

Abstract
Background
Lactococcus lactis has been safely consumed in fermented foods for millennia. This Gram-positive bacterium has now become of industrial importance as an expression host for the overproduction of lipopolysaccharide-free recombinant proteins used as food ingredients, therapeutic proteins and biotechnological enzymes.
Results
This paper reports an agmatine-controlled expression (ACE) system for L. lactis, comprising the lactococcal agmatine-sensor/transcriptional activator AguR and its target promoter PaguB. The usefulness and efficiency of this system was checked via the reporter gene gfp and by producing PEP (Myxococcus xanthus prolyl-endopeptidase), an enzyme of biomedical interest able to degrade the immunotoxic peptides produced during the gastrointestinal breakdown of gluten.
Conclusion
The ACE system developed in this work was suitable for the efficient expression of the functional recombinant proteins GFP and PEP. The expression system was tightly regulated by the agmatine concentration and allowed high protein production without leakiness.
Keywords: Lactococcus lactis, Expression vector, Agmatine induction, Myxococcus xanthus, Prolyl-endopeptidase
Background
Heterologous protein production is a multi-billion dollar market of particular importance to manufacturers of biopharmaceuticals and enzymes for industrial use. Microbial production systems are often the best option for making such products given their ease of handling and high synthesis rates [1]. At present, Escherichia coli remains the first choice of host system given its high overexpression yields, ease of genetic handling, and the wealth of information available on this microorganism [2]. However, it is not without its drawbacks, such as the formation of inclusion bodies, the presence of an outer membrane that hampers secretion, its relatively complicated aerobic fermentation system, and the formation of endotoxins such as cell wall lipopolysaccharides [3]. The presence of bacterial endotoxins is one of the major concerns of regulatory agencies, and the need to add downstream steps to ensure their removal can make otherwise simple processes quite costly [4].
The Gram-positive bacterium Lactococcus lactis has emerged as an attractive alternative for the overproduction of recombinant proteins. Due to its long, safe history of use in dairy fermentations, this bacterium has been classified as a food grade microorganism ‘Generally Recognized As Safe’ (GRAS) by the US Food and Drug Administration (FDA), and has led it to receive ‘Qualified Presumption of Safety’ (QPS) status from the European Food Safety Authority (EFSA) [5]. In addition, it is an efficient secretor of extracellular recombinant proteins, has low protease activity (allowing for simplified purification processes), and a very simple metabolism that allows for rapid growth without aeration—all properties that facilitate scaling-up [4]. Moreover, L. lactis is likely to provide a good membrane environment for the overproduction of eukaryotic proteins [6]. Indeed, a number of eukaryotic membrane transporters, yeast mitochondrial proteins and human proteins have been heterologously expressed in this host [7]. Further, L. lactis is an efficient cellular factory able to turn out recombinant viral antigens, interleukins, allergens, virulence factors, bacteriocins and enzymes [8, 9]. It can even be genetically engineered to produce proteins from pathogenic species on its cell surface, and thus be used as a vector for the production and delivery of oral vaccines against HIV, cholera, malaria, human papillomavirus, stomach ulcers, tetanus and brucellosis [10–18].
For most of these applications, the availability of vectors that allow the cloning and expression of foreign genes is of paramount importance. Although the genetic accessibility and ease of handling of L. lactis lags far behind that of E. coli, the molecular biology techniques and genetic tools available for use with this bacterium have increased over recent years [9, 19]. So far, a number of inducible expression systems regulated by environmental factors have been documented, including the chloride-inducible expression cassette [20], the zinc-inducible expression system [21], the lactate-inducible P170 system [22], the heat shock-inducible system [23], systems based on sugar or peptide concentration-regulated promoters, and bacteriophage-derived promoters [24, 25]. However, some of these systems are less useful since they are controllable only to a limited extent, show low efficiency, or are associated with some degree of basal expression [21, 25, 26]. These limitations may be due to (1) the corresponding inducer being an essential nutrient or metabolite, the concentration of which in the culture cannot be fully controlled, (2) by being strongly sensitive to catabolite repression (i.e., certain sugar-inducible systems) [21, 25, 26], or (3) the promoter showing leaky activity. To date, the most widely used and potent gene expression system in L. lactis has been the nisin-inducible controlled expression (NICE) system. When added to the medium as an inducer, nisin binds to the membrane receptor NisK, which subsequently activates NisR by phosphorylation, and the activated NisR induces the nisin A promoter [27–30].
It has been shown that the agmatine deiminase (AGDI) cluster of L. lactis subsp. cremoris CECT 8666 (formerly GE2-14) encodes the enzymatic activities responsible for the catabolism of agmatine to putrescine [31]. Briefly, aguD codes for the agmatine/putrescine antiporter, aguA encodes agmatine deiminase, aguB encodes putrescine transcarbamylase, and aguC encodes a specific carbamate kinase [32, 33]. Transcriptional analyses of these genes has shown the expression of aguB, aguD, aguA and aguC to be driven by the aguB promoter, and has confirmed the four genes to be co-transcribed as a single polycistronic mRNA [32, 33]. A cre site exists in the promoter of aguB which is transcriptionally regulated by carbon catabolite repression (CCR) mediated by the catabolite control protein CcpA [32, 33]. Also included in the AGDI cluster, upstream of the aguBDAC genes, is the aguR gene, which is transcribed constitutively under its own promoter (PaguR) (Fig. 1). We previously showed that aguR is a regulatory gene encoding a transmembrane protein (AguR) that acts as a one-component signal transduction system able to sense the extracellular agmatine concentration and trigger transcriptional activation of the aguB promoter (which drives expression of the aguBDAC operon) [34].
Fig. 1.
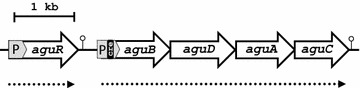
Genetic organization of the AGDI cluster of L. lactis CECT 8666. Physical map showing the cluster to be composed of five genes: aguR which encodes a transcription regulator, followed by aguB, aguD, aguA and aguC, which encode the proteins involved in the putrescine biosynthesis pathway (GenBank: AZSI00000000.1). The PaguR and PaguB promoters are shaded in grey, and the terminators indicated. The predicted transcripts are indicated below (dotted arrows). The cre site is shaded in black
The present work reports the adaptation of the ACE system—an inducible gene expression system that involves aguR and the PaguB promoter (the latter with its natural ribosome binding site)—to L. lactis. This was successfully tested via the production of green fluorescent protein (GFP) and Myxococcus xanthus prolyl-endopeptidase (PEP) [35, 36].
Results
Site-directed mutagenesis of the cre site of PaguB
In previous work [34] we showed that PaguB of L. lactis CECT 8666 drives expression of the aguBDAC operon in response to agmatine supplementation. Moreover, aguBDAC expression is regulated by CCR, mediated by the catabolite control protein A (CcpA) [33]. To develop an efficient gene expression system, CCR had to be inactivated so that glucose could be used as a carbon source. This allowed high bacterial cell densities to be obtained, and in turn high recombinant protein yields. For this we introduced arbitrarily directed mutations into the cre site (sequence 5′-TGAAATCGTTCCCA-3′; the nucleotides matching the cre consensus sequence are underlined) within PaguB (see “Methods”). The pAGDI plasmid [33], containing the PaguR-aguR-PaguB cassette fused to the reporter gene gfp (encoding green fluorescent protein [GFP]), was used to construct new plasmids to assess the effect of cre mutation on PaguB activity. Using this plasmid as a template, and the primers indicated in Table 1, three new plasmids were generated which contained specific mutations in the cre site: pAGDIcre1 (containing 10 nucleotide mutations from positions 5–14 of the cre site), pAGDIcre2 (containing 3 nucleotide mutations from positions 6–8 of the cre site) and pAGDIcre3 (containing 1 nucleotide mutation at position 5) (Fig. 2a). The 10 and 3 nucleotide mutations (plasmids pAGDIcre1 and pAGDIcre2 respectively) both had a dramatic effect on PaguB: no activity was detected with either construct at any glucose concentration (Fig. 2c, d; compare with Fig. 2b [wild type]). The introduction of a single mutation (A > T) in the cre site (pAGDIcre3 construct) gave the expected result, i.e., expression was not repressed at 120 mM glucose (Fig. 2e). This mutated promoter was therefore selected for the construction of the expression vector.
Table 1.
Oligonucleotides used in this study
| Primer | Function | Sequence (5′ to 3′) |
|---|---|---|
| mutf | Mutation of pAGDIcre1 (F) | CACACACACGAATTCTTGGAGTGGGAAGTCAAATAACTATTT |
| mutr | Mutation of pAGDIcre1 (R) | CACACACACGAATTCTTCAGTATAACAAGGTTGATTTCT |
| mutf1 | Mutation of pAGDIcre2 (F) | CACACACACGAATTCCCAAGTGGGAAGTCAAATAACTATTT |
| mutr1 | Mutation of pAGDIcre2 (R) | CACACACACGAATTCTTTCAGTATAACAAGGTTGATTTCTT |
| mutf4 | Mutation of pAGDIcre3 (F) | CACACACACGAATTCGTTCCCAAGTGGGAAGTCAAATAAC |
| mutr4 | Mutation pAGDIcre3 (R) | CACACACACGAATTCAGTATAACAAGGTTGATTTCTTAAAAC |
| prolF | Cloning of pep (F) | CACACACACCCATGGCTTATCCAGCTACACGTGC |
| prolR | Cloning of pep (R) | CACACACACTCTAGATTAACGTCCTTGTGCAGC |
| AgurBamHI | Cloning PaguR-AguR-PaguB cassette (F) | CCCCCCGGATCCGACAAGTTTGGCTCAGATTGCTTG |
| PtcNco | Cloning PaguR-AguR-PaguB cassette (R) | CCCCCCATGGTGTTTATTCCTCCTGAATAAAATAG |
| Expvfor1 | Insertion of His-Tag (F) | CACACACACCCATGGCTAATCGACTGCAGGAAAATTTATACTTCCAAGGTC |
| Expvrev1 | Insertion of His-Tag (R) | CTATCAATCAAAGCAACACGTG |
| GfF1 | Cloning of gfp (F) | CACACACACCCATGGAATTCAGTAAGGGAGAAGAACTTTTC |
| GfR1 | Cloning of gfp (R) | CACACACACCTGCAGACTAGTTTTGTAGAGCTCATCCATGC |
Fig. 2.
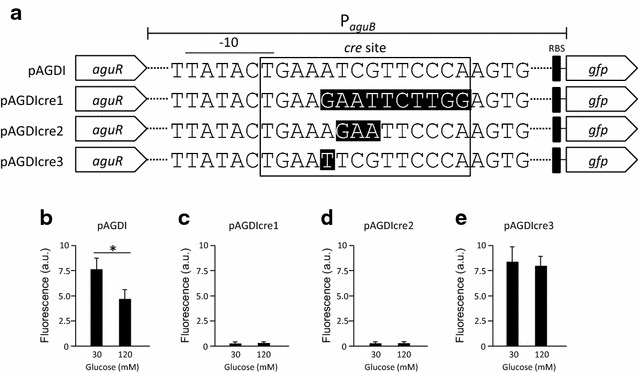
Generation of mutations in the cre site of PaguB in the AGDI cluster of L. lactis CECT 8666, and their effect on promoter activity. a Genetic detail of the different gfp fusions made with the wild type PaguB region (fusion pAGDI) and the derived promoters carrying different mutations in the cre site (fusions pAGDIcre1, pAGDIcre2 and pAGDIcre3). The introduced mutations are highlighted in black. Dashed lines indicate sequence discontinuities. b–e Promoter activity reported by GFP fluorescence (arbitrary units) for the wild type PaguB and mutants assayed in the presence of 30 and 120 mM glucose under induction by 20 mM agmatine. Data represent the average of three independent experiments. Bars indicate standard deviations (*p < 0.05)
Agmatine-induced heterologous expression of gfp
To verify the usefulness of the ACE system in the expression of heterologous genes, the reporter gene gfp was cloned into the pACE vector under the control of PaguB, thus generating the plasmid pACE-gfp. L. lactis NZ9000—a strain without the AGDI cluster—was transformed with pACE-gfp, thus resulting in L. lactis pACE-gfp. The presence of 10 mM agmatine in a culture of L. lactis pACE-gfp induced the expression of gfp, which was measured in terms of the fluorescence produced (7.87 arbitrary units) (Fig. 3). It should be noted that the expression of gfp was undetectable (<0.5 arbitrary units) in agmatine-uninduced cultures of L. lactis pACE-gfp. Fluorescence was also undetectable (<0.5 arbitrary units) in parallel agmatine-induced cultures of L. lactis NZ9000 harbouring the pACE vector (L. lactis pACE).
Fig. 3.
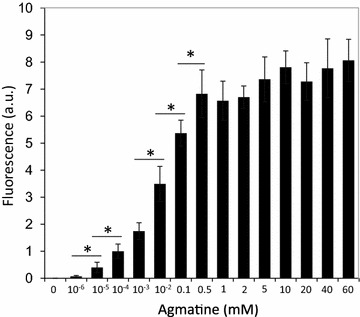
Sensitivity of the ACE system to agmatine. Strength of induction of gfp in L. lactis NZ9000 pACE-GFP. GFP protein activity was determined under a range of agmatine concentrations. Data represent the average of three independent experiments. Bars indicate standard deviations (*p < 0.05)
Sensitivity of the ACE system to the inducer: dose–response curve
The production of GFP in cultures of L. lactis pACE-gfp induced using a range of agmatine concentrations (between 0 and 60 mM) was analysed by whole-cell fluorescence. The ACE expression system showed great sensitivity to very low agmatine levels; a significant increase in fluorescence was seen after induction with agmatine at concentrations as low as 10−5 mM (Fig. 3). Above this concentration, the induction levels increased in line with the agmatine concentration until a maximum induction level (fluorescence ~8 arbitrary units) was reached at 0.5 mM agmatine (no significant increases in induction were seen with concentrations of >0.5 mM). The absence of any leaky activity of the promoter PaguB, as verified by the absence of fluorescence in uninduced cultures (0 mM agmatine, Fig. 3), is remarkable.
Heterologous production of GFP using the ACE system
The efficiency of the ACE system in overexpressing recombinant protein was tested with the reporter protein GFP. The expressed His-tagged GFP protein was purified using immobilized metal affinity chromatography (IMAC). The eluted protein fractions were examined by SDS-PAGE (data not shown) and their GFP activity. Pure protein with GFP activity was obtained in fraction two (of the four obtained); the yield was 47 % (Table 2). Fluorescence was found in the soluble fraction only (data not shown).
Table 2.
Purification of GFP protein using the ACE system
| Step | Total protein (µg) | Protein (µg ml−1) | Total activity (U mg−1) | Protein yield (%)a |
|---|---|---|---|---|
| Lysate | 63,927 | 15,981 | 80 | |
| Flow through | 53,463 | 13,365 | 0 | |
| Wash 1 | 3244 | 811 | 0 | |
| Wash 2 | 4220 | 1055 | 0 | |
| Elution 1 | 3049 | 6098 | 0 | |
| Elution 2 | 235 | 471 | 171 | 47 |
| Elution 3 | 55 | 111 | 0 | |
| Elution 4 | 488 | 977 | 0 |
aRecovery of fluorescence activity relative to the total activity of the soluble extract
Heterologous overproduction of M. xanthus prolyl-endopeptidase and comparison with the NICE system
To confirm the usefulness and efficiency of the ACE system, the pep gene of M. xanthus, which encodes a prolyl-endopeptidase of biomedical interest, was cloned into appropriate plasmids for introduction into L. lactis NZ9000. The resulting L. lactis NZ9000 pACE-pep was induced with different agmatine concentrations (0, 0.001, 0.01, 0.1, 0.5, 1, 2, 5, 10, 20, 40 and 60 mM) and the PEP activity assayed. No activity was detected in cultures without agmatine, but was observed even with the lowest agmatine concentration tested (0.001 mM). Above this concentration, the induction level increased with the agmatine concentration until 0.1 mM agmatine (21.04 mU mg−1) (no significant increase in PEP activity was obtained with concentrations of >0.1 mM) (Fig. 4). PEP activity was sought in the soluble and insoluble fractions, but was only seen in the former.
Fig. 4.
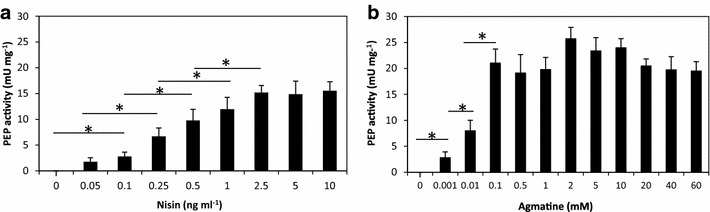
Comparison of PEP activity. a In the NICE system, PEP activity was monitored in L. lactis NZ9000 pNZ8048-pep cells induced at different nisin concentrations for 3 h after reaching OD600, while b in the ACE system, PEP activity was monitored in L. lactis NZ9000 pACE-pep cells induced with different agmatine concentrations (added to the culture medium before inoculation) after 7 h of growth. Data represent the mean of three independent experiments. Bars indicate standard deviations (*p < 0.05)
For comparison, PEP was also produced using the NICE system at different nisin concentrations (0, 0.05, 0.1, 0.25, 0.5, 1, 2.5, 5, 10 ng ml−1) (Fig. 4). The activity increased with the nisin concentration until 2.5 ng ml−1. The highest specific activity obtained (15.2 mU mg−1) was lower than that obtained with the ACE system (21.04 mU mg−1). Again, all PEP activity was observed in the soluble fraction.
Influence of agmatine on Lactococcus lactis growth
Since high induction concentrations of agmatine were tested in the present work, assays were performed to see whether these affected bacterial fitness. L. lactis cultures were grown in liquid GM17 supplemented with 0, 0.001, 0.01, 0.1, 0.5, 1, 2, 5, 10, 20, 40 or 60 mM agmatine. Similar growth curves were obtained (data not shown). Table 3 shows the OD600, μmax and pH values and lactate concentrations reached after 24 h of incubation. Growth slightly decreased with increasing concentration of agmatine. Although the 0.5 and 0.1 mM agmatine concentrations led to the highest GFP and PEP activities, no differences in OD600 were observed compared to the uninduced cultures. Organic acids and sugars were analyzed by HPLC, and no differences observed in the presence or absence of agmatine (data not show). Agmatine had a weak effect on L. lactis cell viability after 24 h of exposure; optical density values suitable for protein production were obtained even at high agmatine concentrations.
Table 3.
Effect of agmatine on growth, μmax, pH and production of lactic acid
| Agmatine (mM) | OD600 | μmax | pH | Lactic acid (mM) |
|---|---|---|---|---|
| 0 | 5.12 | 0.70 | 4.67 | 80.21 |
| 0.0001 | 4.86 | 0.69 | 4.69 | 81.15 |
| 0.001 | 4.44 | 0.67 | 4.7 | 80.10 |
| 0.01 | 3.9 | 0.62 | 4.72 | 80.35 |
| 0.1 | 4.5 | 0.65 | 4.77 | 79.83 |
| 0.5 | 4.11 | 0.65 | 4.79 | 79.07 |
| 1 | 4.11 | 0.66 | 4.79 | 78.55 |
| 2 | 4.11 | 0.63 | 4.79 | 78.22 |
| 5 | 4.31 | 0.60 | 4.77 | 78.62 |
| 10 | 3.89 | 0.61 | 4.77 | 78.89 |
| 20 | 4.12 | 0.61 | 4.78 | 76.14 |
| 40 | 3.65 | 0.58 | 4.8 | 73.49 |
| 60 | 3.85 | 0.59 | 4.78 | 67.40 |
Discussion
Lactococcus lactis has long been used in the food industry and has emerged as a cost-effective cellular factory for the production of proteins of interest [4]. At present, the genome sequences of several strains have been elucidated, and the genetic tools available for use with lactic acid bacteria (LAB) are ever increasing the number of fully lipopolysaccharide-free recombinant proteins that can be produced [3, 37].
The control of gene expression is critical in achieving high protein yields. For example, it is essential for ensuring the conservation of energy for the production of biomass prior to the directed overproduction of the target protein, and in controlling the production of products that might be toxic to the host cell (i.e., membrane proteins, autolysins, lysis-related proteins from phages) [3, 38]. Systems that allow for well controlled inducible expression, and that allow no basal expression, are essential in setting production rates. A number of inducible expression systems for L. lactis are available [20–22, 25, 39], however, the use of some of them, especially in large-scale fermentations, may be hampered by the low-level induction achieved, high background expression before induction (leaky activity), and/or a lack of control over the inducer [21, 25, 26, 39]. Such drawbacks are not suffered with the NICE system, probably the most commonly used regulated expression system for Gram-positive bacteria [27–29, 40]. However, when using a host strain other than L. lactis NZ9000, the NICE system requires the regulatory genes nisR and nisK to be supplied in trans. Thus, although the NICE system has been optimised to be used as a single-plasmid expression vector [41], its exploitation nearly always requires the integration of nisR and nisK, either in the chromosome of the host strain or via their cloning into a separate plasmid (the dual-plasmid strategy).
The present work describes an alternative system based on the aguR/PaguB cassette (the regulatory part of the AGDI cluster of L. lactis CECT 8666) for agmatine-controlled gene expression in L. lactis using a vector that includes all the elements required. It should be noted that this system requires the expression of no additional genes supplied in trans. The developed vector relies on the regulatory transmembrane protein AguR, which responds to extracellular agmatine, and in so doing triggers the induction of gene expression via PaguB [34]. It was previously shown that PaguB of L. lactis CECT 8666 contains a cre site involved in CCR and that this is repressed by glucose concentrations of >30 mM [33]. Since higher concentrations of glucose may be required in culture media to obtain the densities of bacterial cells needed to provide high recombinant protein yields, this repression mechanism was eliminated in the present work through the introduction of a single A > T mutation at position 5 in the cre site. Larger mutations of either 3 or 10 nucleotides completely impaired the activity of the promoter, most likely by preventing some additional regulatory signal. Other authors have also shown CCR to be relieved when single mutations occur in the cre site of CCR-controlled genes in L. lactis. For example, a single mutation in the cre site of the celB promoter allows fully active transcription of the cryptic cel cluster involved in lactose utilization in L. lactis MG1363 [42]. Similarly, two single mutations in the cre site of the ptcC promoter do away with the glucose-repressor effect and allow cells to constitutively metabolize cellobiose [43].
An expression vector combining the one-component signal transduction system, i.e., aguR and the aguB promoter, followed by convenient cloning sites for introducing the gene of interest, was constructed. An important feature of the developed pACE vector is the possibility of fusing a His-tag to the protein of interest by cloning the encoding gene in frame into the NcoI-PstI sites. His-tags have been used in other L. lactis expression vectors previously shown to perform efficiently in the immunodetection and purification of proteins [6, 44]. We here confirm the functionality of the His-tag in purifying the GFP (protein yield 47 %).
The control of the ACE system was assessed via the expression of gfp, the reporter gene coding for GFP. Strong fluorescence was seen in the presence of ≥0.5 mM agmatine (8 arbitrary units compared to 0 in uninduced cells). Interestingly, the system was associated with no basal expression, indicating PaguB to have no leaky activity. Further, agmatine is not present in common culture media, thus allowing for tight control over the gene to be expressed. Neither is it found in milk nor any derivative dairy environment where L. lactis occurs [45]. The optimal moment of induction, which can change from one overexpressed gene to another, needs to be evaluated. In the ACE system, the inducer agmatine can be added when the culture medium is prepared. The AguR/PaguB cassette, on which this expression system is based, is the regulatory part of the AGDI cluster of L. lactis [33] and is not active until the transition between the exponential and stationary phases is reached (5–6 h of culture) [46]. Thus, even when agmatine is supplied to the culture medium, the time of net expression would lie between 5 and 7 h after culturing began. This induction time is comparable to that associated with the NICE system (2–3 h after adding nisin). The addition of the inducer at the beginning of culturing avoids the problem of monitoring the culture’s optical density to determine the optimal moment for induction. Moreover, the abolition of sampling and inducer addition steps may prevent contamination, which could have serious economic consequences in industrial protein production. As seen for nisin, agmatine affected cell viability and caused a 20 % reduction in bacterial yield. However, the final OD (>3.3) of agmatine-induced cultures was still optimal for industrial protein production.
The price of the inducer is important in large scale fermentations. That of agmatine varies widely, depending on the supplier, but even the cheapest found (marketed as dietary supplement) worked properly as an inducer (data not shown). Certainly, it was much cheaper than nisin.
An agmatine induction system combining AguR and PaguB of Enterococcus faecalis was earlier used to develop an expression vector suitable for the latter species [47]. However, the proposed lactococcal ACE system shows some differences to the E. faecalis system: (1) the aguR gene is in the same orientation as PaguB (reflecting the corresponding organization of the AGDI cluster in each system), (2) the highest expression rate is reached at ≥0.5 mM agmatine in the ACE system but at ≥60 mM in the E. faecalis system, and (3) the level of induction (as determined by fluorescence) is less than that achieved with the E. faecalis system.
In the present work, the performance of the ACE system for the heterologous production of the M. xanthus PEP was compared to that obtained using the NICE system, the most widely used and potent gene expression system in L. lactis [40]. The proposed system achieved higher PEP activity (circa 38 %) under similar laboratory conditions. The observed differences might be related to the characteristics of each promoter, or to the effect on signal transduction of the two-component NICE system compared to the one-component ACE system. More studies are required to understand how the agmatine signal is transduced.
In summary, the present results confirm the ACE system as an attractive candidate for high level recombinant protein production. The lactococcal aguR/PaguB system can effectively control the expression of genes in response to agmatine in L. lactis without any basal expression, and combines both the expression cassette and regulatory gene in one plasmid. This vector expands the genetic toolbox available for this species, and could be a powerful and straightforward alternative system for overexpressing proteins in lactococcal strains lacking nisR and nisK. It might also be used to complement the NICE system and be used in co-expression.
Conclusions
The present work describes the construction of a L. lactis agmatine-controlled expression system based on the aguR/PaguB cassette of the putrescine biosynthesis gene cluster. A single mutation of the cre site in PaguB abolished the CCR of this promoter. This system was assessed by expressing the reporter gene gfp, and fluorescence was found strictly dependent on the agmatine concentration added to the culture medium, with maximum induction occurring at 0.5 mM agmatine (7 arbitrary units compared to 0 in uninduced cells). An important potential benefit of this system is the lack of leaky activity associated with it, and the fact that gene expression can be tightly controlled via the addition of the appropriate concentration of agmatine. The pACE vector allowed the agmatine-inducible expression of the gene encoding M. xanthus PEP, an enzyme that can degrade the immunotoxic peptides of gluten breakdown. Moreover, enzymatic activity was greater than that obtained with the NICE expression system. The addition of a His-tag to the pACE vector renders the system suitable for protein purification and immunodetection purposes. Together, these findings suggest that the ACE expression system could be a very valuable addition to the L. lactis genetic toolbox, and offers a straightforward, alternative inducible gene expression system that to be used in functional studies and in the large-scale production of recombinant proteins.
Methods
Bacterial strains and growth conditions
L. lactis CECT 8666 (formerly GE2-14) and L. lactis NZ9000 were grown at 30 °C in M17 medium (Oxoid, Basingstoke, United Kingdom) supplemented with 30 mM glucose (GM17). When required, agmatine (Sigma-Aldrich, St. Louis, MO, USA) was added to the medium. Chloramphenicol (5 μg ml−1) was added as required for plasmid maintenance. For overexpression using the NICE system, cultures of L. lactis NZ9000 in exponential phase (OD600 = 0.4–0.5), grown in GM17, were induced for 3 h with various nisin concentrations (0, 0.05, 0.1, 0.25, 0.5, 1, 2.5, 5 and 10 ng ml−1) (Sigma-Aldrich). Solid media were prepared by adding 2 % (w/v) agar (Merck, Darmstad, Germany). Microbial growth was examined in all cultures by measuring absorbance at 600 nm (OD600) using a spectrophotometer (Eppendorf, NY, USA). The pH of the samples was measured using a CRISON miCropH 2001 pH meter (Crison Instruments S.A., Barcelona, Spain). The maximum specific growth rate (μmax) was determined experimentally in the exponential growth phase, as described by O´Sullivan and Condon [48].
DNA manipulation
The procedures used for DNA manipulation and recombination were essentially those described by Sambrook et al. [49]. Table 1 lists the sequences of primers used. Genetic constructs were achieved in L. lactis NZ9000. The isolation of L. lactis plasmids and total DNA, and the transformation procedures followed, were as previously described [50]. All plasmid constructs were verified by nucleotide sequencing at Macrogen Inc. (Seoul, Republic of Korea). All enzymes for DNA technology were used according to the manufacturer’s specifications.
Construction of plasmids
The core of the lactococcal vector pNZ8048 [29]—which includes the replication cassette and the chloramphenicol resistance marker—was used as a starting point for the construction of the pACE vector. First, a fragment of the AGDI cluster from L. lactis CECT 8666 (Table 4), including the aguR promoter (PaguR), the aguR gene, and the aguB promoter (PaguB) carrying the mutation in the cre site, was PCR-amplified (using pAGDIcre3 as a template) and cloned into the BglII-NcoI sites of pNZ8048. Subsequently, a fragment including the multicloning site and a histidine tag encoding 10 consecutives histidines (His-tag) was amplified from plasmid pNZErmC [6] and cloned into the NcoI-XbaI sites of the previous construct, thus yielding vector pACE (Fig. 5). This vector offers the option to fuse in-frame the gene encoding the protein of interest to a C-terminal His-Tag by cloning the target gene into the NcoI-PstI sites. Thus, this vector could be used for immunodetection or purification of the proteins encoded by overexpressed genes. The target gene could also be cloned without the His-tag for use in functional studies.
Table 4.
Strains and plasmids
| Strain/plasmid | Characteristics | References |
|---|---|---|
| Strains | ||
| L. lactis CECT8666 (formerly GE2-24) | Isolated from an artisanal cheese, containing AGDI cluster | [31] |
| L. lactis NZ9000 | Expression host for NICE system, lacking AGDI cluster | [29] |
| Plasmids | ||
| pNZ8048 | Lactococcal plasmid containing the nisA promoter and the Cmr | [29] |
| pNZErmC | pNZ8048 derivative containing the His-tag and the Cmr | [6] |
| pUC57-pep | pUC57 derivative containing the pep gene from M. Xanthus and the Apr | [36] |
| pAGDI | pNZ8048 derivative bearing the PaguR-aguR-PaguB-gfp fusion and the Cmr | [34] |
| pAG2 | pNZ8048 derivative bearing the PaguB-gfp fusion and the Cmr | [33] |
| pAGDIcre1 | pAGDI derivative bearing the mutated PaguB and the Cmr | This work |
| pAGDIcre2 | pAGDI derivative bearing the mutated PaguB and the Cmr | This work |
| pAGDIcre3 | pAGDI derivative bearing the mutated PaguB and the Cmr | This work |
| pACE | Vector for ACE system containing the PaguR-aguR-PaguB cassette from pAGDIcre3 and the Cmr | This work |
| pACE-gfp | pACE derivative harbouring the gfp gene from pAG2 and the Cmr | This work |
| pNZ8048-pep | pNZ8048 derivative harbouring the pep gene from pUC57-PEPand the Cmr | This work |
| pACE-pep | pACE derivative harbouring the pep gene from pNZ8048-PEP and the Cmr | This work |
PaguR, aguR promoter; PaguB, aguB promoter; pep, prolyl endopeptidase gene; Cmr, chloramphenicol resistance marker; Apr, ampicillin resistance marker
Fig. 5.
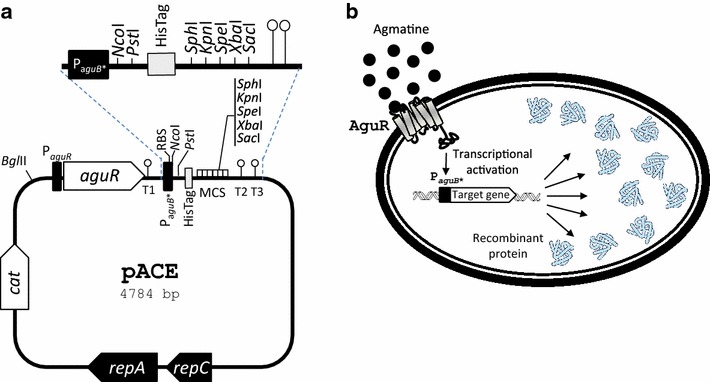
a Genetic map of the pACE expression vector. repC and repA, replication genes; cat, chloramphenicol resistance gene; aguR, gene encoding the regulatory agmatine-sensor-regulator protein AguR; PaguR, aguR promoter; RBS, ribosome binding site; T1, T2 and T3, transcription terminators (ΔG = −10.3, −9.7 and −8.3 kcal/mol respectively); PaguB*, aguB agmatine-inducible promoter carrying the A > T mutation in the fifth nucleotide of the cre site; MCS, multicloning site; HisTag; C-terminal histidine tag. Representative restriction sites are indicated. b Overview of the AguR/PaguB expression system including its components and role. AguR is the transmembrane agmatine (black filled circles) sensor and response regulator that activates PaguB, thereby triggering the massive overproduction of the recombinant target protein
For the heterologous expression of GFP, the gfp gene (amplified from pAG2 [34]) was inserted into the NcoI-PstI sites of pACE, thus generating the plasmid pACE-gfp. To produce PEP using the ACE system, the pep gene was PCR-amplified from pNZ8048-pep and cloned into the NcoI-XbaI sites in the pACE vector, resulting in the pACE-pep plasmid. To produce PEP using the NICE system, the pep gene was released from plasmid pUC57-pep [36] as a NcoI-XbaI fragment and cloned into the same sites in pNZ8048 under the control of the nisA promoter [29], thus generating plasmid pNZ8048-pep.
Directed mutagenesis of the cre site of PaguB
Modification of the cre site was achieved by in vitro site-directed mutagenesis. Mutation(s) were introduced by PCR using two divergent primers (Table 1) spanning the cre site of PaguB and containing the desired mutation(s). Each primer was complementary to the opposite strand of the pAGDI vector, which was used as template to generate plasmids pAGDIcre1, pAGDIcre2 and pAGDIcre3 containing the specific mutations in the cre site. The pAGDI plasmid was first methylated with Dam methylase and S-adenosyl methionine following the manufacturer’s instructions (New England Biolabs, Hertfordshire, UK). Phusion High-Fidelity DNA Polymerase (New England Biolabs) was used to amplify both plasmid strands with high fidelity. The PCR thermoycling conditions were as follows: initial denaturation (98 °C for 30 s), 32 cycles of amplification (98 °C for 10 s; 55 °C for 30 s; and 72 °C for 2.5 min) plus a final extension step (72 °C for 7 min). An EcoRI target site was included in the primers so that the obtained amplicons could be digested with EcoRI and self-ligated. Before transformation in L. lactis NZ9000, the ligation mixture was treated with DpnI order to digest the original, Dam methylated pAGDI plasmid used as a template.
Measurement of green fluorescence
For whole-cell fluorescence measurements, overnight cultures of L. lactis NZ9000 harbouring either pACE or pACE-gfp were transferred (1 %) to fresh medium (GM17) supplemented with different agmatine concentrations (0–60 mM) and grown for 7 h. Equal amounts of cells were harvested, washed, and then resuspended in 50 mM potassium phosphate buffer, pH 7.2, as previously described [6]. GFP emission was measured in a volume of 200 μl of cells, using a Cary Eclipse fluorescence spectrophotometer (Varian Inc., Palo Alto, CA, USA) at an excitation wavelength of 485 nm and an emission wavelength 530 nm. To facilitate direct comparisons, the bacterial cultures used for GFP fluorescence measurements contained similar amounts of cells (estimation was made based on OD600). Background fluorescence levels were assessed using non-fluorescent control cells (lacking the gfp gene), and these values subtracted from the experimental results.
Prolyl-endopeptidase assay
PEP activity was determined using a synthetic substrate, succinyl-Ala-Pro-p-NA (NA, nitroanilide) (Bachem, Bubendorf, Switzerland), as previously described [36] with slight modifications. Bacterial cultures (10 ml) of L.lactis pNZ8048-pep (induced with nisin for 3 h after the cells reached an OD600 of 0.6) and L. lactis pACE-pep (induced with 20 mM agmatine [added when the culture medium was prepared] and grown for 7 h) were harvested by centrifugation (8000g for 10 min), washed twice, and resuspended in 2 ml of 50 mM phosphate buffer, 0.2 M NaCl, pH 7.5. The samples were then disrupted using 200 mg glass beads (<106 µm) (Sigma-Aldrich) in a Fast-Prep FP120 Instrument (Thermo Savant-BIO101/Q-Biogen, CA, USA) for 6 × 30 s at power setting 4.5 (with intermittent cooling). Cell debris was removed by centrifugation (10,000g for 30 min at 4 °C) and the supernatant used in activity assays. The assay mixture contained 625 μl of 50 mM phosphate buffer (pH 7.5), 0.2 M NaCl, 125 μl substrate (1.2 mM), and 250 μl of cell extracts. The reaction was stopped by adding 500 μl of 20 % trichloroacetic acid. The samples were then centrifuged (10,000g for 10 min) and the release of the p-NA spectrophotometrically detected at 410 nm in a U-2800 Digilab Hitachi spectrophotometer (HitachiHigh-Technologies Corporation, Tokyo, Japan). One activity unit was defined as the amount of enzyme required to release 1 μmol of p-NA per min under the assay conditions. Assays were performed in triplicate. Specific enzyme activity was expressed as milliunits per milligram of protein. The protein concentration was measured using a Pierce BCA Assay Kit (Thermo Fisher scientific) following the manufacturer’s indications.
Purification His-tagged protein
Purification of the His-tagged GFP protein was performed by immobilized metal ion affinity chromatography (IMAC). Cells (200 ml) induced with 10 mM agmatine were harvested by centrifugation at 8000g, at 4 °C for 10 min after 7 h of growth. The supernatant was discarded and cells washed twice and resuspended in 4 ml phosphate buffer (50 mM pH 7.5). They were then disrupted using a French Press operating at 2.3 kbar [Constant Cell Disruption Systems (Low March, Daventry, Norttants, UK)]. Cell debris was removed by centrifugation (10,000g for 30 min at 4 °C). Imidazole was then added to a concentration of 10 mM, and the resulting samples employed in protein purification using the QIAexpressionist kit (Quiagen, Madrid, Spain) following the manufacturer’s instructions. Aliquots of collected fractions were analyzed by SDS-PAGE using 12 % acrylamide gels to determine the purity of the His6-taggeted proteins. Their GFP activities and protein concentrations were determined using the protocols mentioned above to determine the protein yield.
Soluble and insoluble protein fractions
Soluble and insoluble proteins fractions were prepared following the protocol described by Cano-Garrido et al. [51]. Samples (10 ml) of bacterial cultures grown at different agmatine concentrations were pelleted by centrifugation at 8000g at 4 °C for 10 min and the sediment resuspended in 1 ml of the appropriate buffer depending on the protein expressed (GFP—phosphate buffer 50 mM, pH 7.5; PEP—phosphate buffer 50 mM, pH 7.5, 0.2 M NaCl). The samples were then disrupted with 200 mg glass beads (<106 µm) (Sigma-Aldrich) in a Fast-Prep FP120 Instrument (Thermo Savant-BIO101/Q-Biogen, CA, USA) for 6 × 30 s at a power setting of 4.5 (with intermittent cooling). Total cell extracts were centrifuged at 15,000g at 4 °C for 15 min. Finally, the insoluble fractions were resuspended in 1 ml of the appropriate buffer and fluorescence and PEP activity monitored in both the soluble and insoluble-resuspended fractions.
Determination of organic acids and sugars by HPLC
Sugar and organic acid concentrations were determined using a chromatographic system composed of an Alliance 2690 module injector, a Photodiode Array PDA 996 detector, and a 410 Differential Refractometer detector, all controlled with Millennium 32 software (Waters, Milford, MA, USA). Supernatants (50 μl) were isocratically separated in a 300 × 7.8 mm HPX-87H Aminex ion-exchange column (Hewlett Packard, Palo Alto, CA, USA) protected by a cation H+ Microguard cartridge (BioRad, Laboratories, Richmond, CA, USA), at a flow rate of 0.7 ml/min and a temperature of 65 °C. Sulphuric acid (3 mM) was used as the mobile phase. A PDA 996 detector at 210 nm was used to identify and quantify the organic acids detected, whereas the sugars were analyzed with a 410 Refractometer. Solutions of lactic and acetic acids, glucose, galactose, lactose, and sucrose were used as standards in the identification and quantification procedure.
Statistical analysis
The Student t test was used to examine differences between groups. Significance was set at p < 0.05.
Authors’ contributions
DML designed and performed some experiments and drafted the manuscript; PAS and BR performed some experiments; VL, BdR, MC and MF participated in the design of the study and helped to write the manuscript; MAG provided the general concept and supervised the work and the manuscript. All authors contributed to the discussion of the research. All authors read and approved the final manuscript.
Acknowledgements
This work was performed with the financial support of the Spanish Ministry of Economy and Competitiveness (AGL2013-45431-R), the Principality of Asturias Plan for Science, Technology and Innovation 2013-2017, and FEDER funds (GRUPIN14-137). We are grateful to Bert Poolman for providing the GFP-based cloning vectors and Adrian Burton for linguistic assistance. L. lactis NZ9000 and plasmid pNZ8048 were kindly provided by NIZO Food Research. We acknowledge support of the publication fee by the CSIC Open Access Publication Support Initiative through its Unit of Information Resources for Research (URICI). P.A.S. was the beneficiary of a fellowship from the FICYT, Principality of Asturias, Spain (BP09093). D.M.L. and B.d.R. were beneficiaries of JAE DOC contracts (CSIC).
Competing interests
The authors declare that they have no competing interests.
Abbreviations
- ACE
agmatine-controlled expression
- AGDI
agmatine deiminase
- CCR
carbon catabolite repression
- GFP
green fluorescence protein
- NICE
nisin-inducible controlled expression
- PEP
prolyl-endopeptidase
Contributor Information
Daniel M. Linares, Email: daniml@ipla.csic.es
Patricia Alvarez-Sieiro, Email: p.a.sieiro@gmail.com.
Beatriz del Rio, Email: beadelrio@ipla.csic.es.
Victor Ladero, Email: ladero@ipla.csic.es.
Begoña Redruello, Email: bredruel@ipla.csic.es.
Mª Cruz Martin, Email: mcm@ipla.csic.es.
Maria Fernandez, Phone: +34 985 89 21 31, Email: mfernandez@ipla.csic.es.
Miguel A. Alvarez, Email: maag@ipla.csic.es
References
- 1.Nocon J, Steiger MG, Pfeffer M, Sohn SB, Kim TY, Maurer M, Rußmayer H, Pflügl S, Ask M, Haberhauer-Troyer C, Ortmayr K, Hann S, Koellensperger G, Gasser B, Lee SY, Mattanovich D. Model based engineering of Pichia pastoris central metabolism enhances recombinant protein production. Metab Eng. 2014;24:129–138. doi: 10.1016/j.ymben.2014.05.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Rosano GL, Ceccarelli EA. Recombinant protein expression in Escherichia coli: advances and challenges. Front Microbiol. 2014;5:172. doi: 10.3389/fmicb.2014.00172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Mierau I, Olieman K, Mond J, Smid EJ. Optimization of the Lactococcus lactis nisin-controlled gene expression system NICE for industrial applications. Microb Cell Fact. 2005;4:16. doi: 10.1186/1475-2859-4-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Cano-Garrido O, Rueda FL, Sànchez-García L, Ruiz-Ávila L, Bosser R, Villaverde A, García-Fruitós E. Expanding the recombinant protein quality in Lactococcus lactis. Microb Cell Fact. 2014;13:167. doi: 10.1186/s12934-014-0167-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bourdichon F, Casaregola S, Farrokh C, Frisvad JC, Gerds ML, Hammes WP, Harnett J, Huys G, Laulund S, Ouwehand A, Powell IB, Prajapati JB, Seto Y, Ter Schure E, Van Boven A, Vankerckhoven V, Zgoda A, Tuijtelaars S, Hansen EB. Food fermentations: microorganisms with technological beneficial use. Int J Food Microbiol. 2012;154:87–97. doi: 10.1016/j.ijfoodmicro.2011.12.030. [DOI] [PubMed] [Google Scholar]
- 6.Linares DM, Geertsma ER, Poolman B. Evolved Lactococcus lactis strains for enhanced expression of recombinant membrane proteins. J Mol Biol. 2010;401:45–55. doi: 10.1016/j.jmb.2010.06.002. [DOI] [PubMed] [Google Scholar]
- 7.Kunji ER, Chan KW, Slotboom DJ, Floyd S, O’Connor R, Monné M. Eukaryotic membrane protein overproduction in Lactococcus lactis. Curr Opin Biotechnol. 2005;16:546–551. doi: 10.1016/j.copbio.2005.08.006. [DOI] [PubMed] [Google Scholar]
- 8.Bermúdez-Humarán LG, Kharrat P, Chatel JM, Langella P. Lactococci and lactobacilli as mucosal delivery vectors for therapeutic proteins and DNA vaccines. Microb Cell Fact. 2011;10:S4. doi: 10.1186/1475-2859-10-S1-S4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.D’Souza R, Pandeya DR, Hong ST. Lactococcus lactis: an efficient Gram positive cell factory for the production and secretion of recombinant protein. Biomed Res. 2012;23:1–7. [Google Scholar]
- 10.Lee MH, Roussel Y, Wilks M, Tabaqchali S. Expression of Helicobacter pylori urease subunit B gene in Lactococcus lactis MG1363 and its use as a vaccine delivery system against H. pylori infection in mice. Vaccine. 2001;19:3927–3931. doi: 10.1016/S0264-410X(01)00119-0. [DOI] [PubMed] [Google Scholar]
- 11.Ribeiro LA, Azevedo V, Le Loir Y, Oliveira SC, Dieye Y, Piard JC, Gruss A, Langella P. Production and targeting of the Brucella abortus antigen L7/L12 in Lactococcus lactis: a first step towards food-grade live vaccines against brucellosis. Appl Environ Microbiol. 2002;68:910–916. doi: 10.1128/AEM.68.2.910-916.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Xin KQ, Hoshino Y, Toda Y, Igimi S, Kojima Y, Jounai N, Ohba K, Kushiro A, Kiwaki M, Hamajima K, Klinman D, Okuda K. Immunogenicity and protective efficacy of orally administered recombinant Lactococcus lactis expressing surface-bound HIV Env. Blood. 2003;10:223–228. doi: 10.1182/blood-2003-01-0110. [DOI] [PubMed] [Google Scholar]
- 13.Robinson K, Chamberlain LM, Lopez MC, Rush CM, Marcotte H, Le Page RW, Wells JM. Mucosal and cellular immune responses elicited by recombinant strains of Lactococcus lactis expressing tetanus toxin fragment C. Infect Immun. 2004;72:2753–2756. doi: 10.1128/IAI.72.5.2753-2761.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Bermúdez-Humarán LG, Cortes-Perez NG, Lefèvre F, Guimarães V, Rabot S, Alcocer-Gonzalez JM, Gratadoux JJ, Rodriguez-Padilla C, Tamez-Guerra RS, Corthier G, Gruss A, Langella P. A novel mucosal vaccine based on live lactococci expressing E7 antigen and IL-12 Induces systemic and mucosal immune responses and protects mice against human papillomavirus type 16-Induced tumors. J Immunol. 2005;175:7297–7302. doi: 10.4049/jimmunol.175.11.7297. [DOI] [PubMed] [Google Scholar]
- 15.Ramasamy R, Yasawardena S, Zomer A, Venema G, Kok J, Leenhouts K. Immunogenicity of a malaria parasite antigen displayed by Lactococcus lactis in oral immunisations. Vaccine. 2006;24:3900–3908. doi: 10.1016/j.vaccine.2006.02.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Van Roosmalen ML, Kanninga R, El Khattabi M, Neef J, Audouy S, Bosma T, Kuipers A, Post E, Steen A, Kok J, Buist G, Kuipers OP, Robillard G, Leenhouts K. Mucosal vaccine delivery of antigens tightly bound to an adjuvant particle made from food-grade bacteria. Methods. 2006;38:144–149. doi: 10.1016/j.ymeth.2005.09.015. [DOI] [PubMed] [Google Scholar]
- 17.Mercenier A, Müller-Alouf H, Grangette C. Lactic acid bacteria as live vaccines. Curr Issues Mol Biol. 2000;2:17–25. [PubMed] [Google Scholar]
- 18.Zamri HF, Shamsudin MN, Rahim RA, Neela V. Oral vaccination with Lactococcus lactis expressing the Vibrio cholerae Wzm protein to enhance mucosal and systemic immunity. Vaccine. 2012;30:3231–3238. doi: 10.1016/j.vaccine.2012.02.012. [DOI] [PubMed] [Google Scholar]
- 19.Douillard FP, O’Connell-Motherway M, Cambillau C, van Sinderen D. Expanding the molecular toolbox for Lactococcus lactis: construction of an inducible thioredoxin gene fusion expression system. Microb Cell Fact. 2011;10:66. doi: 10.1186/1475-2859-10-66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Sanders JW, Venema G, Kok J. A chloride-inducible gene expression cassette and its use in induced lysis of Lactococcus lactis. Appl Environ Microbiol. 1997;63:4877–4882. doi: 10.1128/aem.63.12.4877-4882.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Mu D, Montalbán-López M, Masuda Y, Kuipers OP. Zirex: a novel zinc-regulated expression system for Lactococcus lactis. Appl Environ Microbiol. 2013;79:4503–4508. doi: 10.1128/AEM.00866-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Jorgensen CM, Vrang A, Madsen SM. Recombinant protein expression in Lactococcus lactis using the P170 expression system. FEMS Microbiol Lett. 2014;351:170–178. doi: 10.1111/1574-6968.12351. [DOI] [PubMed] [Google Scholar]
- 23.van Asseldonk M, Simons A, Visser H, de Vos WM, Simons G. Cloning, nucleotide sequence, and regulatory analysis of the LactococcuslactisdnaJ gene. J Bacteriol. 1993;175:1637–1644. doi: 10.1128/jb.175.6.1637-1644.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Marugg JD, Meijer W, van Kranenburg R, Laverman P, Bruinenberg PG, de Vos WM. Medium-dependent regulation of proteinase gene expression in Lactococcus lactis: control of transcription initiation by specific dipeptides. J Bacteriol. 1995;177:2982–2989. doi: 10.1128/jb.177.11.2982-2989.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.de Vos WM. Gene expression systems for lactic acid bacteria. Curr Opin Microbiol. 1999;2:289–295. doi: 10.1016/S1369-5274(99)80050-2. [DOI] [PubMed] [Google Scholar]
- 26.de Ruyter PG, Kuipers OP, de Vos WM. Controlled gene expression systems for Lactococcus lactis with the food-grade inducer nisin. Appl Environ Microbiol. 1996;62:3662–3667. doi: 10.1128/aem.62.10.3662-3667.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kuipers OP, Beerthuyzen MM, de Ruyter PG, Luesink EJ, de Vos WM. Autoregulation of nisin biosynthesis in Lactococcus lactis by signal transduction. J Biol Chem. 1995;270:27299–27304. doi: 10.1074/jbc.270.45.27299. [DOI] [PubMed] [Google Scholar]
- 28.Kuipers OP, Rollema HS, Beerthuyzen MM, Siezen RJ, de Vos WM. Protein engineering and biosynthesis of nisin and regulation of transcription of the structural nisA gene. Int Dairy J. 1995;5:785–795. doi: 10.1016/0958-6946(95)00032-1. [DOI] [Google Scholar]
- 29.Kuipers OP, de Ruyter PG, Kleerebezem M, de Vos WM. Quorum sensing-controlled gene expression in lactic acid bacteria. J Biotechnol. 1998;64:15–21. doi: 10.1016/S0168-1656(98)00100-X. [DOI] [Google Scholar]
- 30.Mierau I, Kleerebezem M. 10 years of the nisin-controlled gene expression system (NICE) in Lactococcus lactis. Appl Microbiol Biotechnol. 2005;68:705–717. doi: 10.1007/s00253-005-0107-6. [DOI] [PubMed] [Google Scholar]
- 31.Ladero V, Del Rio B, Linares DM, Fernandez M, Mayo B, Martin MC, Alvarez MA. Genome sequence analysis of the biogenic amine-producing strain Lactococcus lactis subsp. cremoris CECT 8666 (Formerly GE2-14) Genome Announc. 2014;2:e01088–e01114. doi: 10.1128/genomeA.01088-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ladero V, Rattray FP, Mayo B, Martín MC, Fernández M, Alvarez MA. Sequencing and transcriptional analysis of the biosynthesis gene cluster of putrescine-producing Lactococcus lactis. Appl Environ Microbiol. 2011;77:6409–6418. doi: 10.1128/AEM.05507-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Linares DM, del Río B, Ladero V, Redruello B, Martín MC, Fernández M, Alvarez MA. The putrescine biosynthesis pathway in Lactococcus lactis is transcriptionally regulated by carbon catabolic repression, mediated by CcpA. Int J Food Microbiol. 2013;165:43–50. doi: 10.1016/j.ijfoodmicro.2013.04.021. [DOI] [PubMed] [Google Scholar]
- 34.Linares DM, del Rio B, Redruello B, Ladero V, Martin MC, de Jong A, Kuipers OP, Fernandez M, Alvarez MA. AguR, a transmembrane transcription activator of the putrescine biosynthesis operon in Lactococcus lactis, acts in response to agmatine concentration. Appl Environ Microbiol. 2015;81(18). doi:10.1128/AEM.00959-15. [DOI] [PMC free article] [PubMed]
- 35.Plugis NM, Khosla C. Therapeutic approaches for celiac disease. Best Pract Res Clin Gastroenterol. 2015;29:503–521. doi: 10.1016/j.bpg.2015.04.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Alvarez-Sieiro P, Martin MC, Redruello B, Del Rio B, Ladero V, Palanski BA, Khosla C, Fernandez M, Alvarez MA. Generation of food-grade recombinant Lactobacillus casei delivering Myxococcus xanthus prolyl endopeptidase. Appl Microbiol Biotechnol. 2014;98:6689–6700. doi: 10.1007/s00253-014-5730-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Morello E, Bermúdez-Humarán LG, Llull D, Solé V, Miraglio N, Langella P, Poquet I. Lactococcus lactis, an efficient cell factory for recombinant protein production and secretion. J Mol Microbiol Biotechnol. 2008;14:48–58. doi: 10.1159/000106082. [DOI] [PubMed] [Google Scholar]
- 38.Kunji ER, Slotboom DJ, Poolman B. Lactococcus lactis as host for overproduction of functional membrane proteins. Biochim Biophys Acta. 2003;1610:97–108. doi: 10.1016/S0005-2736(02)00712-5. [DOI] [PubMed] [Google Scholar]
- 39.Llull D, Poquet I. New expression system tightly controlled by zinc availability in Lactococcus lactis. Appl Environ Microbiol. 2004;70:5398–5406. doi: 10.1128/AEM.70.9.5398-5406.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kuipers OP, de Ruyter PG, Kleerebezem M, de Vos WM. Controlled overproduction of proteins by lactic acid bacteria. Trends Biotechnol. 1997;15:135–140. doi: 10.1016/S0167-7799(97)01029-9. [DOI] [PubMed] [Google Scholar]
- 41.Bryan EM, Bae T, Kleerebezem M, Dunny GM. Improved vectors for nisin-controlled expression in gram-positive bacteria. Plasmid. 2000;44:183–190. doi: 10.1006/plas.2000.1484. [DOI] [PubMed] [Google Scholar]
- 42.Solopova A, Bachmann H, Teusink B, Kok J, Neves AR, Kuipers OP. A specific mutation in the promoter region of the silent cel cluster accounts for the appearance of lactose-utilizing Lactococcus lactis MG1363. Appl Environ Microbiol. 2012;78:5612–5621. doi: 10.1128/AEM.00455-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Linares DM, Kok J, Poolman B. Genome sequences of Lactococcus lactis MG1363 (revised) and NZ9000 and comparative physiological studies. J Bacteriol. 2010;192:5806–5812. doi: 10.1128/JB.00533-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Geertsma ER, Groeneveld M, Slotboom DJ, Poolman B. Quality control of overexpressed membrane proteins. Proc Natl Acad Sci USA. 2008;105:5722–5727. doi: 10.1073/pnas.0802190105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Galgano F, Caruso M, Condelli N, Favati F. Focused review: agmatine in fermented foods. Front Microbiol. 2012;3:199. doi: 10.3389/fmicb.2012.00199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.del Rio B, Ladero V, Redruello B, Linares DM, Fernández M, Martín MC, Alvarez MA. Lactose-mediated carbon catabolite repression of putrescine production in dairy Lactococcus lactis is strain dependent. Food Microbiol. 2015;48:163–170. doi: 10.1016/j.fm.2014.11.018. [DOI] [PubMed] [Google Scholar]
- 47.Linares DM, Perez M, Ladero V, Del Rio B, Redruello B, Martin MC, Fernandez M, Alvarez MA. An agmatine-inducible system for the expression of recombinant proteins in Enterococcus faecalis. Microb Cell Fact. 2014;13:169. doi: 10.1186/s12934-014-0169-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.O’Sullivan E. Condon S Relationship between acid 499 tolerance, cytoplasmic pH, and ATP and H+ -ATPase levels in chemostat cultures of Lactococcus lactis. Appl Environ Microbiol. 1999;65:2287–2293. doi: 10.1128/aem.65.6.2287-2293.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Sambrook JD, Russell D. Molecular cloning a laboratory manual. Cold Spring Harbor: Cold Spring Harbor Laboratory; 2001. [Google Scholar]
- 50.de Vos WM, Vos P, Dehaard H, Boerritger I. Cloning and expression of the Lactococcus lactis ssp cremoris SK11 gene encoding an extracellular serine proteinase. Gene. 1989;85:169–176. doi: 10.1016/0378-1119(89)90477-0. [DOI] [PubMed] [Google Scholar]
- 51.Cano-Garrido O, Rueda FL, Sànchez-García L, Ruiz-Ávila L, Bosser R, Villaverde A, García-Fruitós E. Expanding the recombinant protein quality in Lactococcus lactis. Microb Cell Fact. 2014;13:167. doi: 10.1186/s12934-014-0167-3. [DOI] [PMC free article] [PubMed] [Google Scholar]