

ABSTRACT
The arthropod-borne West Nile virus (WNV) emerged in New York State in 1999 and quickly spread throughout the United States. Transmission is maintained in an enzootic cycle in which infected mosquitoes transmit the virus to susceptible hosts during probing and feeding. Arthropod-derived components within the viral inoculum are increasingly acknowledged to play a role in infection of vertebrate hosts. We previously showed that Culex tarsalis mosquito saliva and salivary gland extract (SGE) enhance the in vivo replication of WNV. Here, we characterized the effective dose, timing, and proximity of saliva and SGE administration necessary for enhancement of WNV viremia using a mouse model. Mosquito saliva and SGE enhanced viremia in a dose-dependent manner, and a single mosquito bite or as little as 0.01 μg of SGE was effective at enhancing viremia, suggesting a potent active salivary factor. Viremia was enhanced when SGE was injected in the same location as virus inoculation from 24 h before virus inoculation through 12 h after virus inoculation. These results were confirmed with mosquito saliva deposited by uninfected mosquitoes. When salivary treatment and virus inoculation were spatially separated, viremia was not enhanced. In summary, the effects of mosquito saliva and SGE were potent, long lasting, and localized, and these studies have implications for virus transmission in nature, where vertebrate hosts are fed upon by both infected and uninfected mosquitoes over time. Furthermore, our model provides a robust system to identify the salivary factor(s) responsible for enhancement of WNV replication.
IMPORTANCE Mosquito-borne viruses are a significant class of agents causing emerging infectious diseases. WNV has caused over 18,000 cases of neuroinvasive disease in the United States since its emergence. We have shown that Culex tarsalis mosquito saliva and SGE enhance the replication of WNV. We now demonstrate that saliva and SGE have potent, long-lasting, and localized effects. Our model provides a robust system to identify the salivary factor(s) and characterize the mechanism responsible for enhancement of WNV replication. These studies could lead to the identification of novel prophylactic or treatment options useful in limiting the spread of WNV, other mosquito-borne viruses, and the diseases that they cause.
INTRODUCTION
West Nile virus (WNV) emerged in New York State in 1999 (1, 2) and spread throughout the continental United States within 5 years (3). While infection in humans is primarily asymptomatic, it can develop into a mild febrile disease or a more severe neuroinvasive disease. Over 18,000 cases of neuroinvasive disease were documented from 1999 to 2014 (Centers for Disease Control and Prevention). About 10% of patients who develop neuroinvasive disease succumb to infection; many others cope with persistent sequelae, such as fatigue, muscle weakness, and memory problems, for months to years after resolution of the primary infection (4).
WNV is maintained in an enzootic cycle between its mosquito vector and birds, its amplifying host. WNV can infect a variety of mosquito species in the laboratory, but Culex mosquitoes are the primary vectors in the field (5). In the United States, Culex pipiens, C. quinquefasciatus, and C. tarsalis are the most important enzootic vectors (5, 6). Culex mosquitoes can feed on birds and mammals and, thus, can serve as bridge vectors between the enzootic cycle and incidental hosts, such as humans (5, 6).
Mosquito saliva contains many pharmacologically active substances that affect hemostasis, inflammation, and the immune response of the vertebrate host (7), and the contribution of mosquito saliva to virus transmission and infection has been recognized for a variety of arthropod-borne viruses. Cache Valley virus infects adult mice when the virus is inoculated after uninfected Aedes or Culex mosquitoes spot feed on the mice but not when the virus is inoculated by needle alone (8). Similarly, vesicular stomatitis virus infects adult mice more efficiently when the mice are fed upon by infected Aedes mosquitoes than when they are inoculated by needle (9). Deer and chipmunks infected with La Crosse virus by infected Aedes mosquitoes have higher levels of viremia than animals inoculated with virus by needle (10). Mice inoculated with Rift Valley Fever virus and salivary gland extract (SGE) or with Rift Valley Fever virus by infected Aedes aegypti mosquitoes have increased levels of virus in the blood, liver, and brain and enhanced mortality (11). Mice inoculated with dengue virus (DENV) in areas of A. aegypti probing (12, 13) or by DENV-infected A. aegypti mosquitoes (13) have higher levels of viremia than mice inoculated by needle. Our group and others have demonstrated that WNV replication is enhanced by saliva from C. pipiens and C. tarsalis mosquitoes (14, 15) and A. aegypti mosquitoes (16). Virus levels in the blood are higher, virus spread to distant tissues such as the lymph nodes and spleen is accelerated, and neuroinvasion is increased when mice are infected with WNV by WNV-infected C. tarsalis mosquitoes than when mice are infected with the virus by needle inoculation (15). A similar enhancement of infection is observed when uninfected mosquitoes feed at the injection site immediately prior to WNV infection or when SGE is injected with the virus (15, 16). Our current goal is to further define three parameters of enhancement of WNV replication by mosquito saliva or SGE: dose, timing, and proximity.
MATERIALS AND METHODS
Cells and virus.
Vero African green monkey kidney cells (CCL-81; ATCC, Manassas, VA) were grown in minimal essential medium (MEM) with 10% fetal bovine serum (FBS) at 37°C in 5% CO2. A. albopictus mosquito C6/36 cells (CRL-1660; ATCC) were grown in MEM with 10% FBS at 28°C in 5% CO2. WNV stocks were derived from an infectious clone (17) using C6/36 cells as previously described (15). Briefly, the DNA plasmid was digested with XbaI (New England BioLabs, Ipswich, MA), and RNA was generated by in vitro transcription (mMessage mMachine; Invitrogen, Grand Island, NY). C6/36 cells were electroporated with 10 μg RNA, and virus was harvested 3 to 4 days postelectroporation upon observation of a cytopathic effect. Viral supernatants were clarified and stored at −80°C. The viral titers of the stocks were determined by plaque assay on Vero cells as described previously (18).
Animals.
Experiments conducted at the Wadsworth Center used a C. tarsalis mosquito colony established in 2003 from mosquitoes collected in the Coachella Valley, CA (15). The University of Wisconsin—Madison C. tarsalis colony was established in 2014 from mosquitoes collected in 2002 from the Kern National Wildlife Refuge in California (both kind gifts of William Reisen, University of California, Davis). Mosquitoes were maintained on an 18-h light, 6-h dark cycle; food (10% sucrose) and water were provided ad libitum. Female C57BL/6 mice were obtained from The Jackson Laboratory (Bar Harbor, ME) and allowed to acclimate for at least 1 week in a biosafety level 3 facility. All mice were 6 to 7 weeks old at the start of the studies. Food and water were provided ad libitum. All animal work was approved by the appropriate institutional animal care and use committees at the University of Wisconsin—Madison and at the Wadsworth Center and conducted in accordance with AAALAC standards.
SGE preparation.
Salivary glands were dissected from female C. tarsalis mosquitoes at 7 to 10 days postemergence and added to low-endotoxin phosphate-buffered saline (PBS) at a concentration of 200 glands/ml. The solution was frozen and thawed three times and sonicated on ice with three 20-s bursts at 100 mV, with each burst being separated by a 1-min incubation on ice. The solution was centrifuged at 5,000 × g for 10 min at 4°C. The supernatant was removed, and the protein concentration was determined by the use of bicinchoninic acid (Thermo Scientific, Waltham, MA). Extracts typically contained 150 to 200 μg/ml protein, which corresponds to approximately 0.75 to 1 μg protein per salivary gland or 1.5 to 2 μg protein per salivary gland pair.
Mouse studies. (i) Mosquito spot feeding.
Mice were fed upon by uninfected C. tarsalis mosquitoes (spot feeding) at 7 to 10 days postemergence as previously described (15). Briefly, the mosquitoes were divided into groups of five mosquitoes each and starved for approximately 40 h. The mice were anesthetized with isoflurane. The toes and heel of the left rear foot were covered with tape, exposing only the distal footpad. One group of five starved mosquitoes was allowed to probe and feed on the exposed footpad for 10 min or until they were fully engorged. The number of mosquitoes probing and feeding on the footpad over this period was recorded (one to five mosquitoes probed with or without feeding). The mosquitoes were examined afterward for the presence of blood in the abdomen, which was indicative of a blood meal. Control mice were anesthetized in parallel but were not exposed to mosquitoes. The mice were inoculated subcutaneously (s.c.) at the feeding site with 105 PFU WNV, the median dose inoculated by mosquitoes (19). Mice were immediately subjected to spot feeding by mosquitoes prior to inoculation (see Fig. 1A and B) or at various times relative to the time of infection (16 h prior to, immediately before, or 3 h after), as indicated below (see Fig. 2C).
FIG 1.
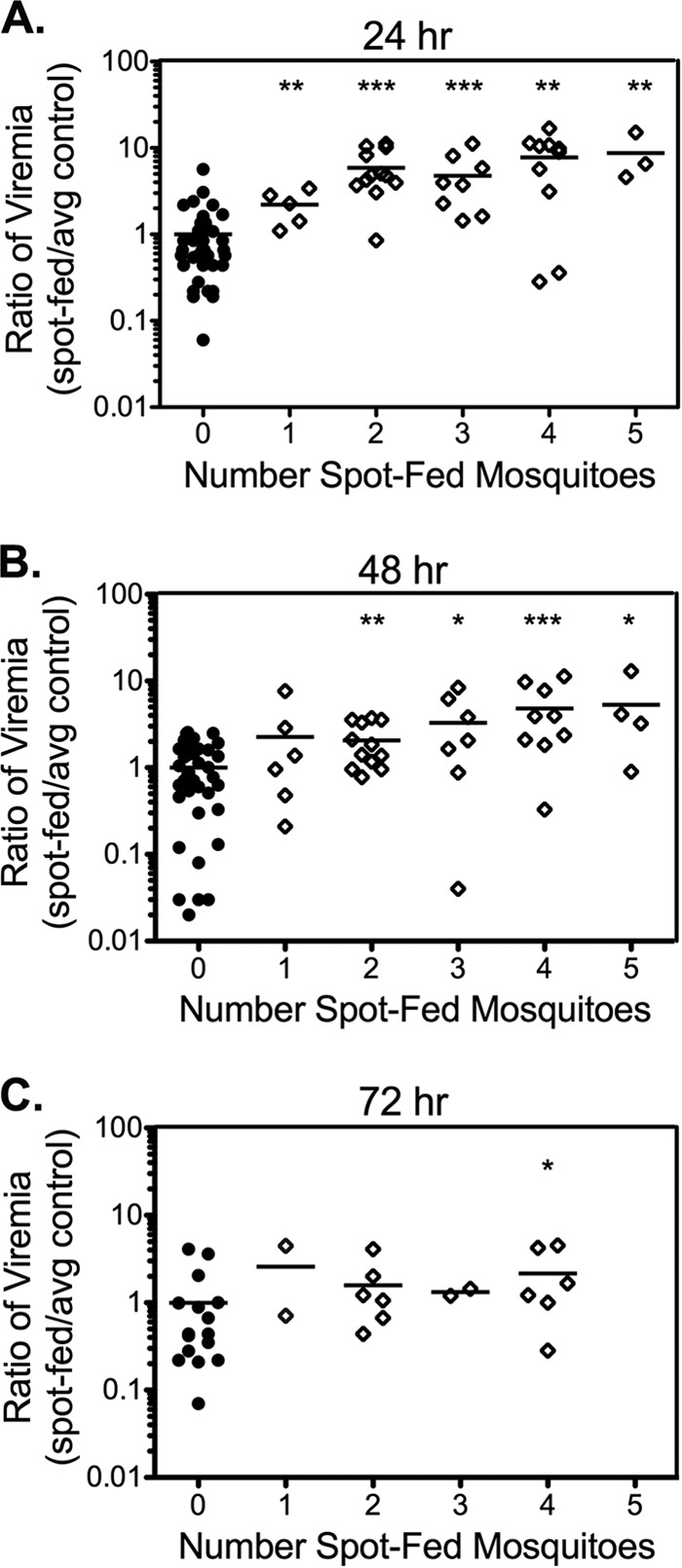
Mosquito spot feeding enhances WNV viremia in a dose-dependent manner. Up to five uninfected C. tarsalis mosquitoes were allowed to probe on the left rear footpad of mice (open diamonds). Control mice were not exposed to mosquitoes (solid circles). Mice were immediately inoculated s.c. with WNV at the site of feeding. Viremia was measured at 24 (A), 48 (B), or 72 (C) hpi. Note that no mice were fed upon by five mosquitoes at the 72-hpi time point (C). The data shown were combined from multiple experiments. Each symbol represents a single mouse (n = 2 to 38 mice), and bars indicate the means. Statistical significance between saliva-exposed and control mice was determined by the Mann-Whitney U test and is indicated as follows: *, P < 0.05; **, P < 0.01; ***, P < 0.001.
FIG 2.
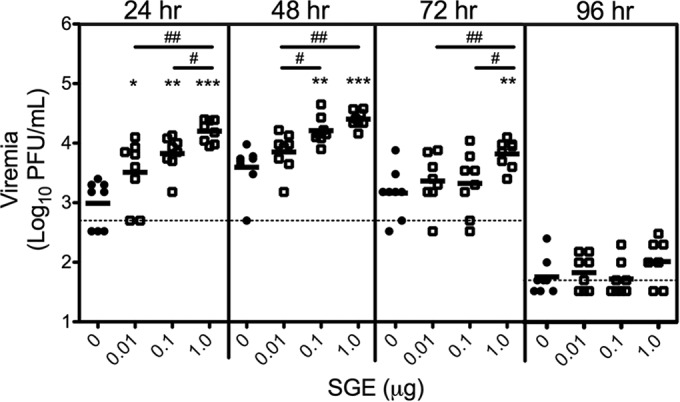
SGE enhances WNV viremia in a dose-dependent manner. Mice were inoculated s.c. in the left rear footpad with WNV mixed with diluent (0 μg SGE; solid circles) or the indicated dose of SGE (open squares). Data are from a representative study. Each symbol represents a single mouse (n = 8 mice), and bars indicate the means. Statistical significance was determined by the Mann-Whitney U test. Statistically significant differences between SGE-treated and control mice are indicated as follows: *, P < 0.05; **, P < 0.01; ***, P < 0.001. Statistically significant differences between SGE doses are indicated as follows: #, P < 0.05; ##, P < 0.01. Dashed line, limit of detection of 500 PFU/ml at 24 to 72 hpi or 50 PFU/ml at 96 hpi.
(ii) SGE studies.
For dose dependence studies, WNV (105 PFU) was mixed with diluent (low-endotoxin PBS with 1% FBS) or 0.01 μg, 0.1 μg, or 1 μg SGE in a final volume of 10 μl and injected s.c. into the left rear footpads of the mice.
For SGE timing studies, mice were injected with 105 PFU WNV in 5 μl diluent s.c. in the left rear footpad at time zero. At various times relative to the time of virus inoculation (time zero), the mice were injected s.c. in the same area with 0.5 μg SGE in 5 μl diluent. Control mice received diluent and virus in the left rear footpad at time zero.
For SGE localization studies, SGE (0.5 μg in 5 μl) was injected s.c. into the left or right rear footpad at 16 h prior to inoculation with virus (see Fig. 3A). Alternatively, 0.5 μg SGE in 5 μl was injected s.c. into the left rear thigh, calf, or footpad immediately before virus inoculation (Fig. 3B). Control mice received diluent in the left rear footpad at the time corresponding to the time of SGE administration. Mice were inoculated with 105 PFU WNV in 5 μl s.c. in the left rear footpad at time zero.
FIG 3.

SGE or mosquito spot feeding enhances WNV viremia over a broad time frame. (A to C) Mice were inoculated s.c. with 0.5 μg SGE in the left rear footpad at the indicated times before (negative numbers) or after (positive numbers) inoculation with WNV (time zero). Control mice (identified by C on the x axis) received diluent and WNV at time zero. Viremia was measured at 24 (A), 48 (B), or 72 (C) hpi. (D) One to five uninfected C. tarsalis mosquitoes (average, 1.95) probed on the left rear footpad of mice at the indicated times relative to the time of inoculation with WNV (time zero). Control mice were not exposed to mosquitoes. (A to D) The data shown are combined from multiple experiments. Each symbol represents a single mouse (in panels A to C, n = 4 to 20 mice; in panel D, n = 8 to 16 mice), and bars indicate the means. Statistically significant differences between experimental and control mice were determined by the Mann-Whitney U test and are indicated as follows: *, P < 0.05; **, P < 0.01; ***, P < 0.001.
Blood collection and processing.
Mice were serially bled from the tail or maxillary vein at 24, 48, and/or 72 h postinfection (hpi), and blood was collected by terminal exsanguination at 72 or 96 hpi. Blood was collected in Microtainer serum separator blood collection tubes (BD Biosciences, San Jose, CA) and centrifuged at 5,000 × g for 5 min at 4°C. Serum was removed and stored at −80°C. Viral titers in sera were determined by plaque assay on Vero cells. Viral titers below the limit of detection were assigned the value that would be obtained if one plaque were detected if an additional replicate was included (e.g., if the limit of detection was 500 PFU/ml, then a titer of 333 PFU/ml was assigned).
Data normalization and statistical analysis.
All data were log transformed. Data derived from a single experiment (see Fig. 2 and 4B) are graphed as the log10 level of viremia (number of PFU per milliliter). Raw data from multiple experiments were not combined due to variation between experiments. Rather, data were standardized before they were combined. To do this, the average value for the control group was determined at each time point. Each control and experimental value was divided by the average control value for the corresponding time point. The resulting value is defined as a ratio of viremia (experimental value/average control value). The ratio of viremia standardizes the results of an experiment by controlling for the natural variation that exists in animal studies, allowing data from multiple experiments to be directly compared. Statistical analysis of these data was performed with normalized data. Graphs containing data combined from multiple experiments are indicated by a y axis labeled ratio of viremia (see Fig. 1, 3, and 4A). All data are presented as the mean. Statistical significance was determined by the Mann-Whitney U test and defined as a P value of <0.05. In the figures, significance between a control group and an experimental group is indicated with asterisks and significance between experimental groups is indicated with number symbols.
FIG 4.
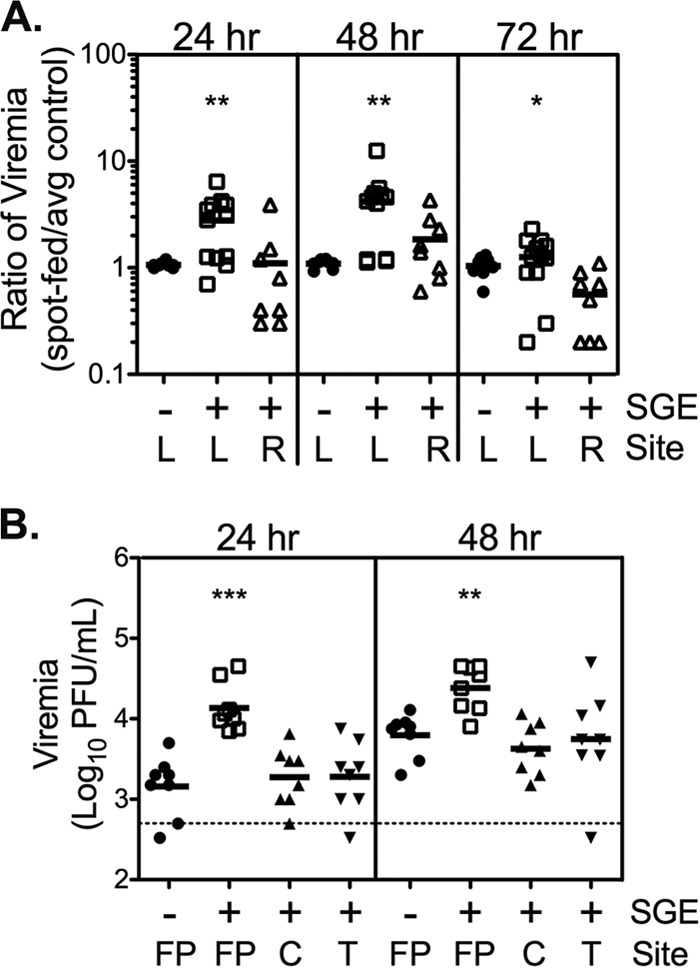
The SGE-mediated enhancement of WNV is highly localized. (A) Mice were inoculated s.c. with diluent (control) or 0.5 μg SGE in the left (L) or right (R) rear footpad. Sixteen hours later, the mice were inoculated s.c. with WNV in the left rear footpad. The data shown are combined from two experiments (n = 8 to 12 mice). (B) Mice were inoculated s.c. with 0.5 μg SGE into the left rear thigh (T), calf (C), or footpad (FP). Control mice received diluent in the left rear footpad. Mice were immediately inoculated s.c. with WNV in the left rear footpad. Data are from a single study (n = 8 mice). (A and B) Each symbol represents a single mouse, and bars indicate the means. Statistically significant differences between experimental and control mice were determined by the Mann-Whitney U test and are indicated as follows: *, P < 0.05; **, P < 0.01; ***, P < 0.001. The dashed line in panel B indicates the limit of detection of 500 PFU/ml.
RESULTS
Dose dependence of saliva enhancement phenotype.
Previous studies by others allowed between 3 and 30 mosquitoes to probe and feed in a single spot to demonstrate enhancement of arboviral infection (8, 12, 13, 16). We used two methods to examine if enhancement of viral replication by mosquito saliva exhibited a dose-response. We first fed different numbers of uninfected C. tarsalis mosquitoes on the left rear footpad of mice (spot feeding) prior to needle inoculation of WNV in the same location. One to five uninfected mosquitoes probed with or without feeding on the left rear footpad. Control mice were anesthetized without exposure to mosquitoes. All mice were immediately injected in the left rear footpad with WNV. Spot feeding by even a single mosquito enhanced the level of viremia at 24 hpi (Fig. 1A). Regression analysis demonstrated a modest correlation (R2 = 0.78) between the number of mosquitoes that spot fed and the virus levels at 24 hpi. Probing by more mosquitoes extended the enhancement phenotype. The levels of viremia at 48 hpi were significantly elevated in mice probed by two or more mosquitoes (Fig. 1B), and the level of viremia remained significantly higher at 72 hpi in mice probed by four mosquitoes (Fig. 1C).
We next treated mice with increasing amounts of SGE to quantitate the relationship between SGE dose and enhancement of WNV replication. A mosquito is estimated to deposit the contents of 0.3 to 0.4 salivary gland pairs each time that it probes or feeds (20). This corresponds to approximately 0.4 to 0.8 μg of protein extract in our system. We tested doses above and below this range by inoculating mice in the left rear footpad with WNV plus diluent or 0.01, 0.1, or 1 μg SGE (Fig. 2). The level of viremia at 24 hpi was significantly elevated in all groups that received SGE compared to that in the control group, demonstrating that as little as 0.01 μg SGE is sufficient for enhancement of viral replication. In addition, a dose-response was observed. At 24 and 48 hpi, the mean level of viremia increased with the dose of SGE. These differences were significant between 0.01 μg and 1 μg SGE at both time points, between 0.1 μg and 1 μg at 24 hpi, and between 0.01 μg and 0.1 μg at 48 hpi. At 72 hpi, virus levels were significantly higher with the 1-μg treatment than with the 0.1-μg and 0.01-μg treatments. Higher doses of SGE also extended the duration of enhancement. Mice receiving 1 or 0.1 μg but not mice receiving 0.01 μg had significantly higher levels of viremia at 48 hpi than the controls. Only mice that received 1 μg of SGE had significantly elevated levels of viremia at 72 hpi. No differences between any treatment groups were observed at 96 hpi when viremia was low or had cleared (19). These data mirror the dose-dependent effects of mosquito saliva observed in spot-feeding experiments and demonstrate that SGE exerts a potent dose-dependent effect on the extent and duration of enhancement of WNV replication.
Temporal separation of mosquito saliva and virus inoculation.
Our previous experiments involved the administration of saliva or SGE at the same time or immediately before WNV inoculation; however, mosquito spot feeding up to 4 h before virus inoculation enhanced Cache Valley virus replication (8). We therefore asked whether SGE could enhance WNV infection if it was administered before or after virus inoculation. SGE was delivered at a range of times relative to the time of WNV inoculation (defined as time zero) from 48 h before to 12 hpi, when WNV production was first detectable in mice (18). Control animals received virus diluent at the time of WNV inoculation. SGE and WNV were inoculated only in the left rear footpad, regardless of timing. SGE treatment enhanced viremia at 24 hpi when it was administered at any time from 24 h before to 12 h after WNV inoculation (Fig. 3A). Enhancement was more variable when SGE was administered at 48 hpi; groups that received SGE 16 h prior to, 3 or 12 h after, or at the time of inoculation demonstrated significant enhancement (Fig. 3B). A trend toward enhancement was often observed in the groups between the time points with statistically significant enhancement (those that received SGE 4 or 8 h prior to or 1 h postinfection). These groups contained fewer mice, suggesting that significance might be achieved with larger sample sizes. No enhancement was observed in any treatment group that received SGE at 72 hpi (Fig. 3C).
Spot-feeding studies with mosquitoes were conducted at select time points to confirm the physiological relevance of the observations made using SGE administration. One to five uninfected C. tarsalis mosquitoes (average, 1.95) were allowed to probe and feed on the left rear footpad 16 h prior to, immediately before (time zero), or 3 h after WNV inoculation. Viremia levels were significantly higher in all groups of mice on which mosquitoes were allowed to spot feed than in control mice at 24 hpi (Fig. 3D). Mice on which mosquitoes were allowed to spot feed immediately prior to inoculation also had significantly higher levels of viremia at 48 hpi. No differences in virus titers between mice on which mosquitoes were allowed to spot feed and mice on which mosquitoes were not allowed to spot feed were observed at 72 hpi. These data corroborate our results with SGE (Fig. 3A to C) and suggest that exposure to probing and feeding by uninfected mosquitoes can enhance the replication of virus transmitted by an infected mosquito in the same location over a broad time frame.
Spatial separation of mosquito saliva and virus inoculation.
We next asked whether SGE was effective if it was administered to a site physically distant from the WNV inoculation site. We previously demonstrated that SGE injected in the footpad opposite that inoculated with WNV immediately prior to WNV inoculation failed to enhance viremia (15). It is possible that SGE triggers a systemic effect favorable for WNV replication that requires time to develop. We tested this possibility by injecting SGE into the right rear footpad 16 h prior to WNV inoculation. Control mice received diluent or SGE in the left rear footpad 16 h prior to inoculation. All mice were inoculated with WNV in the left rear footpad at time zero. WNV titers in the blood at 24, 48, and 72 hpi were significantly enhanced when SGE and WNV were injected into the same (left rear) footpad 16 h apart (Fig. 4A). SGE injected into the right rear footpad did not enhance viremia at any time point. Thus, SGE did not cause a delayed systemic effect to enhance virus replication; its activity was locally restricted. To define the effective local range of SGE activity, SGE was injected into the left rear thigh, calf, or footpad followed immediately by WNV inoculation into the left rear footpad. Control mice received diluent and virus in separate injections in the left rear footpad. The level of viremia was significantly enhanced at 24 and 48 hpi when SGE and WNV were both injected into the footpad (Fig. 4B). SGE injected into the thigh or the calf had no effect on virus levels compared to those in the controls at any time point examined. Together, these data suggest that SGE activity is extremely localized.
DISCUSSION
In previous studies, we demonstrated enhancement of WNV infection in the presence of salivary proteins using a mouse model. The degree and duration of enhancement are similar when (i) WNV-infected mosquitoes feed on or probe mice, (ii) uninfected mosquitoes spot feed on mice prior to WNV inoculation, or (iii) mice are inoculated with WNV mixed with SGE (15). Since the enhancement phenotype occurs under all three experimental conditions, we conclude that viral enhancement is caused by mosquito salivary components and not the virus dose, virus source, location of virus inoculation, or mechanical damage due to mosquito probing. In the current study, we dissected three parameters that influence enhancement of virus replication by mosquito saliva: dose, time of administration, and proximity.
To our knowledge, we are the first to demonstrate that mosquito saliva acts in a dose-dependent manner to enhance virus levels in the blood. The enhancement phenotype persisted for 3 days when five mosquitoes spot fed on mice or when larger doses of SGE were inoculated. In addition, mosquito saliva is very potent, since probing or feeding by even a single mosquito enhanced the levels of viremia in mice in our spot-feeding studies. Previous studies by others allowed at least 3 and up to 30 mosquitoes to probe and feed in a single spot to demonstrate enhancement of arboviral infection (8, 12, 13, 16). A mosquito deposits the contents of 0.3 to 0.4 salivary gland pairs (20), corresponding to approximately 0.4 to 0.8 μg SGE, each time that it probes, but we demonstrated enhancement with as little as 0.01 μg SGE. Due to the potency of SGE and the likelihood that the active component represents only a fraction of the mixture, we hypothesize that the effective dose of the salivary component(s) is very low.
We were surprised to find that SGE enhanced the level of viremia over a broad time frame. A dose of SGE approximately equivalent to the amount of saliva deposited during probing or feeding by a single mosquito (0.5 μg) enhanced the level of viremia if it was administered from 24 h prior to through 12 h after virus inoculation. Previous studies showed that mosquito spot feeding enhances the level of replication of another arbovirus, Cache Valley virus, for 4 h but not 8 h prior to virus inoculation (8). The difference in the duration of enhancement activity could be due to the different mosquito species tested (Aedes triseriatus versus C. tarsalis), intrinsic differences between the viruses, different experimental conditions (the mouse strain used, the endpoint readout used, etc.), or a combination of these factors. We confirmed our results on timing with mosquito spot-feeding studies, in which the level of WNV viremia was enhanced when mosquito saliva was delivered 16 h before and 3 h after virus inoculation. To our knowledge, we are the first to show enhancement of virus replication when mosquito saliva is delivered after virus inoculation, and our results suggest that probing and feeding by uninfected mosquitoes can enhance WNV infection before or after virus transmission in nature. It will be interesting to see if trends for the time frame of enhancement emerge as more groups investigate this parameter.
Although the time frame for enhancement was broad, the location required for enhancement was localized. SGE did not enhance WNV replication unless it was administered in the same immediate location, whether SGE administration was separated by 16 h from the time of virus inoculation or SGE was administered at different locations on the same leg. Cache Valley virus also requires colocalization of virus and uninfected A. triseriatus mosquito bites (8). In contrast, DENV replication is enhanced when uninfected A. aegypti mosquitoes bite the host at a site distant from virus inoculation in a humanized mouse model (13). Further studies with other virus-mosquito-host systems are needed to investigate the requirement of colocalization of mosquito saliva and virus.
In the current studies, we did not use infected mosquitoes, which inoculate a wide range of WNV doses (19), in order to control for viral dose as an experimental parameter. Furthermore, we previously showed viral replication enhancement similar to that found in the present study when we use infected mosquitoes, spot feeding by uninfected mosquitoes, and inoculation of SGE (15). We used the last two methods, spot feeding by uninfected mosquitoes and inoculation of SGE, in our current investigations; however, each method has its limitations. First, mosquitoes deposit variable amounts of saliva and probe/feed for variable lengths of time (19), which introduces variability and prevents us from conducting actual saliva dose-response studies. In addition, it is difficult to conduct closely spaced timing and location studies with live mosquitoes. Thus, we limited studies with live mosquitoes to confirm the findings of our studies with SGE. Inoculation of SGE with virus allows precise control of the protein dose and the timing and location of inoculation. On the other hand, SGE is an extract of all proteins in the salivary gland and not just the secreted salivary proteins. We fully acknowledge this limitation; however, we chose not to use mosquito saliva, which can be collected artificially by placing the mosquito's proboscis into a capillary tube or by allowing mosquitoes to feed on a sucrose solution through a membrane. Such saliva preparations differ qualitatively and quantitatively from mosquito saliva that is inoculated into the host during a natural blood meal. The mixture of salivary proteins in saliva varies between blood and sucrose feeding (21). This is most likely due to different salivary gland gene expression and salivary secretion in the three lobes of the salivary gland. Genes associated with blood feeding are expressed primarily in the medial lobe (22, 23), which has unique innervation (24, 25). Proteins present in the medial lobe are found in saliva collected during blood feeding but are absent from saliva collected during sugar feeding. In addition, the proteins present in artificially collected saliva are substantially diluted (20) and must be precipitated prior to use (26). We published evidence of this dilution in previous studies, which showed that the amount of WNV in artificially collected mosquito saliva is 100-fold lower than the amount of virus inoculated by a mosquito into a live host (19). For these reasons, artificially collected mosquito saliva is infrequently used for in vivo studies, and SGE has been used as a substitute for mosquito saliva in numerous experimental systems (11, 16, 20, 27–35). We thus used SGE, which contains secreted salivary factors representative of those obtained by both blood and sugar feeding and which can be produced in sufficient quantities with minimal manipulations that could reduce or eliminate functional activity. Finally, we confirmed our results with SGE by conducting mosquito spot-feeding studies with one to five mosquitoes, in which the mosquito saliva composition and deposition mimic those associated with a natural blood meal.
Saliva from female mosquitoes is a complex mixture of molecules required for blood feeding and sugar feeding and includes mucins, proteases, vasodilators, anticoagulants, and immune modulators. The specific composition differs for each genus, with unique proteins being found in Culex, Aedes, and Anopheles spp., and the function of the majority of the proteins is unknown (reviewed in reference 7). For example, transcriptome analysis of female C. tarsalis salivary glands reveals 80 putatively secreted salivary proteins, but the functions for less than half of them are known (36). Thus, it is difficult to speculate on the identity of the WNV replication-enhancing component(s). Recently, Conway et al. (27) demonstrated that a serine protease in saliva from A. aegypti and C. tarsalis mosquitoes enhances WNV levels in vivo. The enhancement activity is abolished by heat treatment and also occurs in cell culture (27). Neither of these is true for our system (unpublished data), suggesting that a different active component enhances viremia in our system. It is possible that more than one salivary factor contributes to the enhancement of WNV replication. We plan to examine the replication enhancement ability of different species of mosquitoes, identify common salivary components, and test candidate factors for their ability to enhance WNV replication in vivo. This methodical approach will provide insight into currently uncharacterized factors and inform the mechanism of enhancement of WNV replication.
The mechanism of enhancement is unclear. Viral enhancement occurs even when virus and saliva or SGE are administered separately, suggesting an indirect effect on the virus. The saliva perhaps stimulates a persistent local host response conducive to enhanced viral replication and spread, as saliva is known to alter immune responses (37). Mosquito saliva induces TH2 polarization in mouse skin, leading to higher levels of interleukin-4 (IL-4) (38) or IL-10 (30), and in cultured splenocytes, leading to higher levels of IL-4 and IL-10 and decreased gamma interferon (IFN-γ) levels as long as 10 days after exposure (39). Saliva may also recruit host cells to the feeding site. Anopheles saliva increases the number of neutrophils at the feeding site and the number of dendritic cells in draining lymph nodes (40); both of these cell types support WNV replication (41, 42). Thus, the observed increase in virus levels in our system may be due to an increased number of infected cells rather than an increase in the level of viral replication per cell. Alternatively, mosquito saliva may prevent the early detection and clearance of virus. A. aegypti saliva has been shown to reduce the amount of IFN-β and inducible nitric oxide synthase in macrophages as well as to decrease the influx of T cells (30). The dampening of the normal immune response may provide the virus time to gain an early foothold for replication and spread.
The immune response to mosquito saliva and salivary proteins clearly plays a complex role in infection. Sensitization of mice to A. aegypti saliva enhances WNV pathogenicity (43). In contrast, sensitization of mice to C. tarsalis saliva has no effect on virus levels or morbidity in our previous studies (15). Immunization of mice with C. tarsalis SGE reduces the rates of morbidity and mortality after WNV infection (28), but vaccination against D7, a prominent mosquito salivary protein, increases the rate of mortality during WNV infection (44). Further work is needed to map the interplay between the host immune response, mosquito saliva, and virus replication.
Another possible mechanism for enhancement is the lingering of the active salivary component(s) at the site of infection. Salivary proteins can linger at a mosquito feeding site for up to 18 hpi (45, 46), and we have found that SGE incubated overnight at 37°C retains its viral replication enhancement activity (data not shown). Perhaps any remaining components prime host cells for infection. SGE could stabilize infected cells, preventing virus-induced cell death and allowing viral replication to continue longer than it otherwise would. Alternatively, the residual salivary components could act as a bridge for infection, transforming previously nonpermissive cells into permissive cells.
Arthropod saliva plays an intimate role in the transmission and replication of vector-borne viruses. Future studies will be aimed at identifying the active salivary component(s) and determining the underlying mechanism responsible for enhancement of WNV replication. C. tarsalis saliva increases WNV levels in the blood, enhances spread to distant tissues, and accelerates central nervous system invasion, but it has no effect on morbidity in mice (15). Virus levels in the blood are, however, a predictor of morbidity and mortality from infections caused by other mosquito-borne viruses, such as dengue virus (47). Furthermore, higher levels of viremia in amplifying hosts lead to higher levels of transmission in nature (48). Thus, studies in the WNV model system will provide insight into the interactions between vector, host, and pathogen. Characterization of these interactions could lead to the identification of novel prophylactic or treatment options useful in limiting not only WNV pathogenesis but also the transmission of other mosquito-borne viruses and the diseases that they cause as well.
ACKNOWLEDGMENTS
We gratefully acknowledge the technical support of Fernando Moreira and Brendan Boylan at the University of Wisconsin—Madison and Karen Louie, Rebecca Albright, and Kim Appler at the Wadsworth Center. We also thank William Reisen for providing C. tarsalis mosquitoes to establish our mosquito colony and Lyric Bartholomay for use of the insectary at the University of Wisconsin—Madison.
Funding Statement
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
REFERENCES
- 1.Briese T, Jordan I, Rambaut A, Chi HC, MacKenzie JS, Roy A, Scherret JH, Lipkin WI. 1999. Genetic analysis of West Nile New York 1999 encephalitis virus (research letters). Lancet 354:1971–1972. doi: 10.1016/S0140-6736(99)05384-2. [DOI] [PubMed] [Google Scholar]
- 2.Lanciotti RS, Roehrig JT, Deubel V, Smith J, Parker M, Steele K, Crise B, Volpe KE, Crabtree MB, Scherret JH, Hall RA, MacKenzie JS, Cropp CB, Panigrahy B, Ostlund E, Schmitt B, Malkinson M, Banet C, Weissman J, Komar N, Savage HM, Stone W, McNamara T, Gubler DJ. 1999. Origin of the West Nile virus responsible for an outbreak of encephalitis in the northeastern United States. Science 286:2333–2337. doi: 10.1126/science.286.5448.2333. [DOI] [PubMed] [Google Scholar]
- 3.Petersen LR, Hayes EB. 2008. West Nile virus in the Americas. Med Clin North Am 92:1307–1322, ix. doi: 10.1016/j.mcna.2008.07.004. [DOI] [PubMed] [Google Scholar]
- 4.Sejvar JJ. 2007. The long-term outcomes of human West Nile virus infection. Clin Infect Dis 44:1617–1624. doi: 10.1086/518281. [DOI] [PubMed] [Google Scholar]
- 5.Hayes EB, Komar N, Nasci RS, Montgomery SP, O'Leary DR, Campbell GL. 2005. Epidemiology and transmission dynamics of West Nile virus disease. Emerg Infect Dis 11:1167–1173. doi: 10.3201/eid1108.050289a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Turell MJ, Dohm DJ, Sardelis MR, Oguinn ML, Andreadis TG, Blow JA. 2005. An update on the potential of North American mosquitoes (Diptera: Culicidae) to transmit West Nile virus. J Med Entomol 42:57–62. doi: 10.1093/jmedent/42.1.57. [DOI] [PubMed] [Google Scholar]
- 7.Ribeiro JM, Mans BJ, Arca B. 2010. An insight into the sialome of blood-feeding Nematocera. Insect Biochem Mol Biol 40:767–784. doi: 10.1016/j.ibmb.2010.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Edwards JF, Higgs S, Beaty BJ. 1998. Mosquito feeding-induced enhancement of Cache Valley Virus (Bunyaviridae) infection in mice. J Med Entomol 35:261–265. doi: 10.1093/jmedent/35.3.261. [DOI] [PubMed] [Google Scholar]
- 9.Limesand KH, Higgs S, Pearson LD, Beaty BJ. 2000. Potentiation of vesicular stomatitis New Jersey virus infection in mice by mosquito saliva. Parasite Immunol 22:461–467. doi: 10.1046/j.1365-3024.2000.00326.x. [DOI] [PubMed] [Google Scholar]
- 10.Osorio JE, Godsey MS, DeFoliart GR, Yuill TM. 1996. La Crosse viremias in white-tailed deer and chipmunks exposed by injection or mosquito bite. Am J Trop Med Hyg 54:338–342. [DOI] [PubMed] [Google Scholar]
- 11.Le Coupanec A, Babin D, Fiette L, Jouvion G, Ave P, Misse D, Bouloy M, Choumet V. 2013. Aedes mosquito saliva modulates Rift Valley fever virus pathogenicity. PLoS Negl Trop Dis 7:e2237. doi: 10.1371/journal.pntd.0002237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.McCracken MK, Christofferson RC, Chisenhall DM, Mores CN. 2014. Analysis of early dengue virus infection in mice as modulated by Aedes aegypti probing. J Virol 88:1881–1889. doi: 10.1128/JVI.01218-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cox J, Mota J, Sukupolvi-Petty S, Diamond MS, Rico-Hesse R. 2012. Mosquito bite delivery of dengue virus enhances immunogenicity and pathogenesis in humanized mice. J Virol 86:7637–7649. doi: 10.1128/JVI.00534-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Styer LM, Bernard KA, Kramer LD. 2006. Enhanced early West Nile virus infection in young chickens infected by mosquito bite: effect of viral dose. Am J Trop Med Hyg 75:337–345. [PubMed] [Google Scholar]
- 15.Styer LM, Lim PY, Louie KL, Albright RG, Kramer LD, Bernard KA. 2011. Mosquito saliva causes enhancement of West Nile virus infection in mice. J Virol 85:1517–1527. doi: 10.1128/JVI.01112-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Schneider BS, Soong L, Girard YA, Campbell G, Mason P, Higgs S. 2006. Potentiation of West Nile encephalitis by mosquito feeding. Viral Immunol 19:74–82. doi: 10.1089/vim.2006.19.74. [DOI] [PubMed] [Google Scholar]
- 17.Shi PY, Tilgner M, Lo MK, Kent KA, Bernard KA. 2002. Infectious cDNA clone of the epidemic West Nile virus from New York City. J Virol 76:5847–5856. doi: 10.1128/JVI.76.12.5847-5856.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Brown AN, Kent KA, Bennett CJ, Bernard KA. 2007. Tissue tropism and neuroinvasion of West Nile virus do not differ for two mouse strains with different survival rates. Virology 368:422–430. doi: 10.1016/j.virol.2007.06.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Styer LM, Kent KA, Albright RG, Bennett CJ, Kramer LD, Bernard KA. 2007. Mosquitoes inoculate high doses of West Nile virus as they probe and feed on live hosts. PLoS Pathog 3:e132. doi: 10.1371/journal.ppat.0030132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wasserman HA, Singh S, Champagne DE. 2004. Saliva of the yellow fever mosquito, Aedes aegypti, modulates murine lymphocyte function. Parasite Immunol 26:295–306. doi: 10.1111/j.0141-9838.2004.00712.x. [DOI] [PubMed] [Google Scholar]
- 21.Marinotti O, James AA, Ribeiro JMC. 1990. Diet and salivation in female Aedes aegypti mosquitoes. J Insect Physiol 36:545–548. doi: 10.1016/0022-1910(90)90021-7. [DOI] [Google Scholar]
- 22.Juhn J, Naeem-Ullah U, Maciel Guedes BA, Majid A, Coleman J, Paolucci Pimenta PF, Akram W, James AA, Marinotti O. 2011. Spatial mapping of gene expression in the salivary glands of the dengue vector mosquito, Aedes aegypti. Parasit Vectors 4:1. doi: 10.1186/1756-3305-4-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Poehling HM. 1979. Distribution of specific proteins in the salivary-gland lobes of Culcidae and their relation to age and blood sucking. J Insect Physiol 25:3–8. doi: 10.1016/0022-1910(79)90029-5. [DOI] [Google Scholar]
- 24.Novak MG, Ribeiro JM, Hildebrand JG. 1995. 5-Hydroxytryptamine in the salivary glands of adult female Aedes aegypti and its role in regulation of salivation. J Exp Biol 198:167–174. [DOI] [PubMed] [Google Scholar]
- 25.Barrow PM, McIver SB, Wright KA. 1975. Salivary glands of female Culex pipiens—morphological changes associated with maturation and blood-feeding. Can Entomol 107:1153–1160. doi: 10.4039/Ent1071153-11. [DOI] [Google Scholar]
- 26.Machain-Williams C, Mammen MP Jr, Zeidner NS, Beaty BJ, Prenni JE, Nisalak A, Blair CD. 2012. Association of human immune response to Aedes aegypti salivary proteins with dengue disease severity. Parasite Immunol 34:15–22. doi: 10.1111/j.1365-3024.2011.01339.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Conway MJ, Watson AM, Colpitts TM, Dragovic SM, Li Z, Wang P, Feitosa F, Shepherd DT, Ryman KD, Klimstra WB, Anderson JF, Fikrig E. 2014. Mosquito saliva serine protease enhances dissemination of dengue virus into the mammalian host. J Virol 88:164–175. doi: 10.1128/JVI.02235-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Machain-Williams C, Reagan K, Wang T, Zeidner NS, Blair CD. 2013. Immunization with Culex tarsalis mosquito salivary gland extract modulates West Nile virus infection and disease in mice. Viral Immunol 26:84–92. doi: 10.1089/vim.2012.0051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Schneider BS, Soong L, Zeidner NS, Higgs S. 2004. Aedes aegypti salivary gland extracts modulate anti-viral and TH1/TH2 cytokine responses to Sindbis virus infection. Viral Immunol 17:565–573. doi: 10.1089/vim.2004.17.565. [DOI] [PubMed] [Google Scholar]
- 30.Schneider BS, Soong L, Coffey LL, Stevenson HL, McGee CE, Higgs S. 2010. Aedes aegypti saliva alters leukocyte recruitment and cytokine signaling by antigen-presenting cells during West Nile virus infection. PLoS One 5:e11704. doi: 10.1371/journal.pone.0011704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Limesand KH, Higgs S, Pearson LD, Beaty BJ. 2003. Effect of mosquito salivary gland treatment on vesicular stomatitis New Jersey virus replication and interferon alpha/beta expression in vitro. J Med Entomol 40:199–205. doi: 10.1603/0022-2585-40.2.199. [DOI] [PubMed] [Google Scholar]
- 32.Wanasen N, Nussenzveig RH, Champagne DE, Soong L, Higgs S. 2004. Differential modulation of murine host immune response by salivary gland extracts from the mosquitoes Aedes aegypti and Culex quinquefasciatus. Med Vet Entomol 18:191–199. doi: 10.1111/j.1365-2915.2004.00498.x. [DOI] [PubMed] [Google Scholar]
- 33.Surasombatpattana P, Patramool S, Luplertlop N, Yssel H, Misse D. 2012. Aedes aegypti saliva enhances dengue virus infection of human keratinocytes by suppressing innate immune responses. J Investig Dermatol 132:2103–2105. doi: 10.1038/jid.2012.76. [DOI] [PubMed] [Google Scholar]
- 34.Bissonnette EY, Rossignol PA, Befus AD. 1993. Extracts of mosquito salivary gland inhibit tumour necrosis factor alpha release from mast cells. Parasite Immunol 15:27–33. doi: 10.1111/j.1365-3024.1993.tb00569.x. [DOI] [PubMed] [Google Scholar]
- 35.Cross ML, Cupp EW, Enriquez FJ. 1994. Differential modulation of murine cellular immune responses by salivary gland extract of Aedes aegypti. Am J Trop Med Hyg 51:690–696. [DOI] [PubMed] [Google Scholar]
- 36.Calvo E, Sanchez-Vargas I, Favreau AJ, Barbian KD, Pham VM, Olson KE, Ribeiro JM. 2010. An insight into the sialotranscriptome of the West Nile mosquito vector, Culex tarsalis. BMC Genomics 11:51. doi: 10.1186/1471-2164-11-51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Schneider BS, Higgs S. 2008. The enhancement of arbovirus transmission and disease by mosquito saliva is associated with modulation of the host immune response. Trans R Soc Trop Med Hyg 102:400–408. doi: 10.1016/j.trstmh.2008.01.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Thangamani S, Higgs S, Ziegler S, Vanlandingham D, Tesh R, Wikel S. 2010. Host immune response to mosquito-transmitted chikungunya virus differs from that elicited by needle inoculated virus. PLoS One 5:e12137. doi: 10.1371/journal.pone.0012137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Zeidner NS, Higgs S, Happ CM, Beaty BJ, Miller BR. 1999. Mosquito feeding modulates Th1 and Th2 cytokines in flavivirus susceptible mice: an effect mimicked by injection of sialokinins, but not demonstrated in flavivirus resistant mice. Parasite Immunol 21:35–44. doi: 10.1046/j.1365-3024.1999.00199.x. [DOI] [PubMed] [Google Scholar]
- 40.Demeure CE, Brahimi K, Hacini F, Marchand F, Peronet R, Huerre M, St-Mezard P, Nicolas JF, Brey P, Delespesse G, Mecheri S. 2005. Anopheles mosquito bites activate cutaneous mast cells leading to a local inflammatory response and lymph node hyperplasia. J Immunol 174:3932–3940. doi: 10.4049/jimmunol.174.7.3932. [DOI] [PubMed] [Google Scholar]
- 41.Lim PY, Behr MJ, Chadwick CM, Shi PY, Bernard KA. 2011. Keratinocytes are cell targets of West Nile virus in vivo. J Virol 85:5197–5201. doi: 10.1128/JVI.02692-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Bai F, Kong KF, Dai J, Qian F, Zhang L, Brown CR, Fikrig E, Montgomery RR. 2010. A paradoxical role for neutrophils in the pathogenesis of West Nile virus. J Infect Dis 202:1804–1812. doi: 10.1086/657416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Schneider BS, McGee CE, Jordan JM, Stevenson HL, Soong L, Higgs S. 2007. Prior exposure to uninfected mosquitoes enhances mortality in naturally-transmitted West Nile virus infection. PLoS One 2:e1171. doi: 10.1371/journal.pone.0001171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Reagan KL, Machain-Williams C, Wang T, Blair CD. 2012. Immunization of mice with recombinant mosquito salivary protein D7 enhances mortality from subsequent West Nile virus infection via mosquito bite. PLoS Negl Trop Dis 6:e1935. doi: 10.1371/journal.pntd.0001935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Choumet V, Attout T, Chartier L, Khun H, Sautereau J, Robbe-Vincent A, Brey P, Huerre M, Bain O. 2012. Visualizing non infectious and infectious Anopheles gambiae blood feedings in naive and saliva-immunized mice. PLoS One 7:e50464. doi: 10.1371/journal.pone.0050464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Yamamoto DS, Yokomine T, Sumitani M, Yagi K, Matsuoka H, Yoshida S. 2013. Visualization and live imaging analysis of a mosquito saliva protein in host animal skin using a transgenic mosquito with a secreted luciferase reporter system. Insect Mol Biol 22:685–693. doi: 10.1111/imb.12055. [DOI] [PubMed] [Google Scholar]
- 47.Yacoub S, Wills B. 2014. Predicting outcome from dengue. BMC Med 12:147. doi: 10.1186/s12916-014-0147-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Komar N, Langevin S, Hinten S, Nemeth N, Edwards E, Hettler D, Davis B, Bowen R, Bunning M. 2003. Experimental infection of North American birds with the New York 1999 strain of West Nile virus. Emerg Infect Dis 9:311–322. doi: 10.3201/eid0903.020628. [DOI] [PMC free article] [PubMed] [Google Scholar]