity also increased. Then the plasma lipid and the plasma PAI-1 activity increased more significantly at the 12th week. Moreover, it was shown in the RT-PCR that PAI-1 mRNA was markedly intensein the liver with steatosis.
It is known that various factors could influence the expression and translation of hepatic PAI-1 gene. In the non-alcoholic fatty liver with obesity and hyper lipidemia, many accompanying factors such as lipoprotein (VLDL, LDL), hyperinsul inemia or gut-derived endotoxemia can stimulate hepatic PAI-1 production[19-22]. When the cytokines such as IL-1, TNF, PDGF, TGF-β or IGF-1 exist simultaneously, hepatic PAI-1 production will be further increased[23-28]. Besides, other factors such as dexamethasone, prothrombin, or angiotensin II (Ang II) can also accelerate the secretion of PAI-1[29-32]. Subsequently, the increase in PAI-1 caused the imbalance of tPA and PAI-1, which leads to the decrease in fibrinolysis activity of hepatic microcirculation and degrading of hepatic ECM. Then the dysfunction of hepatic microcirculation, even the microthrombosis could be induced and the occurrence and progression of liver fibrosis promoted[5-11]. However, some studies have indicated that PAI-1 can inhibit the conversion of inactive TGF-β (transforming growth factor-β) into active TGF-β, so that PAI-1 might inhibit liver fibrogenesis since TGF-β can promote the production of ECM and inhibit its degrading[4]. It is therefore, necessary to reinforce the study of the relationship between PAI-1 and fibrosis in fatty liver in order to find an effective method for the prevention and treatment of liver fibrosis.
Footnotes
Edited by Ma JY
References
- 1.Juhan-Vague I, Alessi MC, Morange PE. Hypofibrinolysis and increased PAI-1 are linked to atherothrombosis via insulin resistance and obesity. Ann Med. 2000;32 Suppl 1:78–84. [PubMed] [Google Scholar]
- 2.Nordt TK, Peter K, Ruef J, Kübler W, Bode C. Plasminogen activator inhibitor type-1 (PAI-1) and its role in cardiovascular disease. Thromb Haemost. 1999;82 Suppl 1:14–18. [PubMed] [Google Scholar]
- 3.Mezzano D, Muñoz X, Martínez C, Cuevas A, Panes O, Aranda E, Guasch V, Strobel P, Muñoz B, Rodríguez S, et al. Vegetarians and cardiovascular risk factors: hemostasis, inflammatory markers and plasma homocysteine. Thromb Haemost. 1999;81:913–917. [PubMed] [Google Scholar]
- 4.Inuzuka S, Ueno T, Torimura T, Tamaki S, Sugawara H, Sakata R, Kusaba N, Sata M, Tanikawa K. The significance of colocalization of plasminogen activator inhibitor-1 and vitronectin in hepatic fibrosis. Scand J Gastroenterol. 1997;32:1052–1060. doi: 10.3109/00365529709011224. [DOI] [PubMed] [Google Scholar]
- 5.Piscaglia F, Siringo S, Hermida RC, Legnani C, Valgimigli M, Donati G, Palareti G, Gramantieri L, Gaiani S, Burroughs AK, et al. Diurnal changes of fibrinolysis in patients with liver cirrhosis and esophageal varices. Hepatology. 2000;31:349–357. doi: 10.1002/hep.510310214. [DOI] [PubMed] [Google Scholar]
- 6.Zhang LP, Takahara T, Yata Y, Furui K, Jin B, Kawada N, Watanabe A. Increased expression of plasminogen activator and plasminogen activator inhibitor during liver fibrogenesis of rats: role of stellate cells. J Hepatol. 1999;31:703–711. doi: 10.1016/s0168-8278(99)80351-1. [DOI] [PubMed] [Google Scholar]
- 7.Hayashi T, Kamogawa A, Ro S, Yamaguchi K, Kobayashi Y, Takahashi Y, Murayama M. Plasma from patients with cirrhosis increases tissue plasminogen activator release from vascular endothelial cells in vitro. Liver. 1998;18:186–190. doi: 10.1111/j.1600-0676.1998.tb00148.x. [DOI] [PubMed] [Google Scholar]
- 8.Raya-Sánchez JM, González-Reimers E, Rodríguez-Martín JM, Santolaria-Fernández F, Molina-Pérez M, Rodríguez-Moreno F, Martínez-Riera A. Coagulation inhibitors in alcoholic liver cirrhosis. Alcohol. 1998;15:19–23. doi: 10.1016/s0741-8329(97)00082-7. [DOI] [PubMed] [Google Scholar]
- 9.Fan J, Chen L, Zeng M. [Effects of pravastatin on hepatic plasminogen activator inhibitor 1 mRNA expression in rabbits with fatty liver] Zhonghua Gan Zang Bing Za Zhi. 2000;8:70–72. [PubMed] [Google Scholar]
- 10.Fan JG. Steatohepatitis studies in China. Shijie Huaren Xiaohua Zazhi. 2001;9:6–10. [Google Scholar]
- 11.Urano T, Suzuki Y, Arakida M, Kanamori M, Takada A. The expression of exercise-induced tPA activity in blood is regulated by the basal level of PAI-1. Thromb Haemost. 2001;85:751–752. [PubMed] [Google Scholar]
- 12.Inagaki Y, Mamura M, Kanamaru Y, Greenwel P, Nemoto T, Takehara K, Ten Dijke P, Nakao A. Constitutive phosphorylation and nuclear localization of Smad3 are correlated with increased collagen gene transcription in activated hepatic stellate cells. J Cell Physiol. 2001;187:117–123. doi: 10.1002/1097-4652(2001)9999:9999<00::AID-JCP1059>3.0.CO;2-S. [DOI] [PubMed] [Google Scholar]
- 13.Suzuki Y, Urano T, Ihara H, Nakajima T, Nagai N, Takada Y, Taminato T, Takada A. Bezafibrate attenuates the overexpression of plasminogen activator inhibitor-1 messenger RNA by a combination of mono-unsaturated fatty acid and insulin in hepG2 cells. Life Sci. 2001;68:1827–1837. doi: 10.1016/s0024-3205(01)00976-6. [DOI] [PubMed] [Google Scholar]
- 14.Shimizu M, Hara A, Okuno M, Matsuno H, Okada K, Ueshima S, Matsuo O, Niwa M, Akita K, Yamada Y, et al. Mechanism of retarded liver regeneration in plasminogen activator-deficient mice: impaired activation of hepatocyte growth factor after Fas-mediated massive hepatic apoptosis. Hepatology. 2001;33:569–576. doi: 10.1053/jhep.2001.22650. [DOI] [PubMed] [Google Scholar]
- 15.Uno S, Nakamura M, Ohomagari Y, Matsuyama S, Seki T, Ariga T. Regulation of tissue-type plasminogen activator (tPA) and type-1 plasminogen activator inhibitor (PAI-1) gene expression in rat hepatocytes in primary culture. J Biochem. 1998;123:806–812. doi: 10.1093/oxfordjournals.jbchem.a022008. [DOI] [PubMed] [Google Scholar]
- 16.Lu LG, Zeng MD, Li JQ, Hua J, Fan JG, Fan ZP, Qiu DK. Effect of lipid on proliferation and activation of rat hepatic stellate cells (I) World J Gastroenterol. 1998;4:497–499. doi: 10.3748/wjg.v4.i6.497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Lu LG, Zeng MD, Li JQ, Hua J, Fan JG, Qiu DK. Study on the role of free fatty acids in proliferation of rat hepatic stellate cells (II) World J Gastroenterol. 1998;4:500–502. [PubMed] [Google Scholar]
- 18.Fan JG, Zeng MD, Wang GL. Pathogenesis of fatty liver. Shijie Huaren Xiaohua Zazhi. 1999;7:75–76. [Google Scholar]
- 19.Bastard JP, Piéroni L, Hainque B. Relationship between plasma plasminogen activator inhibitor 1 and insulin resistance. Diabetes Metab Res Rev. 2000;16:192–201. doi: 10.1002/1520-7560(200005/06)16:3<192::aid-dmrr114>3.0.co;2-g. [DOI] [PubMed] [Google Scholar]
- 20.Testa R, Bonfigli AR, Sirolla C, Pieri C, Marra M, Antonicelli R, Manfrini S, Compagnucci P, Testa I. A strong inverse relationship between PAI-1 and Lp (a) in hypertensive Type 2 diabetic patients. Diabetes Nutr Metab. 1999;12:400–406. [PubMed] [Google Scholar]
- 21.Nordt TK, Sawa H, Fujii S, Bode C, Sobel BE. Augmentation of arterial endothelial cell expression of the plasminogen activator inhibitor type-1 (PAI-1) gene by proinsulin and insulin in vivo. J Mol Cell Cardiol. 1998;30:1535–1543. doi: 10.1006/jmcc.1998.0719. [DOI] [PubMed] [Google Scholar]
- 22.Nordt TK, Peter K, Bode C, Sobel BE. Differential regulation by troglitazone of plasminogen activator inhibitor type 1 in human hepatic and vascular cells. J Clin Endocrinol Metab. 2000;85:1563–1568. doi: 10.1210/jcem.85.4.6525. [DOI] [PubMed] [Google Scholar]
- 23.Inoue K, Sugawara Y, Kubota K, Takayama T, Makuuchi M. Induction of type 1 plasminogen activator inhibitor in human liver ischemia and reperfusion. J Hepatol. 2000;33:407–414. doi: 10.1016/s0168-8278(00)80276-7. [DOI] [PubMed] [Google Scholar]
- 24.Zhou L, Hayashi Y, Itoh T, Wang W, Rui J, Itoh H. Expression of urokinase-type plasminogen activator, urokinase-type plasminogen activator receptor, and plasminogen activator inhibitor-1 and -2 in hepatocellular carcinoma. Pathol Int. 2000;50:392–397. doi: 10.1046/j.1440-1827.2000.01059.x. [DOI] [PubMed] [Google Scholar]
- 25.Itoh T, Hayashi Y, Kanamaru T, Morita Y, Suzuki S, Wang W, Zhou L, Rui JA, Yamamoto M, Kuroda Y, et al. Clinical significance of urokinase-type plasminogen activator activity in hepatocellular carcinoma. J Gastroenterol Hepatol. 2000;15:422–430. doi: 10.1046/j.1440-1746.2000.02150.x. [DOI] [PubMed] [Google Scholar]
- 26.Matsuzaki K, Date M, Furukawa F, Tahashi Y, Matsushita M, Sakitani K, Yamashiki N, Seki T, Saito H, Nishizawa M, et al. Autocrine stimulatory mechanism by transforming growth factor beta in human hepatocellular carcinoma. Cancer Res. 2000;60:1394–1402. [PubMed] [Google Scholar]
- 27.Crookston KP, Marsh CL, Chandler WL. A kinetic model of the circulatory regulation of tissue plasminogen activator during orthotopic liver transplantation. Blood Coagul Fibrinolysis. 2000;11:79–88. [PubMed] [Google Scholar]
- 28.Kietzmann T, Roth U, Jungermann K. Induction of the plasminogen activator inhibitor-1 gene expression by mild hypoxia via a hypoxia response element binding the hypoxia-inducible factor-1 in rat hepatocytes. Blood. 1999;94:4177–4185. [PubMed] [Google Scholar]
- 29.Seki T, Healy AM, Fletcher DS, Noguchi T, Gelehrter TD. IL-1beta mediates induction of hepatic type 1 plasminogen activator inhibitor in response to local tissue injury. Am J Physiol. 1999;277:G801–G809. doi: 10.1152/ajpgi.1999.277.4.G801. [DOI] [PubMed] [Google Scholar]
- 30.Kockx M, Princen HM, Kooistra T. Fibrate-modulated expression of fibrinogen, plasminogen activator inhibitor-1 and apolipoprotein A-I in cultured cynomolgus monkey hepatocytes -- role of the peroxisome proliferator-activated receptor-alpha. Thromb Haemost. 1998;80:942–948. [PubMed] [Google Scholar]
- 31.Bisgaard HC, Santoni-Rugiu E, Nagy P, Thorgeirsson SS. Modulation of the plasminogen activator/plasmin system in rat liver regenerating by recruitment of oval cells. Lab Invest. 1998;78:237–246. [PubMed] [Google Scholar]
- 32.Nakamura S, Nakamura I, Ma L, Vaughan DE, Fogo AB. Plasminogen activator inhibitor-1 expression is regulated by the angiotensin type 1 receptor in vivo. Kidney Int. 2000;58:251–259. doi: 10.1046/j.1523-1755.2000.00160.x. [DOI] [PubMed] [Google Scholar]