

Abstract
Faecalibacterium prausnitzii strain A2-165 was previously reported to have anti-inflammatory properties and prevent colitis in a TNBS model. We compared the immunomodulatory properties of strain A2-165 to four different F. prausnitzii isolates and eight abundant intestinal commensals using human dendritic cells (DCs) and mouse BMDCs in vitro. Principal component analysis revealed that the cytokine response to F. prausnitzii A2-165 is distinct from the other strains in eliciting high amounts of IL-10 secretion. The mouse DNBS model of relapsing IBD was used to compare the protective effects of F. prausnitzii A2-165 and Clostridium hathewayi, a low secretor of IL-10, on the Th1-driven inflammatory response to DNBS; attenuation of disease parameters was only observed with F. prausnitzii. In an in vivo mouse model of nasal tolerance to ovalbumin, F. prausnitzii A2-165 enhanced ovalbumin-specific T cell proliferation and reduced the proportion of IFN-γ+ T cells in CLNs. Similarly, in vitro F. prausnitzii A2-165 stimulated BMDCs increased ovalbumin-specific T cell proliferation and reduced the number of IFN-γ+ T cells. These mechanisms may contribute to the anti-inflammatory effects of F. prausnitzii in colitis and support the notion that this abundant bacterium might contribute to immune homeostasis in the intestine via its anti-inflammatory properties.
The human gastrointestinal tract is colonized by several hundred different species of commensal bacteria, referred to as the microbiota, which reaches densities of 1012 bacteria per gram of luminal content in the colon1,2. Major changes in the microbiota are associated with a number of intestinal-related disorders including inflammatory bowel disease (IBD), irritable bowel syndrome, autoimmune disease, diabetes and colorectal cancer3.
In the healthy intestine, commensal bacteria are predominantly compartmentalized to the lumen, but can interact with immune cells associated with the epithelium or in mucosa-associated lymphoid tissues such as Peyer’s patches (PPs) or gut-draining mesenteric lymph nodes (MLNs). At these sites, antigen-presenting cells (APCs), such as dendritic cells (DCs), can sample microbes and food proteins and prime mucosal naive T cells driving their activation, clonal expansion and differentiation. In steady-state conditions, regulatory T cell (Treg) conversion in response to the presentation of harmless protein antigens occurs with a higher frequency in PPs and MLNs when compared to non-mucosa-draining lymphoid tissue. It has long been known that this preferential tolerance induction at mucosal sites is dependent on microbial sensing4,5.
Over the past years, it has become apparent that not all components of the microbiota are equal in terms of their impact on host physiology. For example, colonization of germ-free mice with the mouse commensal segmented filamentous bacteria induces an increase in the number of CD4+ T cells, in particular T helper 1 cells (Th1), Th17 and Foxp3+ Tregs in the small intestine and colon6. Conversely, colonization of germ-free mice with 17 Clostridia isolated from the human microbiota induced Tregs expansion and differentiation by inducing an increase in TGF-β7. Accumulation of colonic IL-10 secreting, Foxp3+ Tregs is also induced by colonization of germ-free mice with Bacteroides fragilis8. This implies that commensal bacteria may act upon mucosal T cells at different stages during their activation, differentiation as well as accumulation and maintenance in the lamina propria (LP).
The microbiota of patients with active Crohn’s disease (CD) and ulcerative colitis (UC), both forms of IBD, have increased abundance of Proteobacteria and lower faecal counts of Firmicutes compared to healthy individuals9. The Proteobacteria phylum includes pathobionts of Escherichia coli that contribute to the pathophysiology of the disease10. Among the Firmicutes, Faecalibacterium prausnitzii (Fp), one of the most abundant intestinal bacteria in humans11,12,13, is reduced in CD and UC patients14,15. Fp strain A2-165 has been shown to attenuate colitis induced by 2,4,6-trinitrobenzenesulfonic acid (TNBS) or dinitrobenzenesulfonic acid (DNBS) in mice and to induce a relatively high IL-10 to IL-12p70 cytokine ratio in human peripheral blood mononuclear cells (hPBMCs)14,16. Fp along with certain members of the Clostridium cluster XIVa are prominent butyrate producers in the intestine17. Butyrate is a primary energy source for the epithelial cells lining the colon18 and appears to have both anti-inflammatory and cancer chemopreventive activities17,18,19.
DCs and T cells are instrumental in regulating tolerance and immunity in the intestinal mucosa but the effects of Fp on these cells have not yet been described. It is not known if the Fp strain A2-165 tested in the mouse colitis models is characteristic of other Fp strains or if responses differ to other members of Clostridium cluster IV and cluster XIVa isolated from the human colon. Furthermore, the mechanisms of the protective and anti-inflammatory effects of Fp have not yet been fully unravelled.
To address these questions, we have investigated and compared the immunomodulatory properties of Fp with other commensal bacteria on DCs and T cells in vitro. The immunomodulatory and TLR signalling properties of 13 commensal strains of the Firmicutes and Actinobacteria phyla, including five isolates of Fp were first evaluated using human monocyte-derived DCs (hDCs) and TLR expressing cell lines. Further immune response studies were then performed on a smaller number of strains using mouse bone marrow-derived DCs (BMDCs). Subsequently, Fp strain A2-165 and another butyrate producing Clostridium, Clostridium hathewayi (Ch) 82-B were tested in vivo for their anti-inflammatory effects in the DNBS model of chronic relapsing colitis and on mucosal T cell differentiation using a well characterized model for nasal tolerance induction to the antigen ovalbumin (OVA).
Results
Colonic bacteria of Clostridium clusters IV and XIVa induce differential cytokine responses in hDCs
Incubation of hDCs with the commensal bacteria (Table 1) induced hDC maturation and activation and increased surface expression of the markers CD83 and CD86 when compared with the control immature non-stimulated hDCs (Supplementary Fig. S1). Incubation with the bacteria did not significantly alter hDC viability compared to the unstimulated control cells (60–80% viability, not shown).
Table 1. Phylum, family and phylogenetic cluster of the bacteria used.
| Bacterial strain | Phylum | Family | Phylogenetic cluster | Ref. |
|---|---|---|---|---|
| F. prausnitzii A2-165 | Firmicutes | Ruminococcaceae | Clostr. cluster IV | 12 |
| F. prausnitzii M21/2 | Firmicutes | Ruminococcaceae | Clostr. cluster IV | 34 |
| F. prausnitzii S3L/3 | Firmicutes | Ruminococcaceae | Clostr. cluster IV | 34 |
| F. prausnitzii L2-6 | Firmicutes | Ruminococcaceae | Clostr. cluster IV | 12 |
| F. prausnitzii HTF-F | Firmicutes | Ruminococcaceae | Clostr. cluster IV | 2 |
| R. bromii L2-63 | Firmicutes | Ruminococcaceae | Clostr. cluster IV | 13 |
| E. rectale A1-86 | Firmicutes | Lachnospiraceae | Clostr. cluster XIVa | 13 |
| E. hallii L2-7 | Firmicutes | Lachnospiraceae | Clostr. cluster XIVa | 35 |
| B. adolescentis L2-32 | Actinobacteria | Bifidobacteriaceae | 13 | |
| Megamonas sp. Mag1B 95,0% homology* with M. rupellensis | Firmicutes | Veillonellaceae | This work | |
| Clostridium sp. 82B 95,4% homology* with C. hathewayi | Firmicutes | Clostridiaceae | Clostr. cluster XIVa | This work |
| Clostridium sp. Lac1D 90,5% homology* with C. xylanovorans | Firmicutes | Clostridiaceae | Clostr. cluster XIVa | This work |
| L. plantarum WCFS1 | Firmicutes | Lactobacillaceae |
(Clostr, Clostridium); *based on the sequence homology of 850 nucleotides of the 16S rRNA.
Cytokines elicited by these strains result from the combination of TLR and NLR recognition of bacterial microbial associated molecular patterns (MAMPs) on the cell membrane as well as intra-cellularly after phagocytosis. Thus immune activation of hDCs was assessed by measuring the levels of IL-10, IL-12p70, IL-6, IL-1β and TNF-α secreted in the culture supernatant after incubation of the bacteria with hDCs from three to four different donors (Fig. 1). The largest amounts of IL-10 were induced by the Fp strains, Eubacterium hallii (Eh) L2-7 and Megamonas rupellenesis (Mr) Mag1B. In addition, these strains induced the largest amounts of TNF-α and IL-6. The Fp strains, Eh L2-7 and Eubacterium rectale (Er) A1-86 also induced relatively large amounts of IL-1β. Lactobacillus plantarum (Lp) WCFS1, Mr Mag1B and Eh L2-7 were among the most potent inducers of IL-12p70, whereas the Fp strains were among the poorer inducers (Fig. 1). Surprisingly, Ruminococcus bromii (Rb) L2-63, Ch 82-B and Clostridium xylanovorans (Cx) Lac1D induced small or undetectable amounts of some cytokines (Fig. 1) despite having effectively induced hDCs maturation (Supplementary Fig. S1). The five Fp strains had very different cytokines profiles. The highest IL-10 producer was strain A2-165, strains L2-6 and M21/2 induced high amounts of IL-10 but also IL-12p70 while strains S3L/3 and HTF-F induced intermediate to low amounts of both cytokines. The amounts of IL-10 induced by Fp strain A2-165 was 4.7 ng/ml in Donor 1, 1. 9 ng/ml in Donor 2 and 1.7 ng/ml in Donor 3. These amounts are relatively high compared to similar studies with probiotics and DCs or PBMCs20,21.
Figure 1. Cytokine secretion by hDCs. IL-10, IL-12p70, TNF-α, IL-1β and IL-6 were measured in the supernatant of hDCs from 3 to 4 donors after 48 h of incubation with the bacteria (bacterium to DC ratio 10:1, 106 DCs/well in 24w plate).

Each donor is represented with a different symbol and the red lines indicate the average of the donors.
Bacteria and hDCs were incubated in the presence of penicillin and streptomycin to prevent contaminations and bacterial overgrowth so no short-chain fatty acids or other metabolites would be produced. However, we independently tested whether the Fp A2-165 cell free culture supernatants, bacterial medium or butyrate at concentrations similar to that measured in the bacterial culture supernatant would activate hDCs. In an initial experiment, the bacterial supernatants were found to be toxic for the hDCs at a concentration of 10% v/v (data not shown). This toxicity can be attributed to the high concentration of butyrate produced during bacterial fermentation22. Fp A2-165 culture supernatant (1.25 and 2.5% v/v) and butyrate (25 to 500 μM) did not affect cell viability or cytokine secretion by hDCs compared to non-stimulated cells (data not shown).
Differential activation of TLRs by commensal bacteria
To investigate the potential relationship between cytokine profiles in hDCs and TLR signalling, bacteria were incubated with HEK293 reporter cell lines expressing different human TLRs and a reporter plasmid harbouring the luciferase gene under the control of the NF-κB promoter.
The Fp strains, Rb L2-63, Er A1-86, Eh L2-7 and Bifidobacterium adolescentis (Ba) L2-32 induced high NF-κB activation in the TLR2/6 and TLR2 reporter cell lines (Fig. 2). Agonists of TLR2 include acylated lipoprotein anchors and potentially lipoteichoic acid present in Gram-positive and Gram-negative bacterial envelopes. As previously reported23, the flagellated strain Er A1-8624 induced NF-κB activation via TLR5 signalling. Surprisingly, Eh L2-7 and Mr Mag1B activated TLR4 signalling although they are considered Gram-positive. Mr Mag1B however, has been described previously as Gram-negative to Gram-variable25. Ch 82-B and Cx Lac1D did not induce NF-κB activation in any of the TLR assays. The relative capacity of the strains to elicit NF-κB activation through TLR signalling pathways correlated with the induction of cytokines in hDCs.
Figure 2. TLR signalling properties of the commensal bacteria. NF-κB activation was measured using a luminescence reporter in HEK293 cell lines expressing TLR2, TLR2/6, TLR5 and TLR4 after incubation with the bacteria (bacterium: cell ratio, 10:1).

NF-κB activation is expressed as percentage of the positive control. Error bars represent SEM, n = 6, *indicates p < 0.05, **p < 0.01, ***p < 0.001 compared to the control.
Principal component analysis
Principal component analysis (PCA) was performed to describe the variance of the cytokine levels (IL-10, IL-12p70, IL-6, IL-1β and TNF-α) induced by the bacteria after incubation with hDCs. The PC 1 and PC 2 of the PCA describe 99.9% of the variance between the cytokine levels induced by the bacteria. The coefficients of PC 1 revealed that the contribution of the different cytokines to the variance was almost equal, the coefficient associated with IL-12p70 was 0.2 while for the other cytokines the coefficient was in average 0.48. The PC 1, PC 2 score plot (Fig. 3) revealed that the majority of the bacteria are grouped in the centre of the plot, while Fp A2-165, Eh L2-7, Mr Mag1B and Lp WCFS1 are clearly separated from the other bacteria and from each other reflecting their capacity to induce distinct cytokine profiles in hDCs. Furthermore, PCA revealed that the immune response induced by Fp A2-165 is very different from the other Fp strains tested.
Figure 3. Principal component analysis (PCA) of the cytokine profiles induced by the commensal bacteria in hDCs.
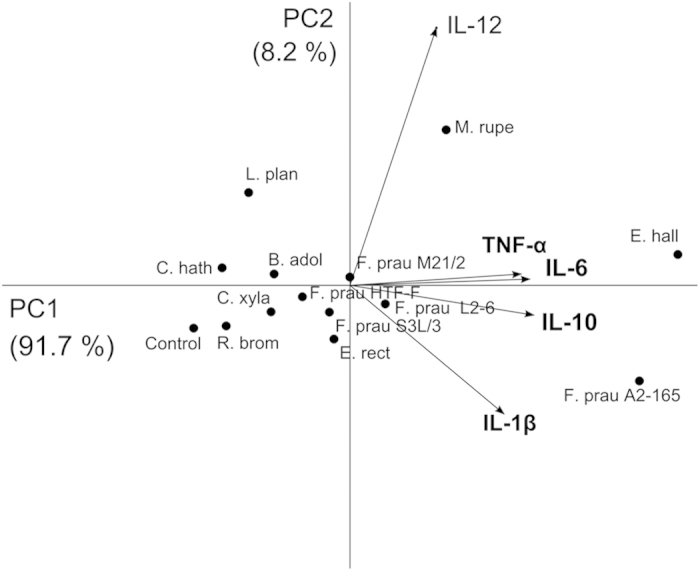
Each dot represents the average of cytokine levels of a commensal strain in hDCs. The variables (level of cytokines) are represented by vectors. The direction and length of a vector indicates how the variable contributes to the two principal components in the plot. (F. prau, F. prausnitzii; R. brom, R. bromii L2-63; E. rect, E. rectale A1-86; E. hall, E. halli L2-7; M. rupe, M. rupellensis Mag1B; C. hath, C. hathewayi 82-B; C. xyla, C. xylanovorans Lac1D; L. plan, L. plantarum WCFS1).
Mouse BMDC cytokine responses to commensal bacteria
In order to select a strain to compare with Fp A2-165 in subsequent in vivo experiments, the immune response of mouse BMDCs incubated with Fp A2-165 and five other human commensal bacterial strains was measured. The candidate strains were chosen on the basis of cytokines produced in culture with hDCs. Fp A2-165 was the highest inducer of IL-10 release by hDCs, Ch 82-B and Cx Lac1D induced low or undetectable cytokine responses, Lp WCFS1 was one of the highest inducers of IL-12p70 release and Er A1-86 and Ba L2-32 induced intermediate levels of cytokines compared to the other strains (Fig. 1).
All the strains induced increased surface expression of the co-stimulatory molecules CD86 and CD40 by BMDCs compared to untreated BMDCs (Supplementary Fig. S2). Culture of BMDCs with Fp A2-165 induced the largest amount of IL-10 release, followed by Er A1-86 and Ba L2-32. IL-12p70 release by BMDCs was mostly induced by Fp A2-165 and Lp WCFS1. However, for all the strains tested the levels of IL-12p70 were around 10 fold lower than those obtained using hDCs (Fig. 4). The amounts of IFN-γ in the culture supernatants of stimulated BMDCs were low but high levels of TNF release were induced by virtually all bacterial strains with Lp WCFS1 and Fp A2-165 being the strongest inducers while Ch 82-B and Cx Lac1D were low to weak inducers. Ch 82-B was selected for comparison to Fp A2-165 in subsequent experiments in vitro and in vivo as it is capable of activating immune cells but a weak inducer of IL-10 and other cytokines.
Figure 4. Cytokine secretion by mouse BMDCs. IL-10, IL-12p70, TNF and IFN-γ were measured in the supernatant of BMDCs (5 × 105 BMDCs/well in 24w plate) after 24 h of incubation with bacteria (bacterium: BMDC, 10:1).

Error bars represent SEM, n = 3, ***indicates p < 0.001, **p < 0.01, *p < 0.05 compared to the control.
F. prausnitzii A2-165 but not C. hathewayi 82-B attenuates inflammation in a chronic relapsing model of colitis
The capacity of Fp A2-165 to attenuate parameters of inflammatory colitis was investigated using a recently described model of chronic relapsing colitis16 and compared to the one of Ch 82-B. Ch 82-B was selected as it also produces butyrate but does not induce significant levels of IL-10 in BMDCs, or hDCs compared to Fp A2-165. Mice were administered a first dose of DNBS, and, after full recovery (day 10), were administered either Fp A2-165, Ch 82-B or ethanol intragastrically for other 10 days. On day 20 mice received a second lower dose of DNBS to induce a Th1-driven inflammatory response to the hapten. The weight loss of the Fp A2-165 treated mice was significantly smaller than the one of the DNBS control mice at the endpoint of the experiment (Fig. 5a). Additionally, the Fp A2-165 treated mice gained body weight significantly more rapidly than Ch 82B treated or DNBS control mice after the second administration of DNBS (Supplementary Fig. S4). In the Fp A2-165 treated mice, the macroscopic disease scores were significantly lower than in the DNBS control and in the Ch 82-B groups (Fig. 5b). Additionally, myeloperoxidase (MPO), an enzyme produced mainly by neutrophils and used as a marker of neutrophilic infiltration, was significantly lower in the Fp A2-165 treated group than in the other groups, suggesting reduced inflammatory responses leading to the recruitment of neutrophils (Fig. 5c). Fp A2-165 and Ch 82-B had no effect on histological damages (Fig. 5d).
Figure 5. Effects of F. prausnitzii A2-165 and C. hathewayi 82-B on DNBS induced colitis.

Percentage of weight loss at the end of the experiment (a), macroscopic scores (b), MPO activity (c) and histological scores (d) in control non-inflamed (PBS-Gly no DNBS), control inflamed (PBS-Gly), F. prausnitzii A2-165, C. hathewayi 82-B treated mice. Error bars represent SEM, n = 8, *indicates p < 0.05 compared to the control PBS-DNBS.
In vivo CD4+ T cell responses
To investigate whether the attenuation of certain disease scores in the DNBS model was due to the modulation of the mucosal T cell response by Fp A2-165, we used an in vivo model of nasal tolerance induction26. Nasal tolerance is known to have many similarities to oral tolerance induction26,27 and application of bacteria via the nasal route effectively targets responses in the nose-draining CLNs. In short, BALB/c acceptor mice were adoptively transferred with CFSE labelled naive DO11.10 OVA T cell receptor (TCR) transgenic T cells and after 24 h, received an intranasal (i.n.) application of either the bacteria together with OVA or OVA alone. After 72 h, single cell suspensions isolated from CLNs and spleens were analysed to assess the proliferation and phenotype of OVA-specific T cells (OVA-T cells).
In Fp A2-165 treated mice, the percentage of dividing OVA-T cells in the CLNs was increased compared to the control mice administered OVA alone. In the spleen of control mice, that received OVA i.n., a population of OVA-T cells was found in the third, fourth or fifth division. In agreement with previous observations, no CFSE fluorescence was detected corresponding with T cells in first or second division. Therefore, this population reflects cells that have differentiated in the CLNs and entered the circulation upon exiting the CLNs. Administration of Fp A2-165 together with OVA significantly increased the number of divided OVA-T cells isolated from spleens compared to control mice (Fig. 6a and Supplementary Fig. S5). Together, these data demonstrate that i.n. administration of Fp A2-165 enhances the number of OVA-T cells that divide in the CLNs. In contrast, Ch 82-B administration significantly decreased the proliferation of OVA-T cells isolated from CLNs compared to the control and Fp A2-165 (Fig. 6a). This effect was also observed in a second independent experiment although the differences were not statistically significant (Supplementary Fig. S3). Increased numbers of divided OVA-T cells were not detected in the spleen of mice administered Ch 82-B (Fig. 6a).
Figure 6. Percentage of dividing and IFN-γ+ OVA-T cells in vivo.

CFSE labelled naive OVA-T cells (KJ1-26+/CD4+) were adoptively transferred in BALB/c mice, after 24 h, mice were administered i.n. with bacteria plus OVA and after additional 72 h, OVA-T cells were isolated from cervical lymph nodes (CLNs) and spleens and analysed. (a) Percentage of dividing OVA-T cells (KJ1-26+/CD4+) isolated from CLNs or spleens. (b) Percentage of IFN-γ+ OVA-T cells (KJ1-26+/CD4+) isolated from CLNs. *indicates p < 0.05, **indicates p < 0.01 compared to the control administered OVA alone.
I.n. administration of either Fp A2-165 or Ch 82-B together with OVA significantly decreased the number of IFN-γ secreting OVA-T cells isolated from the CLNs. In particular, Fp A2-165 induced an approximately three-fold reduction in the percentage of IFN-γ secreting cells (Fig. 6b). However, these bacteria did not affect the intracellular expression of Foxp3, IL-17, or IL-10 in OVA-T cells isolated from the CLNs (not shown).
Overall re-stimulation of CLN cells led to low and variable levels of cytokine release. This is likely to be due to the low frequency of OVA-T cells within the total CLN cell population. Notably, CLN cells of Ch 82-B treated mice released lower amounts of cytokines but this was only significant for IL-10 and MCP-1 (Fig. 7).
Figure 7. Cytokine secretion by OVA-T cells after ex vivo re-stimulation.

CFSE labelled naive OVA-T cells were adoptively transferred in BALB/c mice, after 24 h, mice were administered i.n. with bacteria plus OVA and after additional 72 h, OVA-T cells were isolated from CLNs and re-stimulated with OVA for additional 24 h and then analysed. *indicates p < 0.05 compared to the control administered OVA alone.
Effects of F. prausnitzii A2-165 and C. hathewayi 82-B on T cell differentiation in BMDC-T cell co-cultures
In order to investigate whether the effects of Fp A2-165 and Ch 82-B on activation, differentiation and modulation of T cell responses in vivo could be observed in an in vitro assay we cultured BMDCs in the presence or absence of the bacteria or LPS for 24 h and then OVA was added for a further 6 h. Finally, naive CFSE labelled CD4+ OVA-T cells were added for an additional 72 h.
All conditions of BMDC stimulation increased the number of OVA-T cells undergoing proliferation compared to the unstimulated BMDC control. No significant differences in the percentage of dividing T cells were observed between BMDC stimulation with LPS, Fp A2-165 or Ch 82-B (Fig. 8a and Supplementary Fig. S6).
Figure 8. OVA-T cells (KJ1-26+/CD4+) division (a), IFN-γ+, IL-17+, Foxp3+ cells (b) and cytokine secretion (c) after incubation with BMDCs pre-stimulated with bacteria (bacterium: BMDC, 10: 1).

BMDCs (5 × 104 BMDCs/well in 96w plate) were loaded with OVA (0.5 mg/ml) and cultured in the presence or absence of the bacteria or LPS and after 24 h, CFSE labelled OVA-T cells (5 × 105 T cells/well) were added, after 72 h cytokine secretion in the supernatant and intracellular markers were measured. Error bars represent SEM, n = 3, *** indicates p < 0.001 compared to the control, *p < 0.05 compared to the control. Intracellular IL-10 staining was negative for all samples.
As observed in vivo the intracellular staining of OVA-T cells revealed that Fp A2-165 decreased the percentage of total IFN-γ+ T cells compared to the control but had no effect on IL-17+ and Foxp3+ T cells (Fig. 8b). Notably, BMDCs pre-incubated with Fp A2-165 induced increased IL-10 secretion during OVA-T cell proliferation (Fig. 8c) compared to the control. However, no significant changes were observed in the amounts of IFN-γ and IL-17 in any of the culture supernatants compared to the control (Fig. 8c). None of the bacteria altered the levels of secreted IL-2, IL-6 or TNF compared to the control (not shown). In contrast to Fp, Ch 82-B induced an increase in the percentage of IFN-γ+, IL-17+ and Foxp3+ T cells compared to the control (Fig. 8b) which is not consistent with results with this strain in vivo. It may be that the in vitro immune assays are not predictive for the complex situation in vivo where stromal cytokines and factors also influence APC phenotypes. Another possibility is that this inconsistency is due to the fact that Ch 82-B possesses almost no TLR signalling activity (Fig. 2) but can strongly activate hDCs and BMDCs in vitro (Supplementary Fig. S1 and S2) resulting in no or low amounts of cytokine secretion. Thus activation of hDCs and BMDCs by Ch 82-B is presumably the consequence of phagocytosis and intracellular NLR activation.
Discussion
Members of the Firmicutes, including members of the Clostridium clusters IV and XIVa, are reduced in abundance in the fecal microbiota of IBD patients28,29. Additionally, mucosal abundance of Fp, a member of Clostridium cluster IV, has been correlated with increased recurrence of colitis in CD patients14. Fp strain A2-165 and its culture supernatant were previously shown to attenuate colitis in mouse models. The incubation of hPBMCs with Fp A2-165 induced a relatively high IL-10 to IL-12p70 cytokine ratio, suggesting that the bacterium might have an effect on DCs and induction of T cells in the mucosal lymphoid tissue14,16. Furthermore, IL-10 produced by APCs in the LP has been reported to enhance the suppressive activity of Foxp3+ Tregs in the mucosa30.
We investigated the immunomodulatory properties of five Fp strains and eight other abundant intestinal commensal bacteria of Clostridium clusters IV and XIVa. Other Clostridia, when administered as a spore preparation have been shown to induce the expansion and activation of Tregs after colonization of germ-free mice31. All the bacteria tested were able to induce efficient maturation and activation of hDCs (Supplementary Fig. S1). In hDCs, the highest levels of IL-10 were induced by Fp A2-165, Eh L2-7 and Mr Mag1B. Fp A2-165 was among the weakest inducers of IL-12p70 while Mr Mag1B and Eh L2-7 were among the strongest inducers. Additionally, these strains induced the largest amounts of TNF-α and IL-6. The Fp strains, Eh L2-7 and Er A1-86 induced also relatively large amounts of IL-1β (Fig. 1). PCA revealed that Fp A2-165, Mr Mag1B and Eh L2-7 cause distinct effects on the immune cells when compared to the other strains. These differential effects were due to the high levels of IL-10 induced by Fp A2-165, the high levels of IL-12p70 and TNF-α induced by Mr Mag1B and the high levels of IL-10 and IL-12p70 induced by Eh L2-7. Furthermore, Fp A2-165 is separated from the other Fp strains (Fig. 3).
In general, high TLR signalling activity was associated with the ability of commensal bacteria to induce cytokines in the DC assays. TLR4 signalling was induced by Eh L2-7 and Mr Mag1B, belonging to the Firmicutes lineage of bacteria, which are generally considered as Gram-positive. However, due to their unique cell wall composition, upon Gram staining these bacteria appeared to be Gram-negative to Gram-variable24,25. Ch 82-B and Cx Lac1D induced low NF-κB activation in the TLR assays and low or undetectable cytokines in the DC assay (Figs 1 and 2) despite the fact that they induced activation and maturation of DCs (Supplementary Fig. S1). These strains might express MAMPs that are shielded or not recognized by the TLRs tested or might possess immunomodulatory components that inhibit activation of the NF-κB pathway. This appears to be a unique property and may be an evolutionary adaptation to avoid immune recognition by the host as proposed for Bacteroides fragilis32.
Fp A2-165 has been previously reported to be anti-inflammatory and attenuate inflammation in mouse models of colitis14,16, therefore we compared its protective effects in the DNBS model of chronic relapsing colitis with that of Ch 82-B, a butyrate producing organism that induces only low amounts of IL-10 secretion in hDCs and BMDCs. In the Fp A2-165 treated mice, body weight loss, tissue MPO and macroscopic disease scores were attenuated compared to the control and Ch 82-B groups (Fig. 5). To investigate whether the attenuation of inflammation parameters in the DNBS model was due to modulation of the mucosal T cell response by Fp A2-165, we i.n. inoculated OVA or OVA together with the bacteria into BALB/c mice adoptively transferred with naive OVA-TCR transgenic T cells. After 72 h, single cell suspensions isolated from CLNs and spleens were analysed to assess the proliferation and phenotype of OVA-T cells. Fp A2-165 enhanced the number of OVA-T cells that divide in the CLNs and decreased the percentage of IFN-γ secreting T cells (Fig. 6). The effects of Fp on T cell differentiation in vivo may be due to the uptake of bacteria or microbial factors (including MAMPs) resulting in increased DC activation as seen in the in vitro experiments (Supplementary Fig. S1 and S2). The activated DCs may then migrate from PPs or LP to drive enhanced antigen presentation of OVA in the CLNs. Although Fp A2-165 administration increased the percentage of dividing T cells in vivo, it decreased the percentage of IFN-γ+ T cells without affecting Foxp3+ or IL-17+ T cells (Fig. 6). This finding suggests that Fp A2-165 modulated the phenotype of mucosal DCs that migrate from the LP to suppress IFN-γ differentiation in the draining CLNs. Unfortunately we were not able to detect intracellular IL-10 staining in CLNs cells. IL-10 may suppress differentiation of IFN-γ secreting cells in the CLNs once the cells have proliferated and re-express their IL-10R. Future experiments using in vivo blocking with anti-IL-10R antibodies would provide an answer to this hypothesis.
Strikingly, the effects of Fp A2-165 on T cell proliferation and differentiation of IFN-γ secreting T cells in vivo were also observed in vitro using Fp A2-165 stimulated BMDCs co-cultured with naive D011.10 T cells. In these co-cultures, secretion of IL-10 was significantly higher than the control when the BMDCs were pre-stimulated with Fp A2-165 but not with other stimuli (Fig. 8). The concomitant enhanced proliferation and high levels of IL-10 secretion in BMDC-T cell co-cultures may seem counter intuitive as IL-10 is known to suppress T cell proliferation. However, the onset of T cell proliferation may not be sensitive to IL-10 suppression because the cells down regulate expression of the IL-10Rα chain33.
In contrast to Fp A2-165, Ch 82-B reduced the number of proliferating OVA-T cells in the CLNs in vivo (Fig. 6). This finding was in contrast to the in vitro results showing that Ch 82-B stimulated BMDCs were highly effective in stimulating T cell proliferation. Indeed, the percentage of T cells staining positive for intracellular markers of different T cell subsets (i.e. IFN-γ, IL-17 and Foxp3) were significantly higher for T cells incubated with BMDCs pre-stimulated with Ch 82-B than BMDCs pre-stimulated with Fp A2-165, LPS or left unstimulated (Fig. 8). The reasons for this are not clear but may be due to inhibitory effects of Ch on the migration of DCs from the mucosa to the draining lymph nodes, which is not mimicked in vitro.
In conclusion, we demonstrated that Fp A2-165 has a strong capacity to induce IL-10 in human and murine DCs and influence T cell differentiation in vitro and in vivo. The reduction in IFN-γ+ cells in vivo and the IL-10 induction in the LP cells of damaged intestinal tissue could be a mechanisms contributing to the suppressive effect on inflammation in mouse colitis models14,16. Moreover, these findings suggest that this normally abundant bacterium might contribute to immune homeostasis in the intestine via its anti-inflammatory properties.
Materials and Methods
Bacterial strains and culturing conditions
All the commensal bacteria used (Table 1) were isolated from human faeces and identified by 16S rRNA sequencing as previously described2. L. plantarum WCFS1 was isolated from human saliva. The commensal bacteria were maintained at 37 °C on YCFAG in an anaerobic tent. Bacteria were grown in YCFAG broth to an optical density at 600 nm (A600) of 0.8, which corresponds to the late exponential or early stationary growth phase. L. plantarum WCFS1 was cultured overnight to stationary phase in MRS broth (MRS, Merck, Darmstadt, Germany) at 37 °C. Bacteria were harvested by centrifugation, washed in PBS, resuspended in PBS containing 20% glycerol and stored at −80 °C prior to use in the assays. Bacteria were quantified by fluorescent in situ hybridization (FISH) or phase contrast microscopy. All buffers and media used for the anaerobic bacteria were deoxygenated by flushing with oxygen free nitrogen for 30 minutes.
Human DCs assays
This study was approved by Wageningen University Ethical Committee and was conducted in accordance with the principles of the ‘Declaration of Helsinki’ (52nd WMA General Assembly, Edinburgh, Scotland, October 2000). Buffy coats from healthy blood donors were obtained from the Sanquin Blood bank in Nijmegen (The Netherlands). A written informed consent was obtained from each volunteer before sample collection. See Supplementary Methods.
TLR signalling assays
Mice
For the DNBS induced colitis, animal experiments were approved by the Ethic Committee of Jouy en Josas research centre. Male C57BL/6 mice (6 weeks old) were purchased from Janvier Labs and maintained at the animal facility under specific pathogen-free conditions. Mice were housed under standard conditions for a minimum of 1 week before experimentation.
For the in vivo nasal tolerance and the in vitro BMDCs and T cells assays, animal experiments were approved by the Animal Experimental Committee of the Erasmus Medical Centre. BALB/c mice were obtained from Charles River Laboratories (Maastricht, The Netherlands) and kept in the Erasmus University Medical Centre. DO11.10 transgenic mice on BALB/c background, which have a T cell receptor (TCR) specific for the OVA 323-339 peptide, were bred at the Erasmus University Medical Centre. Mice were kept under specific pathogen-free housing conditions.
Animals were handled in accordance with the general principles and guidelines recommended by the European Community (Directive 86/609/EEC) and current French legislation (Law 87–848) or Dutch legislation (The Experiments on Animals Act, 1997).
Mouse BMDCs assays
Induction of DNBS colitis and bacteria administration
The DNBS induced chronic colitis model was described by Martin et al.16. Briefly, mice were anesthetized and a 4 cm long catheter (Ecimed, France) was attached to a 1 ml-syringe and inserted 3.5 cm into the colon. Colitis was induced by intrarectal injection of 150 mg/kg of DNBS solution (Sigma) in 30% ethanol (EtOH). Control mice (that did not develop colitis) received only EtOH. Mice were supplied with 6% sucrose in the drinking water for the first 3 days after DNBS injection to prevent dehydration (DNBS period). Ten days following DNBS injection, 200 μL containing either 1 × 109 bacteria or 200 μL of PBS were administered intragastrically daily for 10 days. Colitis was reactivated 21 days after the first DNBS injection (recovery period) with a second injection of 150 mg/kg of DNBS solution, control mice received only EtOH. Mice weight was recorded through all the experiment, each group consisted of 8 mice.
Macroscopic scores, colonic histology and myeloperoxidase activity
Severity of colitis was determined as previously described16. See Supplementary Methods.
Purification of DO11.10 T-cells
In vitro assays with DO11.10 T cells
Adoptive transfer of DO11.10 T cells and intranasal challenge
For the in vivo nasal tolerance study, acceptor BALB/c mice received 6 × 106 CFSE labelled KJ1-26+ CD4+ T cells in 200 μl saline (NaCl 0.9%, Brown) by injection in the tail vein. DO11.10xRag-/- T cells were isolated as described above. Twenty-four h after the adoptive transfer, mice were administered i.n. with 2 × 108 bacteria in 15 μl of saline containing 400 μg of OVA or 15 μl of saline with OVA as a control. Throughout the in vivo experiment, groups of mice that received different treatments were housed separately. After 72 h, mice were sacrificed, nose-draining superficial and deep cervical lymph nodes (CLNs) and spleens were collected and single cell suspensions were analysed by flow cytometry for cell division and stained for surface or intracellular protein detection or used for ex vivo T-cells restimulation.
Ex vivo DO11.10 T-cell restimulation
Flow cytometry
Statistics and principal component analysis
Differences in NF-κB activation in the TLR assays, cytokine secretion, surface marker expression and percentage of dividing or positive cells were analysed comparing samples treated with the bacteria or LPS with the untreated control. Data were analysed by one way Anova followed by Dunnett’s multiple comparison test for the in vitro assays or Newman-Keuls multiple comparison test for the in vivo assays using the GraphPad PRISM (GraphPad Software). PCA was used to analyse the variance of the level of cytokines along the different types of bacteria studied. PCA was computed on the standardized data by using the function princomp of the software Matlab.
Additional Information
How to cite this article: Rossi, O. et al. Faecalibacterium prausnitzii A2-165 has a high capacity to induce IL-10 in human and murine dendritic cells and modulates T cell responses. Sci. Rep. 6, 18507; doi: 10.1038/srep18507 (2016).
Supplementary Material
Acknowledgments
This work was financially supported by the EC FP7 Cross-talk project (PITN-GA-2008-215553). The authors thank the Histology Platform from GABI research unit and especially Abdelhak Boukadiri for their technical support in the histology sample preparation and Marlène Héry, Charline Pontlevoy, Jerome Pottier and André Tiffoche (UE0907 IERP, Jouy en Josas) for their help during animal experiments. The authors thank Rafael Muñoz-Tamayo (INRA) for his help in performing the PCA.
Footnotes
Author Contributions O.R., J.W., S.H.D., H.J.F., H.J.M.H., P.L., J.N.S. and J.M.W. conceived the experiments, O.R., L.v.B., F.C., M.T.K. and N.T., conducted the experiments, O.R., L.v.B., F.C., M.T.K. and H.S. analysed the results. O.R., J.W. and J.N.S. wrote the manuscript, all authors reviewed the manuscript.
References
- Whitman W. B., Coleman D. C. & Wiebe W. J. Prokaryotes: the unseen majority. Proc Natl Acad Sci USA 95, 6578–6583 (1998). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopez-Siles M. et al. Cultured representatives of two major phylogroups of human colonic Faecalibacterium prausnitzii can utilize pectin, uronic acids, and host-derived substrates for growth. Applied and environmental microbiology 78, 420–428, doi: 10.1128/AEM.06858-11 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sekirov I., Russell S. L., Antunes L. C. & Finlay B. B. Gut microbiota in health and disease. Physiological reviews 90, 859–904, doi: 10.1152/physrev.00045.2009 (2010). [DOI] [PubMed] [Google Scholar]
- Kiyono H., McGhee J. R., Wannemuehler M. J. & Michalek S. M. Lack of oral tolerance in C3H/HeJ mice. J Exp Med 155, 605–610 (1982). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanaka K. & Ishikawa H. Role of intestinal bacterial flora in oral tolerance induction. Histology and histopathology 19, 907–914 (2004). [DOI] [PubMed] [Google Scholar]
- Gaboriau-Routhiau V. et al. The key role of segmented filamentous bacteria in the coordinated maturation of gut helper T cell responses. Immunity 31, 677–689, doi: 10.1016/j.immuni.2009.08.020 (2009). [DOI] [PubMed] [Google Scholar]
- Atarashi K. et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 500, 232–236, doi: 10.1038/nature12331 (2013). [DOI] [PubMed] [Google Scholar]
- Round J. L. & Mazmanian S. K. Inducible Foxp3 + regulatory T-cell development by a commensal bacterium of the intestinal microbiota. Proc Natl Acad Sci USA 107, 12204–12209, doi: 10.1073/pnas.0909122107 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sokol H. et al. Low counts of Faecalibacterium prausnitzii in colitis microbiota. Inflammatory bowel diseases 15, 1183–1189, doi: 10.1002/ibd.20903 (2009). [DOI] [PubMed] [Google Scholar]
- Chassaing B. & Darfeuille-Michaud A. The commensal microbiota and enteropathogens in the pathogenesis of inflammatory bowel diseases. Gastroenterology 140, 1720–1728, doi: 10.1053/j.gastro.2011.01.054 (2011). [DOI] [PubMed] [Google Scholar]
- Flint H. J., Scott K. P., Duncan S. H., Louis P. & Forano E. Microbial degradation of complex carbohydrates in the gut. Gut microbes 3, 289–306, doi: 10.4161/gmic.19897 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duncan S. H. Growth requirements and fermentation products of Fusobacterium prausnitzii, and a proposal to reclassify it as Faecalibacterium prausnitzii gen. nov., comb. nov. International Journal of Systematic and Evolutionary Microbiology 52, 2141–2146, doi: 10.1099/ijs.0.02241-0 (2002). [DOI] [PubMed] [Google Scholar]
- Barcenilla A. et al. Phylogenetic relationships of butyrate-producing bacteria from the human gut. Applied and environmental microbiology 66, 1654–1661 (2000). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sokol H. et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc Natl Acad Sci U S A 105, 16731–16736, doi: 10.1073/pnas.0804812105 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varela E. et al. Colonisation by Faecalibacterium prausnitzii and maintenance of clinical remission in patients with ulcerative colitis. Alimentary pharmacology & therapeutics 38, 151–161, doi: 10.1111/apt.12365 (2013). [DOI] [PubMed] [Google Scholar]
- Martin R. et al. The commensal bacterium Faecalibacterium prausnitzii is protective in DNBS-induced chronic moderate and severe colitis models. Inflammatory bowel diseases 20, 417–430, doi: 10.1097/01.MIB.0000440815.76627.64 (2014). [DOI] [PubMed] [Google Scholar]
- Pryde S. E., Duncan S. H., Hold G. L., Stewart C. S. & Flint H. J. The microbiology of butyrate formation in the human colon. FEMS microbiology letters 217, 133–139 (2002). [DOI] [PubMed] [Google Scholar]
- Yin L., Laevsky G. & Giardina C. Butyrate suppression of colonocyte NF-kappa B activation and cellular proteasome activity. J Biol Chem 276, 44641–44646, doi: 10.1074/jbc.M105170200 (2001). [DOI] [PubMed] [Google Scholar]
- Place R. F., Noonan E. J. & Giardina C. HDAC inhibition prevents NF-kappa B activation by suppressing proteasome activity: down-regulation of proteasome subunit expression stabilizes I kappa B alpha. Biochemical pharmacology 70, 394–406, doi: 10.1016/j.bcp.2005.04.030 (2005). [DOI] [PubMed] [Google Scholar]
- Meijerink M. et al. Identification of genetic loci in Lactobacillus plantarum that modulate the immune response of dendritic cells using comparative genome hybridization. PLoS One 5, e10632, doi: 10.1371/journal.pone.0010632 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Foligne B. et al. Correlation between in vitro and in vivo immunomodulatory properties of lactic acid bacteria. World journal of gastroenterology : WJG 13, 236–243 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang B., Morinobu A., Horiuchi M., Liu J. & Kumagai S. Butyrate inhibits functional differentiation of human monocyte-derived dendritic cells. Cellular immunology 253, 54–58, doi: 10.1016/j.cellimm.2008.04.016 (2008). [DOI] [PubMed] [Google Scholar]
- Erridge C., Duncan S. H., Bereswill S. & Heimesaat M. M. The induction of colitis and ileitis in mice is associated with marked increases in intestinal concentrations of stimulants of TLRs 2, 4, and 5. PLoS One 5, e9125, doi: 10.1371/journal.pone.0009125 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duncan S. H. & Flint H. J. Proposal of a neotype strain (A1-86) for Eubacterium rectale. Request for an opinion. Int J Syst Evol Microbiol 58, 1735–1736, doi: 10.1099/ijs.0.2008/004580-0 (2008). [DOI] [PubMed] [Google Scholar]
- Chevrot R., Carlotti A., Sopena V., Marchand P. & Rosenfeld E. Megamonas rupellensis sp. nov., an anaerobe isolated from the caecum of a duck. Int J Syst Evol Microbiol 58, 2921–2924, doi: 10.1099/ijs.0.2008/001297-0 (2008). [DOI] [PubMed] [Google Scholar]
- Unger W. W. et al. Early events in peripheral regulatory T cell induction via the nasal mucosa. J Immunol 171, 4592–4603 (2003). [DOI] [PubMed] [Google Scholar]
- Samsom J. N. Regulation of antigen-specific regulatory T-cell induction via nasal and oral mucosa. Critical reviews in immunology 24, 157–177 (2004). [DOI] [PubMed] [Google Scholar]
- Seksik P. et al. Alterations of the dominant faecal bacterial groups in patients with Crohn’s disease of the colon. Gut 52, 237–242 (2003). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macfarlane G. T., Blackett K. L., Nakayama T., Steed H. & Macfarlane S. The gut microbiota in inflammatory bowel disease. Current pharmaceutical design 15, 1528–1536 (2009). [DOI] [PubMed] [Google Scholar]
- Murai M. et al. Interleukin 10 acts on regulatory T cells to maintain expression of the transcription factor Foxp3 and suppressive function in mice with colitis. Nat Immunol 10, 1178–1184, doi: 10.1038/ni.1791 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Atarashi K. et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 331, 337–341, doi: 10.1126/science.1198469 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weintraub A., Zahringer U., Wollenweber H. W., Seydel U. & Rietschel E. T. Structural characterization of the lipid A component of Bacteroides fragilis strain NCTC 9343 lipopolysaccharide. Eur J Biochem 183, 425–431 (1989). [DOI] [PubMed] [Google Scholar]
- Liu Y., Michalopoulos G. K. & Zarnegar R. Structural and functional characterization of the mouse hepatocyte growth factor gene promoter. J Biol Chem 269, 4152–4160 (1994). [PubMed] [Google Scholar]
- Louis P. et al. Restricted distribution of the butyrate kinase pathway among butyrate-producing bacteria from the human colon. Journal of bacteriology 186, 2099–2106 (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duncan S. H., Louis P. & Flint F. J. Lactate-utilizing bacteria, isolated from human feces, that produce butyrate as a major fermentation product. Applied and environmental microbiology 70, 5810–5817, doi: 10.1128/AEM.70.10.5810-5817.2004 (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.