

Abstract
Background
Carbapenem-resistant Pseudomonas aeruginosa (CRPA) and Acinetobacter baumannii (CRAB) are the leading causes of nosocomial infections. A rapid and sensitive test to detect CRPA and CRAB is required for appropriate antibiotic treatment. We optimized a loop-mediated isothermal amplification (LAMP) assay to detect the presence of blaVIM-2, blaIMP-1, and blaOXA-23, which are critical components for carbapenem resistance.
Methods
Two sets of primers, inner and outer primers, were manually designed as previously described. The LAMP buffer was optimized (at 2mM MgSO4) by testing different concentrations of MgSO4. The optimal reaction temperature and incubation time were determined by using a gradient thermocycler. Then, the optimized blaVIM-2, blaIMP-1, and blaOXA-23 LAMP reactions were evaluated by using 120 P. aeruginosa and 99 A. baumannii clinical isolates.
Results
Only one strain of the 100 CRPA isolates harbored blaIMP-1, whereas none of them harbored blaVIM-2. These results indicate that the acquisition of blaVIM-2 or blaIMP-1 may not play a major role in carbapenem resistance in Korea. Fifty two strains of the 75 CRAB isolates contained blaOXA-23, but none contained blaVIM-2 and blaIMP-1 alleles.
Conclusions
Our results demonstrate the usefulness of LAMP for the diagnosis of CRPA and CRAB.
Keywords: Loop-mediated isothermal amplification (LAMP), Carbapenem resistance, blaVIM-2, blaIMP-1, blaOXA-23, Pseudomonas aeruginosa, Acinetobacter baumannii
INTRODUCTION
Pseudomonas aeruginosa and Acinetobacter baumannii cause a wide range of opportunistic infections and nosocomial outbreaks with high mortality [1,2,3]. Nosocomial infections are usually caused by multidrug-resistant P. aeruginosa (MRPA) and A. baumannii (MRAB), and their multidrug resistance makes curing infected patients very difficult. Although carbapenems are the last line of treatment for infections caused by multidrug-resistant bacteria, some organisms can acquire carbapenem resistance. Based on recent reports, carbapenemase-producing clinical isolates of P. aeruginosa and A. baumannii have been steadily reported around the world. Even in Korea, healthcare-associated infections caused by multidrug-resistant pathogens have become a significant clinical concern [4,5,6,7,8]. Following this trend, since 2012, six multidrug-resistant organisms were designated as the source of healthcare-associated infections by the Korea Centers for Disease Control and Prevention, and MRPA and MRAB strains associated with carbapenem resistance were included among these six strains [9].
Carbapenem resistance is induced by the expression of carbapenemase. Carbapenemases are versatile β-lactamases that have the ability to hydrolyze penicillins, cephalosporins, monobactams. Carbapenems exhibit a broader spectrum of antibacterial activity towards Gram-positive and Gram-negative bacteria than other β-lactams [10]. To detect carbapenemase-producing P. aeruginosa and A. baumannii strains, blaVIM-2, blaIMP-1, and blaOXA-23 were chosen as target genes. blaVIM-2 and blaIMP-1 encode class B metallo-β-lactamases, and these genes have been detected in P. aeruginosa and A. baumannii. In contrast, blaOXA-23, which encodes a carbapenemase belonging to the class D β-lactamases, has been detected only in A. baumannii [1]. Testing for blaVIM-2, blaIMP-1, and blaOXA-23 is important for the timely detection and treatment of carbapenem-resistant P. aeruginosa (CRPA) and A. baumannii (CRAB) infections.
Routine diagnostic methods for the detection of CRPA and CRAB have mainly included antibiotic sensitivity testing (e.g., modified Hodge test [MHT]), carbapenemase-inhibition testing (e.g., double disk synergy test [DDST]), and the use of automated instruments to determine the minimum inhibitory concentration (MIC) of antibiotics (e.g., Vitek, MicroScan, and Phoenix) [11,12,13,14]. However, most routine diagnostic methods are culture-based assays that require 16-24 hr to complete. This causes a delay in diagnosis and subsequent effective treatment of infected patients.
Loop-mediated isothermal amplification (LAMP) has been widely used in microbial diagnoses for several reasons, including its simple methodology, short reaction time, and naked-eye detection of positive reactions [15]. In this study, we explored whether or not LAMP could be used for fast and convenient detection of CRPA or CRAB in clinical specimens. To this end, we optimized the reaction conditions for LAMP-based detection of blaVIM-2, blaIMP-1, and blaOXA-23 alleles. The sensitivity and specificity of the current LAMP protocol were superior to those of conventional PCR methods.
METHODS
1. Bacterial strains and DNA extraction
All the bacterial strains used in this study were clinical isolates obtained from the Department of Laboratory Medicine, Yonsei University College of Medicine, Seoul, Korea. The positive control DNAs used in the optimization test were extracted from carbapenem-resistant strains that were confirmed to possess carbapenem-resistance genes by PCR. DNA from sputum isolates of CRPA (n=100) and carbapenem-susceptible P. aeruginosa (CSPA, n=20) and those of CRAB (n=75) and carbapenem-susceptible A. baumannii (CSAB, n=24) were recovered by using the Geno-Auto Bacterial DNA Prep Kit (Genolution, Seoul, Korea, Catalogue No. RE1702). This kit includes unique magnetic beads with a DNA binding affinity that varies according to the pH of the buffer used. The magnetic beads bind DNA in low pH buffer and release DNA in high pH buffer. After setup, the time required to extract the DNA is 15-30 min, and the yield from these clinical specimens was quite high.
2. PCR assay for blaVIM-2, blaIMP-1, and blaOXA-23 alleles
Before the LAMP assay, the presence of carbapenemase-producing genes in isolates was confirmed by PCR. Primers for blaVIM-2, blaIMP-1, and blaOXA-23 LAMP reactions were designed based on the NCBI database sequences of blaVIM-2 (801 bp, GenBank ID: NC_020452.1), blaIMP-1 (741 bp, GenBank ID: AP012280.1), and blaOXA-23 (822 bp, GenBank ID: GQ861438.1). PCR of blaVIM-2 and blaIMP-1 was performed by using the respective outer LAMP primers (Table 1). PCR primers for blaOXA-23 were generated by Primer3 software (http://bioinfo.ut.ee/primer3/) by using custom designed primer sets. All PCR primers were designed in house, including the annealing sites of the LAMP primers. The primers used in the study are shown in Table 1. PCR reaction was performed as previously described [15].
Table 1. Primers used in this study.
| Target gent | Primer | Sequence (5' to 3') | |
|---|---|---|---|
| LAMP | blaVIM-2 | FIP | 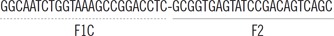 |
| BIP |  |
||
| F3* | CATGGCTATTGCGAGTCCGCTCG | ||
| B3* | GCCGCTGTGTTTTTCGCACCCCA | ||
| blaIMP-1 | FIP |  |
|
| BIP | 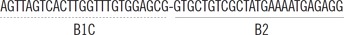 |
||
| F3* | TGTTTTGCAGCATTGCTACCGCAGC | ||
| B3* | CGAGAATTAAGCCACTCTATTCCGC | ||
| blaOXA-23 | FIP | 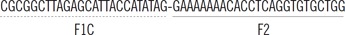 |
|
| BIP | 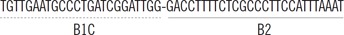 |
||
| F3 | AACCCCGAGTCAGATTGTTCAAGG | ||
| B3 | GCTTCATGGCTTCTCCTAGTGTC | ||
| PCR | blaOXA-23 | F | AAATGAAACCCCGAGTCAGA |
| R | CCCAACCAGTCTTTCCAAAA |
*The LAMP outer primers F3 and B3 were used in the PCRs for both blaVIM-2 and blaIMP-1.
Abbreviations: LAMP, loop-mediated isothermal amplification; FIP, forward inner primer; BIP, backward inner primer; F, forward; R, reverse.
3. LAMP reactions for the detection of blaVIM-2, blaIMP-1, and blaOXA-23 genes
The target gene sequences used to design the LAMP primers were identical to that used to design the PCR primers. However, the LAMP primers were different from the PCR primers. For the LAMP reactions, two sets of primers, inner and outer, are required. Although loop primers can also be included to shorten the reaction time, in this study, only inner and outer primers were used. The LAMP primer design method was identical to that described in a previous study [15]. Fig. 1 shows the DNA sequences of blaVIM-2, blaIMP-1, and blaOXA-23 and the locations of both the forward and backward inner and outer primers. The primer sets (shown in Table 1) amplified 147 bp (89-235), 187 bp (116-302), and 191 bp (136-326) target sequences of blaVIM-2, blaIMP-1, and blaOXA-23 between the F2 and B2 regions, respectively (Fig. 1). The template for the LAMP reactions was genomic DNA that contains blaVIM-2, blaIMP-1, or blaOXA-23.
Fig. 1. Primers designed for blaVIM-2, blaIMP-1, and blaOXA-23 loop-mediated isothermal amplification (LAMP) assays. Nucleotide sequences of blaVIM-2 (A), blaIMP-1 (B), and blaOXA-23 (C) and the location of LAMP primers. The forward and backward inner primers are F1c-F2 and B1c-B2 sequences, respectively. The forward and backward outer primers are F3 and B3, respectively.

To determine the most efficient conditions for the blaVIM-2, blaIMP-1, and blaOXA-23 LAMP reactions, a range of reaction temperatures (57-68℃) was tested. To determine the optimal concentration of MgSO4, 10× LAMP buffer was made as described previously [15] containing various concentrations of MgSO4 (Sigma-Aldrich Co., St. Louis, MO, USA). The 10× LAMP buffer with MgSO4 was diluted to 1× before use. The optimized reaction buffer was also used to determine the lowest template DNA concentration that yielded detectable amplification products. Formation of a white precipitate (magnesium pyrophosphate [Mg2P2O7]) after centrifuging the PCR tubes at 15,328g for 2 min was a criterion for a positive LAMP reaction. A positive reaction was confirmed by running the reaction products on a 1.5% agarose gel. All LAMP experiments were performed in triplicate.
RESULTS
1. Optimization of the blaVIM-2, blaIMP-1, and blaOXA-23 LAMP reactions
To determine the optimal reaction temperature, LAMP reactions were performed at a range of temperatures from 57℃ to 68℃ for 1 hr. A distinct ladder-like band pattern, an indication of a successful LAMP amplification, was observed in all LAMP reactions (Fig. 2A-C). Under our experimental conditions, positive LAMP reactions for blaVIM-2 and blaIMP-1 were observed at 57-63℃, whereas for blaOXA-23, positive LAMP reactions were observed at 57-62℃.
Fig. 2. Optimization of the reaction temperature and MgSO4 concentration for the blaVIM-2, blaIMP-1, and blaOXA-23 loop-mediated isothermal amplification (LAMP) assays. (A-C) Ten ng of DNA was used as a template, and the reaction was performed for 1 hr at the temperatures indicated at the top. The 'DW' reaction was performed at 62℃. (D-F) Ten ng DNA was used as a template, and the reaction was performed for 1 hr with MgSO4 concentrations indicated at the top. LAMP reaction was performed at 62℃. 'DW' means distilled water used in replace of the template DNA as a negative control. The reaction products were loaded onto 1.5% agarose gel for analysis.

The optimal MgSO4 concentration for each reaction was also determined. At 63℃, the best amplification was observed at 2-5 mM MgSO4 for blaVIM-2 and at 2-3mM MgSO4 for blaIMP-1 (Fig. 2D and E). The best amplification of blaOXA-23 was observed at 62℃ and 1-2mM MgSO4 (Fig. 2F). These results indicated that the optimal MgSO4 concentration using our primers was 2mM for all reactions and that the optimal reaction temperatures were 63℃ for blaVIM-2 and blaIMP-1 and 62℃ for blaOXA-23.
2. Sensitivity of the LAMP reaction
To assess the limit of detection (LOD) for each LAMP reaction, LAMP assays were performed by using a series of DNA template samples prepared from 10-fold serial dilutions of a stock with an initial concentration of 10 ng/µL. A typical ladder-like pattern was detected by using as little as 1 pg of target DNA for blaVIM-2 and 10 pg of target DNA for blaIMP-1 and blaOXA-23 (Fig. 3A-C). We then compared the LOD for each target gene in conventional PCR.
Fig. 3. Determination of the detection limit of the loop-mediated isothermal amplification (LAMP) assays and the minimum reaction time. (A-C) Various amounts of template DNA (1 fg, 10 fg, 100 fg, 1 pg, 10 pg, 100 pg, 1 ng, or 10 ng) were used in the LAMP reactions. The amplification was performed with 2mM MgSO4 at 62℃ for 1 hr. (D-F) LAMP reactions were performed for a range of reaction times. After the reaction time indicated on top of the gels, the LAMP reaction was terminated by inactivating Bst DNA polymerase by incubation at 95℃ for 3 min. The reaction products were loaded onto a 1.5% agarose gel for analysis.

As shown in Fig. 3A, 1 ng of DNA was required for PCR amplification of blaVIM-2. This indicates that for blaVIM-2, using our primer sets, a positive LAMP reaction could be obtained with 1,000-fold less DNA template. Likewise, for blaIMP-1 and blaOXA-23, 10-fold or 100-fold less DNA template was required to obtain a positive LAMP reaction (Fig. 3B and C).
Next, we assessed how fast a LAMP reaction can yield products that were detectable by the naked eye when using 10 ng of DNA template. Under our reaction conditions, a positive LAMP reaction for blaVIM-2 was achieved after a 20-min reaction (Fig. 3D). Importantly, magnesium pyrophosphate (Mg2P2O7) precipitate was clearly visible after a 25-min reaction (data not shown). For blaIMP-1 and blaOXA-23, positive reactions were detectable after 35 min and 40 min, respectively (Fig. 3E and F). This finding is consistent with the sensitivity result shown in Fig. 3C. Among the three LAMP targets, blaVIM-2 was detectable with the smallest amount of template DNA.
3. Application of blaVIM-2, blaIMP-1, and blaOXA-23 LAMP reactions for efficient detection of CRPA or CRAB in clinical isolates
The optimized blaVIM-2, blaIMP-1, and blaOXA-23 LAMP reactions were evaluated by using clinical isolates. Of the 120 sputum isolates of P. aeruginosa tested, 100 were determined to be CRPA based on their growth capabilities, while the other 20 isolates were determined to be CSPA. Sputum isolates of A. baumannii (N=99) were also tested; 75 were determined to be CRAB based on their growth capabilities, while the other 24 strains were determined to be CSAB. DNA extracted by a magnetic bead-based method was used as a template. Among the 100 CRPA strains, only one contained blaIMP-1, and none of them contained blaVIM-2 (Table 2). This result clearly suggests that in the P. aeruginosa clinical isolates tested in the current study, carbapenem resistance is not mediated by the presence of blaVIM-2 or blaIMP-1.
Table 2. Detection of blaVIM-2 and blaIMP-1 by conventional PCR and LAMP assays in clinical isolates of Pseudomonas aeruginosa.
| Strain (N) |
blaVIM-2 PCR |
blaVIM-2 LAMP |
blaIMP-1 PCR |
blaIMP-1 LAMP |
||||
|---|---|---|---|---|---|---|---|---|
| + | - | + | - | |||||
| CRPA (100) |
+ | 0 | 0 | 0 | + | 1 | 1 | 0 |
| - | 100 | 0 | 100 | - | 99 | 0 | 99 | |
| CSPA (20) |
+ | 0 | 0 | 0 | + | 0 | 0 | 0 |
| - | 20 | 0 | 20 | - | 20 | 0 | 20 | |
+, positive reaction; -, negative reaction. P. aeruginosa clinical isolates were not tested for the presence of blaOXA-23.
Abbreviations: CRPA, carbapenem-resistant Pseudomonas aeruginosa; CSPA, carbapenem-susceptible Pseudomonas aeruginosa.
In addition, neither PCR nor LAMP reactions for blaVIM-2 and blaIMP-1 using the 75 CRAB and 24 CSAB isolates as templates produced amplicons. In contrast, a large number of A. baumannii clinical isolates were found to have blaOXA-23, indicating that blaOXA-23 is a major genetic determinant of carbapenem resistance in A. baumannii [16]. Among the 75 CRAB sputum specimens, 52 were positive for the presence of blaOXA-23 by both PCR and LAMP (Table 3). There were 23 CRAB clinical specimens that were negative for blaOXA-23 by PCR. Of these 23 PCR-negative CRAB specimens, two had blaOXA-23 by LAMP. This result supports the notion that LAMP detection of antibiotic resistance genes is more sensitive than conventional PCR. Among the 24 CSAB specimens, 13 were positive for blaOXA-23 by PCR and LAMP, demonstrating that the presence of blaOXA-23 would not always result in carbapenem resistance. In general, the results from the blaVIM-2, blaIMP-1, and blaOXA-23 PCR and LAMP assays agreed with each other, although the LAMP assay for blaOXA-23 was more sensitive than the PCR assay.
Table 3. Detection of blaVIM-2, blaIMP-1, and blaOXA-23 by conventional PCR and LAMP assays in clinical isolates of A. baumannii.
| Strain (N) |
blaVIM-2 PCR |
blaVIM-2 LAMP |
blaIMP-1 PCR |
blaIMP-1 LAMP |
blaOXA-23 PCR |
blaOXA-23 LAMP |
||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| + | - | + | - | + | - | |||||||
| CRPA (75) |
+ | 0 | 0 | 0 | + | 0 | 0 | 0 | + | 52 | 52 | 0 |
| - | 75 | 0 | 75 | - | 75 | 0 | 75 | - | 23 | 2 | 21 | |
| CSPA (24) |
+ | 0 | 0 | 0 | + | 0 | 0 | 0 | + | 13 | 13 | 0 |
| - | 24 | 0 | 24 | - | 24 | 0 | 24 | - | 11 | 2 | 9 | |
+, positive reaction; -, negative reaction.
Abbreviations: CRAB, carbapenem-resistant A. baumannii; CSAB, carbapenem-susceptible A. baumannii.
DISCUSSION
Carbapenemase-producing strains are a worldwide concern, as some of these strains have acquired broad spectrum antibiotic resistance. In Korea, carbapenem-resistant bacteria frequently cause nosocomial infections, especially in intensive care units (ICUs). Among the deaths caused by infection, 54.2% were due to nosocomial infection [17]. Infections caused by multidrug-resistant bacteria prolong the length of hospital stays and increase the medical expenses of immunocompromised patients in the ICU. To timely prescribe proper antibiotics for the treatment of multidrug-resistant infections, rapid and accurate diagnostic methods are required.
Growing a culture of the infecting bacteria must precede the routine diagnostic methods currently used to detect carbapenem-resistant organisms. For phenotype testing using antibiotic sensitivity, the two main methods are MHT and DDST [11, 13]. Additionally, various automated instruments that measure the MIC of resistant bacteria are used as diagnostic tests. Typically, the Vitek, MicroScan, and Phoenix are used [14]. Phenotype testing and automated instruments require cultured bacteria; therefore, the time required to complete these tests is usually 16-20 hr. Although culture-dependent diagnostic tests are easy to perform, they are time-consuming and often require additional confirmation tests [18].
Molecular diagnostic methods are used when conventional culture-based methods are not sensitive enough, cost too much money, or take too long to complete. The most common molecular diagnostic method is PCR. PCR-based diagnosis has also been used due to its ability to detect specific target genes. However, quantitative measurements of target genes are not always straightforward in PCR analyses, and the presence of the amplified products has to be confirmed by gel electrophoresis.
Rapid and reliable detection of CRPA and CRAB is very important in clinical environments. It is of note that Mg2+ concentration optimal for each LAMP reaction was different. Furthermore, amplification of blaVIM-2 was achieved with only 20 min reaction time, while 35 min and 40 min reactions were required to detect amplifications of blaIMP-1 and blaOXA-23, respectively. These results suggest that our reaction conditions are better optimized, compared with those described previously [19]. Based on our protocols, the minimum amount of DNA template for successful LAMP reaction was 100-1,000 fold less than that for PCR. This demonstrates that LAMP is certainly a superb diagnostic method, when fast and sensitive decision is necessary.
Our results using clinical isolates showed that carbapenem resistance in P. aeruginosa and A. baumannii is not related to the presence of blaVIM-2 or blaIMP-1. Lee et al. [20] reported that the prevalence of metallo-β-lactamase-producing P. aeruginosa decreased from 9% to 3% during the period between 2008 and 2012. Likewise, infections caused by metallo-β-lactamase-producing Acinetobacter spp. also decreased from 20% to 2% during the period between 2005 and 2012 [20]. These findings suggest that carbapenem resistance of P. aeruginosa and Acinetobacter spp. is likely mediated by other mechanisms that may involve activation of AmpC-type, OXA-type, or extended spectrum β-lactamases with porin loss [21, 22].
In this study, we present LAMP as a useful molecular diagnosis assay for CRPA and CRAB. For rapid detection, DNA samples were extracted from sputum specimens by using a magnetic bead-based method, which took less than 30 min. Then, the LAMP assay was immediately performed, and a positive reaction was produced within 60 min. Our results also demonstrated the usefulness of LAMP for detecting CRPA and CRAB infections in clinical specimens by targeting the blaVIM-2, blaIMP-1, and blaOXA-23 genes, which are critical for carbapenem resistance.
Acknowledgments
This work was supported by a grant from the Korea Healthcare Technology R&D Project, of the Ministry for Health, Welfare, and Family Affairs (HI12C1251-00013).
Footnotes
Authors' Disclosures of Potential Conflicts of Interest: No potential conflicts of interest relevant to this article were reported.
References
- 1.Poirel L, Nordmann P. Carbapenem resistance in Acinetobacter baumannii: mechanisms and epidemiology. Clin Microbiol Infect. 2006;12:826–836. doi: 10.1111/j.1469-0691.2006.01456.x. [DOI] [PubMed] [Google Scholar]
- 2.Sevillano E, Valderrey C, Canduela MJ, Umaran A, Calvo F, Gallego L. Resistance to antibiotics in clinical isolates of Pseudomonas aeruginosa. Pathol Biol (Paris) 2006;54:493–497. doi: 10.1016/j.patbio.2006.07.030. [DOI] [PubMed] [Google Scholar]
- 3.Oh EJ, Lee S, Park YJ, Park JJ, Park K, Kim SI, et al. Prevalence of metallo-beta-lactamase among Pseudomonas aeruginosa and Acinetobacter baumannii in a Korean university hospital and comparison of screening methods for detecting metallo-beta-lactamase. J Microbiol Methods. 2003;54:411–418. doi: 10.1016/s0167-7012(03)00090-3. [DOI] [PubMed] [Google Scholar]
- 4.Jeong SH, Bae IK, Park KO, An YJ, Sohn SG, Jang SJ, et al. Outbreaks of imipenem-resistant Acinetobacter baumannii producing carbapenemases in Korea. J Microbiol. 2006;44:423–431. [PubMed] [Google Scholar]
- 5.Lee K, Park AJ, Kim MY, Lee HJ, Cho JH, Kang JO, et al. Metallo-beta-lactamase-producing Pseudomonas spp. in Korea: high prevalence of isolates with VIM-2 type and emergence of isolates with IMP-1 type. Yonsei Med J. 2009;50:335–339. doi: 10.3349/ymj.2009.50.3.335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Yong D, Choi YS, Roh KH, Kim CK, Park YH, Yum JH, et al. Increasing prevalence and diversity of metallo-beta-lactamases in Pseudomonas spp., Acinetobacter spp., and Enterobacteriaceae from Korea. Antimicrob Agents Chemother. 2006;50:1884–1886. doi: 10.1128/AAC.50.5.1884-1886.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sung JY, Koo SH, Cho HH, Kwon KC. Dissemination of an AbaR-type resistance island in multidrug-resistant Acinetobacter baumannii global clone 2 in Daejeon of Korea. Ann Clin Microbiol. 2013;16:75–80. [Google Scholar]
- 8.Chung H-S, Lee Y, Park ES, Lee DS, Ha EJ, Kim M, et al. Characterization of the multidrug-resistant Acinetobacter species causing a nosocomial outbreak at intensive care units in a Korean teaching hospital: suggesting the correlations with the clinical and environmental samples, including respiratory tract-related instruments. Ann Clin Microbiol. 2014;17:29–34. [Google Scholar]
- 9.Lee J. Status on sentinel surveillance system of multidrug-resistant organisms in Korea. [Updated in 2012]. http://www.cdc.go.kr/CDC/info/CdcKrInfo0301.jsp?menuIds=HOME001-MNU1132-MNU1138-MNU0037-MNU1380&cid=12713.
- 10.Queenan AM, Bush K. Carbapenemases: the versatile beta-lactamases. Clin Microbiol Rev. 2007;20:440–458. doi: 10.1128/CMR.00001-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lee K, Chong Y, Shin HB, Kim YA, Yong D, Yum JH. Modified Hodge and EDTA-disk synergy tests to screen metallo-beta-lactamase-producing strains of Pseudomonas and Acinetobacter species. Clin Microbiol Infect. 2001;7:88–91. doi: 10.1046/j.1469-0691.2001.00204.x. [DOI] [PubMed] [Google Scholar]
- 12.Lee K, Lim YS, Yong D, Yum JH, Chong Y. Evaluation of the Hodge test and the imipenem-EDTA double-disk synergy test for differentiating metallo-beta-lactamase-producing isolates of Pseudomonas spp. and Acinetobacter spp. J Clin Microbiol. 2003;41:4623–4629. doi: 10.1128/JCM.41.10.4623-4629.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Amjad A, Mirza I, Abbasi S, Farwa U, Malik N, Zia F. Modified Hodge test: A simple and effective test for detection of carbapenemase production. Iran J Microbiol. 2011;3:189–193. [PMC free article] [PubMed] [Google Scholar]
- 14.Markelz AE, Mende K, Murray CK, Yu X, Zera WC, Hospenthal DR, et al. Carbapenem susceptibility testing errors using three automated systems, disk diffusion, Etest, and broth microdilution and carbapenem resistance genes in isolates of Acinetobacter baumannii-calcoaceticus complex. Antimicrob Agents Chemother. 2011;55:4707–4711. doi: 10.1128/AAC.00112-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kim HJ, Kim YJ, Yong DE, Lee K, Park JH, Lee JM, et al. Loop-mediated isothermal amplification of vanA gene enables a rapid and naked-eye detection of vancomycin-resistant enterococci infection. J Microbiol Methods. 2014;104:61–66. doi: 10.1016/j.mimet.2014.05.021. [DOI] [PubMed] [Google Scholar]
- 16.Labarca JA, Salles MJ, Seas C, Guzmán-Blanco M. Carbapenem resistance in Pseudomonas aeruginosa and Acinetobacter baumannii in the nosocomial setting in Latin America. Crit Rev Microbiol. 2014:1–17. doi: 10.3109/1040841X.2014.940494. [DOI] [PubMed] [Google Scholar]
- 17.Kim BM, Jeon EJ, Jang JY, Chung JW, Park J, Choi JC, et al. Four year trend of carbapenem-resistance in newly opened ICUs of a university-affiliated hospital of South Korea. Tuberc Respir Dis (Seoul) 2012;72:360–366. doi: 10.4046/trd.2012.72.4.360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Tripathi A, Shukla SK, Singh A, Prasad KN. A new approach of real time polymerase chain reaction in detection of vancomycin-resistant enterococci and its comparison with other methods. Indian J Med Microbiol. 2013;31:47–52. doi: 10.4103/0255-0857.108721. [DOI] [PubMed] [Google Scholar]
- 19.Kojima T, Shibata N, Ikedo M, Arakawa Y. Development and evaluation of novel loop-mediated isothermal amplification for rapid detection of bla(IMP-1) and bla(VIM-2) genes. Kansenshogaku Zasshi. 2006;80:405–412. doi: 10.11150/kansenshogakuzasshi1970.80.405. [DOI] [PubMed] [Google Scholar]
- 20.Lee Y, Kim C-K, Chung H-S, Yong D, Jeong SH, Lee K, et al. Increasing carbapenem-resistant gram-negative bacilli and decreasing metallo-β-lactamase producers over eight years from Korea. Yonsei Med J. 2015;56:572–577. doi: 10.3349/ymj.2015.56.2.572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Rodríguez-Martínez JM, Poirel L, Nordmann P. Molecular epidemiology and mechanisms of carbapenem resistance in Pseudomonas aeruginosa. Antimicrob Agents Chemother. 2009;53:4783–4788. doi: 10.1128/AAC.00574-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Quale J, Bratu S, Gupta J, Landman D. Interplay of efflux system, ampC, and oprD expression in carbapenem resistance of Pseudomonas aeruginosa clinical isolates. Antimicrob Agents Chemother. 2006;50:1633–1641. doi: 10.1128/AAC.50.5.1633-1641.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]