

Abstract
The incidence of esophageal cancer is rising, mostly because the increasing incidence of esophageal adenocarcinoma in Western countries. Despite improvements in diagnosis and treatment, the overall 5-year survival rates remain low. MicroRNAs (miRNAs) are small non-coding RNA molecules that regulate the expression of target genes. Recently, disease specific miRNAs have been identified, which act as tumor suppressors or oncogenes. In this review, we will summarize the current knowledge about the function of aberrantly expressed miRNAs in esophageal cancer. We selected 5 miRNAs (miRNA-21, -143, -145, -196a and let-7) based on the available literature, and described their potential role in regulating pathways that are deregulated in esophageal cancer. Finally we will highlight the current achievements of using and targeting miRNAs. Because these miRNAs likely have important regulatory roles in cancer development, they open a therapeutic window for new treatment modalities.
Keywords: Esophageal cancer, Esophageal squamous cell carcinoma, Esophageal adenocarcinoma, MicroRNAs, Target genes
Core tip: MicroRNAs (miRNAs) likely have important regulatory roles in the development of cancer as they target essential pathways. Here, we review the function of aberrantly expressed miRNAs in esophageal cancer, with the aim to provide guidelines for future studies focusing on the function of miRNAs and the development of new treatment modalities.
INTRODUCTION
Esophageal cancer is the 8th most common cancer and the 6th most lethal cancer worldwide[1]. Esophageal cancer is divided into two main histopathological subtypes; esophageal squamous cell carcinoma (ESCC) and esophageal adenocarcinoma (EAC). ESCC develops in esophageal squamous cells while EAC develops in intestinal type epithelium containing goblet cells, which is also known as Barrett’s esophagus (BE). Both cancer types are different in development, etiology and treatment. Due to fact that most patients have already distant metastases at the time of diagnosis, the prognosis of esophageal cancer remains poor. Despite improvements in diagnosis and treatment, the overall 5-year survival rate is 15%-20%, and only after treatment with a curative intent, the survival rate increases to 47%[2].
MicroRNAs (miRNAs) are small non-coding RNA molecules of 20-24 nucleotides long that modulate the expression of target genes. They are first transcribed from DNA as parts of longer molecules (pre-miRNA) and undergo final processing by dicer in the cytoplasm to form mature miRNAs[3]. miRNAs are present in tissue, blood and other body fluids and have emerged as critical components of complex functional pathways involved in processes such as differentiation, apoptosis and proliferation[4]. Recently, numerous studies have identified tissue or disease specific miRNAs by extensive miRNA-profiling. These studies have identified miRNAs which are aberrantly expressed in esophageal cancer for example miR-21, -145, -192, which are upregulated and miRNA-31, -203, -205 and let-7 which are often downregulated compared to normal esophageal tissue[5,6]. These findings have led to the identification of miRNAs that act as tumor suppressors or oncogenes. In addition, several studies have shown that aberrant expression of specific miRNAs correlates with the survival of patients with esophageal cancer, presence of metastasis and response to neo-adjuvant therapies[5]. However, to date little is known about the cellular function of these differently expressed miRNAs.
In this review, we will summarize the function of miRNAs in the carcinogenic process of the esophagus. Using PubMed, we identified studies that evaluated the effects of miRNAs in ESCC or EAC and selected five miRNAs, miRNA-21, -143, -145, -196a and let-7, which are known to be up- or downregulated in esophageal cancer. Moreover, we will describe the current development of novel anti-cancer therapy by targeting miRNAs.
MIRNA-21 ACTS AS AN ONCOGENE IN ESOPHAGEAL SQUAMOUS CELL CARCINOMA
MiRNA-21 is reported to be an oncogene and is highly expressed in various malignancies[6]. Next to colon and gastric cancer[7], miRNA-21 expression is also upregulated in esophageal cancer. In ESCC, miRNA-21 expression is consistently reported to be higher expressed compared to normal adjacent squamous epithelium[8-10]. Moreover, increased expression of miRNA-21 is associated with more advanced stages of ESCC[8,9]. Profiling studies of EAC have also shown upregulated miRNA-21 expression in EAC compared to normal adjacent squamous epithelium[11-13]. In addition, high miRNA-21 expression is already observed in the known precursor of EAC, BE[12,14,15]. This indicates that miRNA-21 plays an important role in the carcinogenic process that occurs in the esophagus. Several studies have focused on functional roles of miRNA-21 during cancer development of the esophagus.
In vitro studies using ESCC cell lines showed increased cell viability upon miRNA-21 precursor transfection[8,9]. This increase in cell viability could be the effect of miRNA-21 targeting the PI3K pathway. This pathway regulates various cellular processes including metabolism, proliferation and cell migration and is one of the most critical cancer-promoting pathways. In short, PI3K catalyzes the formation of PIP3, which transduces activating signals to the serine-threonine kinase AKT, which in his turn is able to phosphorylate a wide array of additional substrates that also induces proliferation and survival. Phosphatase and tensin homolog deleted on chromosome 10 (PTEN) is an antagonist of this pathway; it dephosphorylates PIP3 and subsequently inhibits activation of AKT[16]. Dysregulation of the PI3K pathway through selective mutations have been reported in numerous cancers[16].
In ESCC, Ma et al[9] observed an inverse correlation, however not statistically significant, between miRNA-21 expression and PTEN protein expression (Table 1 summarizes the function of all miRNAs described in this review). In vitro transfection experiments overexpressing miRNA-21 showed no significant effect on PTEN mRNA expression, but a downregulation of PTEN protein expression was observed. This suggests that miRNA-21 targets PTEN at a post-transcriptional level. In addition, knockdown of miRNA-21 leads to significantly upregulated PTEN expression[9,17,18]. Moreover, Huang et al[17] showed, that after PTEN downregulation, pAKT is dephosphorylated. Based on these studies, it can be concluded that miRNA-21 inhibits PTEN thereby inhibiting the PI3K pathway leading to increased proliferation and cancer cell survival.
Table 1.
MiRNAs and their targets in esophageal cancer
| miRNA | Expression profile | ESCC | EAC | Related biological function | Ref. |
| miRNA-21 | ↑ | PTEN | Cell proliferation | [8,17,18] | |
| PDCD4 | Cell migration, apoptosis | [9] | |||
| Let-7 | ↓ | HMGA2 | Cell proliferation | [25,26] | |
| IL-6 | Cytokine activity | [27] | |||
| miRNA-143 and -145 | ↓ | FSCN | FSCN | Cell migration | [29,32] |
| ERK5 | Cell proliferation, cell migration | [30] | |||
| miRNA-196a | ↑ | Annexin A1 | Apoptosis | [38,39] | |
| SPRR2C | Pseudogene | [38] | |||
| S100A9 | Cell differentiation, cell proliferation | [38] | |||
| KRT5 | Cell architecture | [38] | |||
| RAP1A | Cell proliferation, cell adhesion and cell migration | [41] |
miRNAs with corresponding expression profile (↑ = upregulated, ↓ = downregulated in esophageal cancer), described targets and related biological function in esophageal squamous cells carcinoma (ESCC) and esophageal adenocarcinoma (EAC).
Next to regulating the PI3K pathway, Liu et al[10] reported that miRNA-21 targets programmed cell death 4 (PDCD4) gene in ESCC. This is in line with other studies, which have shown that miRNA-21 directly targets PDCD4 in colorectal cancer, hepatocellular carcinoma and breast cancer[19-21]. PDCD4 is a recently discovered tumor suppressor which controls cell migration, directs apoptosis and regulates the cellular response to DNA damage. In various types of cancer tissue, PDCD4 expression is downregulated. This was also observed in ESCC, where high expression of PDCD4 was detected in normal squamous epithelium in contrast to ESCC[10,22]. In addition, in ESCC an inverse correlation of PDCD4 with miRNA-21 was reported[10]. Future studies should determine if miRNA-21 also targets PDCD4 during the carcinogenic processes that occur in the transition from BE to EAC.
LET-7 ACTS AS A TUMOR SUPPRESSOR IN ESOPHAGEAL CANCER
One of the first miRNAs discovered, the let-7 family is comprised of 12 family members and have an overlapping set of target genes[23]. In the earliest phases of embryology, the let-7 family is not expressed while expression is upregulated during later stages of development. During neoplastic progression, let-7 is often downregulated and therefore considered to be a tumor suppressor[24]. In both ESCC and EAC, downregulated expression of let-7 has been reported and downregulated expression is associated with a poor prognosis[5,6].
Overexpression of let-7 in an ESCC cell line resulted in decreased cell viability compared to cells transfected with a let-7 inhibitor[25]. Several studies have identified high mobility group AT-Hook (HMGA2) as a putative target of let-7[24]. In contrast to let-7, expression of HMGA2 is prominent during development and absent in adult human tissues. However, during cancer development HMGA2 is re-expressed and acts as an oncogene by regulating cell proliferation[24]. An in vitro study of Liu et al[25] observed decreased HMGA2 protein expression after let-7 overexpression in an ESCC cell line. However, no difference was observed in mRNA expression among the different groups suggesting that let-7 targets HMGA2 at a post-transcriptional level. Another paper from the same group confirmed with a luciferase assay that let-7 directly targets HMGA2 in the esophagus[26]. To our knowledge, no studies regarding HMGA2 expression have been performed in BE or EAC. It would be interesting to determine if downregulated expression of let-7 also results in upregulation of HGMA2 in EAC. Future studies are required to elucidate this in detail.
Besides HGMA2 as putative target of let-7, Sugimura et al[27] reported an inverse association between both let-7b/c and IL6 expression in ESCC tissue. IL-6 was identified as potential target using the Target scan database. In addition, in vitro studies have shown that let-7c overexpression significantly reduced IL-6 expression in ESCC cell lines. Moreover, expression of STAT3, a downstream target of IL-6, was also reduced after let-7c overexpression[27]. Findings by Sung et al[28] supported the results of this study as they showed that let-7 directly targets IL6 in cancer associated mesenchymal stem cells involved in prostate cancer.
miRNA-143 AND miRNA-145 JOINTLY ACT AS TUMOR SUPPRESSORS
miRNA-143 and miRNA-145 are positioned in close proximity with each other on chromosome 5 and this suggests that they have similar biological functions. The expression of miRNA-143 and miRNA-145 is downregulated in both ESCC and EAC[29-31]. Furthermore, downregulation of these miRNAs is reported to be associated with increased invasion depth and lymph node metastasis[29,30] suggesting a tumor suppressive role for these miRNAs. In vitro studies have shown that overexpression of both miR-143 and miR-145 significantly reduced cell proliferation and migration while apoptosis was increased[30-33]. In addition, overexpression of miRNA-143 decreased invasive properties of ESCC cell lines[30].
Two different groups have shown that miRNA-145 directly targets fascin homolog (FSCN). Following overexpression of miRNA-145, a significant downregulation in FSCN expression was observed[29,32]. In addition, Liu et al[29] showed that miRNA-143 also directly targets FSCN, with even a 2-fold higher inhibition efficacy compared to miRNA-145. FSCN is an important regulatory element in the maintenance and stability of filamentous actin. It organizes filamentous actin in well-ordered parallel bundles and plays a crucial role in the formation of membrane protrusions. It has been suggested that FSCN has an oncogenic role as it promotes cell motility and migration[29,32]. Knockdown of FSCN in an ESCC cell line resulted in decreased cell growth and cell invasion[32]. In addition, high expression of FSCN correlates with poor survival in ESCC[34].
Besides the effect on FSCN expression, it was also suggested that miRNA-143 targets extracellular-signal-regulated kinase 5 (ERK5) expression. This kinase is a member of the mitogen activated protein kinase family and is important for cell proliferation and angiogenesis. Some reports have shown a potential role for ERK5 in cancer progression[35]. To our knowledge, only one study investigated ERK5 expression in ESCC on protein level. After overexpression of miRNA-143 in an ESCC cell line, ERK5 expression was downregulated[30]. In bladder cancer, overexpression of both miRNA-143 and -145 reduced ERK5 expression[36]. However, further studies are necessary to evaluate if ERK5 is a direct target of miRNA-143 and -145 and if the downregulation is a downstream effect of targeting another gene in the MEK5/ERK5 pathway in the esophagus.
MIRNA-196a ACTS AS AN ONCOGENE
The miRNA-196 family contains three miRNAs: miRNA-196a-1, miRNA-196a-2 and miRNA196b. miRNA-196a-1 is located on chromosome 17 while miRNA-196a-2 is located on chromosome 12. When these miRNAs are processed to mature miRNAs, both have identical mature nucleotide sequences (miRNA196a)[37]. In BE, the precursor of EAC, the expression of miRNA-196a is already significantly upregulated compared to the normal squamous epithelium. This increased expression is also observed in EAC[38]. Further studies are needed to specifically identify whether this is miRNA-196a-1 or -196a-2 that is upregulated in BE and EAC. For ESCC, publications concerning miRNA-196 expression are limited.
Functional assays have shown that overexpression of miR-196a in EAC cells resulted in increased cell proliferation compared to control cells[39]. However, this study used SEG-1 cells, which later found to be a cell line derived from lung cancer[40]. The first study which evaluated the effect of increased miRNA-196a expression in EAC found an inverse correlation comparing the expression of Annexin A1 and miRNA-196a. As expected, miRNA-196a was upregulated while Annexin A1 was downregulated in EAC compared to normal squamous epithelium[38]. Annexin A1 is a crucial factor regulating apoptosis and suppression of Annexin is often reported in malignancies[39]. To confirm that annexin A1 was a direct target of miRNA-196a, luciferase assays were performed, showing that miRNA-196a directly targets annexin A1[39]. The same group also showed an inverse correlation between the expression of miRNA-196a and SPRR2C, S100A9 and Keratin 5. To confirm these putative targets, in vitro miRNA-196a overexpression and luciferase-based assays were performed in EAC cell lines[38].
In addition, Wang et al[41] showed in ESCC cell lines that miRNA-196a directly inhibits RAP1A. RAP1A belongs to the family of RAS-related proteins regulating a wide range of biological processes, i.e., cell proliferation, cell adhesion and cell mobility[41]. In patients with ESCC, high expression of RAP1a is associated with lymph node metastasis. Overexpression of RAP1A in an ESCC cell line resulted in increased migration and invasion. In addition knockdown of RAP1A resulted in decreased migration and invasion[41]. To our knowledge, this is the only study which have reported RAP1A as a target for miRNA-196a. It would be interesting if other groups can confirm these results and to further evaluate the effect of reduced expression of RAP1A in ESCC.
CLINICAL APPLICATIONS OF MIRNAS
Results of the studies summarized above highlight the important regulatory roles of miRNAs in mediating changes in gene expression during the development of esophageal cancer. These miRNAs are excellent candidates for the development of novel treatment modalities. Depending on the oncogenic or tumor suppressive role(s) of the specific miRNA, it may be possible to inhibit or replace its function through the use of miRNA mimics and inhibitors. However until now, the research field has mainly focused on the identification of down-stream targets of miRNAs using in vitro studies (Figure 1). Animal studies should be the next step in order to elucidate whether the manipulation of specific miRNAs could provide a new therapeutic window for esophageal cancer. In a mouse model of ESCC, knockdown of miRNA-21 reduced tumor size and weight, suggesting a potential role for miRNA-21 as therapeutic target in the treatment of ESCC[9]. This study indicates that it should be possible to translate the in vitro results to animal studies.
Figure 1.
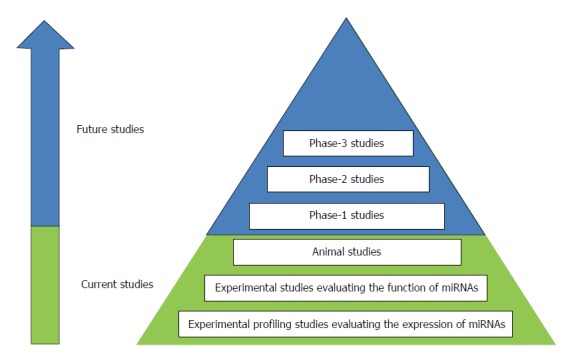
Overview of study phases for developing miRNA-based therapies for esophageal cancer. An overview of the current and future studies for developing miRNA-based therapies. The research field of esophageal cancer has mainly focused on the identification of miRNAs (profiling studies) and evaluating their specific functions by identifying the targets using in vitro experiments. Animal studies provide insight whether the manipulation of miRNAs provide new therapeutic windows. For the future, phase-1, -2 and -3 studies should be conducted in order to determine whether the manipulation of miRNAs could provide a new therapeutic window for esophageal cancer.
Recently, the first human study for the evaluation of miRNA-based therapy was published. In patients with chronic hepatitis C virus (HCV), Miravirsen, an antisense oligonucleotide which binds and thereby blocks miRNA-122, was randomly compared with placebo. Treatment with Miravirsen was found to dose-dependently reduce HCV RNA levels compared to placebo[42]. In addition, MRX34, a mimic of the tumor suppressor miRNA-34, is currently evaluated in an open-label phase 1 clinical trial in patients with unresectable liver cancer (ClinicalTrials.gov number: NCT01829971). These crucial studies will pave the way for other phase 1 studies for miRNA-based therapies.
Moreover the expression of specific miRNAs could be used in order to individualize the treatment for patients with esophageal cancer. For example, a low expression of let-7c was found to correlate with a poor response to chemotherapy[27]. Sensitivity to cisplatin, which is commonly used as chemotherapy in esophageal cancer, increased after transfection with let-7c in ESCC cell lines[27]. Furthermore, Hummel et al[43] compared the miRNA signature of chemoresistant esophageal cell lines (both EAC as ESCC) to chemotherapy sensitive controls and identified 18 miRNAs that were significantly dysregulated compared to controls. Q-RT-PCR validation of the microarray experiments has identified numerous miRNAs aberrantly expressed in cisplatin resistant esophageal cancer cell lines and 5-FU resistant esophageal cancer cell lines It would be of high interest to further investigate if these results can be translated into a clinical setting in order to modify the treatment for patients with esophageal cancer and increase disease survival.
CONCLUSION
This review describes the current knowledge regarding the function of aberrantly expressed miRNAs in esophageal cancer. Overall there is a limited number of studies published evaluating the effect of miRNAs in esophageal cancer; however, the knowledge in this field is rapidly expanding. Further studies to identify putative targets of miRNAs will improve our understanding of their function in the development and progression of esophageal cancer. This knowledge will improve the ability to utilize miRNAs clinically as therapeutic targets and/or as prognostic markers.
Footnotes
Conflict-of-interest statement: No potential conflicts of interest.
Open-Access: This article is an open-access article which was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Peer-review started: August 19, 2015
First decision: October 14, 2015
Article in press: November 9, 2015
P- Reviewer: Alshehabi Z, Goral V S- Editor: Qi Y L- Editor: A E- Editor: Wang CH
References
- 1.Ferlay J, Shin HR, Bray F, Forman D, Mathers C, Parkin DM. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int J Cancer. 2010;127:2893–2917. doi: 10.1002/ijc.25516. [DOI] [PubMed] [Google Scholar]
- 2.Dikken JL, Lemmens VE, Wouters MW, Wijnhoven BP, Siersema PD, Nieuwenhuijzen GA, van Sandick JW, Cats A, Verheij M, Coebergh JW, et al. Increased incidence and survival for oesophageal cancer but not for gastric cardia cancer in the Netherlands. Eur J Cancer. 2012;48:1624–1632. doi: 10.1016/j.ejca.2012.01.009. [DOI] [PubMed] [Google Scholar]
- 3.Huang J, Zhang SY, Gao YM, Liu YF, Liu YB, Zhao ZG, Yang K. MicroRNAs as oncogenes or tumour suppressors in oesophageal cancer: potential biomarkers and therapeutic targets. Cell Prolif. 2014;47:277–286. doi: 10.1111/cpr.12109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Calin GA, Croce CM. MicroRNA signatures in human cancers. Nat Rev Cancer. 2006;6:857–866. doi: 10.1038/nrc1997. [DOI] [PubMed] [Google Scholar]
- 5.Amin M, Lam AK. Current perspectives of mi-RNA in oesophageal adenocarcinoma: Roles in predicting carcinogenesis, progression and values in clinical management. Exp Mol Pathol. 2015;98:411–418. doi: 10.1016/j.yexmp.2015.03.002. [DOI] [PubMed] [Google Scholar]
- 6.Sakai NS, Samia-Aly E, Barbera M, Fitzgerald RC. A review of the current understanding and clinical utility of miRNAs in esophageal cancer. Semin Cancer Biol. 2013;23:512–521. doi: 10.1016/j.semcancer.2013.08.005. [DOI] [PubMed] [Google Scholar]
- 7.Volinia S, Calin GA, Liu CG, Ambs S, Cimmino A, Petrocca F, Visone R, Iorio M, Roldo C, Ferracin M, et al. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc Natl Acad Sci USA. 2006;103:2257–2261. doi: 10.1073/pnas.0510565103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Mori Y, Ishiguro H, Kuwabara Y, Kimura M, Mitsui A, Ogawa R, Katada T, Harata K, Tanaka T, Shiozaki M, et al. MicroRNA-21 induces cell proliferation and invasion in esophageal squamous cell carcinoma. Mol Med Rep. 2009;2:235–239. doi: 10.3892/mmr_00000089. [DOI] [PubMed] [Google Scholar]
- 9.Ma WJ, Lv GD, Tuersun A, Liu Q, Liu H, Zheng ST, Huang CG, Feng JG, Wang X, Lin RY, et al. Role of microRNA-21 and effect on PTEN in Kazakh’s esophageal squamous cell carcinoma. Mol Biol Rep. 2011;38:3253–3260. doi: 10.1007/s11033-010-0480-9. [DOI] [PubMed] [Google Scholar]
- 10.Liu T, Liu Q, Zheng S, Gao X, Lu M, Yang C, Dai F, Sheyhidin I, Lu X. MicroRNA-21 promotes cell growth and migration by targeting programmed cell death 4 gene in Kazakh’s esophageal squamous cell carcinoma. Dis Markers. 2014;2014:232837. doi: 10.1155/2014/232837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Chen Z, Saad R, Jia P, Peng D, Zhu S, Washington MK, Zhao Z, Xu Z, El-Rifai W. Gastric adenocarcinoma has a unique microRNA signature not present in esophageal adenocarcinoma. Cancer. 2013;119:1985–1993. doi: 10.1002/cncr.28002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Saad R, Chen Z, Zhu S, Jia P, Zhao Z, Washington MK, Belkhiri A, El-Rifai W. Deciphering the unique microRNA signature in human esophageal adenocarcinoma. PLoS One. 2013;8:e64463. doi: 10.1371/journal.pone.0064463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Mathé EA, Nguyen GH, Bowman ED, Zhao Y, Budhu A, Schetter AJ, Braun R, Reimers M, Kumamoto K, Hughes D, et al. MicroRNA expression in squamous cell carcinoma and adenocarcinoma of the esophagus: associations with survival. Clin Cancer Res. 2009;15:6192–6200. doi: 10.1158/1078-0432.CCR-09-1467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wu X, Ajani JA, Gu J, Chang DW, Tan W, Hildebrandt MA, Huang M, Wang KK, Hawk E. MicroRNA expression signatures during malignant progression from Barrett’s esophagus to esophageal adenocarcinoma. Cancer Prev Res (Phila) 2013;6:196–205. doi: 10.1158/1940-6207.CAPR-12-0276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wijnhoven BP, Hussey DJ, Watson DI, Tsykin A, Smith CM, Michael MZ. MicroRNA profiling of Barrett’s oesophagus and oesophageal adenocarcinoma. Br J Surg. 2010;97:853–861. doi: 10.1002/bjs.7000. [DOI] [PubMed] [Google Scholar]
- 16.Martini M, De Santis MC, Braccini L, Gulluni F, Hirsch E. PI3K/AKT signaling pathway and cancer: an updated review. Ann Med. 2014;46:372–383. doi: 10.3109/07853890.2014.912836. [DOI] [PubMed] [Google Scholar]
- 17.Huang S, Li XQ, Chen X, Che SM, Chen W, Zhang XZ. Inhibition of microRNA-21 increases radiosensitivity of esophageal cancer cells through phosphatase and tensin homolog deleted on chromosome 10 activation. Dis Esophagus. 2013;26:823–831. doi: 10.1111/j.1442-2050.2012.01389.x. [DOI] [PubMed] [Google Scholar]
- 18.Weng W, Wu Q, Yu Y, Mei W, Wang X. A novel chemotherapeutic arene ruthenium(II) drug Rawq01 altered the effect of microRNA-21 on PTEN/AKT signaling pathway in esophageal cancer cells. Anticancer Res. 2013;33:5407–5414. [PubMed] [Google Scholar]
- 19.Lu Z, Liu M, Stribinskis V, Klinge CM, Ramos KS, Colburn NH, Li Y. MicroRNA-21 promotes cell transformation by targeting the programmed cell death 4 gene. Oncogene. 2008;27:4373–4379. doi: 10.1038/onc.2008.72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Asangani IA, Rasheed SA, Nikolova DA, Leupold JH, Colburn NH, Post S, Allgayer H. MicroRNA-21 (miR-21) post-transcriptionally downregulates tumor suppressor Pdcd4 and stimulates invasion, intravasation and metastasis in colorectal cancer. Oncogene. 2008;27:2128–2136. doi: 10.1038/sj.onc.1210856. [DOI] [PubMed] [Google Scholar]
- 21.Qiu X, Dong S, Qiao F, Lu S, Song Y, Lao Y, Li Y, Zeng T, Hu J, Zhang L, et al. HBx-mediated miR-21 upregulation represses tumor-suppressor function of PDCD4 in hepatocellular carcinoma. Oncogene. 2013;32:3296–3305. doi: 10.1038/onc.2013.150. [DOI] [PubMed] [Google Scholar]
- 22.Fassan M, Realdon S, Pizzi M, Balistreri M, Battaglia G, Zaninotto G, Ancona E, Rugge M. Programmed cell death 4 nuclear loss and miR-21 or activated Akt overexpression in esophageal squamous cell carcinogenesis. Dis Esophagus. 2012;25:263–268. doi: 10.1111/j.1442-2050.2011.01236.x. [DOI] [PubMed] [Google Scholar]
- 23.Peter ME. Let-7 and miR-200 microRNAs: guardians against pluripotency and cancer progression. Cell Cycle. 2009;8:843–852. doi: 10.4161/cc.8.6.7907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Park SM, Shell S, Radjabi AR, Schickel R, Feig C, Boyerinas B, Dinulescu DM, Lengyel E, Peter ME. Let-7 prevents early cancer progression by suppressing expression of the embryonic gene HMGA2. Cell Cycle. 2007;6:2585–2590. doi: 10.4161/cc.6.21.4845. [DOI] [PubMed] [Google Scholar]
- 25.Liu Q, Lv GD, Qin X, Gen YH, Zheng ST, Liu T, Lu XM. Role of microRNA let-7 and effect to HMGA2 in esophageal squamous cell carcinoma. Mol Biol Rep. 2012;39:1239–1246. doi: 10.1007/s11033-011-0854-7. [DOI] [PubMed] [Google Scholar]
- 26.Liu Q, Liu T, Zheng S, Gao X, Lu M, Sheyhidin I, Lu X. HMGA2 is down-regulated by microRNA let-7 and associated with epithelial-mesenchymal transition in oesophageal squamous cell carcinomas of Kazakhs. Histopathology. 2014;65:408–417. doi: 10.1111/his.12401. [DOI] [PubMed] [Google Scholar]
- 27.Sugimura K, Miyata H, Tanaka K, Hamano R, Takahashi T, Kurokawa Y, Yamasaki M, Nakajima K, Takiguchi S, Mori M, et al. Let-7 expression is a significant determinant of response to chemotherapy through the regulation of IL-6/STAT3 pathway in esophageal squamous cell carcinoma. Clin Cancer Res. 2012;18:5144–5153. doi: 10.1158/1078-0432.CCR-12-0701. [DOI] [PubMed] [Google Scholar]
- 28.Sung SY, Liao CH, Wu HP, Hsiao WC, Wu IH, Jinpu SH, Hsieh CL. Loss of let-7 microRNA upregulates IL-6 in bone marrow-derived mesenchymal stem cells triggering a reactive stromal response to prostate cancer. PLoS One. 2013;8:e71637. doi: 10.1371/journal.pone.0071637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Liu R, Liao J, Yang M, Sheng J, Yang H, Wang Y, Pan E, Guo W, Pu Y, Kim SJ, et al. The cluster of miR-143 and miR-145 affects the risk for esophageal squamous cell carcinoma through co-regulating fascin homolog 1. PLoS One. 2012;7:e33987. doi: 10.1371/journal.pone.0033987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ni Y, Meng L, Wang L, Dong W, Shen H, Wang G, Liu Q, Du J. MicroRNA-143 functions as a tumor suppressor in human esophageal squamous cell carcinoma. Gene. 2013;517:197–204. doi: 10.1016/j.gene.2012.12.031. [DOI] [PubMed] [Google Scholar]
- 31.van Baal JW, Verbeek RE, Bus P, Fassan M, Souza RF, Rugge M, ten Kate FJ, Vleggaar FP, Siersema PD. microRNA-145 in Barrett’s oesophagus: regulating BMP4 signalling via GATA6. Gut. 2013;62:664–675. doi: 10.1136/gutjnl-2011-301061. [DOI] [PubMed] [Google Scholar]
- 32.Kano M, Seki N, Kikkawa N, Fujimura L, Hoshino I, Akutsu Y, Chiyomaru T, Enokida H, Nakagawa M, Matsubara H. miR-145, miR-133a and miR-133b: Tumor-suppressive miRNAs target FSCN1 in esophageal squamous cell carcinoma. Int J Cancer. 2010;127:2804–2814. doi: 10.1002/ijc.25284. [DOI] [PubMed] [Google Scholar]
- 33.Derouet MF, Liu G, Darling GE. MiR-145 expression accelerates esophageal adenocarcinoma progression by enhancing cell invasion and anoikis resistance. PLoS One. 2014;9:e115589. doi: 10.1371/journal.pone.0115589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Hashimoto Y, Ito T, Inoue H, Okumura T, Tanaka E, Tsunoda S, Higashiyama M, Watanabe G, Imamura M, Shimada Y. Prognostic significance of fascin overexpression in human esophageal squamous cell carcinoma. Clin Cancer Res. 2005;11:2597–2605. doi: 10.1158/1078-0432.CCR-04-1378. [DOI] [PubMed] [Google Scholar]
- 35.Nithianandarajah-Jones GN, Wilm B, Goldring CE, Müller J, Cross MJ. ERK5: structure, regulation and function. Cell Signal. 2012;24:2187–2196. doi: 10.1016/j.cellsig.2012.07.007. [DOI] [PubMed] [Google Scholar]
- 36.Noguchi S, Yasui Y, Iwasaki J, Kumazaki M, Yamada N, Naito S, Akao Y. Replacement treatment with microRNA-143 and -145 induces synergistic inhibition of the growth of human bladder cancer cells by regulating PI3K/Akt and MAPK signaling pathways. Cancer Lett. 2013;328:353–361. doi: 10.1016/j.canlet.2012.10.017. [DOI] [PubMed] [Google Scholar]
- 37.Chen C, Zhang Y, Zhang L, Weakley SM, Yao Q. MicroRNA-196: critical roles and clinical applications in development and cancer. J Cell Mol Med. 2011;15:14–23. doi: 10.1111/j.1582-4934.2010.01219.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Maru DM, Singh RR, Hannah C, Albarracin CT, Li YX, Abraham R, Romans AM, Yao H, Luthra MG, Anandasabapathy S, et al. MicroRNA-196a is a potential marker of progression during Barrett’s metaplasia-dysplasia-invasive adenocarcinoma sequence in esophagus. Am J Pathol. 2009;174:1940–1948. doi: 10.2353/ajpath.2009.080718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Luthra R, Singh RR, Luthra MG, Li YX, Hannah C, Romans AM, Barkoh BA, Chen SS, Ensor J, Maru DM, et al. MicroRNA-196a targets annexin A1: a microRNA-mediated mechanism of annexin A1 downregulation in cancers. Oncogene. 2008;27:6667–6678. doi: 10.1038/onc.2008.256. [DOI] [PubMed] [Google Scholar]
- 40.Boonstra JJ, van Marion R, Beer DG, Lin L, Chaves P, Ribeiro C, Pereira AD, Roque L, Darnton SJ, Altorki NK, et al. Verification and unmasking of widely used human esophageal adenocarcinoma cell lines. J Natl Cancer Inst. 2010;102:271–274. doi: 10.1093/jnci/djp499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wang K, Li J, Guo H, Xu X, Xiong G, Guan X, Liu B, Li J, Chen X, Yang K, et al. MiR-196a binding-site SNP regulates RAP1A expression contributing to esophageal squamous cell carcinoma risk and metastasis. Carcinogenesis. 2012;33:2147–2154. doi: 10.1093/carcin/bgs259. [DOI] [PubMed] [Google Scholar]
- 42.Janssen HL, Reesink HW, Lawitz EJ, Zeuzem S, Rodriguez-Torres M, Patel K, van der Meer AJ, Patick AK, Chen A, Zhou Y, et al. Treatment of HCV infection by targeting microRNA. N Engl J Med. 2013;368:1685–1694. doi: 10.1056/NEJMoa1209026. [DOI] [PubMed] [Google Scholar]
- 43.Hummel R, Sie C, Watson DI, Wang T, Ansar A, Michael MZ, Van der Hoek M, Haier J, Hussey DJ. MicroRNA signatures in chemotherapy resistant esophageal cancer cell lines. World J Gastroenterol. 2014;20:14904–14912. doi: 10.3748/wjg.v20.i40.14904. [DOI] [PMC free article] [PubMed] [Google Scholar]