

Abstract
Biofilms are complex communities of microorganisms in organized structures attached to surfaces. Importantly, biofilms are a major cause of bacterial infections in humans, and remain one of the most significant challenges to modern medical practice. Unfortunately, conventional therapies have shown to be inadequate in the treatment of most chronic biofilm infections based on the extraordinary innate tolerance of biofilms to antibiotics. Antagonists of quorum sensing signaling molecules have been used as means to control biofilms. QS and other cell-cell communication molecules are able to revert biofilm tolerance, prevent biofilm formation and disrupt fully developed biofilms, albeit with restricted effectiveness. Recently however, it has been demonstrated that Pseudomonas aeruginosa produces a small messenger molecule cis-2-decenoic acid (cis-DA) that shows significant promise as an effective adjunctive to antimicrobial treatment of biofilms. This molecule is responsible for induction of the native biofilm dispersion response in a range of Gram-negative and Gram-positive bacteria and in yeast, and has been shown to reverse persistence, increase microbial metabolic activity and significantly enhance the cidal effects of conventional antimicrobial agents. In this manuscript, the use of cis-2-decenoic acid as a novel agent for biofilm control is discussed. Stimulating the biofilm dispersion response as a novel antimicrobial strategy holds significant promise for enhanced treatment of infections and in the prevention of biofilm formation.
Keywords: biofilms, biofilm dispersion, persister cells, signaling molecules, bacterial physiology, cis-2-decenoic acid
1. Introduction
Biofilms are dynamic communities of closely associated microbial cells embedded within a hydrated extracellular polymeric substance (EPS) on an air-liquid interface, or adherent to inert (abiotic) or living surfaces and constitute the major proportion of bacterial biomass in nature [1]. The EPS of biofilm communities is composed of any combination of polysaccharides, nucleic acids, proteins, lipids, and under certain conditions, humic substances [2,3,4,5,6]. Biofilms are formed in a sequential manner [7,8,9,10], initiated by the reversible attachment of free-floating cells to a surface [11]. A transition to irreversible attachment is mediated by a variety of direct interactions with the substratum and is generally associated with the onset of the production of EPS which entraps bacteria and results in cell-cell aggregation and the formation of microcolonies [12,13,14,15,16]. As microcolonies grow, their resident bacteria alter their physiological state toward a biofilm-specific phenotype, described as maturation stages I and II [7,8,9,17,18]. When biofilm microcolonies begin to experience limited transport with the bulk liquid, the bacteria within the microcolony may undergo a final switch in phenotype from a sessile to a mobile state, releasing cells through breaches formed in the microcolony wall in a process called dispersion [7,19].
As three dimensionally structured communities, biofilms encompass a variety of environmental microniches characterized by bacteria with differences in metabolic activity, growth rate, nutrient utilization, and the expression of surface molecules and virulence factors [20,21,22,23,24,25,26,27,28,29]. The behavior of bacteria within a biofilm has been shown in many cases to be governed by secreted chemical signals that are used in cell-to-cell communication. An example of such behavioral modification is demonstrated by Pseudomonas aeruginosa, which as a wild-type forms highly structured heterogeneous biofilms, but in strains lacking the ability to produce the quorum sensing (QS) inducer 3-oxododecanoyl-HSL, forms biofilms displaying a flat, densely packed and homogeneous architecture [17]. Likewise, a P. aeruginosa pqs mutant, deficient in the production of 2-heptyl-3-hydroxy-4-quinolone (Pseudomonas quinolone signal—PQS) has been observed to form thin biofilms containing reduced levels of extracellular DNA, compared to the wild-type strain [6,30,31,32]. In addition to influencing biofilm architecture, bacterial cell-to-cell signaling within a biofilm can provide cues about local environmental conditions, modulate inter-species and intra-species interactions with other individuals within the community, and allow coordinated responses to enable group-level expression of virulence and bacterial persistence. Cell-cell signaling in bacteria is accomplished through the production and detection of small diffusible signaling molecules of different chemical classes, that can be produced by Gram-negative bacteria, Gram-positive bacteria, or both Gram-positive and Gram-negative. To date, more than 50 such molecules have been identified and include: autoinducer-1 (AI-1) also known as N-acylhomoserine lactones (N-AHL), autoinducer-2 (AI-2) a furanosyl borate, PQS, oligopeptides (5–10 amino acid cyclic thiolactone) known as autoinducer peptides (AIP), and short-chain fatty acids which are typically unsaturated at the number 2 carbon in a cis configuration [33,34,35,36,37]. Secondary messengers provide a further level of regulation and include c-di-GMP, known to regulate biofilm development, with high intercellular levels of this molecule inducing the production of adhesins and EPS, and low levels having the opposite effect [38,39,40]. In P. aeruginosa, biofilm formation has been linked to elevated c-di-GMP levels, while dispersion is coincident with significantly reduced c-di-GMP [38,41].
One of the hallmark characteristics of biofilms is their profound tolerance to antimicrobial agents [42,43,44], which results in persistent infections and renders biofilms difficult to control [42,45]. Biofilms are typically 10 to 1000 times less susceptible to antimicrobials when compared to their planktonic counterparts [22,46,47,48]. This resistance to killing, referred to here as “biofilm tolerance” is distinct from specific resistance conferred by mutation of an antimicrobial target or the acquisition of a plasmid-borne resistance marker [46,49,50,51,52]. Instead, biofilm tolerance is observed as a generalized reduction in susceptibility to killing by essentially any antimicrobial agent. This tolerance can be overcome, in part, by significantly increasing the concentration of antimicrobial, suggesting that specific resistance is not a component of biofilm tolerance, and that cells derived from biofilms do not exhibit specific tolerance. Furthermore, biofilm tolerance may be overcome by inducing biofilm bacteria to mount a dispersion response, resulting in the evacuation of bacterial cells from the biofilm; a behavior that re-establishes susceptibility of the participating bacteria to antimicrobial agents [53,54,55].
Biofilm tolerance arises from a combination of factors that are associated with the unique environment bacteria experience within the confines of a biofilm, including: failure of the antimicrobial to penetrate throughout the biofilm matrix [21,56,57,58], reduced growth rates by the resident bacteria, a wide range of non-specific protective adaptations (such as enhanced efflux) associated with the biofilm phenotype [59,60,61,62,63], and the formation of a persister cell sub-population [64,65,66,67]. Biofilm tolerance further extends to resistance to killing by the immune system, with bacteria deriving protection against immune cells by being embedded within the mesh-like network of EPS [68].
Considering the multifactorial nature of biofilm resistance, it is apparent that strategies to eradicate biofilms have to address all the above mechanisms simultaneously [21]. While there are numerous ongoing efforts to address the biofilm problem, the most promising strategies include those aimed at manipulating the mode of growth, by either preventing biofilms from forming, or by disrupting existing biofilms. This is further supported by the finding that dispersion, the evacuation of bacterial cells from the biofilm, coincides with the reinstatement of antibiotic sensitivity [53,54,55]; suggesting that the potential to overcome the biofilm resistance mechanisms or to induce the transition of biofilm bacteria from a resistant to a susceptible phenotype would likely result in enhanced treatment options in fighting biofilm infections.
In this review, we will discuss a novel strategy to enhance the susceptibility of biofilm cells by subverting the normal biological processes that maintain biofilm integrity and promote biofilm tolerance. This strategy takes advantage of the regulatory networks that are controlled by inter and intra-species signaling and will focus primarily upon the cross-kingdom signaling molecule cis-2-decenoic acid, a fatty acid signal that has been shown to prevent biofilm development, induce biofilm dispersion, cause global changes in cellular phenotype, modulate bacterial virulence and override microbial persistence.
2. Jamming Bacterial Communication
Intercellular signaling regulates functions contributing to persistence and virulence of many bacterial pathogens. Thus, signaling interference holds significant promise as the basis of novel therapeutic strategies to improve the outcome of bacterial, and in particular biofilm infections [69,70,71]. QS plays a vital role in the regulation of virulence and biofilm-related behaviors both in the natural environment and in persistent infections [72,73], and is currently considered to be the main target as means of control of biofilm infections in the lung infections of cystic fibrosis patients, as well as other infections. Disruption of QS can be achieved by reducing the N-acylhomoserine lactone (N-AHL) synthase activity, inhibiting the production of the QS molecules, degrading N-AHLs (the most used), and using synthetic compounds that mimic QS inducer molecules [33]. Below, we provide several examples of quorum quenching, where interference with QS signalling occurs, leading to the arrest of biofilm development, disaggregation of established biofilms, and reduction of virulence.
P. aeruginosa, is an opportunistic microorganism found in a variety of niches and able to cause acute and chronic infections such as those found in the lungs of cystic fibrosis patients [74,75,76,77]. It is thus, a principal model microorganism used in the study of biofilms and biofilm cell behavior, has one of the most extensively investigated QS systems, and has been found to have approximately 10% of its genome dedicated to signalling [78]. P. aeruginosa contains one of the most complex QS systems known in bacteria, where an interconnected signaling cascade coordinates virulence, persistence, the transition to sessile growth, and biofilm development [79]. Three separate and inter-related QS systems are used by P. aeruginosa. These systems are regulated in a hierarchical manner with LasI/LasR positively regulating the RhlI/RhlR system, while the PQS system is regulated by both the LasI/LasR and RhlI/RhlR systems [30,73,80]. Considering that establishment of infections by P. aeruginosa is dependent on the LasI/LasR system, with LasR being required for QS signal binding and activation of the regulatory cascade, it is not surprising that the identification of LasR inhibitors has become a major focus in P. aeruginosa research [34]. The LasR inhibitors comprise N-AHL signaling molecules that have been chemically altered with respect to the acyl side chain, and the lactone ring [34]. In addition to inhibiting LasR, some of these molecules have been found to be potent inhibitors of RhlR [81,82]. For instance, meta-bromothiolactone was shown to inhibit pyocyanin production (a virulence factor) and biofilm formation, and to protect Caenorhabditis elegans and human lung epithelial cells from killing by P. aeruginosa [83]. Additional N-AHL inhibitors comprise various naturally occurring products such as furanones and patulin and their derivatives [83,84,85,86,87,88], with synthetic furanones having been demonstrated to significantly attenuate P. aeruginosa lung infections [83].
Modulators of the secondary messenger molecule c-di-GMP have also previously been described. The molecules LP 3134, LP 3145, LP 4010 and LP 1062 were found to inhibit the diguanylate cyclases (DCG), WspR from P. aeruginosa and tDGC from Thermotoga maritime; both required for c-di-GMP synthesis [89]. The inactivation of these DGCs resulted in biofilm inhibition of P. aeruginosa and Acinetobacter baumannii [89]. In addition, the compounds ebselen and ebselen oxide were also found to inhibit c-di-GMP by covalently modifying cysteine residues and inhibiting the binding of c-di-GMP to particular receptors, ultimately regulating biofilm development [90].
Most of these QS inhibitors are naturally occurring and show promise as therapeutic agents due to their cross species activity. For example, the QS modulators curcumin and its derivatives have been shown to reduce biofilm growth of P. aeruginosa, Escherichia coli and Vibrio harveyi, while also reducing pathogenicity and the accumulation of microorganisms on wet surfaces (biofouling) [85]. A combination of quorum quenching metabolites (inhibitors of QS) with antimicrobials has been demonstrated to enhance the susceptibility of P. aeruginosa and other microorganisms [86,91]. In Staphylococcus aureus, a Gram-positive organism, an RNAIII-inhibiting peptide suppressed the staphylococcal TRAP/agr two-component regulatory systems [involved in biofilm formation and virulence), reduced biofilm formation in vivo, and prevented infections by methicillin-resistant S. aureus (MRSA) in rats [92]. Several anti-QS peptides have also led to the inhibition of biofilm formation in Gram-positive oral bacteria [93].
These examples underscore the importance of QS quenching in the control of biofilm infections. However, molecules involved in QS have different chemical compositions, different targets, and vary significantly from species to species, and to date, no universal molecule that is able to quench (or inhibit) all different QS systems has been found. While QS quenching shows promise as a strategy for combatting biofilms [94], this approach is limited due to the complex interactions between the signals of the various species, and the various mechanisms that control QS sensing [95].
3. Fatty Acid Signaling Systems
Fatty acid signals comprise a growing group of recently identified structurally related inducer molecules, which regulate a wide range of cellular functions. These signals have been identified in a range of Gram-positive and Gram-negative bacteria, as well as in the yeast Candida albicans (Table 1), and are now recognized as a unique chemical class of QS signals [96]. To readily distinguish these molecules from one another, a new nomenclature has been proposed where the methyl (Me) substitution, if present, and its position are indicated first, the number of carbons in the fatty acyl side-chain is indicated second, and the position of the double bond (Δ) is indicated last, for example: cis-11-methyl-2-dodecenoic acid is named 11-Me-C12:Δ2, where the methyl group is on carbon 11, in a 12 carbon fatty acid, with a double bond in the cis configuration between the number 2 and number 3 carbons (Table 1) [97].
Table 1.
Fatty acid signaling molecules with known functions in the various microorganisms.
| Compound | New Nomenclature | Structure | Bacterial Species | Function | Reference |
|---|---|---|---|---|---|
| cis-11-Methyl-2-dodecenoic acid (DSF) | 11-Me-C12:Δ2 | 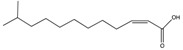 |
Xanthomonas campestris, Xanthomonas oryzae, Stenotrophomonas maltophilia, Burkholderia multivorans | Virulence, biofilm formation, floc disaggregation, microcolony formation, tolerance to antibiotics, detoxification, hyphal growth inhibition | [96,97,98,99,104,105] |
| cis-2-Dodecenoic acid (BDSF) | C12:Δ2 | 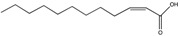 |
Burkholderia cenocepacia, Burkholderia lata Burkholderia stabilis Burkholderia vietnamiensis Burkholderia dolorosa Burkholderia ambifaria Burkholderia anthina Burkholderia pyrrocinia B. multivorans, X. oryzae | Virulence, hyphal growth inhibition | [97,104,106,107] |
| cis-2-Decenoic acid (cis-DA) | C10:Δ2 | 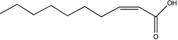 |
Pseudomonas aeruginosa | Biofilm formation, biofilm dispersion, persister cell formation, persister cell awakening, tolerance to antimicrobials. | [19,65,108,109,110] |
| cis-2-Tetradecenoic acid | C14:Δ2 | 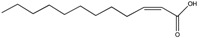 |
Xylella fastidiosa | Virulence and aggregation | [100] |
| trans-2-Decenoic acid (SDSF) | C10:Δ2t | 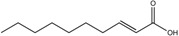 |
Streptococcus mutans | Hyphal growth inhibition | [111] |
| cis-11-Methyldodeca-2,5-dienoic acid (CDSF) | 11-Me-C12:Δ2,5 | 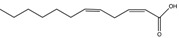 |
B. multivorans, B. stabilis B. anthina, B. pyrrocinia, X. oryzae | Hyphal growth inhibition | [97,104] |
| 10-Methyldodecanoic acid | 10-Me-C12 | 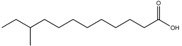 |
S. maltophilia | Stress tolerance and antibiotic tolerance | [105] |
| 11-Methyldodecanoic acid | 11-Me-C12 | 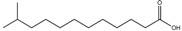 |
S. maltophilia | Stress tolerance and antibiotic tolerance | [105] |
| 12-Methyltetradecanoic acid | 12-Me-C14 | 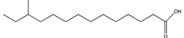 |
Xylella fastidiosa | Virulence, biofilm formation, motility | [112,113] |
| 3-Hydroxypalmitic acid | 3OH-PAME | 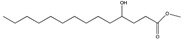 |
Ralstonia solanacearum | Virulence | [114] |
| Farnesoic acid | 3,7,11-Me-C12:Δ2t,6t,1°t | 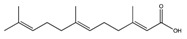 |
Candida albicans | Inhibition of germ tube formation | [115] |
Akin to N-AHLs and other cell-cell signaling molecules, fatty acid signals are involved in intra-species, inter-species, and cross-kingdom communication where they regulate bacterial growth, virulence, motility, polymer production, biofilm development, biofilm dispersion and persistence. The fatty acid signal 11-Me-C12:Δ2 in Xanthomonas campestris was the first member of this sub-class of cis-2-unsaturated fatty acids to be characterized and was named DSF for diffusible signal factor [98,99]. Interspecies signaling using fatty acids has been shown to occur between members of the Burkholderia cepacia complex [97], and between the members of Xanthomonads [96]. The Xyllela fastidiosa signal C14:Δ2, and the Burkholderia cenocepacia signals C12:Δ2 and 11-Me-C12:Δ2,5 are functional homologues of 11-Me-C12:Δ2 and both regulate biofilm formation and production of virulence factors [97,100]. Additionally, both signaling molecules contribute to the regulation of virulence factors, in response to elevated concentrations of 11-Me-C12:Δ2 produced by Xanthomonas campestris, Stenotrophomonas maltophilia and Xylella sp., leading to the disaggregation of flocs (clumps) present in planktonic cultures, acriflavin resistance and detoxification [101,102,103].
In S. maltophilia, 11-Me-C12:Δ2 production has been linked to changes in the lipopolysaccharide composition of LPS, increased protease synthesis, enhanced cell aggregation, biofilm microcolony formation and increased tolerance to antibiotics and heavy metals [116]. However, fatty acid signals do not only increase aggregative behavior or biofilm formation capability. For instance, 12-Me-C14 which was originally isolated from Xylella fastidiosa, has been found to abolish swarming motility and to reduce biofilm formation by X. fastidiosa and in P. aeruginosa [117]. The various fatty acids produced by S. maltophilia (Table 1) have also been demonstrated to have interspecies activity, and have been found to increase the stress tolerance and resistance of P. aeruginosa to antimicrobial peptides [118]. Production of fatty acid signals is not limited to bacteria, but is also found within the fungi where C. albicans has been shown to produce farnesoic acid which regulates the morphological transition from a yeast-form to a hyphal-form by inhibiting germ tube formation [96,115]. In addition to responding to its own fatty acid signal, C. albicans has been shown to respond to a variety of signals from other organisms, including C12:Δ2 from B. cenocepacia, C10:Δ2t from S. mutans, 11-Me-C12:Δ2 from X. campestris and 11-Me-C12:Δ2,5 from Burkholderia multivorans, demonstrating the cross-kingdom capabilities of fatty acid signals [96,97,106,111].
4. The Fatty Acid System in P. aeruginosa and Its Role in Biofilm Control
The fatty acid signal cis-2-decenoic acid (C10:Δ2), which will be referred to subsequently as cis-DA, was recently identified in P. aeruginosa and found to act as a biofilm dispersion autoinducer [19]. The cis-DA signaling system has been shown to regulate 666 genes encoding proteins involved in motility, chemotaxis, cell attachment, TCA cycle, EPS and LPS synthesis and secretion, virulence, iron uptake, and respiration [119]. Current findings have shown the dspI gene (PA14_54640), a PA0745 ortholog, to be required for the production of cis-DA [120]. Mutation of dspI in P. aeruginosa leads to a reduction of pyoverdine (a virulence factor) production, and to defective swarming motility [121]. However, the complete signaling transduction mechanism of the cis-DA system has yet to be fully elucidated. Most importantly and unlike other fatty acids and other signaling systems, cis-DA has been demonstrated to control biofilms by three different mechanisms: by impairing biofilm formation, by inducing biofilm dispersion [19], and by modulating the persister cell sub-population [65]. Additionally, cis-DA shows interspecies activity and also induces a dispersion response similar to that observed by P. aeruginosa in biofilms formed by C. albicans and various Gram-negative and Gram-positive bacteria, including other Pseudomonads, E. coli, Streptococcus pyogenes, S. aureus, Klebsiella pneumoniae, Proteus mirabilis and Bacillus subtilis [19]. Furthermore, cis-DA has been shown to induce biofilm dispersion in Propionibacterium acnes [122], Actimomyces naeslundii, Lactobacillus casei and Streptococcus mutans [123], when grown as single or mixed species biofilms. To date no other fatty acid has been shown to induce biofilm dispersion. However, 11-Me-C12:Δ2 has been demonstrated to disaggregate bacterial flocs of X. campestris [101].
4.1. Dispersion Induced by cis-2-Decenoic Acid as Means of Biofilm Control
Dispersion is one of the only bacterial behaviors known to be unique to biofilms, and is modulated by cell-to-cell communication. Biofilm dispersion is characterized by the release of bacteria from established biofilms, presumably to escape overcrowding or degraded environmental conditions. Autoinduction of dispersion occurs when microcolonies within a biofilm have attained a critical maximum size, and provides a mechanism by which bacteria can evacuate a microcolony, enter the bulk liquid environment, and disseminate to new sites for attachment [19]. The biofilm-specific nature of the dispersion response implies that drug development focusing on this behavior, could lead to an effective treatment of biofilm infections. The principal advantage to using drugs that target biofilm dispersion is that as bacteria (and fungi) transition from a biofilm to planktonic phenotype, the biofilm tolerance is reversed, enhancing susceptibility to antimicrobial agents and eliminating problems associated with antimicrobial penetration, the biofilm-specific phenotype and reduced growth rates [19,124]. Hence, exposure of biofilms to a dispersion inducer before and/or in combination with conventional antimicrobials would provide a novel mechanism for enhancing the activity of the treatments, through the disruption of existing biofilms [19,124]. Recent studies have shown that when compared to treatment with antimicrobials alone, targeting mature biofilms with antimicrobials in combination with cis-DA significantly reduces and removes the overall biofilm biomass in bacteria that are associated with biofilm infections and contamination of foods [108,109,110,125]. Exposure of pre-formed biofilms to different antimicrobials in the presence of cis-DA has been shown to lead to a reduction of the viability of MRSA S. aureus biofilm cells [109], and to significantly remove the biofilm biomass of B. subtilis, Salmonella enterica, S. aureus and E. coli from stainless steel and polystyrene surfaces, where they are known to cause food contamination [108]. In addition, co-exposure of P. aeruginosa PAO1 biofilm cells to antibiotics with cis-DA enhanced the killing efficacy of tobramycin or ciprofloxacin by greater than 1.5 Log, compared to treatment with the antibiotics alone [125]. It is notable that exposure to cis-DA alone shows no reduction in viability of bacteria [126], and has no cytotoxic effects on fibroblasts [109].
4.2. Persister Cell Control by Exposure to cis-DA
Persister cells are a subgroup (typically 0.0001%–1%) of a bacterial clonal population, consisting of dormant/semi-dormant specialised survivor cells that fail to succumb to treatment with antimicrobial agents, when the majority of the population shows susceptibility [64,65,66,127,128]. Various mechanisms have been proposed to explain persister cell formation, including down-regulation of genes involved in energy generation and cell maintenance [129,130], activation of the stringent response [131], and increased expression of toxin-antitoxin (TA) modules [64,132,133]. The growth rate distribution of cells within the differing microniches of a biofilm likely accounts for persister cell survival, with dormant or semi-dormant sub-populations existing at locations within the biofilm that are nutrient-limited, inducing a bacterial stress response [83,84].
The control of persister cells could be accomplished if this population were to be resuscitated to a metabolically active state, rendering them susceptible to challenge with antimicrobials. Exposure to metabolic stimuli such as—mannitol, glucose, fructose and pyruvate—have been shown to increase bacterial growth of S. aureus persister cells by more than 600 fold [134]. In addition to experiencing increased growth, exposure of S. aureus and E. coli persister cells to mannitol, glucose, fructose and pyruvate, also increased central metabolic activity as measured by increased respiration rates and membrane permeability [134]. This finding suggests that metabolic stimuli have the potential to resuscitate persister cells [134]. This is further supported by the finding that exposure to metabolic stimuli coincided with the eradication of E. coli persister cells by gentamicin [135]. In addition to metabolic stimuli, resuscitation of persister cells has been accomplished using QS inhibitors. The QS inhibitor (Z)-4-bromo-5-(bromomethylene)-3-methylfuran-2(5H)-one (BF8), known to inhibit AI-1 mediated QS through the inhibition of lasB expression, was shown to sensitize P. aeruginosa persister cells to ciprofloxacin and tobramycin, however the efficacy was reduced compared to exposure to metabolic stimuli has it did not result in complete cell eradication [136]. Although not used as a carbon source [65], cis-DA was found to significantly increase the number of cells recovered in agar plates [126], suggesting cis-DA to somehow trigger a transition from a dormant to an awake, likely more metabolically active susceptible state. This is supported by the finding that P. aeruginosa and E. coli persister cells obtained from planktonic and biofilm populations experience an additional decrease in viability (greater than 2 Log) to the point of eradication upon exposure to antimicrobials in combination with cis-DA compared to treatment with antimicrobials alone [65]. The effect of cis-DA is not limited to persister cells formed by Gram-negative organisms, as exposure of persister cells by the Gram-positive S. aureus to antibiotics in the presence of cis-DA also led to a significant decrease in cell viability, while antibiotics alone had no effect on the persister population [137]. In addition to increasing the metabolic status of persister cells and reverting their tolerant state, cis-DA also prevents persister cell formation. This is supported by the finding that the presence of cis-DA during persister cell isolation resulted in a 2-Log decrease in persister cell numbers in both P. aeruginosa and E. coli [65]. Thus, under the conditions tested challenge with cis-DA leads to a change in the cell’s metabolic status, resulting in a reduction of persister cell formation and a reversion of the persister cell’s tolerant state.
4.3. Prevention of Biofilm Formation by Exposure to cis-DA
It has been shown that certain fatty acid signals can act as agents to reduce, or negatively impact biofilm formation in several bacteria as demonstrated with 11-Me-C12:Δ2, 11-Me-C12:Δ2,5, and 12-Me-C14 [97,99,117]. It is therefore not surprising that cis-DA can also prevent biofilm formation, likely by keeping the cells in a constant metabolically active and/or dispersive growth state, as cis-DA also induces dispersion and increases the persister cells’ metabolic status. In a S. aureus MRSA strain, inhibition of bacterial growth was achieved in the presence of 2.94 mM of cis-DA, while as little as 734 μM cis-DA prevented biofilm formation [109]. Similarly, cis-DA prevented biofilm formation by single and dual-species biofilms of E. coli and K. pneumoniae in catheters [110], and in biofilms of P. aeruginosa PAO1 grown in flow cell reactors [19], when used at concentrations of 310 nM and 2.5 nM respectively. Continuous exposure of P. aeruginosa PAO1 expressing green fluorescent protein (GFP), in flow cell reactors to Electric Power Research Institute (EPRI) medium supplemented with 1 nM, 1 μM or 1 mM of cis-DA (Figure 1B–D, Table 2) under the conditions described by Davies and Marques [19], resulted in a significant reduction of maximum biofilm thickness (2-fold), average biofilm thickness (2.2-, 1.4- and 8-fold lower, respectively), roughness coefficient (1.5-fold lower, 6.4-fold lower and no-fold change, respectively), and total biomass (2.4-, 1.7- and 8.5-fold lower respectively), when compared to the absence of cis-DA (Figure 1, Table 2). Interestingly, cis-DA effectiveness does not increase with increasing concentration. Challenge of bacteria with 1 μM cis-DA (Figure 1C) actually resulted in higher biomass and increased microcolony average thickness than 1 nM cis-DA (Figure 1B), demonstrating that the effects of this fatty acid are due to its activity as a signaling molecule and not due to direct effects of the fatty acid as a detergent. However, increasing the cis-DA concentration to the mM range (Figure 1D), its effectiveness is once again present, indicating that at high concentrations cis-DA might have a detergent effect.
Figure 1.

Biofilms of P. aeruginosa PA01 harboring the green fluorescence protein (GFP) expressing vector pMRP9-1 were grown in flow cell reactors in EPRI media, in the continuous presence of different concentrations of cis-2-decenoic acid (cis-DA). (A) Control (media); (B) 1 nM of cis-DA; (C) 1 μM of cis-DA; (D) 1 mM cis-DA. Following 96 h of culture, biofilms were observed using a confocal microscope at a 400× magnification.
Table 2.
COMSTAT analysis of P. aeruginosa PAO1 harboring the green fluorescence protein expressing vector pMRP9-1, grown in flow cell reactors in EPRI media, in the continuous presence of different concentrations of cis-DA. Results are the average of at least 4 replicates.
| Biofilm Structure Quantification | Control | cis-DA 1 nM | cis-DA 1 μM | cis-DA 1 mM |
|---|---|---|---|---|
| Average ± SD | Average ± SD | Average ± SD | Average ± SD | |
| Total biomass (μm3/μm2) | 20.66 ± 7.02 | 8.49 ± 2.38 | 12.29 ± 3.5 | 2.42 ± 0.86 |
| Average thickness (μm) | 26.46 ± 14.99 | 12.21 ± 3.73 | 19.37 ± 1.92 | 3.31 ± 1.19 |
| Maximum thickness (μm) | 146.13 ± 27.69 | 61.38 ± 13.22 | 50.80 ± 6.35 | 59.27 ± 15.98 |
| Roughness coefficient (dimensionless, range: zero-infinity) | 1.40 ± 0.38 | 0.90 ± 0.27 | 0.22 ± 0.05 | 1.86 ± 0.07 |
5. Conclusions
During the past 15 years, efforts to develop new and effective anti-biofilm chemotherapeutics has focused to a large degree on blocking the mechanism of signal transduction involved in cell-to-cell communication via quorum sensing, and has led to the development of several QS antagonists. The potential chemotherapeutic use of cis-DA differs significantly from existing anti-biofilm strategies, all of which exert a negative effect upon biofilm bacteria and are likely to be both limited in the spectrum of bacteria they act upon and likely to lead to some form of resistance. The biofilm dispersion response is undoubtedly necessary for the long-term survival of bacterial biofilm populations/communities. Thus, the development of resistance to autoinduction of biofilm dispersion is not likely to be an advantage to a bacterial population. This approach to infection management, therefore, presents a new direction in microbial control. One of the key advantages to the potential use of cis-DA in therapeutic contexts is the multiple desirable effects it induces in bacteria, inhibiting biofilm development, altering virulence, inducing the disaggregation of the biofilm by inducing a dispersion response, and reverting persister cells to a susceptible state. Studies with this signal have also shown that cis-DA has no bactericidal activity nor does it induce cytotoxic effects in fibroblasts. Instead, cis-DA increases cell recovery on agar plates while reducing the number of persister cells formed in a population. Using cis-DA as an adjunctive to antimicrobial agents enhances killing of biofilm bacteria and persister cells, in some instances to the point of eradication. This effect is achieved by overcoming the mechanisms that lead to biofilm tolerance including: penetration failure, reduced metabolic rates and slow growth, biofilm-specific resistance (such as is mediated by the activation of efflux pumps), and the existence of a persister cell sub-population. Considering the urgent need to find new therapies to effectively treat chronic and unresolved infections, cis-DA provides a promising new potential chemotherapeutic to treat biofilm infections. Further research on this novel cell-to-cell signal is likely to elucidate the mechanism of signal transduction and reversion of persistence in cells challenged with cis-DA, as well as, reveal other molecules with similar activity and extend the list of microorganisms that use cell-to-cell signaling to induce biofilm dispersion.
Acknowledgments
The authors would like to acknowledge NIH grant 1 R15 AI094485-01 for the funding support and all the students that worked in their laboratories throughout the years.
Author Contributions
C.N.H.M., D.G.D. and K.S. wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- 1.De Kievit T.R., Gillis R., Marx S., Brown C., Iglewski B.H., Kievit T.R.D.E. Quorum-sensing genes in Pseudomonas aeruginosa biofilms: Their role and expression patterns. Appl. Environ. Microbiol. 2001;67:1865–1873. doi: 10.1128/AEM.67.4.1865-1873.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Costerton J.W. Introduction to biofilm. Int. J. Antimicrob. Agents. 1999;11:217–219. doi: 10.1016/S0924-8579(99)00018-7. [DOI] [PubMed] [Google Scholar]
- 3.Donlan R.M. Biofilms: Microbial life on surfaces. Emerg. Infect. Dis. 2002;8:881–890. doi: 10.3201/eid0809.020063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Wimpenny J.W. An overview of biofilms as functional communities. In: Allison Gilbert P., Lappin-Scott H., Wilson M.D., editors. Community Structure and Cooperation in Biofilms. Cambridge University Press; Cambridge, UK: 2000. pp. 1–24. [Google Scholar]
- 5.Flemming H.-C., Neu T.R., Wozniak D.J. The EPS matrix: The “house of biofilm cells”. J. Bacteriol. 2007;189:7945–7947. doi: 10.1128/JB.00858-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Allesen-Holm M., Barken K.B., Yang L., Klausen M., Webb J.S., Kjelleberg S., Molin S., Givskov M., Tolker-Nielsen T. A characterization of DNA release in Pseudomonas aeruginosa cultures and biofilms. Mol. Microbiol. 2006;59:1114–1128. doi: 10.1111/j.1365-2958.2005.05008.x. [DOI] [PubMed] [Google Scholar]
- 7.Sauer K., Camper A., Ehrlich G., Costerton J., Davies D. Pseudomonas aeruginosa displays multiple phenotypes during development as a biofilm. J. Bacteriol. 2002;184:1140–1154. doi: 10.1128/jb.184.4.1140-1154.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Stoodley P., Sauer K., Davies D.G., Costerton J.W. Biofilms as complex differentiated communities. Annu. Rev. Microbiol. 2002;56:187–209. doi: 10.1146/annurev.micro.56.012302.160705. [DOI] [PubMed] [Google Scholar]
- 9.Allegrucci M., Hu F.Z., Shen K., Hayes J., Ehrlich G.D., Post J.C., Sauer K. Phenotypic characterization of Streptococcus pneumoniae biofilm development. J. Bacteriol. 2006;188:2325–2335. doi: 10.1128/JB.188.7.2325-2335.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.O’Toole G., Kaplan H.B., Kolter R. Biofilm formation as microbial development. Annu. Rev. Microbiol. 2000;54:49–79. doi: 10.1146/annurev.micro.54.1.49. [DOI] [PubMed] [Google Scholar]
- 11.Watnick P., Kolter R. Biofilm, city of microbes. J. Bacteriol. 2000;182:2675–2679. doi: 10.1128/JB.182.10.2675-2679.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hinsa S.M., Espinosa-Urgel M., Ramos J.L., O’Toole G.A. Transition from reversible to irreversible attachment during biofilm formation by Pseudomonas fluorescens WCS365 requires an ABC transporter and a large secreted protein. Mol. Microbiol. 2003;49:905–918. doi: 10.1046/j.1365-2958.2003.03615.x. [DOI] [PubMed] [Google Scholar]
- 13.Caiazza N.C., O’Toole G.A. SadB is required for the transition from reversible to irreversible attachment during biofilm formation by Pseudomonas aeruginosa PA14. J. Bacteriol. 2004;186:4476–4485. doi: 10.1128/JB.186.14.4476-4485.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Chiang P., Burrows L.L. Biofilm formation by hyperpiliated mutants of Pseudomonas aeruginosa. J. Bacteriol. 2003;185:2374–2378. doi: 10.1128/JB.185.7.2374-2378.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Sutherland I.W. The biofilm matrix—An immobilized but dynamic microbial environment. Trends Microbiol. 2001;9:222–227. doi: 10.1016/S0966-842X(01)02012-1. [DOI] [PubMed] [Google Scholar]
- 16.Gómez-Suárez C., Pasma J., van der Borden A.J., Wingender J., Flemming H.-C., Busscher H.J., van der Mei H.C. Influence of extracellular polymeric substances on deposition and redeposition of Pseudomonas aeruginosa to surfaces. Microbiology. 2002;148:1161–1169. doi: 10.1099/00221287-148-4-1161. [DOI] [PubMed] [Google Scholar]
- 17.Davies D.G., Parsek M.R., Pearson J.P., Iglewski B.H., Costerton J.W., Greenberg E.P. The involvement of cell-to-cell signals in the development of a bacterial biofilm. Science. 1998;280:295–298. doi: 10.1126/science.280.5361.295. [DOI] [PubMed] [Google Scholar]
- 18.Harmsen M., Yang L., Pamp S.J., Tolker-Nielsen T. An update on Pseudomonas aeruginosa biofilm formation, tolerance, and dispersal. FEMS Immunol. Med. Microbiol. 2010;59:253–268. doi: 10.1111/j.1574-695X.2010.00690.x. [DOI] [PubMed] [Google Scholar]
- 19.Davies D.G., Marques C.N.H. A fatty acid messenger is responsible for inducing dispersion in microbial biofilms. J. Bacteriol. 2009;191:1393–1403. doi: 10.1128/JB.01214-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kinniment S.L., Wimpenny J.W. Measurements of the distribution of adenylate concentrations and adenylate energy charge across Pseudomonas aeruginosa biofilms. Appl. Environ. Microbiol. 1992;58:1629–1635. doi: 10.1128/aem.58.5.1629-1635.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Schierholz J.M., Beuth J., Konig D., Nurnberger A., Pulverer G. Antimicrobial substances and effects on sessile bacteria. Zentralblatt Bakteriol. 1999;289:165–177. doi: 10.1016/S0934-8840(99)80101-7. [DOI] [PubMed] [Google Scholar]
- 22.Gilbert P., Collier P.J., Brown M.R. Influence of growth rate on susceptibility to antimicrobial agents: Biofilms, cell cycle, dormancy, and stringent response. Antimicrob. Agents Chemother. 1990;34:1865–1868. doi: 10.1128/AAC.34.10.1865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Bagge N., Hentzer M., Andersen J.B., Ciofu O., Givskov M., Høiby N. Dynamics and spatial distribution of b-lactamase expression in Pseudomonas aeruginosa biofilms. Antimicrob. Agents Chemother. 2004;48:1168–1174. doi: 10.1128/AAC.48.4.1168-1174.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lenz A.P., Williamson K.S., Pitts B., Stewart P.S., Franklin M.J. Localized gene expression in Pseudomonas aeruginosa biofilms. Appl. Environ. Microbiol. 2008;74:4463–4471. doi: 10.1128/AEM.00710-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Vuong C., Whitney A.R., Deleo F.R. Polysaccharide intercellular adhesin (PIA) protects Staphylococcus epidermidis against major components of the human innate immune system. Cell. Microbiol. 2004;6:269–275. doi: 10.1046/j.1462-5822.2004.00367.x. [DOI] [PubMed] [Google Scholar]
- 26.Pearson M.M., Laurence C.A., Guinn S.E., Hansen E.J. Biofilm formation by Moraxella catarrhalis in vitro: Roles of the UspA1 adhesin and the Hag hemagglutinin. Infect. Immun. 2006;74:1588–1596. doi: 10.1128/IAI.74.3.1588-1596.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Jurcisek J.A., Bakaletz L.O. Biofilms formed by nontypeable Haemophilus influenzae in vivo contain both double-stranded DNA and type IV pilin protein. J. Bacteriol. 2007;189:3868–3875. doi: 10.1128/JB.01935-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Zhang L., Fritsch M., Hammond L., Landreville R., Slatculescu C., Colavita A., Mah T.-F. Identification of genes involved in Pseudomonas aeruginosa biofilm-specific resistance to antibiotics. PLoS ONE. 2013;8:e61625. doi: 10.1371/journal.pone.0061625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Zhang L., Mah T.-F. Involvement of a novel efflux system in biofilm-specific resistance to antibiotics. J. Bacteriol. 2008;190:4447–4452. doi: 10.1128/JB.01655-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Diggle S.P., Winzer K., Chhabra S.R., Worrall K.E., Camara M., Williams P. The Pseudomonas aeruginosa quinolone signal molecule overcomes the cell density-dependency of the quorum sensing hierarchy, regulates rhl-dependent genes at the onset of stationary phase and can be produced in the absence of LasR. Mol. Microbiol. 2003;50:29–43. doi: 10.1046/j.1365-2958.2003.03672.x. [DOI] [PubMed] [Google Scholar]
- 31.Yang L., Barken K.B., Skindersoe M.E., Christensen A.B., Givskov M., Tolker-Nielsen T. Effects of iron on DNA release and biofilm development by Pseudomonas aeruginosa. Microbiology. 2007;153:1318–1328. doi: 10.1099/mic.0.2006/004911-0. [DOI] [PubMed] [Google Scholar]
- 32.Yang L., Nilsson M., Gjermansen M., Givskov M., Tolker-Nielsen T. Pyoverdine and PQS mediated subpopulation interactions involved in Pseudomonas aeruginosa biofilm formation. Mol. Microbiol. 2009;74:1380–1392. doi: 10.1111/j.1365-2958.2009.06934.x. [DOI] [PubMed] [Google Scholar]
- 33.Kalia V.C. Quorum sensing inhibitors: An overview. Biotechnol. Adv. 2013;31:224–245. doi: 10.1016/j.biotechadv.2012.10.004. [DOI] [PubMed] [Google Scholar]
- 34.Rutherford S.T., Bassler B.L. Bacterial quorum sensing: Its role in virulence and possibilities for its control. Cold Spring Harb. Perspect. Med. 2012;2:1–26. doi: 10.1101/cshperspect.a012427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Bassler B.L. How bacteria talk to each other: Regulation of gene expression by quorum sensing. Curr. Opin. Microbiol. 1999;2:582–587. doi: 10.1016/S1369-5274(99)00025-9. [DOI] [PubMed] [Google Scholar]
- 36.Parsek M.R., Greenberg E.P. Sociomicrobiology: The connections between quorum sensing and biofilms. Trends Microbiol. 2005;13:27–33. doi: 10.1016/j.tim.2004.11.007. [DOI] [PubMed] [Google Scholar]
- 37.Ryan R.P., Dow J.M. Communication with a growing family: Diffusible signal factor (DSF) signaling in bacteria. Trends Microbiol. 2011;19:145–152. doi: 10.1016/j.tim.2010.12.003. [DOI] [PubMed] [Google Scholar]
- 38.Hengge R. Principles of c-di-GMP signalling in bacteria. Nat. Rev. Microbiol. 2009;7:263–273. doi: 10.1038/nrmicro2109. [DOI] [PubMed] [Google Scholar]
- 39.Romling U., Amikam D. Cyclic di-GMP as a second messenger. Curr. Opin. Microbiol. 2006;9:218–228. doi: 10.1016/j.mib.2006.02.010. [DOI] [PubMed] [Google Scholar]
- 40.Ryan R.P., Fouhy Y., Lucey J.F., Dow J.M. Cyclic di-GMP signaling in bacteria: Recent advances and new puzzles. J. Bacteriol. 2006;188:8327–8334. doi: 10.1128/JB.01079-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Simm R., Morr M., Kader A., Nimtz M., Romling U. GGDEF and EAL domains inversely regulate cyclic di-GMP levels and transition from sessility to motility. Mol. Microbiol. 2004;53:1123–1134. doi: 10.1111/j.1365-2958.2004.04206.x. [DOI] [PubMed] [Google Scholar]
- 42.Costerton J.W., Stewart P.S., Greenberg E.P. Bacterial biofilms: A common cause of persistent infections. Science. 1999;284:1318–1322. doi: 10.1126/science.284.5418.1318. [DOI] [PubMed] [Google Scholar]
- 43.Lewis K. Multidrug tolerance of biofilms and persister cells. Curr. Top. Microbiol. Immunol. 2008;322:107–131. doi: 10.1007/978-3-540-75418-3_6. [DOI] [PubMed] [Google Scholar]
- 44.Spoering A., Lewis K. Biofilms and planktonic cells of Pseudomonas aeruginosa have similar resistance to killing by antimicrobials. J. Bacteriol. 2001;183:6746–6751. doi: 10.1128/JB.183.23.6746-6751.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.O’Toole G.A. Microbiology: A resistance switch. Nature. 2002;416:695–696. doi: 10.1038/416695a. [DOI] [PubMed] [Google Scholar]
- 46.Gilbert P., Maira-Litran T., McBain A.J., Rickard A.H., Whyte F.W. The physiology and collective recalcitrance of microbial biofilm communities. Adv. Microb. Physiol. 2002;46:202–256. [PubMed] [Google Scholar]
- 47.Prosser B.L., Taylor D., Dix B.A., Cleeland R. Method of evaluating effects of antibiotics on bacterial biofilm. Antimicrob. Agents Chemother. 1987;31:1502–1506. doi: 10.1128/AAC.31.10.1502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Gristina A.G., Hobgood C.D., Webb L.X., Myrvik Q.N. Adhesive colonization of biomaterials and antibiotic resistance. Biomaterials. 1987;8:423–426. doi: 10.1016/0142-9612(87)90077-9. [DOI] [PubMed] [Google Scholar]
- 49.Donlan R.M., Costerton J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002;15:167–193. doi: 10.1128/CMR.15.2.167-193.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Lewis K. Riddle of biofilm resistance. Antimicrob. Agents Chemother. 2001;45:999–1007. doi: 10.1128/AAC.45.4.999-1007.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Mah T.F., O’Toole G.A. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001;9:34–39. doi: 10.1016/S0966-842X(00)01913-2. [DOI] [PubMed] [Google Scholar]
- 52.Stewart P.S., William Costerton J. Antibiotic resistance of bacteria in biofilms. Lancet. 2001;358:135–138. doi: 10.1016/S0140-6736(01)05321-1. [DOI] [PubMed] [Google Scholar]
- 53.Stewart P.S. Mechanisms of antibiotic resistance in bacterial biofilms. Int. J. Med. Microbiol. 2002;292:107–113. doi: 10.1078/1438-4221-00196. [DOI] [PubMed] [Google Scholar]
- 54.Gupta K., Marques C.N.H., Petrova O.E., Sauer K. Antimicrobial tolerance of Pseudomonas aeruginosa biofilms is activated during an early developmental stage and requires the two-component hybrid SagS. J. Bacteriol. 2013;195:4975–4987. doi: 10.1128/JB.00732-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Davies D. Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2003;2:114–122. doi: 10.1038/nrd1008. [DOI] [PubMed] [Google Scholar]
- 56.Yassien M., Khardori N., Ahmedy A., Toama M. Modulation of biofilms of Pseudomonas aeruginosa by quinolones. Antimicrob. Agents Chemother. 1995;39:2262–2268. doi: 10.1128/AAC.39.10.2262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Stewart P.S. Theoretical aspects of antibiotic diffusion into microbial biofilms. Antimicrob. Agents Chemother. 1996;40:2517–2522. doi: 10.1128/aac.40.11.2517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Allison D.G. The biofilm matrix. Biofouling. 2003;19:139–150. doi: 10.1080/0892701031000072190. [DOI] [PubMed] [Google Scholar]
- 59.Liao J., Sauer K. The MerR-like transcriptional regulator BrlR contributes to Pseudomonas aeruginosa biofilm tolerance. J. Bacteriol. 2012;194:4823–4836. doi: 10.1128/JB.00765-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Newberry K.J., Brennan R.G. The structural mechanism for transcription activation by MerR family member multidrug transporter activation, N terminus. J. Biol. Chem. 2004;279:20356–20362. doi: 10.1074/jbc.M400960200. [DOI] [PubMed] [Google Scholar]
- 61.Heldwein E.E., Brennan R.G. Crystal structure of the transcription activator BmrR bound to DNA and a drug. Nature. 2001;409:378–382. doi: 10.1038/35053138. [DOI] [PubMed] [Google Scholar]
- 62.Zheleznova E.E., Markham P.N., Neyfakh A.A., Brennan R.G. Structural basis of multidrug recognition by BmrR, a transcription activator of a multidrug transporter. Cell. 1999;96:353–362. doi: 10.1016/S0092-8674(00)80548-6. [DOI] [PubMed] [Google Scholar]
- 63.Drenkard E., Ausubel F.M. Pseudomonas biofilm formation and antibiotic resistance are linked to phenotypic variation. Nature. 2002;416:740–743. doi: 10.1038/416740a. [DOI] [PubMed] [Google Scholar]
- 64.Keren I., Kaldalu N., Spoering A., Wang Y., Lewis K. Persister cells and tolerance to antimicrobials. FEMS Microbiol. Lett. 2004;230:13–18. doi: 10.1016/S0378-1097(03)00856-5. [DOI] [PubMed] [Google Scholar]
- 65.Marques C.N.H., Morozov A., Planzos P., Zelaya H.M. The fatty acid signaling molecule cis-2-decenoic acid increases metabolic activity and reverts persister cells to an antimicrobial susceptible state. Appl. Environ. Microbiol. 2014;80:6976–6991. doi: 10.1128/AEM.01576-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Keren I., Shah D., Spoering A. Specialized persister cells and the mechanism of multidrug tolerance in Escherichia coli. J. Bacteriol. 2004;186:8172–8180. doi: 10.1128/JB.186.24.8172-8180.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Sufya N., Allison D.G., Gilbert P. Clonal variation in maximum specific growth rate and susceptibility towards antimicrobials. J. Appl. Microbiol. 2003;95:1261–1267. doi: 10.1046/j.1365-2672.2003.02079.x. [DOI] [PubMed] [Google Scholar]
- 68.Hoyle B.D., Jass J., Costerton J.W. The biofilm glycocalyx as a resistance factor. J. Antimicrob. Chemother. 1990;26:1–5. doi: 10.1093/jac/26.1.1. [DOI] [PubMed] [Google Scholar]
- 69.McCarthy Y., Yang L., Twomey K.B., Sass A., Tolker-Nielsen T., Mahenthiralingam E., Dow J.M., Ryan R.P. A sensor kinase recognizing the cell-cell signal BDSF (cis-2-dodecenoic acid) regulates virulence in Burkholderia cenocepacia. Mol. Microbiol. 2010;77:1220–1236. doi: 10.1111/j.1365-2958.2010.07285.x. [DOI] [PubMed] [Google Scholar]
- 70.Rasko D.A., Sperandio V. Novel approaches to bacterial infection therapy by interfering with cell-to-cell signaling. Curr. Protoc. Microbiol. 2009;12 doi: 10.1002/9780471729259.mc1703s12. [DOI] [PubMed] [Google Scholar]
- 71.Rasko D.A., Sperandio V. Anti-virulence strategies to combat bacteria-mediated disease. Nat. Rev. Drug Discov. 2010;9:117–128. doi: 10.1038/nrd3013. [DOI] [PubMed] [Google Scholar]
- 72.Singh P.K., Schaefer A.L., Parsek M.R., Moninger T.O., Welsh M.J., Greenberg E.P. Quorum-sensing signals indicate that cystic fibrosis lungs are infected with bacterial biofilms. Nature. 2000;407:762–764. doi: 10.1038/35037627. [DOI] [PubMed] [Google Scholar]
- 73.Chugani S.A., Whiteley M., Lee K.M., D’Argenio D., Manoil C., Greenberg E.P. QscR, a modulator of quorum-sensing signal synthesis and virulence in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA. 2001;98:2752–2757. doi: 10.1073/pnas.051624298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Green S.K., Schroth M.N., Cho J.J., Kominos S.K., Vitanza-jack V.B. Agricultural plants and soil as a reservoir for Pseudomonas aeruginosa. Appl. Microbiol. 1974;28:987–991. doi: 10.1128/am.28.6.987-991.1974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Zimakoff J., Høiby N., Rosendal K., Guilbert J.P. Epidemiology of Pseudomonas aeruginosa infection and the role of contamination of the environment in a cystic fibrosis clinic. J. Hosp. Infect. 1983;4:31–40. doi: 10.1016/0195-6701(83)90062-2. [DOI] [PubMed] [Google Scholar]
- 76.Moreau-Marquis S., Stanton B.A., O’Toole G.A. Pseudomonas aeruginosa biofilm formation in the cystic fibrosis airway. Pulm. Pharmacol. Ther. 2008;21:595–599. doi: 10.1016/j.pupt.2007.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Van Delden C., Iglewski B.H. Cell-to-cell signaling and Pseudomonas aeruginosa infections. Emerg. Infect. Dis. 1998;4:551–560. doi: 10.3201/eid0404.980405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Williams P., Cámara M. Quorum sensing and environmental adaptation in Pseudomonas aeruginosa: A tale of regulatory networks and multifunctional signal molecules. Curr. Opin. Microbiol. 2009;12:182–191. doi: 10.1016/j.mib.2009.01.005. [DOI] [PubMed] [Google Scholar]
- 79.Jimenez P.N., Koch G., Thompson J.A., Xavier K.B., Cool R.H., Quax W.J. The multiple signaling systems regulating virulence in Pseudomonas aeruginosa. Microbiol. Mol. Biol. Rev. 2012;76:46–65. doi: 10.1128/MMBR.05007-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Pearson J.P., Pesci E.C., Iglewski B.H. Roles of Pseudomonas aeruginosa las and rhl quorum-sensing systems in control of elastase and rhamnolipid biosynthesis genes. J. Bacteriol. 1997;179:5756–5767. doi: 10.1128/jb.179.18.5756-5767.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Smith R.S., Iglewski B.H. Pseudomonas aeruginosa quorum sensing as a potential antimicrobial target. J. Clin. Investig. 2003;112:1460–1465. doi: 10.1172/JCI200320364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Ishida T., Ikeda T., Takiguchi N., Kuroda A., Kato J., Ohtake H. Inhibition of quorum sensing in Pseudomonas aeruginosa by N-acyl cyclopentylamides. Appl. Environ. Microbiol. 2007;73:3183–3188. doi: 10.1128/AEM.02233-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Wu H., Song Z., Hentzer M., Andersen J.B., Molin S., Givskov M., Høiby N. Synthetic furanones inhibit quorum-sensing and enhance bacterial clearance in Pseudomonas aeruginosa lung infection in mice. J. Antimicrob. Chemother. 2004;53:1054–1061. doi: 10.1093/jac/dkh223. [DOI] [PubMed] [Google Scholar]
- 84.Hentzer M., Riedel K., Rasmussen T.B., Heydorn A., Andersen J.B., Parsek M.R., Rice S.A., Eberl L., Molin S., Høiby N., et al. Inhibition of quorum sensing in Pseudomonas aeruginosa biofilm bacteria by a halogenated furanone compound. Microbiology. 2002;148:87–102. doi: 10.1099/00221287-148-1-87. [DOI] [PubMed] [Google Scholar]
- 85.Bjarnsholt T., Jensen P.Ø., Rasmussen T.B., Christophersen L., Calum H., Hentzer M., Hougen H.-P., Rygaard J., Moser C., Eberl L., et al. Garlic blocks quorum sensing and promotes rapid clearing of pulmonary Pseudomonas aeruginosa infections. Microbiology. 2005;151:3873–3880. doi: 10.1099/mic.0.27955-0. [DOI] [PubMed] [Google Scholar]
- 86.Ren D., Zuo R., Barrios A.F.G., Bedzyk L.A., Eldridge G.R., Pasmore M.E., Wood T.K. Differential gene expression for investigation of Escherichia coli biofilm inhibition by plant extract ursolic acid. Appl. Environ. Microbiol. 2005;71:4022–4034. doi: 10.1128/AEM.71.7.4022-4034.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Kim C., Kim J., Park H.-Y., Park H.-J., Lee J.H., Kim C.K., Yoon J. Furanone derivatives as quorum-sensing antagonists of Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2008;80:37–47. doi: 10.1007/s00253-008-1474-6. [DOI] [PubMed] [Google Scholar]
- 88.Borlee B.R., Goldman A.D., Murakami K., Samudrala R., Wozniak D.J., Parsek M.R. Pseudomonas aeruginosa uses a cyclic-di-GMP-regulated adhesin to reinforce the biofilm extracellular matrix. Mol. Microbiol. 2010;75:827–842. doi: 10.1111/j.1365-2958.2009.06991.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Sambanthamoorthy K., Luo C., Pattabiraman N., Feng X., Koestler B., Waters C.M., Palys T.J. Identification of small molecules inhibiting diguanylate cyclases to control bacterial biofilm development. Biofouling. 2014;30:17–28. doi: 10.1080/08927014.2013.832224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Lieberman O.O.J., Orr M.M.W., Wang Y., Lee V.T. High-throughput screening using the differential radial capillary action of ligand assay identifies ebselen as an inhibitor of diguanylate cyclases. ACS Chem. Biol. 2014;9:183–192. doi: 10.1021/cb400485k. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Garo E., Eldridge G.R., Goering M.G., Pulcini E.D., Hamilton M.A., Costerton J.W., James G.A. Asiatic acid and corosolic acid enhance the susceptibility of Pseudomonas aeruginosa biofilms to tobramycin. Antimicrob. Agents Chemother. 2007;51:1813–1817. doi: 10.1128/AAC.01037-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Balaban N., Cirioni O., Giacometti A. Treatment of Staphylococcus aureus biofilm infection by the quorum-sensing inhibitor RIP. Antimicrob. Agents Chemother. 2007;51:2226–2229. doi: 10.1128/AAC.01097-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.LoVetri K., Madhyastha S. Antimicrobial and antibiofilm activity of quorum sensing peptides and Peptide analogues against oral biofilm bacteria. Methods Mol. Biol. 2010;618:383–392. doi: 10.1007/978-1-60761-594-1_24. [DOI] [PubMed] [Google Scholar]
- 94.Kjelleberg S., Molin S. Is there a role for quorum sensing signals in bacterial biofilms? Curr. Opin. Microbiol. 2002;5:254–258. doi: 10.1016/S1369-5274(02)00325-9. [DOI] [PubMed] [Google Scholar]
- 95.Estrela A.B., Abraham W.-R. Combining Biofilm-Controlling Compounds and Antibiotics as a Promising New Way to Control Biofilm Infections. Pharmaceuticals. 2010;3:1374–1393. doi: 10.3390/ph3051374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Wang L.H., He Y., Gao Y., Wu J.E., Dong Y.H., He C., Wang S.X., Weng L.X., Xu J.L., Tay L., et al. A bacterial cell-cell communication signal with cross-kingdom structural analogues. Mol. Microbiol. 2004;51:903–912. doi: 10.1046/j.1365-2958.2003.03883.x. [DOI] [PubMed] [Google Scholar]
- 97.Deng Y., Wu J., Eberl L., Zhang L.H. Structural and functional characterization of diffusible signal factor family quorum-sensing signals produced by members of the Burkholderia cepacia complex. Appl. Environ. Microbiol. 2010;76:4675–4683. doi: 10.1128/AEM.00480-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Tang J.L., Liu Y.N., Barber C.E., Dow J.M., Wootton J.C., Daniels M.J. Genetic and molecular analysis of a cluster of rpf genes involved in positive regulation of synthesis of extracellular enzymes and polysaccharide in Xanthomonas campestris pathovar campestris. Mol. Gen. Genet. 1991;226:409–417. doi: 10.1007/BF00260653. [DOI] [PubMed] [Google Scholar]
- 99.Barber C.E., Tang J.L., Feng J.X., Pan M.Q., Wilson T.J.G., Slater H., Dow J.M., Williams P., Daniels M.J. A novel regulatory system required for pathogenicity of Xanthomonas campestris is mediated by a small diffusible signal molecule. Mol. Microbiol. 1997;24:555–566. doi: 10.1046/j.1365-2958.1997.3721736.x. [DOI] [PubMed] [Google Scholar]
- 100.Beaulieu E.D., Ionescu M., Chatterjee S., Yokota K., Trauner D., Lindow S. Characterization of a diffusible signaling factor from Xylella fastidiosa. MBio. 2013;4:9–14. doi: 10.1128/mBio.00539-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Dow J.M., Crossman L., Findlay K., He Y.Q., Feng J.X., Tang J.L. Biofilm dispersal in Xanthomonas campestris is controlled by cell-cell signaling and is required for full virulence to plants. Proc. Natl. Acad. Sci. USA. 2003;100:10995–11000. doi: 10.1073/pnas.1833360100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.He Y.W., Xu M., Lin K., Ng Y.J.A., Wen C.M., Wang L.H., Liu Z.D., Zhang H.B., Dong Y.H., Dow J.M., et al. Genome scale analysis of diffusible signal factor regulon in Xanthomonas campestris pv. campestris: Identification of novel cell-cell communication-dependent genes and functions. Mol. Microbiol. 2006;59:610–622. doi: 10.1111/j.1365-2958.2005.04961.x. [DOI] [PubMed] [Google Scholar]
- 103.Minkwitz A., Berg G. Comparison of antifungal activities and 16S ribosomal DNA sequences of clinical and environmental isolates of Stenotrophomonas maltophilia. J. Clin. Microbiol. 2001;39:139–145. doi: 10.1128/JCM.39.1.139-145.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.He Y.-W., Wu J., Cha J.-S., Zhang L.-H. Rice bacterial blight pathogen Xanthomonas oryzae pv. oryzae produces multiple DSF-family signals in regulation of virulence factor production. BMC Microbiol. 2010;10:187. doi: 10.1186/1471-2180-10-187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Huang T.-P., Lee Wong A.C. Extracellular fatty acids facilitate flagella-independent translocation by Stenotrophomonas maltophilia. Res. Microbiol. 2007;158:702–711. doi: 10.1016/j.resmic.2007.09.002. [DOI] [PubMed] [Google Scholar]
- 106.Boon C., Deng Y., Wang L.-H., He Y., Xu J.-L., Fan Y., Pan S.Q., Zhang L.-H. A novel DSF-like signal from Burkholderia cenocepacia interferes with Candida albicans morphological transition. ISME J. 2008;2:27–36. doi: 10.1038/ismej.2007.76. [DOI] [PubMed] [Google Scholar]
- 107.Ryan R.P., McCarthy Y., Watt S.A., Niehaus K., Dow J.M. Intraspecies signaling involving the diffusible signal factor BDSF (cis-2-dodecenoic acid) influences virulence in Burkholderia cenocepacia. J. Bacteriol. 2009;191:5013–5019. doi: 10.1128/JB.00473-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Sepehr S., Rahmani-Badi A., Babaie-Naiej H., Soudi M.R. Unsaturated fatty acid, cis-2-decenoic acid, in combination with disinfectants or antibiotics removes pre-established biofilms formed by food-related bacteria. PLoS ONE. 2014;9:e101677. doi: 10.1371/journal.pone.0101677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Jennings J., Courtney H., Haggard W. Cis-2-decenoic acid inhibits S. aureus growth and biofilm in vitro: A pilot study. Clin. Orthop. Relat. Res. 2012;470:2663–2670. doi: 10.1007/s11999-012-2388-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Rahmani-Badi A., Sepehr S., Mohammadi P., Soudi M.R., Babaie-Naiej H. A combination of cis-2-decenoic acid and antibiotics eradicates pre- established catheter-associated biofilms. J. Med. Microbiol. 2014;63:1509–1516. doi: 10.1099/jmm.0.075374-0. [DOI] [PubMed] [Google Scholar]
- 111.Vílchez R., Lemme A., Ballhausen B., Thiel V., Schulz S., Jansen R., Wagner-Döbler I., Sztajer H. Streptococcus mutans inhibits Candida albicans hyphal formation by the fatty acid signaling molecule trans-2-decenoic acid (SDSF) Chembiochem. 2010;11:1552–1162. doi: 10.1002/cbic.201000086. [DOI] [PubMed] [Google Scholar]
- 112.Colnaghi Simionato A.V., da Silva D.S., Lambais M.R., Carrilho E. Characterization of a putative Xylella fastidiosa diffusible signal factor by HRGC-EI-MS. J. Mass Spectrom. 2007;42:490–496. doi: 10.1002/jms.1181. [DOI] [PubMed] [Google Scholar]
- 113.Chatterjee S., Wistrom C., Lindow S.E. A cell-cell signaling sensor is required for virulence and insect transmission of Xylella fastidiosa. Proc. Natl. Acad. Sci. USA. 2008;105:2670–2675. doi: 10.1073/pnas.0712236105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Flavier A.B., Clough S.J., Schell M.A., Denny T.P. Identification of 3-hydroxypalmitic acid methyl ester as a novel autoregulator controlling virulence in Ralstonia solanacearum. Mol. Microbiol. 1997;26:251–259. doi: 10.1046/j.1365-2958.1997.5661945.x. [DOI] [PubMed] [Google Scholar]
- 115.Oh K.B., Miyazawa H., Naito T., Matsuoka H. Purification and characterization of an autoregulatory substance capable of regulating the morphological transition in Candida albicans. Proc. Natl. Acad. Sci. USA. 2001;98:4664–4668. doi: 10.1073/pnas.071404698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Fouhy Y., Scanlon K., Schouest K., Spillane C., Crossman L., Avison M.B., Ryan R.P., Dow J.M. Diffusible signal factor-dependent cell-cell signaling and virulence in the nosocomial pathogen Stenotrophomonas maltophilia. J. Bacteriol. 2007;189:4964–4968. doi: 10.1128/JB.00310-07. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 117.Inoue T., Shingaki R., Fukui K. Inhibition of swarming motility of Pseudomonas aeruginosa by branched-chain fatty acids. FEMS Microbiol. Lett. 2008;281:81–86. doi: 10.1111/j.1574-6968.2008.01089.x. [DOI] [PubMed] [Google Scholar]
- 118.Ryan R.P., Fouhy Y., Garcia B.F., Watt S.A., Niehaus K., Yang L., Tolker-Nielsen T., Dow J.M. Interspecies signalling via the Stenotrophomonas maltophilia diffusible signal factor influences biofilm formation and polymyxin tolerance in Pseudomonas aeruginosa. Mol. Microbiol. 2008;68:75–86. doi: 10.1111/j.1365-2958.2008.06132.x. [DOI] [PubMed] [Google Scholar]
- 119.Rahmani-Badi A., Sepehr S., Fallahi H., Heidari-Keshel S. Dissection of the cis-2-decenoic acid signaling network in Pseudomonas aeruginosa using microarray technique. Front. Microbiol. 2015;6:383. doi: 10.3389/fmicb.2015.00383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Amari D.T., Marques C.N.H., Davies D.G. The putative enoyl-coenzyme A hydratase DspI is required for production of the Pseudomonas aeruginosa biofilm dispersion autoinducer cis-2-decenoic acid. J. Bacteriol. 2013;195:4600–4610. doi: 10.1128/JB.00707-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Feinbaum R.L., Urbach J.M., Liberati N.T., Djonovic S., Adonizio A., Carvunis A.-R., Ausubel F.M. Genome-wide identification of Pseudomonas aeruginosa virulence-related genes using a Caenorhabditis elegans infection model. PLoS Pathog. 2012;8:e1002813. doi: 10.1371/journal.ppat.1002813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Marques C.N.H., Amari D.T., Varer M., Richards J.S., Cavender D., Davies D.G. Influence of cis-2-decenoic acid on Propionibacterium acnes biofilm development and antimicrobial susceptibility; Proceedings of the 5th ASM Conference on Biofilms; Cancun, Mexico. 15–19 November 2009; p. 67. [Google Scholar]
- 123.Marques C.N.H., Guttenplan S.B., Okkotsu Y., Davies D.G. Biofilm dispersion in oral bacterial species; Proceedings of the 109th General Meeting of the American Society for Microbiology; Philadelphia, PA, USA. 17–21 May 2009; p. 158. [Google Scholar]
- 124.Barraud N., Schleheck D., Klebensberger J., Webb J.S., Hassett D.J., Rice S.A., Kjelleberg S. Nitric oxide signaling in Pseudomonas aeruginosa biofilms mediates phosphodiesterase activity, decreased cyclic di-GMP levels, and enhanced dispersal. J. Bacteriol. 2009;191:7333–7342. doi: 10.1128/JB.00975-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Marques C.N.H., Dolacky S.D., Guttenplan S.B., Payabyab E.C., Davies D.G. Pseudomonas aeruginosa antimicrobial challenge in combination with a biofilm dispersal agent; Proceedings of the 4th ASM conference on Biofilms; Quebec, QC, Canada. 25–29 March 2007; p. 166. [Google Scholar]
- 126.Amari D.T., Marques C.N.H., Ferreira A., Silverman I., Davies D.G. Resuscitation of bacteria by exposure to the intercellular communication molecule, cis-2-decenoic acid; Proceedings of the 110th General Meeting of the American Society for Microbiology; San Diego, CA, USA. 23–27 May 2010; p. 162. [Google Scholar]
- 127.Fauvart M., de Groote V.N., Michiels J. Role of persister cells in chronic infections: Clinical relevance and perspectives on anti-persister therapies. J. Med. Microbiol. 2011;60:699–709. doi: 10.1099/jmm.0.030932-0. [DOI] [PubMed] [Google Scholar]
- 128.Mulcahy L.R., Burns J.L., Lory S., Lewis K. Emergence of Pseudomonas aeruginosa strains producing high levels of persister cells in patients with cystic fibrosis. J. Bacteriol. 2010;192:6191–6199. doi: 10.1128/JB.01651-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Balaban N.Q., Merrin J., Chait R., Kowalik L., Leibler S. Bacterial persistence as a phenotypic switch. Science. 2004;305:1622–1625. doi: 10.1126/science.1099390. [DOI] [PubMed] [Google Scholar]
- 130.Lewis K. Persister cells and the riddle of biofilm survival. Biochemistry. 2005;70:267–274. doi: 10.1007/s10541-005-0111-6. [DOI] [PubMed] [Google Scholar]
- 131.Kwan B.W., Valenta J.A., Benedik M.J., Wood T.K. Arrested protein synthesis increases persister-like cell formation. Antimicrob. Agents Chemother. 2013;57:1468–1473. doi: 10.1128/AAC.02135-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Korch S.B., Henderson T.A., Hill T.M. Characterization of the hipA7 allele of Escherichia coli and evidence that high persistence is governed by (p)ppGpp synthesis. Mol. Microbiol. 2003;50:1199–1213. doi: 10.1046/j.1365-2958.2003.03779.x. [DOI] [PubMed] [Google Scholar]
- 133.Scherrer R., Moyed H.S. Conditional impairment of cell division and altered lethality in hipA mutants of Escherichia coli K-12. J. Bacteriol. 1988;170:3321–3326. doi: 10.1128/jb.170.8.3321-3326.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Pascoe B., Dams L., Wilkinson T.S., Harris L.G., Bodger O., Mack D., Davies A.P. Dormant cells of Staphylococcus aureus are resuscitated by spent culture supernatant. PLoS ONE. 2014;9:e85998. doi: 10.1371/journal.pone.0085998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Allison K.R., Brynildsen M.P., Collins J.J. Metabolite-enabled eradication of bacterial persisters by aminoglycosides. Nature. 2011;473:216–220. doi: 10.1038/nature10069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Pan J., Bahar A.A., Syed H., Ren D. Reverting antibiotic tolerance of Pseudomonas aeruginosa PAO1 persister cells by (Z)-4-bromo-5-(bromomethylene)-3-methylfuran-2(5H)-one. PLoS ONE. 2012;7:e45778. doi: 10.1371/journal.pone.0045778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Mina E., Marques C.N.H. Effect of cis-2-decenoic acid on virulence of Staphylococcus aureus persister cells; Proceedings of the 113th General Meeting of the American Society for Microbiology; Denver, CO, USA. 18–21 May 2013; p. 121. [Google Scholar]