

Abstract
Wnt signaling regulates multiple aspects of vertebrate central nervous system (CNS) development, including neurogenesis. However, vertebrate genomes can contain up to 25 Wnt genes, the functions of which are poorly characterized partly due to redundancy in their expression. To identify candidate Wnt genes as candidate mediators of pathway activity in specific brain progenitor zones, we have performed a comprehensive expression analysis at three different stages during zebrafish development. Antisense RNA probes for 21 Wnt genes were generated from existing and newly synthesized cDNA clones and used for in situ hybridization on whole embryos and dissected brains. As in other species, we found that Wnt expression patterns in the embryonic zebrafish CNS are complex and often redundant. We observed that progenitor zones in the telencephalon, dorsal diencephalon, hypothalamus, midbrain, midbrain-hindbrain boundary, cerebellum and retina all express multiple Wnt genes. Our data identify 12 specific ligands that can now be tested using loss-of-function approaches.
Introduction
Wnt/β-Catenin signaling is known to act in multiple ways to regulate vertebrate central nervous system (CNS) development, including as a mitogen [1], and in neural specification and differentiation [2–4]. The pathway also functions in post-embryonic neurogenesis, to promote the differentiation of neural progenitor cells in the dentate gyrus of the hippocampus [5], the rostral migratory stream [6], and the hypothalamus [4]. However with a few exceptions such as wnt7a in the dentate gyrus [7], it has been difficult to link functions in defined neural progenitor populations to specific Wnt ligands, possibly due to extensive redundancy within the Wnt family [8, 9]. In addition different Wnt ligands can activate multiple downstream pathways in the same tissue, such as in the zebrafish fin where wnt10a and wnt5b are both required for regeneration through ß-catenin-dependent and independent signaling, respectively [10]. It is therefore important to first precisely identify the specific Wnt ligands expressed in each neural progenitor population in order to test their function in neurogenesis.
Our laboratory is interested in the role of Wnt/ß-catenin signaling in hypothalamic neurogenesis, where we have previously shown a requirement for Lef1-mediated transcription in progenitor differentiation [4]. While we have identified one candidate ligand (wnt8b) in this region, knockout of this gene in mouse does not produce significant defects in brain development [11]. We thus decided to systematically examine the expression of the entire Wnt gene family in the developing zebrafish CNS in order to identify other candidates that may regulate hypothalamic neurogenesis. While many of these genes have been previously reported to have expression in specific brain regions (zfin.org), others have not been characterized and no single study has compared all the patterns at multiple stages.
For this work we examined the expression of 21 Wnt genes that either had known brain expression or were previously unexamined. While a comprehensive analysis of Wnt gene expression during early developmental stages has been previously performed [12], we carried out our experiments at 24, 48 and 72 hours post-fertilization (hpf), to cover both embryonic and post-embryonic CNS progenitor populations. At 48 and 72 hpf, we specifically focused on known progenitor zones including the telencephalic pallium/subpallium [13], the dorsal diencephalon (epithalimus and thalamus [14]), the ventral diencephalon (hypothalamus [4]), the midbrain [15], the midbrain/hindbrain boundary [16], the cerebellum [17], and the ciliary marginal zone (CMZ) of the retina [18]. Ultimately we were able to identify 12 genes with specific brain expression at all stages, most of which were localized to progenitor zones, and we found 3 genes (wnt8b, wnt11r, and wnt16) expressed in the hypothalamic posterior ventricular recess. Our results highlight the redundancy of Wnt ligand expression during zebrafish development, and lay the foundation for future functional analysis of Wnt signaling throughout the CNS.
Materials and Methods
Zebrafish embryo maintenance
This research was approved by the University of Utah IACUC Committee. All experiments were conducted on zebrafish embryos at 72 hours post-fertilization or younger, which are not considered to be vertebrate animals by IACUC guidelines. Zebrafish embryos were euthanized in tricaine. Fertilized wild-type (AB*) zebrafish embryos were and staged according to Kimmel et al. [19], and raised until 24, 48 and 72 hours post-fertilization (hpf). Embryos were fixed overnight at 4°C in 4% paraformaldehyde with 5% sucrose. Brains were manually dissected for in situ hybridization at 48 and 72 hpf.
Cloning of zebrafish Wnt genes
The Ensembl Danio rerio genome database was used to identify genomic loci for all unpublished genes. Primers were designed to amplify ~500bp cDNA fragments for each gene (Table 1), and RT-PCR was performed on total RNA extracted from 24 hpf embryos using a Superscript II kit (Invitrogen). Amplicons were then subcloned into PCRII-TOPO (Invitrogen), and sequenced to verify gene identity as well as to confirm orientation for generation of antisense RNA probes.
Table 1. Sources of published in situ probe templates or primers used to amplify cDNA.
| Gene | Reference or primers |
|---|---|
| wnt1 | [20] |
| wnt2 | [21] |
| wnt2ba | [21] |
| wnt2bb | [22] |
| wnt3 | [23] |
| wnt3a | [20] |
| wnt4a | [24] |
| wnt4b | F: TGTATTTGATGTGTCGGCCA R: ACGCAGACACTTTGCCTTTT |
| wnt5a | F: ATGATGCTGCTGAAGCTGAAGT R: CTTACAGGTGTAAACCTCTTCTTTTTGT |
| wnt5b | F: GGAAGGATGGATGTGAGAATGAA R: CGTCTGCTACTTGCACACAAACT |
| wnt7aa | F: ATGAGCAGGAAAACGCGC R: TCACTTGCATGTGTACACTTCTGTC |
| wnt7ba | [25] |
| wnt7bb | [25] |
| wnt8b | [26] |
| wnt9a | F: GGAGAAGAAGCAGCGCAGAA R: CTTACAGGTGTAAACCTCTTCTTTTTGT |
| wnt9b | F: GGGATTTCAACACGGACAGATAG R: AAGCGCGTGAGACAATGCT |
| wnt10a | F: ATGAGCTCTCACGACATCAGTTG R: TGCTTGCTTATTTCATTTGCAGA |
| wnt10b | F: GTTCGACGCAATGGAGTTACC R: TGCTGCTCACTTGCACACATTA |
| wnt11 | [27] |
| wnt11r | [28] |
| wnt16 | Gift from Gilbert Weidinger |
In situ hybridization
Antisense riboprobes were synthesized and in situ hybridization was performed as previously described [29], with the addition of 5% dextran sulfate (Sigma-Aldrich D8906) to the hybridization buffer. To increase visualization of staining, some 24 hpf embryos were also incubated in 3%H2O2/0.5%KOH to remove pigmentation. Post-hybridization washes were carried out using a Biolane HTI in situ machine (Huller and Huttner AG). After staining, embryos were stored in MeOH and cleared in 70% glycerol for dissection and imaging. For sectioning, embryos were post-fixed in 4% paraformaldehyde for 1–2 hours at room temp, washed in PBS, and cryoprotected in 30% sucrose in PBS. Embryos were then embedded in OCT and sectioned at 20μm thickness on a Leica cryostat. Images were acquired using an Olympus BX51WI compound microscope and an Olympus Microfire camera. Digital images were cropped and aligned using Adobe Photoshop.
Results
Expression in the 24 hpf brain
We generated antisense probes for 21 zebrafish Wnt genes either from previously published DNA templates, or by RT-PCR amplification and subcloning (Table 1). The only known Wnt genes that we did not examine were wnt7ab and wnt8a due to their reported lack of CNS expression after somitogenesis [26, 30], and wnt6a and wnt6b due to their annotation after the initiation of this project. At 24 hpf much of the CNS is still rapidly proliferating and undergoing neurogenesis, and we observed expression of multiple Wnt genes expression throughout the brain (Fig 1 and Table 2). We found only two genes (wnt7ba and wnt8b) expressed in the telencephalon (Fig 1L' and 1N'). Eleven genes were expressed in the diencephalon, including seven in the epithalamus (Fig 1A', 1E', 1F', 1G', 1K', 1L', 1M' and 1N'), one (wnt4b) in the thalamus (Fig 1H'), and three (wnt8b, wnt11r, and wnt16) with expression in the hypothalamus (Fig 1N', 1T' and 1U'). Ten genes were expressed in the midbrain (Fig 1A', 1E', 1F', 1G', 1H', 1K', 1M', 1N', 1R' and 1T'), including four at the midbrain-hindbrain boundary (Fig 1A', 1E, 1N' and 1R'). Four genes were expressed in the rostral hindbrain (cerebellum) (Fig 1A', 1E', 1K' and 1R'). We also observed one gene (wnt4b) with expression in the floor plate of the hindbrain and spinal cord (Fig 1H). Nine other genes showed low-level ubiquitous or non-specific expression throughout the brain (Fig 1B–1D', 1I, 1J', 1O–1Q', 1S and 1S'), however we did observe specific expression of one gene (wnt2) in the retinal margin (Fig 1B').
Fig 1. Wnt gene expression at 24 hpf.

Lateral views of whole embryos are shown in left panels, and dorsal views of dissected brains are shown in right panels. Genes with nonspecific brain expression are indicated with grey text. tel, telencephalon; et, epithalamus; th, thalamus; hy, hypothalamus; mb, midbrain; mhb, midbrain-hindbrain boundary; cb, cerebellum; fp, floor plate.
Table 2. Wnt expression in the developing zebrafish brain.
| Telencephalon | Dorsal Diencephalon | Hypothalamus | Midbrain | Cerebellum | Retina (CMZ) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 24 hpf | 48 hpf | 72 hpf | 24 hpf | 48 hpf | 72 hpf | 24 hpf | 48 hpf | 72 hpf | 24 hpf | 48 hpf | 72 hpf | 24 hpf | 48 hpf | 72 hpf | 24 hpf | 48 hpf | |
| wnt1 | et | et | et | m | dm | dm | X | X | |||||||||
| wnt2 | X | X | |||||||||||||||
| wnt2ba | |||||||||||||||||
| wnt2bb | |||||||||||||||||
| wnt3 | et | et | et | m | dm | dm | X | X | X | ||||||||
| wnt3a | et | et th | et | X | d | d | |||||||||||
| wnt4a | et | et | et | X | v | v | X | ||||||||||
| wnt4b | th | th | X | v | v | X | |||||||||||
| wnt5a | |||||||||||||||||
| wnt5b | |||||||||||||||||
| wnt7aa | p | p | et | et | et | X | d | d | X | X | X | ||||||
| wnt7ba | X | p | p | et | et | et | d | d | |||||||||
| wnt7bb | p | et | et | et | X | d | X | ||||||||||
| wnt8b | X | sp | sp | et | X | pr | pr | m | |||||||||
| wnt9a | |||||||||||||||||
| wnt9b | |||||||||||||||||
| wnt10a | |||||||||||||||||
| wnt10b | et | et | m | dm | dm | X | X | X | |||||||||
| wnt11 | |||||||||||||||||
| wnt11r | X | pr | pr | X | X | ||||||||||||
| wnt16 | th | th | X | pr | pr | ||||||||||||
X, expression; p, pallium; sp, subpallium; et, epithalamus; th, thalamus; pr, posterior recess; d, dorsal; v, ventral; m, midbrain/hindbrain boundary.
Expression in the 48 hpf brain
By 48 hpf the CNS has largely completed morphogenesis, and several proliferative neurogenic zones are retained near the ventricles [31]. At this stage and beyond we only analyzed expression patterns of the 12 genes that were specifically expressed at 24 hpf (Fig 2 and Table 2). In the telencephalon, expression of wnt7aa, wnt7ba, and wnt7bb was localized to the dorsal (pallial) region and wnt8b expression was localized to the ventral (subpallial) region (Fig 2F–2I'). In the dorsal diencephalon, eight genes were expressed in the epithalamus (Fig 2A–2D', 2F–2H', 2J and 2J'), and three genes (wnt3a, wnt4b and wnt16) were expressed in the thalamus (Fig 2B, 2B', 2E, 2E', 2L and 2L'). At 48 hpf the third ventricle of the hypothalamus has elaborated into bilateral recesses, and we observed specific expression of wnt8b, wnt11r and wnt16 in the posterior recess as viewed from the ventral brain surface (Fig 2I, 2I' and 2K–2L'). Nine genes were expressed in the midbrain (Fig 2A–2H', 2J and 2J'), including three (wnt1, wnt3, and wnt10b) in the midbrain-hindbrain boundary (Fig 2A, 2B', 2J and 2J'), and two (wnt4a, wnt4b) in the ventral midline as well as the hindbrain floor plate (Fig 2D–2E'). The same four genes were expressed in the cerebellum as at 24hpf, with the addition of wnt7bb (Fig 2A–2B', 2F, 2F', 2H, 2H', 2J and 2J'). Finally, we observed expression of four genes in the ciliary marginal zone (CMZ) of the retina (Fig 3 and Table 2), which is the region containing neural progenitors [18].
Fig 2. Wnt gene expression at 48 hpf.

Lateral views of dissected brains are shown in left panels, and dorsal or ventral views of dissected brains are shown in right panels. p, pallium; sp, subpallium; et, epithalamus; th, thalamus; pr, posterior recess; mb, midbrain; mhb, midbrain-hindbrain boundary; cb, cerebellum; fp, floor plate.
Fig 3. Wnt gene expression in the retina at 48 hpf.
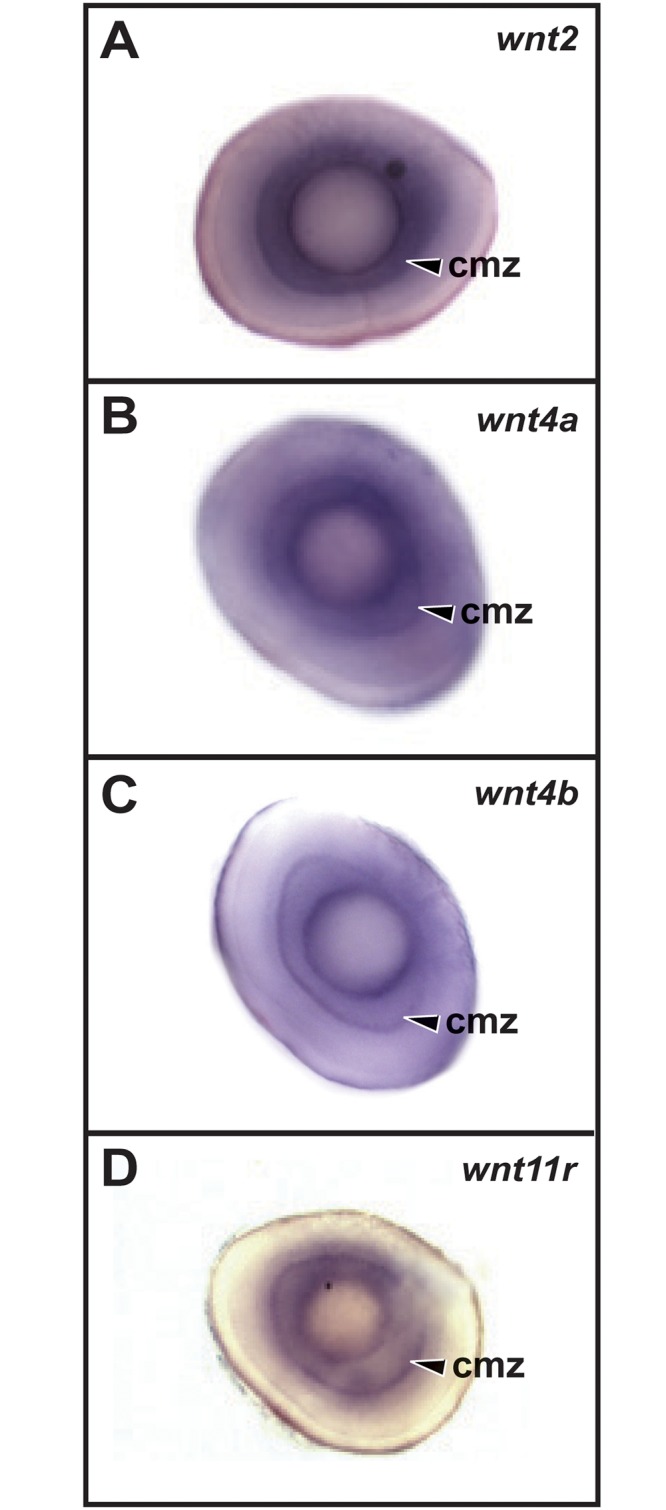
Dissected eyes are shown in all panels. cmz, ciliary marginal zone.
Expression in the 72 hpf brain
Due to the larger brain size at 72 hpf, we were only able to characterize specific expression patterns using dorsal or ventral whole-mount views (Fig 4 and Table 2). We found that wnt7aa and wnt7ba were expressed in the pallium (Figs 4F, 4G, 5A and 5B), and wnt8b was still expressed in the subpallium (Fig 4I). The same eight genes were expressed in the epithalamus as at 48 hpf (Fig 4A–4D, 4F–4H and 4J), and wnt16 continued to be expressed in the thalamus (Fig 4L). Similarly, wnt8b, wnt11r, and wnt16 continued to be expressed in the hypothalamic posterior recess (Figs 4I, 4K, 4L and 5C–5E), and eight genes were expressed in the midbrain and midbrain-hindbrain boundary, (Fig 4A–4G and 4J). Expression of wnt3, wnt7aa, and wnt10b was observed in the cerebellum (Fig 4B, 4F and 4J) and expression of wnt4a and wnt4b was maintained in the floor plate (Fig 4D and 4E).
Fig 4. Wnt gene expression at 72 hpf.

Dorsal or ventral views of dissected brains are shown in all panels. p, pallium; sp, subpallium; et, epithalamus; th, thalamus; pr, posterior recess; mb, midbrain; mhb, midbrain-hindbrain boundary; cb, cerebellum; fp, floor plate.
Fig 5. Specific expression of Wnt genes at 72 hpf and schematic diagram of brain anatomy.

20μm transverse cryosections at 72 hpf in panels A and B show expression in pallium (p). Ventral views of dissected brains in panels C-E show expression in the posterior hypothalamic recess (pr). Diagrams in panels F-H show regions of expression identified in Figs 1, 2 and 4.
Discussion
Our data clearly demonstrate the coincident expression of multiple Wnt genes in progenitor zones of the developing CNS (Fig 5F–5H and Table 2), and support previous observations of gene redundancy. Some of the overlapping gene expression that we observed could be explained by closely related orthologs arising from the teleost genome duplication [32], such as in the cases of wnt4a/b and wnt7ba/bb. However other related genes that are not teleost-specific duplicates, such as wnt2/2b, wnt3/3a, wnt5a/b, wnt10a/b, and wnt11/11r, have clearly different expression patterns indicating that their transcriptional regulation has likely diverged considerably. Investigations using double in situ hybridization and higher resolution analysis of specific CNS tissues can also be used to identify more subtle differences between grossly overlapping gene expression patterns. Expression of Wnt genes encoding ligands that signal through ß-catenin independent pathways also precludes simple predictions of redundant function. Importantly, both ß-catenin dependent and independent pathways play roles in stem cell maintenance and differentiation [33, 34], suggesting that post-embryonic neurogenesis may be regulated by multiple Wnt outputs in the same tissue.
The data presented here will be useful for the identification of candidate Wnt ligands that could mediate specific processes in CNS development, as demonstrated by the distinct subsets of Wnt genes expressed in consistent regional domains. For example, wnt1, wnt3, wnt3a, wnt7aa, and wnt10b are all localized to the roof plate of the diencephalon, midbrain and cerebellum. In contrast, wnt4a and wnt4b show consistent expression in floor plate structures, and wnt8b, wnt11r, and wnt16 are expressed in the hypothalamus. In addition, our analysis offers a starting point for further studies investigating the functions of Wnt ligands in developmental and post-embryonic neurogenesis. The identification of three ligands expressed in the hypothalamic posterior recess provides specific targets for manipulating Wnt pathway activity in a defined model of neural progenitor maintenance and differentiation, especially as mutant alleles are available for all three genes [35]. With the recent development of effective gene targeting methods in zebrafish [36], it will be possible to produce animals carrying null alleles for all the potential Wnt genes expressed in or near a given CNS tissue or cell population.
Acknowledgments
We thank Wilson Clements, Elke Ober, Matthias Carl, Gilbert Weidinger and Arne Lekven for generously sharing plasmid templates for in situ probes. R.I.D. was funded by NIH RO1NS082645 and R.N.D. was funded by NIDCD 5T32 DC008553.
Data Availability
All relevant data are within the paper and figures.
Funding Statement
This work was supported by National Institutes of Health, R01NS082645 (R.I.D.), T32DC008553 (R.N.D.). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Megason SG, McMahon AP. A mitogen gradient of dorsal midline Wnts organizes growth in the CNS. Development. 2002;129(9):2087–98. . [DOI] [PubMed] [Google Scholar]
- 2. Agathocleous M, Iordanova I, Willardsen MI, Xue XY, Vetter ML, Harris WA, et al. A directional Wnt/beta-catenin-Sox2-proneural pathway regulates the transition from proliferation to differentiation in the Xenopus retina. Development. 2009;136(19):3289–99. Epub 2009/09/09. 10.1242/dev.040451 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Munji RN, Choe Y, Li G, Siegenthaler JA, Pleasure SJ. Wnt signaling regulates neuronal differentiation of cortical intermediate progenitors. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2011;31(5):1676–87. Epub 2011/02/04. 10.1523/JNEUROSCI.5404-10.2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Wang X, Kopinke D, Lin J, McPherson AD, Duncan RN, Otsuna H, et al. Wnt signaling regulates postembryonic hypothalamic progenitor differentiation. Developmental cell. 2012;23(3):624–36. Epub 2012/09/15. 10.1016/j.devcel.2012.07.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Lie DC, Colamarino SA, Song HJ, Desire L, Mira H, Consiglio A, et al. Wnt signalling regulates adult hippocampal neurogenesis. Nature. 2005;437(7063):1370–5. Epub 2005/10/28. 10.1038/nature04108 . [DOI] [PubMed] [Google Scholar]
- 6. Imura T, Wang X, Noda T, Sofroniew MV, Fushiki S. Adenomatous polyposis coli is essential for both neuronal differentiation and maintenance of adult neural stem cells in subventricular zone and hippocampus. Stem Cells. 2010;28(11):2053–64. Epub 2010/11/20. 10.1002/stem.524 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Qu Q, Sun G, Li W, Yang S, Ye P, Zhao C, et al. Orphan nuclear receptor TLX activates Wnt/beta-catenin signalling to stimulate neural stem cell proliferation and self-renewal. Nat Cell Biol. 2010;12(1):31–40; sup pp 1–9. 10.1038/ncb2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Farin HF, Van Es JH, Clevers H. Redundant sources of Wnt regulate intestinal stem cells and promote formation of Paneth cells. Gastroenterology. 2012;143(6):1518–29 e7. 10.1053/j.gastro.2012.08.031 . [DOI] [PubMed] [Google Scholar]
- 9. Ikeya M, Lee SMK, Johnson JE, McMahon AP, Takada S. Wnt signalling required for expansion of neural crest and CNS progenitors. Nature. 1997;389(6654):966–70. [DOI] [PubMed] [Google Scholar]
- 10. Stoick-Cooper CL, Weidinger G, Riehle KJ, Hubbert C, Major MB, Fausto N, et al. Distinct Wnt signaling pathways have opposing roles in appendage regeneration. Development. 2007;134(3):479–89. . [DOI] [PubMed] [Google Scholar]
- 11. Fotaki V, Larralde O, Zeng S, McLaughlin D, Nichols J, Price DJ, et al. Loss of Wnt8b has no overt effect on hippocampus development but leads to altered Wnt gene expression levels in dorsomedial telencephalon. Dev Dyn. 2010;239(1):284–96. 10.1002/dvdy.22137 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Lu FI, Thisse C, Thisse B. Identification and mechanism of regulation of the zebrafish dorsal determinant. Proc Natl Acad Sci U S A. 2011;108(38):15876–80. 10.1073/pnas.1106801108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Marz M, Chapouton P, Diotel N, Vaillant C, Hesl B, Takamiya M, et al. Heterogeneity in progenitor cell subtypes in the ventricular zone of the zebrafish adult telencephalon. Glia. 2010;58(7):870–88. 10.1002/glia.20971 . [DOI] [PubMed] [Google Scholar]
- 14. Grandel H, Kaslin J, Ganz J, Wenzel I, Brand M. Neural stem cells and neurogenesis in the adult zebrafish brain: origin, proliferation dynamics, migration and cell fate. Dev Biol. 2006;295(1):263–77. Epub 2006/05/10. S0012-1606(06)00247-8 [pii] 10.1016/j.ydbio.2006.03.040 . [DOI] [PubMed] [Google Scholar]
- 15. Ito Y, Tanaka H, Okamoto H, Ohshima T. Characterization of neural stem cells and their progeny in the adult zebrafish optic tectum. Developmental biology. 2010;342(1):26–38. Epub 2010/03/30. 10.1016/j.ydbio.2010.03.008 [DOI] [PubMed] [Google Scholar]
- 16. Leucht C, Stigloher C, Wizenmann A, Klafke R, Folchert A, Bally-Cuif L. MicroRNA-9 directs late organizer activity of the midbrain-hindbrain boundary. Nat Neurosci. 2008;11(6):641–8. 10.1038/nn.2115 . [DOI] [PubMed] [Google Scholar]
- 17. Kaslin J, Ganz J, Geffarth M, Grandel H, Hans S, Brand M. Stem cells in the adult zebrafish cerebellum: initiation and maintenance of a novel stem cell niche. J Neurosci. 2009;29(19):6142–53. Epub 2009/05/15. 29/19/6142 [pii] 10.1523/JNEUROSCI.0072-09.2009 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Wehman AM, Staub W, Meyers JR, Raymond PA, Baier H. Genetic dissection of the zebrafish retinal stem-cell compartment. Dev Biol. 2005;281(1):53–65. 10.1016/j.ydbio.2005.02.010 . [DOI] [PubMed] [Google Scholar]
- 19. Kimmel CB, Ballard WW, Kimmel SR, Ullmann B, Schilling TF. Stages of embryonic development of the zebrafish. Dev Dyn. 1995;203(3):253–310. . [DOI] [PubMed] [Google Scholar]
- 20. Dorsky RI, Moon RT, Raible DW. Control of neural crest cell fate by the Wnt signalling pathway. Nature. 1998;396(6709):370–3. [DOI] [PubMed] [Google Scholar]
- 21. Veien ES, Rosenthal JS, Kruse-Bend RC, Chien CB, Dorsky RI. Canonical Wnt signaling is required for the maintenance of dorsal retinal identity. Development. 2008;135(24):4101–11. Epub 2008/11/14. 10.1242/dev.027367 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Ober EA, Verkade H, Field HA, Stainier DY. Mesodermal Wnt2b signalling positively regulates liver specification. Nature. 2006;442(7103):688–91. 10.1038/nature04888 . [DOI] [PubMed] [Google Scholar]
- 23. Clements WK, Ong KG, Traver D. Zebrafish wnt3 is expressed in developing neural tissue. Dev Dyn. 2009;238(7):1788–95. 10.1002/dvdy.21977 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Ungar AR, Kelly GM, Moon RT. Wnt4 affects morphogenesis when misexpressed in the zebrafish embryo. Mech Dev. 1995;52(2–3):153–64. [DOI] [PubMed] [Google Scholar]
- 25. Carl M, Bianco IH, Bajoghli B, Aghaallaei N, Czerny T, Wilson SW. Wnt/Axin1/beta-catenin signaling regulates asymmetric nodal activation, elaboration, and concordance of CNS asymmetries. Neuron. 2007;55(3):393–405. Epub 2007/08/07. 10.1016/j.neuron.2007.07.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Kelly GM, Greenstein P, Erezyilmaz DF, Moon RT. Zebrafish wnt8 and wnt8b share a common activity but are involved in distinct developmental pathways. Development. 1995;121(6):1787–99. [DOI] [PubMed] [Google Scholar]
- 27. Ulrich F, Concha ML, Heid PJ, Voss E, Witzel S, Roehl H, et al. Slb/Wnt11 controls hypoblast cell migration and morphogenesis at the onset of zebrafish gastrulation. Development. 2003;130(22):5375–84. 10.1242/dev.00758 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Jing L, Lefebvre JL, Gordon LR, Granato M. Wnt signals organize synaptic prepattern and axon guidance through the zebrafish unplugged/MuSK receptor. Neuron. 2009;61(5):721–33. Epub 2009/03/17. 10.1016/j.neuron.2008.12.025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Thisse C, Thisse B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nature protocols. 2008;3(1):59–69. 10.1038/nprot.2007.514 . [DOI] [PubMed] [Google Scholar]
- 30. Beretta CA, Brinkmann I, Carl M. All four zebrafish Wnt7 genes are expressed during early brain development. Gene Expr Patterns. 2011;11(3–4):277–84. 10.1016/j.gep.2011.01.004 . [DOI] [PubMed] [Google Scholar]
- 31. Mueller T, Wullimann MF. Anatomy of neurogenesis in the early zebrafish brain. Brain Res Dev Brain Res. 2003;140(1):137–55. . [DOI] [PubMed] [Google Scholar]
- 32. Postlethwait JH, Yan YL, Gates MA, Horne S, Amores A, Brownlie A, et al. Vertebrate genome evolution and the zebrafish gene map. Nat Genet. 1998;18(4):345–9. . [DOI] [PubMed] [Google Scholar]
- 33. Clements WK, Kim AD, Ong KG, Moore JC, Lawson ND, Traver D. A somitic Wnt16/Notch pathway specifies haematopoietic stem cells. Nature. 2011;474(7350):220–4. 10.1038/nature10107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Katoh M. WNT signaling in stem cell biology and regenerative medicine. Current drug targets. 2008;9(7):565–70. . [DOI] [PubMed] [Google Scholar]
- 35. Gordon LR, Gribble KD, Syrett CM, Granato M. Initiation of synapse formation by Wnt-induced MuSK endocytosis. Development. 2012;139(5):1023–33. Epub 2012/02/10. 10.1242/dev.071555 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Hwang WY, Fu Y, Reyon D, Maeder ML, Tsai SQ, Sander JD, et al. Efficient genome editing in zebrafish using a CRISPR-Cas system. Nat Biotechnol. 2013;31(3):227–9. 10.1038/nbt.2501 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the paper and figures.