

Abstract
Autophagy is an evolutionarily conserved self-digestion process for the quality control of intracellular entities in eukaryotes. In the past few years, mounting evidence indicates that microRNAs (miRNAs)-mediated post-transcriptional regulation of gene expression represents an integral part of the autophagy regulatory network and may have a substantial effect on autophagy-related physiological and pathological conditions including cancer. Herein, we examine some of the molecular mechanisms by which miRNAs manipulate the autophagic machinery to maintain cellular homeostasis and their biological outputs during cancer development. A better understanding of interaction between miRNAs and cellular autophagy may ultimately benefit future cancer diagnostic and anticancer therapeutics.
Keywords: MicroRNA, Autophagy, Cancer
INTRODUCTION
The discovery of cellular microRNAs (miRNAs), a class of evolutionarily conserved short non-coding RNAs, has revealed an entirely new layer of post-transcriptional gene regulation [1]. miRNAs are present either as single units throughout the genome or in gene clusters and play a critical role in various biological processes. Expression of miRNAs reshape various aspect of cancer-related events such as cell transformation, apoptosis, metastasis, inflammation, and immunity [2]. Depending on the tissue context and target genes, cellular miRNAs can act as oncogenes or tumor suppressors when mis-expressed or de-regulated [2]. The first identified miRNA tumor suppressors are miR-15a and miR-16-1, which target the pro-survival Bcl-2 family members and were frequently lost in chronic lymphocytic leukemia (CLL) [3, 4]. Since then, numerous tumor-associated miRNAs have been uncovered to date, yet only a handful of them have their targets validated and even less have the relevant mechanisms addressed. Given that a single miRNA may target a multitude of transcripts and that one certain gene may influence multiple miRNA expression, elucidating tissue-specific miRNA expression and function remains a challenge in miRNA research. Recent development of high throughput discovery tools such as next-generation sequencing and integrative genomics provide opportunities to identify mechanistic links between deregulated miRNAs and cancer-related pathways [5, 6]. Significantly, it is now being appreciated that miRNAs serve as novel and potent regulators of the autophagy pathway [7]. The miRNA-autophagy crosstalk reprograms the biological functions of autophagy during cancer development, and also sheds lights on new targets for anticancer therapies. In this review, we focus on recent findings about the multifaceted roles of miRNAs in autophagy regulation and its significance in cancer.
THE AUTOPHAGY MACHINERY
Macroautophagy (referred to as autophagy), initially described as a lysosome-dependent degradation of cytoplasmic contents on starvation, has since been implicated in almost every facet of human health and can be linked to a myriad of human diseases including cancer [8]. Over 30 distinct autophagy-related (Atg) genes have been identified so far in yeast, and many of them have orthologs in mammals [9]. Importantly, a growing body of evidence shows that, in addition to their role in regulating autophagy, Atg proteins can function in different metabolic and immunologic pathways, highlighting intricate connection and coordination of autophagy with other signaling networks. While constitutively active at basal levels to maintain homeostasis, autophagy is upregulated under conditions of cellular stress [8]. During autophagy, portions of cytoplasmic proteins and/or organelles are sequestered within characteristic double-membrane-bound autophagosomes and delivered to lysosomes for bulk degradation [10]. A complete autophagic process can be divided into several distinct stages, including induction, vesicle nucleation, autophagosome membrane elongation, fusion, and degradation (Figure 1) [8, 9]. Simply stated, autophagy induction is controlled by the gatekeeper, the mammalian target of rapamycin complex 1 (mTOC1), which can sense and integrates stress signals from various sources including growth factors, amino acids, hypoxia, and energy levels. Under normal conditions, mTORC1 is active and represses autophagy by precluding the assembly of the autophagy-initiating ULK1 (human homolog of ATG1) kinase complex, including ULK1, Atg13, FIP200, and Atg101 [11, 12]. Stress-induced inactivation of mTORC1 enables the assembly and activation of the ULK1 complex to initiate the autophagy cascade. Following induction, autophagy signals are resolved in space to the phagophore-forming sites rich in phosphatidylinositol-3-phosphate (PI3P), which can be derived from multiple pre-existing membrane sources including endoplasmic reticulum, Golgi stacks, endosome, as well as nuclear and plasma membrane [13–16]. Notably, phagophore nucleation is driven by the Beclin1-associated PI3P-kinase class III (PI3KC3) complex, whereby Beclin1 acts as a scaffold to recruit both activating cofactors including Vps15, Atg14L, UVRAG, Bif-1, and Ambra-1, as well as inhibiting factors including Bcl-2 for a balanced regulation [17]. Growth and ultimate sealing of the autophagosome membrane engage two ubiquitin-like conjugation systems, LC3-phophatidylethanolamine (PE) conjugation and Atg12-Atg5 conjugation [18]. After proteolytic cleavage by the Atg4 cysteine protease, LC3 is conjugated to the membrane lipid PE (the conjugated form is termed LC3-II) after sequential processing by Atg7 (E1) and Atg3 (E2). In parallel, ubiquitin-like Atg12 is conjugated to Atg5 in a similar manner except that Atg10, instead of Atg3 serves as the E2 enzyme [18, 19]. Once formed, most of the autophagy machinery can be recycled in the cytosol with the exception of a proportion of LC3-II that is retained on the autophagosome membrane until the final lysosome fusion is executed. Finally, the autophagosome ‘matures’ through multiple transient interaction and membrane fusion with the endosome–lysosome network to form a hybrid-like organelle called the autolysosome, a pivotal checkpoint for the overall autophagy flux and a convergence point of endocytic trafficking. Recent studies have uncovered multiple factors that function in the maturation process of autophagosome including but not limiting to SNARE proteins like VAMP7, tethering factors like the class C vacuolar protein sorting (Vps) complex, Rab family members such as Rab7, Atg9, Rubicon-containing PI3KC3 complex, and lysosome membrane proteins all seem involved to different extent (Figure 1) [20–25]. Consequently, the encapsulated materials, together with the inner membrane of the autophagosome, undergo breakdown by lysosomal hydrolases and retrieval [26]. Importantly, almost every stage of the autophagy pathway is recently found to be exquisitely manipulated by miRNAs, including both oncogenic and tumor suppressive ones, which reflects the substantial significance of miRNA-regulated autophagy during cancer development. For clarity, we discuss the many interactions of cancer-related miRNAs and the autophagy pathway in relation to the linear cascade of the autophagy process (Figure 1).
Figure 1.
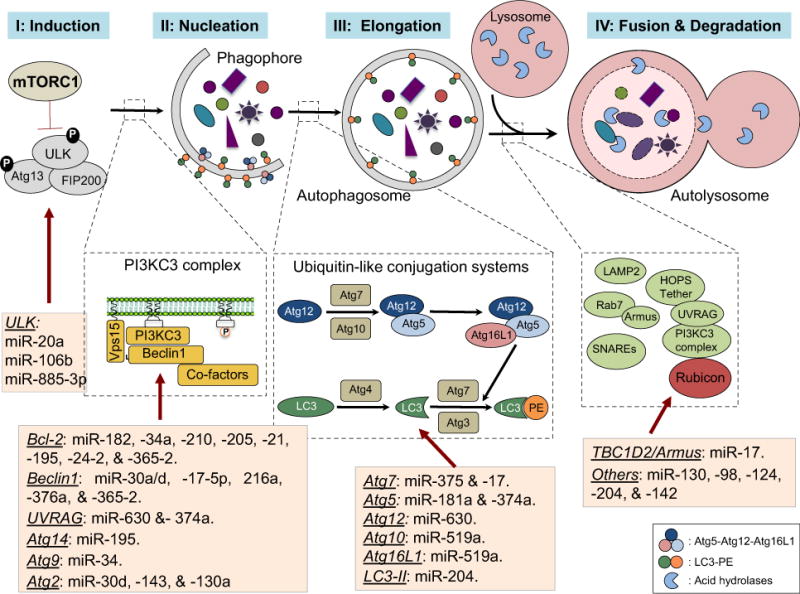
Schematic representation of the core autophagy pathway and its regulation by miRNAs. Autophagy proceeds through a series of steps including induction, vesicle nucleation, membrane elongation, and autophagosome maturation into the lysosome for degradation and recycling. miRNAs regulation occurs at each dynamic step. Autophagy induction is directly and tightly controlled by the ‘nutrient sensor’ mTORC1, which under nutrient-rich condition precludes the assembly and the activation of the autophagy-initiating ULK complex comprising ULK1/2, Atg13, and FIP200 (Step 1). mTORC1 is inactivated in response to various stimuli (for instance, starvation, hypoxia), which enables the activation of the ULK complex and triggers the autophagy cascade. MiR-20a, miR106b, and miR-885-3P target ULK1/2 and suppress autophagy. Phagophore nucleation, which is confined to the phosphatidylinositol-3-phosphate (PI3P)-containing vesicles, is driven by the Beclin1-associated PI3KC3 complex (Step 2). The activity of this kinase complex is suppressed by the anti-apoptotic Bcl-2 proteins, a functional target of multiple miRNAs such as miR-182, miR-34a, miR-210, miR-205, miR-21, miR-195, miR-24-2, and miR-365-2. Beclin1 can also be targeted by miR-30a, miR-30d, miR-17-5p, miR-216a, miR-376a, and miR-365-2. In addition, UVRAG is a direct and functional target of miR-630 and miR-374a, while Atg14 being inhibited by miR-195. Autophagosome membrane elongation involves two ubiquitin-like conjugation systems: one is Atg12-Atg5 conjugation and the other is LC3-phosphatidylethanolamine (PE) conjugation, the latter of which is sequentially processed by Atg4, Atg7, and Atg3 (Step 3). Atg7 can be suppressed by miR-375 and miR-17. LC3-II levels are posttranscriptionally controlled by miR-204. Fusion of the autophagosome with the lysosome signifies the maturation stage of the autophagy pathway, a step that involves multiple membrane fusion factors and is negatively regulated by the Rubicon complex (Rubicon-UVRAG-Beclin1-PI3KC3-p150). Finally, the encapsulated contents, together with the inner membrane of the autophagosome, undergo breakdown by lysosomal hydrolases.
TARGETING AUTOPHAGY INDUCTION
One of the initial events upon autophagy induction is the activation of the ULK1 complex, which is directly controlled by mTORC1 (Figure 1). miRNAs could interfere with upstream autophagy signaling by targeting mTORC1 or the ULK1 complex. Indeed, oncogenic miR-20a and miR-106b have been found to target the 3′UTR of ULK1 and blunt autophagy induced by leucine deprivation in C2C12 myoblasts [27]. More recent findings further established that another Atg1 homologue, ULK2, which can compensate for the loss of ULK1 function in certain cell types [28], was inhibited by miR-885-3p mostly in response to chemotherapeutic drugs [29]. Intriguingly, the miR-885-3p recognition sequences have been detected in multiple apoptosis- and autophagy-related genes including MDM4, Bcl2, Casp2, and Casp3, etc., highlighting a complex tumor cell response to drug treatment [29]. It remains to be determined whether inhibition of autophagy induction by miRNAs serves to preclude prolonged and unleashed autophagy initiation in normal conditions. Nevertheless, targeting mTORC1-ULK1 signaling may be an important negative regulation to suppress the early stage of self-survival autophagy, thereby sensitizing some cancer cells to cell death.
TARGETING VESICLE NUCLEATION
Vesicle nucleation of the phagophore is driven by the activation of the Beclin1-associated PI3KC3 complex (Figure 1). This process is suppressed by the anti-apoptotic BCL-2 proteins such as Bcl-2, Bcl-xL, and Bcl-w, and is promoted by the pro-autophagic proteins Atg14 and UVRAG, all targeting Beclin1 during autophagosome formation [17]. It is thus plausible to speculate that miRNAs inhibiting Bcl-2 may positively regulate autophagy, whereas those targeting Atg14, UVRAG, or Beclin1 may inhibit autophagy. Of note, a growing list of tumor suppressive miRNAs have been identified to regulate Bcl-2 at their respective binding sites in Bcl-2, including miR-182, miR-34a, miR-210, miR-205, miR-21, miR-195, miR-24-2, and miR-365-2 in different tumor contexts [30, 31]. However, most studies on these miRNAs focus on their ability to post-transcriptionally inhibit Bcl-2 expression and thereof facilitate the mitochondrial release of apoptogenic factors, which can augment drug-induced apoptosis [31]. At present, it is not clear whether these anti-Bcl-2 miRNAs have the comparable capability to activate autophagy; however, owing to the fact that apoptosis is not the sole action of Bcl-2 and that autophagy also participates in BCL-2-mediated tumorigenesis [32], it seems likely that these pro-apoptotic miRNAs targeting Bcl-2 also function as pro-autophagic factors. Apart from the tumor suppressive anti-Bcl-2 miRNAs, Bcl-2 expression is also upregulated by the oncogenic miR-21 that is amplified in various tumors including breast cancer [33]. In line with this, recent study reported that targeted elimination of miR-21 triggers autophagy and increase drug sensitivity in leukemia cells [34]. As autophagy attenuation is critical for tumor maintenance, miRNA-mediated dysregulation of autophagy might contribute to the oncogenic mechanism in some cancer.
Unlike Bcl-2 that acts as a negative regulator of autophagy, Beclin1 serves as a core autophagy effector by forming a complex with PI3KC3 to directly mediate autophagosome formation (Figure 1). Not surprisingly, expression of Beclin1 is targeted by several miRNAs, including but likely not limiting to miR-30a [35], miR-30d [36], miR-17 [37], miR-216a [38], miR-376a [39, 40], and miR-365-2 [41]. Among them, miR-376a appears to act in rapamycin- or starvation-induced autophagy by targeting both Atg4 and Beclin1 [40], whereas miR-30a, miR-30d, and miR17 are cancer-related Beclin1 inhibitors and determine the sensitivity of cancer cells to chemotherapy [35–37, 39]. A most recent study by Chatterjee et al., [42] demonstrated that combined expression of both miR-16, which inhibits Bcl-2-mediated anti-apoptosis, and miR-17, which antagonizes Beclin1-mediated cytoprotective autophagy, significantly sensitized paclitaxel-resistant lung cancer cells, highlighting miRNAs a promising target for therapeutic improvement in cancer. Beyond tumor development, miR-216a can control oxidized low-density lipoprotein-induced autophagy in endothelial cells by suppressing intracellular levels of Beclin1 and thus plays a role in some cardiovascular disorders and atherosclerosis [38]. Given that the same Beclin1 target may have distinct functions beyond autophagy in different cellular contexts and disease conditions, it remains to be confirmed whether phenotypic changes associated with Beclin1-targeted miRNAs are due to pathogenic or bystander effects of autophagy.
Additionally, the autophagy promoters, UVRAG and Atg14, are also controlled by miRNAs. Specifically, miR-630 and miR-374a can inhibit the tumor suppressor UVRAG [43]. In line with this, increased miR-630 expression has been observed in gastric cancer and correlated to poor survival [44], which to some extent may explain deficient UVRAG expression in some human cancers. It should be noted, however, UVRAG is not the sole target of miR-630 and several genes in the DNA damage-signaling pathway are also deregulated in miR-630-expressing cells [45]. Atg14, which forms a mutually exclusive complex with Beclin1-PI3KC3 from the one containing UVRAG, was identified as a functional target for miR-195 [46]. Intriguingly, miR-195 inhibited Atg14-mediated autophagic process functions in neuroinflammation in vivo and in vitro [46]. Inhibition of miR-195 can boost autophagy and alleviate neuropathic pain following peripheral nerve injury, pointing to a connection of the autophagy pathway with cells’ inflammatory response.
Finally, Rab5A, a small GTPase in endocytic trafficking, can induce the early stage of autophagosome formation through its interaction with the Beclin1-PI3KC3 complex and is targeted by miR-101 [47]. Other autophagy-related targets of miR-101 also include stathmin 1 (STMN1) and Atg4D [47], suggesting its potent role in autophagy inhibition. Consistent with this, miR-101 was recently found to block chemotherapy-induced autophagy in osteosarcoma cells to increase therapeutic response [48]. Autophagosome biogenesis is also regulated by Atg2-Atg18 and Atg9-mediated membrane retrieval process [49]. Atg2 has been identified as a direct target of miR-30d [50], miR-143 [51], and miR-130a [51], while Atg9 is downregulated by miR-34 in C. elegans [52]. Expression of miR-143 in colon cancer cells caused anti-proliferative and chemo-sensitizing effect to 5-fluorouracil treatment, but it is not certain whether miR-143-mediated Atg2 suppression and autophagy inhibition plays a role in this regard [53].
TARGETING AUTOPHAGOSOME MEMBRANE ELONGATION
In addition to targeting autophagy induction and nucleation, miRNAs also modulate the expansion of autophagosome by targeting both the E1-like (i.e. Atg7) and E2-like enzymes (i.e. Atg10 and Atg3) and ubiquitin-like substrates (i.e. Atg12 and LC3) in the two conjugation processes (Figure 1) [18]. Recent data suggest that miR-375 can suppress autophagy by targeting Atg7, independently of its regulation of oncogenic AKT/mTORC1 signaling [54]. Although autophagy is generally considered as a tumor suppressor mechanism, miR-375-mediated anti-autophagy disrupted the ability of tumor cells to survive metabolic stresses and thereof sensitized sensitizes hepatocellular carcinoma cells to hypoxia-induced cell death [54]. Interestingly, downregulation of another Atg7-targeted miR-17 caused autophagy activation, and paradoxically decreased the threshold resistance in human glioblastoma cells to chemotherapeutic treatments [55]. These seemingly opposite effects of miRNA-mediated autophagy inhibition on tumor growth may reflect the tissue-specific manner in which autophagy is re-programmed by tumors. Although the regulation of most autophagy-related miRNAs expression in tumors remains unclear, it seems evident that drug treatment can considerably reprogram the miRNA expression profile. For instance, Cisplatin has been found to decrease the levels of a panel of autophagy-related miRNAs, such as miR-181a and miR-374a against Atg5, miR-630 against Atg12, and miR-519a against Atg10 and Atg16L1 in a ΔNp63α-dependent manner in squamous cell carcinoma (SCC) cells [43], suggesting that autophagy regulation by miRNAs plays an undoubtedly important role in anti-tumor response.
Being an authentic autophagosome marker, LC3-II levels are regulated by miR-204 and its autophagy-suppressive effect was first recognized in cardiomyocytes during ischemia-reperfusion injury [56], and was later confirmed in the study of renal clear cell carcinoma (ccRCC) [57]. Mikhaylova et al. observed that higher tumor grade of human ccRCC was correlated with a concomitant reduction in miR-204 levels and increase in LC3-II levels, which is controlled by the VHL tumor suppressor [57]. This study revealed the addiction of cancer cells to autophagy, likely to ameliorate stressed conditions for continued survival and proliferation. Yet, whether such change in autophagy represents a by-product of malignant transformation or represents its founding cause remains to be determined. Further investigation in this issue may delineate complex interrelationship between tumor suppressor, miRNA, and autophagy in cancer development. Beside autophagosome-conjugated LC3-II, an inverse correlation of the levels of SQSTM1/p62, an autophagy adaptor and ubiquitin-binding protein, with miR-106 expression was also detected in acute myeloid leukemia [58]. Knockdown of SQSTM1 phenocopied the effect of miR-106 and induced cell expansion of myeloid cells through a mechanism involving the endosome-related but not autophagosome-related protein degradation [58]. It is thus worth noting that miRNAs against autophagy-related factors may confer autophagy-independent tumor suppressive or oncogenic activities.
TARGETING AUTOPHAGOSOME MATURATION
The autophagosome matures by fusion with the late endosome and the lysosome to form the autoloysosome, a process that requires the membrane fusion machinery including the Rab-SNARE system, the tethering HOPS complex, as well as some lysosomal membrane proteins such as LAMP1 and LAMP2 (Figure 1). TBC1D2/Armus, a GAP of the late endosome-related Rab7 GTPase, was recently found to be suppressed by miR-17 in both endocytic and autophagic maturation into the lysosome [59]. Currently, little is known about what “prepares” the autophagosome for fusion with a target vesicle and what relevant “signals” are involved. However, recent studies revealed a negative regulation of autophagosome maturation mediated by Rubicon (RUN domain Beclin1-interacting and cysteine-rich containing protein) in complex with UVRAG, Beclin1 and other members of the PI3KC3 complex (Figure 1). Yet, no Rubicon-targeted miRNAs have been identified to promote autophagic turnover. Notably, a recent system biology-based approach identified several miRNAs such as miR-130, miR-98, miR-124, miR-204, and miR-142 as the putative regulators of the autophagy-lysosomal genes [60], albeit the in vivo relevance of these miRNA-mediated inhibitions of autophagic degradation has not been investigated.
CONCLUSIONS
In summary, the involvement of different miRNAs in various steps of autophagy suggests the importance of autophagy in cellular homeostasis and disease conditions. Although it is still a matter of debate whether miRNAs-regulated autophagy reflects ultimately a protective response or a detrimental process during cancer development, progression, and treatment, it seems clear that the autophagy pathway does not exist in isolation but is integrated with other cell signaling networks cross-regulated by miRNAs. Strikingly, in addition to being regulated by miRNAs, autophagy can selectively regulates the abundance and function of miRNAs by a NDP52-mediated targeted degradation of the miRNA-processing enzyme, DICER1, and the main miRNA effector, AGO [61]. This intriguing result emphasizes the fact that our knowledge of miRNA interaction with autophagy is far from complete. Furthermore, how these new regulatory functions of miRNAs and autophagy are fine-tuned in specific contexts of tumors and how they are translated into to the pathological conditions, remain unknown. Thus, an understanding of the complexity and specificity of miRNAs functions in autophagy regulation and technologies that allow the specific manipulation of miRNA function could have the potential to inform new therapeutic strategies for cancer prevention, diagnosis, and treatment in the future.
Acknowledgments
Work in the authors’ laboratory is funded by the by American Cancer Society (RSG-11-121-01-CCG to C. Liang) and National Institutes of Health grants (CA140964 and CA161436 to C. Liang)
Footnotes
COMPETING INTERESTS
The authors declare no competing interests.
References
- 1.Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–97. doi: 10.1016/S0092-8674(04)00045-5. [DOI] [PubMed] [Google Scholar]
- 2.Di Leva G, Croce CM. Roles of small RNAs in tumor formation. Trends Mol Med. 2010;16:257–67. doi: 10.1016/j.molmed.2010.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Calin GA, Dumitru CD, Shimizu M, Bichi R, Zupo S, Noch E, Aldler H, Rattan S, Keating M, Rai K, Rassenti L, Kipps T, Negrini M, Bullrich F, Croce CM. Nonlinear partial differential equations and applications: frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc Natl Acad Sci USA. 2002;99:15524–9. doi: 10.1073/pnas.242606799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Cimmino A, Calin GA, Fabbri M, Iorio MV, Ferracin M, Shimizu M, Wojcik SE, Aqeilan RI, Zupo S, Dono M, Rassenti L, Alder H, Volinia S, Liu C-G, Kipps TJ, Negrini M, Croce CM. miR-15 and miR-16 induce apoptosis by targeting BCL2. Proc Natl Acad Sci USA. 2005;102:13944–9. doi: 10.1073/pnas.0506654102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Martens-Uzunova ES, Jalava SE, Dits NF, van Leenders GJ, Møller S, Trapman J, Bangma CH, Litman T, Visakorpi T, Jenster G. Diagnostic and prognostic signatures from the small non-coding RNA transcriptome in prostate cancer. Oncogene. 2012;31:978–91. doi: 10.1038/onc.2011.304. [DOI] [PubMed] [Google Scholar]
- 6.Farazi TA, Horlings HM, Ten Hoeve JJ, Mihailovic A, Halfwerk H, Morozov P, Brown M, Hafner M, Reyal F, van Kouwenhove M, Kreike B, Sie D, Hovestadt V, Wessels LFA, Van De Vijver MJ, Tuschl T. MicroRNA sequence and expression analysis in breast tumors by deep sequencing. Cancer Res. 2011;71:4443–53. doi: 10.1158/0008-5472.CAN-11-0608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Fu LL, Wen X, Bao JK, Liu B. MicroRNA-modulated autophagic signaling networks in cancer. Int J Biochem Cell Biol. 2012;44:733–6. doi: 10.1016/j.biocel.2012.02.004. [DOI] [PubMed] [Google Scholar]
- 8.Levine B, Klionsky DJ. Development by self-digestion: molecular mechanisms and biological functions of autophagy. Dev Cell. 2004;6:463–77. doi: 10.1016/S1534-5807(04)00099-1. [DOI] [PubMed] [Google Scholar]
- 9.Klionsky DJ. Cell biology: regulated self-cannibalism. Nature. 2004;431:31–2. doi: 10.1038/431031a. [DOI] [PubMed] [Google Scholar]
- 10.Mizushima N, Komatsu M. Autophagy: renovation of cells and tissues. Cell. 2011;147:728–41. doi: 10.1016/j.cell.2011.10.026. [DOI] [PubMed] [Google Scholar]
- 11.Jung CH, Jun CB, Ro SH, Kim YM, Otto NM, Cao J, Kundu M, Kim DH. ULK-Atg13-FIP200 complexes mediate mTOR signaling to the autophagy machinery. Mol Biol Cell. 2009;20:1992–2003. doi: 10.1091/mbc.E08-12-1249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Mizushima N. The role of the Atg1/ULK1 complex in autophagy regulation. Curr Opin Cell Biol. 2010;22:132–9. doi: 10.1016/j.ceb.2009.12.004. [DOI] [PubMed] [Google Scholar]
- 13.Yen WL, Legakis JE, Nair U, Klionsky DJ. Atg27 Is Required for Autophagy-dependent Cycling of Atg9. Mol Biol Cell. 2007;18:581–93. doi: 10.1091/mbc.E06-07-0612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hailey DW, Rambold AS, Satpute-Krishnan P, Mitra K, Sougrat R, Kim PK, Lippincott-Schwartz J. Mitochondria supply membranes for autophagosome biogenesis during starvation. Cell. 2010;141:656–67. doi: 10.1016/j.cell.2010.04.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ravikumar B, Moreau K, Jahreiss L, Puri C, Rubinsztein DC. Plasma membrane contributes to the formation of pre-autophagosomal structures. Nat Cell Biol. 2010;12:747–57. doi: 10.1038/ncb2078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Reggiori F, Klionsky DJ. Autophagosomes: biogenesis from scratch? Curr Opin Cell Biol. 2005;17:415–22. doi: 10.1016/j.ceb.2005.06.007. [DOI] [PubMed] [Google Scholar]
- 17.He C, Levine B. The Beclin 1 interactome. Curr Opin Cell Biol. 2010;22:140–9. doi: 10.1016/j.ceb.2010.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Yang Z, Klionsky DJ. Mammalian autophagy: core molecular machinery and signaling regulation. Curr Opin Cell Biol. 2010;22:124–31. doi: 10.1016/j.ceb.2009.11.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Levine B, Kroemer G. Autophagy in the pathogenesis of disease. Cell. 2008;132:27–42. doi: 10.1016/j.cell.2007.12.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Eskelinen EL. Maturation of autophagic vacuoles in Mammalian cells. Autophagy. 2005;1:1–10. doi: 10.4161/auto.1.1.1270. [DOI] [PubMed] [Google Scholar]
- 21.Nara A, Mizushima N, Yamamoto A, Kabeya Y, Ohsumi Y, Yoshimori T. SKD1 AAA ATPase-dependent endosomal transport is involved in autolysosome formation. Cell Struct Funct. 2002;27:29–37. doi: 10.1247/csf.27.29. [DOI] [PubMed] [Google Scholar]
- 22.Tamai K, Tanaka N, Nara A, Yamamoto A, Nakagawa I, Yoshimori T, Ueno Y, Shimosegawa T, Sugamura K. Role of Hrs in maturation of autophagosomes in mammalian cells. Biochem Biophys Res Commun. 2007;360:721–7. doi: 10.1016/j.bbrc.2007.06.105. [DOI] [PubMed] [Google Scholar]
- 23.Liang C, Lee JS, Inn KS, Gack MU, Li Q, Roberts EA, Vergne I, Deretic V, Feng P, Akazawa C, Jung JU, et al. Beclin1-binding UVRAG targets the class C Vps complex to coordinate autophagosome maturation and endocytic trafficking. Nat Cell Biol. 2008;10:776–87. doi: 10.1038/ncb1740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Virgin HW, Levine B. Autophagy genes in immunity. Nat Immunol. 2009;10:461–70. doi: 10.1038/ni.1726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Zhong Y, Wang QJ, Li X, Yan Y, Backer JM, Chait BT, Heintz N, Yue Z. Distinct regulation of autophagic activity by Atg14L and Rubicon associated with Beclin 1–phosphatidylinositol-3-kinase complex. Nat Cell Biol. 2009;11:468–76. doi: 10.1038/ncb1854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ganley IG. Autophagosome maturation and lysosomal fusion. Essays Biochem. 2013;55:65–78. doi: 10.1038/nature11910. [DOI] [PubMed] [Google Scholar]
- 27.Wu H, Wang F, Hu S, Yin C, Li X, Zhao S, Wang J, Yan X. MiR-20a and miR-106b negatively regulate autophagy induced by leucine deprivation via suppression of ULK1 expression in C2C12 myoblasts. Cell Signal. 2012;24:2179–86. doi: 10.1016/j.cellsig.2012.07.001. [DOI] [PubMed] [Google Scholar]
- 28.Mizushima N. The role of the Atg1/ULK1 complex in autophagy regulation. Curr Opin Cell Biol. 2010;22:132–9. doi: 10.1016/j.ceb.2009.12.004. [DOI] [PubMed] [Google Scholar]
- 29.Huang Y, Chuang AY, Ratovitski EA. Phospho-DeltaNp63alpha/miR-885-3p axis in tumor cell life and cell death upon cisplatin exposure. Cell Cycle. 2011;10:3938–47. doi: 10.4161/cc.10.22.18107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Jing Z, Han W, Sui X, Xie J, Pan H. Interaction of autophagy with microRNAs and their potential therapeutic implications in human cancers. Cancer Lett. 2015;356:332–8. doi: 10.1016/j.canlet.2014.09.039. [DOI] [PubMed] [Google Scholar]
- 31.Singh R, Saini N. Downregulation of BCL2 by miRNAs augments drug-induced apoptosis – a combined computational and experimental approach. J Cell Sci. 2012;125:1568–78. doi: 10.1242/jcs.095976. [DOI] [PubMed] [Google Scholar]
- 32.Pattingre S, Levine B. Bcl-2 inhibition of autophagy: a new route to cancer? Cancer Res. 2006;66:2885–8. doi: 10.1158/0008-5472.CAN-05-4412. [DOI] [PubMed] [Google Scholar]
- 33.Song B, Wang C, Liu J, Wang X, Lv L, Wei L, Xie L, Zheng Y, Song X. MicroRNA-21 regulates breast cancer invasion partly by targeting tissue inhibitor of metalloproteinase 3 expression. J Exp Clin Cancer Res. 2010;29:29. doi: 10.1186/1756-9966-29-29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Seca H, Lima RT, Lopes-Rodrigues V, Guimaraes JE, Almeida GM, Vasconcelos MH. Targeting miR-21 induces autophagy and chemosensitivity of leukemia cells. Curr Drug Targets. 2013;14:1135–43. doi: 10.2174/13894501113149990185. [DOI] [PubMed] [Google Scholar]
- 35.Zhu H, Wu H, Liu X, Li B, Chen Y, Ren X, Liu CG, Yang JM. Regulation of autophagy by a beclin 1-targeted microRNA, miR-30a, in cancer cells. Autophagy. 2009;5:816–23. doi: 10.4161/auto.9064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Zhang Y, Yang WQ, Zhu H, Qian YY, Zhou L, Ren YJ, Ren XC, Zhang L, Liu XP, Liu CG, Ming ZJ, Li B, Chen B, Wang JR, Liu YB, Yang JM. Regulation of autophagy by miR-30d impacts sensitivity of anaplastic thyroid carcinoma to cisplatin. Biochem Pharmacol. 2014;87:562–70. doi: 10.1016/j.bcp.2013.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Chatterjee A, Chattopadhyay D, Chakrabarti G, Mari B. miR-17-5p downregulation contributes to paclitaxel resistance of lung cancer cells through altering beclin1 expression. PLoS One. 2014;9:e95716. doi: 10.1371/journal.pone.0095716.s007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Menghini R, Casagrande V, Marino A, Marchetti V, Cardellini M, Stoehr R, Rizza S, Martelli E, Greco S, Mauriello A, Ippoliti A, Martelli F, Lauro R, Federici M. MiR-216a: a link between endothelial dysfunction and autophagy. Cell Death Dis. 2014;5:e1029. doi: 10.1038/cddis.2013.556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Korkmaz G, Tekirdag KA, Ozturk DG, Kosar A, Sezerman OU, Gozuacik D, Pfeffer S. MIR376A is a regulator of starvationinduced autophagy. PLoS One. 2013;8:e82556. doi: 10.1371/journal.pone.0082556.s001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Korkmaz G, le Sage C, Tekirdag KA, Agami R, Gozuacik D. miR-376b controls starvation and mTOR inhibition-related autophagy by targeting ATG4C and BECN1. Autophagy. 2012;8:165–76. doi: 10.4161/auto.8.2.18351. [DOI] [PubMed] [Google Scholar]
- 41.Frankel LB, Lund AH. MicroRNA regulation of autophagy. Carcinogenesis. 2012;33:2018–25. doi: 10.1093/carcin/bgs266. [DOI] [PubMed] [Google Scholar]
- 42.Chatterjee A, Chattopadhyay D, Chakrabarti G. miR-16 targets Bcl-2 in paclitaxel-resistant lung cancer cells and overexpres-sion of miR-16 along with miR-17 causes unprecedented sensitivity by simultaneously modulating autophagy and apoptosis. Cell Signal. 2014;27:189–203. doi: 10.1016/j.cellsig.2014.11.023. [DOI] [PubMed] [Google Scholar]
- 43.Huang Y, Guerrero-Preston R, Ratovitski EA. Phospho-DeltaNp63alpha-dependent regulation of autophagic signaling through transcription and micro-RNA modulation. Cell Cycle. 2012;11:1247–59. doi: 10.4161/cc.11.6.19670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Chu D, Zhao Z, Li Y, Li J, Zheng J, Wang W, Zhao Q, Ji G, Navarro A. Increased microRNA-630 expression in gastric cancer is associated with poor overall survival. PLoS One. 2014;9:e90526. doi: 10.1371/journal.pone.0090526.t002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Huan LC, Wu JC, Chiou BH, Chen CH, Ma N, Chang CY, Tsen YK, Chen SC. MicroRNA regulation of DNA repair gene expression in 4-aminobiphenyl-treated HepG2 cells. Toxicology. 2014;322:69–77. doi: 10.1016/j.tox.2014.05.003. [DOI] [PubMed] [Google Scholar]
- 46.Shi G, Shi J, Liu K, Liu N, Wang Y, Fu Z, Ding J, Jia L, Yuan W. Increased miR-195 aggravates neuropathic pain by inhibiting autophagy following peripheral nerve injury. Glia. 2013;61:504–12. doi: 10.1002/glia.22451. [DOI] [PubMed] [Google Scholar]
- 47.Frankel LB, Wen J, Lees M, Høyer-Hansen M, Farkas T, Krogh A, Jäättelä M, Lund AH. MicroRNA-101 is a potent inhibitor of autophagy. Embo J. 2011;30:4628–41. doi: 10.1038/emboj.2011.331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Chang Z, Huo L, Li K, Wu Y, Hu Z. Blocked autophagy by miR-101 enhances osteosarcoma cell chemosensitivity in vitro. Scientific World J. 2014;2014:794756. doi: 10.1155/2014/794756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Xie Z, Klionsky DJ. Autophagosome formation: core machinery and adaptations. Nat Cell Biol. 2007;9:1102–9. doi: 10.1038/ncb1007-1102. [DOI] [PubMed] [Google Scholar]
- 50.Yang X, Zhong X, Tanyi JL, Shen J, Xu C, Gao P, Zheng TM, DeMichele A, Zhang L. mir-30d Regulates multiple genes in the autophagy pathway and impairs autophagy process in human cancer cells. Biochem Biophys Res Commun. 2013;431:617–22. doi: 10.1016/j.bbrc.2012.12.083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Kovaleva V, Mora R, Park YJ, Plass C, Chiramel AI, Bartenschlager R, Dohner H, Stilgenbauer S, Pscherer A, Lichter P, Seiffert M. miRNA-130a targets ATG2B and DICER1 to inhibit autophagy and trigger killing of chronic lymphocytic leukemia cells. Cancer Res. 2012;72:1763–72. doi: 10.1158/0008-5472.CAN-11-3671. [DOI] [PubMed] [Google Scholar]
- 52.Yang J, Chen D, He Y, Melendez A, Feng Z, Hong Q, Bai X, Li Q, Cai G, Wang J, Chen X. MiR-34 modulates Caenorhabditis elegans lifespan via repressing the autophagy gene atg9. Age. 2011;35:11–22. doi: 10.1007/s11357-011-9324-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Borralho PM, Kren BT, Castro RE, da Silva IB, Steer CJ, Rodrigues CM. MicroRNA-143 reduces viability and increases sensitivity to 5-fluorouracil in HCT116 human colorectal cancer cells. Febs J. 2009;276:6689–700. doi: 10.1111/j.1742-4658.2009.07383.x. [DOI] [PubMed] [Google Scholar]
- 54.Chang Y, Yan W, He X, Zhang L, Li C, Huang H, Nace G, Geller DA, Lin J, Tsung A. miR-375 inhibits autophagy and reduces viability of hepatocellular carcinoma cells under hypoxic conditions. Gastroenterology. 2012;143:177–87.e8. doi: 10.1053/j.gastro.2012.04.009. [DOI] [PubMed] [Google Scholar]
- 55.Comincini S, Allavena G, Palumbo S, Morini M, Durando F, Angeletti F, Pirtoli L, Miracco C. MicroRNA-17 regulates the expression of ATG7 and modulates the autophagy process, improving the sensitivity to temozolomide and low-dose ionizing radiation treatments in human glioblastoma cells. Cancer Biol Ther. 2013;14:574–86. doi: 10.4161/cbt.24597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Xiao J, Zhu X, He B, Zhang Y, Kang B, Wang Z, Ni X. MiR-204 regulates cardiomyocyte autophagy induced by ischemia-reperfusion through LC3-II. J Biomed Sci. 2011;18:35. doi: 10.1016/j.ijcard.2011.01.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Mikhaylova O, Stratton Y, Hall D, Kellner E, Ehmer B, Drew AF, Gallo CA, Plas DR, Biesiada J, Meller J, Czyzyk-Krzeska MF. VHL-regulated MiR-204 suppresses tumor growth through inhibition of LC3B-mediated autophagy in renal clear cell carcinoma. Cancer Cell. 2012;21:532–46. doi: 10.1016/j.ccr.2012.02.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Meenhuis A, van Veelen PA, de Looper H, van Boxtel N, van den Berge IJ, Sun SM, Taskesen E, Stern P, de Ru AH, van Adrichem AJ, Demmers J, Jongen-Lavrencic M, Lowenberg B, Touw IP, Sharp PA, Erkeland SJ. MiR-17/20/93/106 promote hematopoietic cell expansion by targeting sequestosome 1-regulated pathways in mice. Blood. 2011;118:916–25. doi: 10.1182/blood-2011-02-336487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Serva A, Knapp B, Tsai YT, Claas C, Lisauskas T, Matula P, Harder N, Kaderali L, Rohr K, Erfle H, Eils R, Braga V, Starkuviene V, Ghoshal K. miR-17-5p regulates endocytic trafficking through targeting TBC1D2/Armus. PLoS One. 2012;7:e52555. doi: 10.1371/journal.pone.0052555.s006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Jegga AG, Schneider L, Ouyang X, Zhang J. Systems biology of the autophagy-lysosomal pathway. Autophagy. 2011;7:477–89. doi: 10.4161/auto.7.5.14811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Gibbings D, Mostowy S, Voinnet O. Autophagy selectively regulates miRNA homeostasis. Autophagy. 2013;9:781–3. doi: 10.4161/auto.23694. [DOI] [PMC free article] [PubMed] [Google Scholar]