

Abstract
Background
The synthetic biology technology which enhances the specificity and efficacy of treatment is a novel try in biomedical therapy during recent years. A high frequency of somatic mutations was shown in the human telomerase reverse transcriptase (hTERT) promoter in bladder cancer, indicating that a mutational hTERT promoter might be a tumor-specific element for bladder cancer therapy. In our study, we aimed to construct a synthetic combination module driven by a super artificial hTERT promoter and to investigate its influence on the malignant phenotypes of bladder cancer.
Methods
The dual luciferase assay system was used to verify the driven efficiency and tumor-specificity of the artificial hTERT promoter and to confirm the relationship between ETS-1 and the driven efficiency of the artificial hTERT promoter. CCK-8 assay and MTT assay were used to test the effects of the Bax-Anti Bcl2 combination module driven by the artificial hTERT promoter on cell proliferation. Simultaneously, the cell apoptosis was detected by the caspase 3ELISA assay and the flow cytometry analysis after transfection. The results of CCK-8 assay and MTT assay were analyzed by ANOVA. The independent samples t-test was used to analyze other data.
Results
We demonstrated that the artificial hTERT promoter had a higher driven efficiency which might be regulated by transcription factor ETS-1 in bladder cancer cells, compared with wild-type hTERT promoter. Meanwhile, the artificial hTERT promoter showed a strong tumor-specific effect. The cell proliferation inhibition and apoptosis induction were observed in artificial hTERT promoter- Bax-Anti Bcl2 combination module -transfected bladder cancer 5637 and T24 cells, but not in the module -transfected normal human fibroblasts.
Conclusion
This module offers us a useful synthetic biology platform to inhibit the malignant phenotypes of bladder cancer in a more specific and effective way.
Keywords: hTERT promoter, Bax, Bcl2, Synthetic biology, Therapy, Bladder cancer
Background
Bladder cancer is one of the most common types of urologic tumors around the world [1]. Despite methods for early diagnosis of bladder cancer have been emerged, effective treatments are still required [2]. Traditional therapies for bladder cancer are mainly surgery, radiation therapy and chemotherapy, which have dissatisfied side-effect respectively on account of non-specificity and inefficiency [3, 4].
For this reason, it should be obligatory to find a new method for enhancing specificity and availability of bladder cancer therapy.
Synthetic biology is a novel discipline that aims to construct fresh and foreseeable modules, circuits or life entities with the standardized bioparts base on engineering principles [5]. The evolution of synthetic biology provides a favorable platform for the cancer therapy [6].
Therefore, using the theory of synthetic biology to design specific elements is feasible for the cancer treatment. In our previous work [7], the frequency of somatic mutations in the human telomerase reverse transcriptase (hTERT) promoter was about 55.6 % in bladder cancer. The mutant hTERT promoter could activate the expression of hTERT by using the transcription factor v-ets avian erythroblastosis virus E26 oncogene homolog 1 (ETS-1). ETS-1was associated with the progression and angiogenesis of several malignancies [8, 9]. Furthermore, it was reported that ETS-1 expression was high in urothelial carcinomas of the urinary bladder, indicating that it might be a marker of aggressiveness [10]. These discoveries inspired us to design an artificial hTERT promoter as a tumor-specific element regulated by ETS-1.
In the apoptotic pathway, Bax and Bcl2 are two important regulator genes [11]. Bcl2 regulates cell apoptosis and Bax promotes cell apoptosis [12]. The ratio of Bcl2/Bax is closely related to the sensitivity of cells apoptosis. When Bcl2 is excessive, cells are protected. On the contrary, when Bax is in excess, cells are susceptible to apoptosis [13]. Moreover, some extracellular or cellular factors were reported to have induced cell apoptosis through reducing Bcl2 levels and elevating Bax levels [14–18]. It indicates that the combination of Bax protein and anti-Bcl2 molecule can be used to reverse the ratio of Bcl2/Bax in bladder cancer.
In this study, we constructed the Bax-Anti Bcl2 combination module driven by artificial hTERT promoter which could over express Bax and knockdown of Bcl2 and tested the ability of this module in selectively indentifying and killing bladder cancer cells. Our results demonstrated that the Bax-Anti Bcl2 combination module driven by artificial hTERT promoter selectively inhibits malignant phenotypes of bladder cancer cells.
Methods
Cell lines and cell culture
Human bladder cancer cell lines (T24, 5637, UMUC-3, RT4, J82, SW780) were purchased from the Institute of Cell Biology, Chinese Academy of Sciences (Shanghai, China). Normal human fiber cell (NHF) was kindly provided by Ting Chen, Peking University Shenzhen Hospital, China. T24, UMUC-3, RT4, J82, SW780 and NHF were maintained in DMEM media supplemented with 10 % fetal bovine serum and 1 % antibiotics (100U/ml penicillin and 100 μg/ml streptomycin sulfates). 5637 was maintained in RPMI-1640 media supplemented with 10 % fetal bovine serum and 1 % antibiotics (100U/ml penicillin and 100 μg/ml streptomycin sulfates). All cells were routinely grown at 37 °C in an atmosphere of 5 % CO2.
Design and construction of the artificial hTERT promoter-driven reporter module
Base on the wild-type hTERT promoter sequence, some mutations were introduced to construct the artificial hTERT promoter. The cytosines (C) which were on the upstream −124, −138, −139, and −146 of initiation codon (ATG) were mutated to thymines (T). Furthermore, the adenines (A) which were on the upstream −57 and −189 of ATG were severally mutated to cytosine (C) and guanine (G), and the cytosine (C) which was on the upstream −89 of ATG was mutated to guanine (G). The artificial hTERT promoter has been chemically synthesized. Then the artificial hTERT promoter fragment was cloned into siCHECKTM-2 luciferase vector (Promerga, Madison, WI, USA) digested with BgIII and Nhel to substitute the SV40 promoter. To construct a negative control, the wild-type hTERT promoter fragment was cloned into siCHECK™-2 luciferase vector (Promerga, Madison, WI, USA) digested with BgIII and Nhel to substitute the SV40 promoter as well. All the detailed sequence information has showed in Table 1.
Table 1.
Related sequences in the modules
| Name | Related sequences |
|---|---|
| Wild-type hTERT promoter | GGCCCCTCCC TCGGGTTACC CCACAGCCTA GGCCGATTCG ACCTCTCTCC GCTGGGGCCC TCGCTGGCGT CCCTGCACCC TGGGAGCGCG AGCGGCGCGC GGGCGGGGAA GCGCGGCCCA GACCCCCGGG TCCGCCCGGA GCAGCTGCGC TGTCGGGGCC AGGCCGGGCT CCCAGTGGAT TCGCGGGCAC AGACGCCCAG GACCGCGCTC CCCACGTGGC GGAGGGACTG GGGACCCGGG CACCCGTCCT GCCCCTTCAC CTTCCAGCTC CGCCTCCTCC GCGCGGACCC CGCCCCGTCC CGACCCCTCC CGGGTCCCCG GCCCAGCCCC CTCCGGGCCC TCCCAGCCCC TCCCCTTCCT TTCCGCGGCC CCGCCCTCTC CTCGCGGCGC GAGTTTCAGG CAGCGCTGCG TCCTGCTGCG CACGTGGGAA GCCCTGGCCC CGGCCACCCC CGCG |
| Artificial hTERT promoter | GGCCCCTCCC TCGGGTTACC CCACAGCCTA GGCCGATTCG ACCTCTCTCC GCTGGGGCCC TCGCTGGCGT CCCTGCACCC TGGGAGCGCG AGCGGCGCGC GGGCGGGGAA GCGCGGCCCA GACCCCCGGG TCCGCCCGGA GCAGCTGCGC TGTCGGGGCC AGGCCGGGCT CCCAGTGGAT TCGCGGGCAC AGACGCCCAG GACCGCGCTC CCCACGTGGC GGAGGGACTG GGGACCCGGG CACCCGTCCT GCCCCTTCAC CTTCCGGCTC CGCCTCCTCC GCGCGGACCC CGCCCCGTCC CGACCCCTTC CGGGTTTCCG GCCCAGCCCC TTCCGGGCCC TCCCAGCCCC TCCCCTTCCT TTCCGGGGCC CCGCCCTCTC CTCGCGGCGC GAGTTTCCGG CAGCGCTGCG TCCTGCTGCG CACGTGGGAA GCCCTGGCCC CGGCCACCCC CGCG |
| Bax | CACCATGGACGGGTC CGGGGAGCAG CCCAGAGGCG GGGGGCCCAC CAGCTCTGAG CAGATCATGA AGACAGGGGC CCTTTTGCTT CAGGGTTTCA TCCAGGATCG AGCAGGGCGA ATGGGGGGGG AGGCACCCGA GCTGGCCCTG GACCCGGTGC CTCAGGATGC GTCCACCAAG AAGCTGAGCG AGTGTCTCAA GCGCATCGGG GACGAACTGG ACAGTAACAT GGAGCTGCAG AGGATGATTG CCGCCGTGGA CACAGACTCC CCCCGAGAGG TCTTTTTCCG AGTGGCAGCT GACATGTTTT CTGACGGCAA CTTCAACTGG GGCCGGGTTG TCGCCCTTTT CTACTTTGCC AGCAAACTGG TGCTCAAGGC CCTGTGCACC AAGGTGCCGG AACTGATCAG AACCATCATG GGCTGGACAT TGGACTTCCT CCGGGAGCGG CTGTTGGGCT GGATCCAAGA CCAGGGTGGT TGGGTGAGAC TCCTCAAGCC TCCTCACCCC CACCACCGCG CCCTCACCAC CGCCCCTGCC CCACCGTCCC TGCCCCCCGC CACTCCTCTG GGACCCTGGG CCTTCTGGAG CAGGTCACAG TGGTGCCCTC TCCCCATCTT CAGATCATCA GATGTGGTCT ATAATGCGTT TTCCTTACGTGTCTGATTCTAGGCGATCG |
| Anti Bcl2 element | AAGGTATATTGCTGTTGACAGTGAGCGCAGGGAGAUAGUGAUGAAGUAATAGTGAAGCCACAGATGTATTACTTCATCACTATCTCCCTTTGCCTACTGCCTCG |
| Negative control element | AAGGTATATTGCTGTTGACAGTGAGCGCAGCGCAUUCCAGCUUACGUAATAGTGAAGCCACAGATGTATTACGTAAGCTGGAATGCGCTTTGCCTACTGCCTCG |
Construction of ETS-1 shRNA vector
Either the small-hairpin RNA (shRNA) targeting ETS-1 or the negative control shRNA targeting no known sequence were cloned into the pGPU/GFP/Neo vector (GenePharma, Shanghai, China). The sequence of ETS-1 shRNA has been previously provided [19]. The ETS-1 shRNA sequence was 5′-CTGATGTAAGGCAATTAAT-3′.
Construction of the artificial hTERT promoter- Bax-Anti Bcl2 combination module
The cDNA sequence of Bax gene was chemically synthesized and cloned into the artificial-hTERT promoter reporter module digested with Nhel and Xhol to substitute the Renilla luciferase gene fragment. Meanwhile, Anti Bcl2 element, which was constructed by using Bcl2 shRNA [20] to replace the mature-miR-30 encoding region of the pre-miR-30 scaffold [21], was inserted into the artificial-hTERT promoter reporter module digested with Pmel and Notl as well. Eventually, the artificial hTERT promoter - Bax-Anti Bcl2 combination module, in which the Bax-Anti Bcl2 combination module was driven by artificial hTERT promoter, has been constructed. Similarly, to construct a negative control, the negative control element, which was also constructed by using the sequence of NC shRNA [20] to replace the corresponding region of the pre-miR-30 scaffold, was inserted into the artificial-hTERT promoter reporter module digested with Pmel and Notl. All the related sequences are shown in Table 1.
Cell transfection
Cells were plated about 20 h prior to transfection to achieve 80–90 % confluency at the time of transfection. Each device was transfected into cells using LipoFiter™ Liposomal Transfection Reagent (Hanbio, Shanghai, China) according to the manufacturer’s instructions in different experiments.
Dual luciferase reporter assay
Cells were seeded in 24-well plates (1 × 105/well) and transfected with the relevant designed device. Luciferase activity was detected using the dual luciferase assay system (Promega, Madison, WI, USA) according to the manufacturer’s instructions at 48 h after transfection. In order to calculate the driven efficiency of artificial or wild-type hTERT promoter in bladder cancer cells and NHF, the renilla luciferase activity was normalized to the firefly luciferase activity. The relative luciferase activity (%) of artificial hTERT promoter-driven reporter module co-transfected with ETS-1 shRNA or negative control shRNA was calculated by the following formula: Relative luciferase activity (%) = (luciferase activity of artificial hTERT promoter-driven reporter module co-transfected with ETS-1 shRNA/luciferase activity of artificial hTERT promoter-driven reporter module co-transfected with negative control shRNA) × 100 %. The experiments were performed in duplicate and repeated at least three times.
CCK-8 assay
The effects of the designed devices on cell proliferation were examined by Cell Counting Kit-8 (Beyotime, Shanghai, China) according to the manufacturer’s instructions. Cells were grown in a 96-well plate (5 × 103/well) for 24 h, transfected with artificial hTERT promoter-Bax-Anti Bcl2 combination module or negative control and cultured in normal medium. Zero, 24, 48, or 72 h post-transfection, 15 μl of CCK-8 was added to each well of a 96-well plate and the cells were seed for 1 h. Absorbance was measured at a wavelength of 450 nm using an ELISA microplate reader (Bio-Rad, Hercules, CA, USA). Assays was repeated at least three times.
MTT assay
The cell proliferation was also examined by using 3-[4, 5-dimethylthiazol-2-yl]-2, 5-diphenyl-tetrazolium bromide (MTT) assay. Cells were seed in a 96-well plate (5 × 103/well) for 24 h, transfected with artificial hTERT promoter-Bax-Anti Bcl2 combination module or negative control and cultured in normal medium. At 0, 24, 48, and 72 h after transfection, cells were cultured in 5 mg/ml MTT for 4 h and lysed in dimethyl sulfoxide (DMSO) at room temperature for 10 min. The absorbance in each well was calculated at a wavelength of 490 nm using an ELISA microplate reader (Bio-Rad, Hercules, CA, USA). The experiments were carried out in duplicate and repeated at least three times.
Caspase 3 ELISA assay
Cells were seeded in 24-well plates (1 × 105/well) and transfected with artificial hTERT promoter-Bax-Anti Bcl2 combination module or negative control. Forty-eight hours after transfection, cell apoptosis were detected by calculating the activity of caspase-3 using the Caspase-3 enzyme-linked immunosorbent assay (ELISA) assay kit (Hcusabio, Wuhan, China) according to the manufacturer’s instructions. Absorbance was measured at a wavelength of 450 nm using a microplate reader (Bio-Rad, Hercules, CA, USA). Data were shown as the ratios between the absorbance of artificial hTERT promoter-Bax-Anti Bcl2 combination module transfected cells and those of negative control transfected cells. Experiments were performed at least three times.
Flow cytometry analysis
Cell apoptosis was detected using an Alexa Fluor®488 Annexin V/Dead Cell Apoptosis Kit (Invitrogen, Carlsbad, CA, USA) according to the supplier’s protocols. Cells were seeded in 6-well plates (5 × 105/well). Forty-eight hours post-transfection, cells were harvested, centrifuged, and washed in 1000 μl cold phosphate-buddered saline (PBS) for two times. Then cells in each tube were resuspended in 100 μl 1 × annexin-binding buffer. Five microliter Alexa Fluor® 488 annexin V and 1 μl PI working solution (100 μg/ml) were added to each tube. The tubes were incubated in the dark on ice for 15 min. Cell apoptosis assay was performed immediately on a flow cytometry (EPICS, XL-4, Beckman, CA, USA). Each experiment was done at least three times.
Statistical analyses
All experimental data from three independent experiments were presented as mean ± standard deviation (SD). All statistical data were analyzed by SPSS 19.0 software (SPSS Inc. Chicago, IL, USA). The data of CCK-8 assay and MTT assay were analyzed by ANOVA. The independent samples t-test was used to analyze other data. A P value of less than 0.05 was considered to be statistically significant.
Results
Artificial hTERT promoter can drive the expression of downstream gene efficiently and selectively in bladder cancer cells
The artificial hTERT promoter was designed and constructed through the approaches of synthetic biology, and the related information was shown in “Methods” section. The designed principle and the action model of synthetic Bax-Anti Bcl2 combination module were shown in Fig. 1.
Fig. 1.

Design and construction of the artificial hTERT promoter-Bax-Anti Bcl2 combination module. In the cancer cell, the expression level of transcription factor ETS-1 is high and thus the artificial promoter is active. In contrast, ETS-1 is absent and the artificial promoter is inactive in the normal cells. The apoptotic markers can only be activated in the cancer cells which have a high ETS-1 level
Bladder cancer cells and normal human fibroblasts (NHF) were cultured and transfected with artificial hTERT promoter-driven reporter module or wide-type hTERT promoter-driven reporter module. Forty-eight hours later, the activity of the artificial hTERT promoter and wild-type hTERT promoter were detected using dual luciferase assay. As shown in Fig. 2, the driven efficiency of artificial hTERT promoter was significantly higher than that of wild-type hTERT promoter in bladder cancer cells 5637, T24, UMUC-3, RT4, J82 and SW780. The average fold-change was about six-fold. Especially in bladder cancer 5637 cells, the fold-change was up to 14-fold. Oppositely, both the activity of artificial hTERT promoter and that of wild-type hTERT promoter were weak in NHF. There was no difference between the activity of artificial hTERT promoter and that of wild-type hTERT promoter in NHF. The data indicated that artificial hTERT promoter could drive the expression of downstream gene efficiently and selectively in bladder cancer cell.
Fig. 2.

Artificial hTERT promoter can drive downstream gene expression efficiently and selectively in bladder cancer cell. Luciferase activity was detected using dual luciferase assay system at 48 h after transfection. The activity of artificial hTERT promoter was significantly higher than that of wild-type hTERT promoter in bladder cancer cells. Nevertheless, both the activity of artificial hTERT promoter and that of wild-type hTERT promoter were weak in NHF. Data are indicated as mean ± S.D.
Silencing ETS-1 inhibited the activity of artificial hTERT promoter in bladder cancer cells
To confirm whether ETS-1 is associated with the driven efficiency of artificial hTERT promoter, bladder cancer cells, 5637, T24, UMUC-3 and RT4, were cultivated and co-transfected with the ETS-1 shRNA and artificial hTERT promoter reporter module or the negative control shRNA and artificial hTERT promoter reporter module. Forty-eight hours post-transfection, dual luciferase assay was used to detect the activity of the artificial hTERT promoter. Compared with the group which was co-transfected with negative control shRNA, the activity of artificial hTERT promoter was respectively decreased by nearly 69 % in bladder cancer 5637 cells, 77 % in bladder cancer T24 cells, 32 % in bladder cancer UMUC-3 cells and 80 % in bladder cancer RT4 cells in the group which was co-transfected with ETS-1 shRNA (Fig. 3). Silencing ETS-1 observably inhibited the activity of artificial hTERT promoter in bladder cancer cell. The data demonstrated that ETS-1 was associated with the driven efficiency of artificial hTERT promoter. ETS-1 is likely to regulate the driven efficiency of artificial hTERT promoter in bladder cancer cells.
Fig. 3.
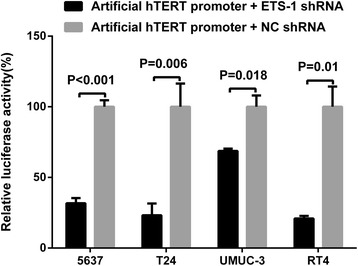
Silencing ETS-1 inhibited the activity of artificial hTERT promoter in bladder cancer cells. Dual luciferase assay system was used to detect the activity of artificial hTERT promoter at 48 h after transfection. The activity of artificial hTERT promoter in the group co-transfected with ETS-1 shRNA was obviously lower than that in the group co-transfected with negative control shRNA. All data are shown as mean ± S.D.
Artificial hTERT promoter-Bax-Anti Bcl2 combination module selectively restrained cell proliferation in bladder cancer cells
Base on the confirmed high efficiency and specificity of artificial hTERT promoter, artificial hTERT promoter-Bax-Anti Bcl2 combination module has been constructed to determine whether this module selectively inhibits cell proliferation in bladder cancer. After transfection with artificial hTERT promoter-Bax-Anti Bcl2 combination module or negative control, bladder cancer 5637 cells, T24 cells and NHF were analyzed by the cell proliferation assay with CCK-8 and MTT. Compared with the negative control, the cell proliferation was significantly suppressed in bladder cancer 5637 cells (Fig. 4a and d) and T24 cells (Fig. 4b and e) transfected with artificial hTERT promoter-Bax-Anti Bcl2 combination module (P < 0.05). As expected, there was no difference in the cell proliferation rate between artificial hTERT promoter-Bax-Anti Bcl2 combination module and negative control transfected group in NHF (Fig. 4c and f) (P > 0.05). These results indicated that artificial hTERT promoter-Bax-Anti Bcl2 combination module selectively suppress cell proliferation in bladder cancer cells.
Fig. 4.
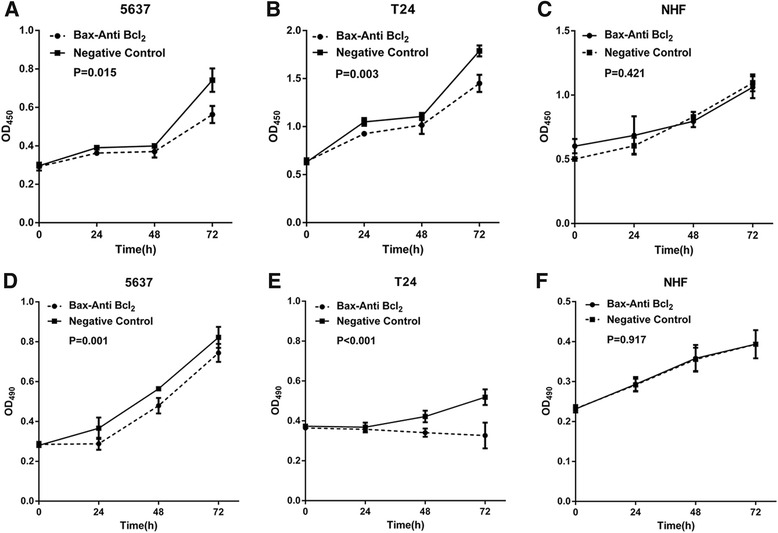
Artificial hTERT promoter-Bax-Anti Bcl2 combination module selectively restrained cell proliferation in bladder cancer cells. Cell proliferation assay (CCK-8 and MTT) were performed as described in Methods. The cell proliferation was significantly suppressed in bladder cancer 5637 cells (a, c) and T24 cells (b, d) which were transfected with artificial hTERT promoter-Bax-Anti Bcl2 combination module (P < 0.05), compared with negative control group. However, no significant change was shown in NHF (c, e) (P > 0.05). Data are indicated as mean ± S.D.
Artificial hTERT promoter-Bax-Anti Bcl2 combination module selectively induced cell apoptosis in bladder cancer cells
Finally, we explored whether cell apoptosis of bladder cancer cells were selectively induced by artificial hTERT promoter-Bax-Anti Bcl2 combination module. At 48 h after transfection of Artificial hTERT promoter-Bax-Anti Bcl2 combination module or negative control, the relative activity of caspase-3 and the apoptosis ratio in bladder cancer 5637 cells, T24 cells, and NHF were detected by the caspase-3 enzyme-linked immunosorbent assay (ELISA) assay and the flow cytometry analysis. Induced cell apoptosis was observed in bladder cancer 5637 cells (Fig. 5a, d and g) and T24 cells (Fig. 5b, e and h) transfected with artificial hTERT promoter-Bax-Anti Bcl2 combination module. On the contrary, there was no difference in the relative activity of caspase-3 and the cell apoptosis rate between artificial hTERT promoter-Bax-Anti Bcl2 combination module and negative control transfected group in NHF (Fig. 5c, f and i). These results confirmed that artificial hTERT promoter-Bax-Anti Bcl2 combination module selectively induced cell apoptosis in bladder cancer cells.
Fig. 5.
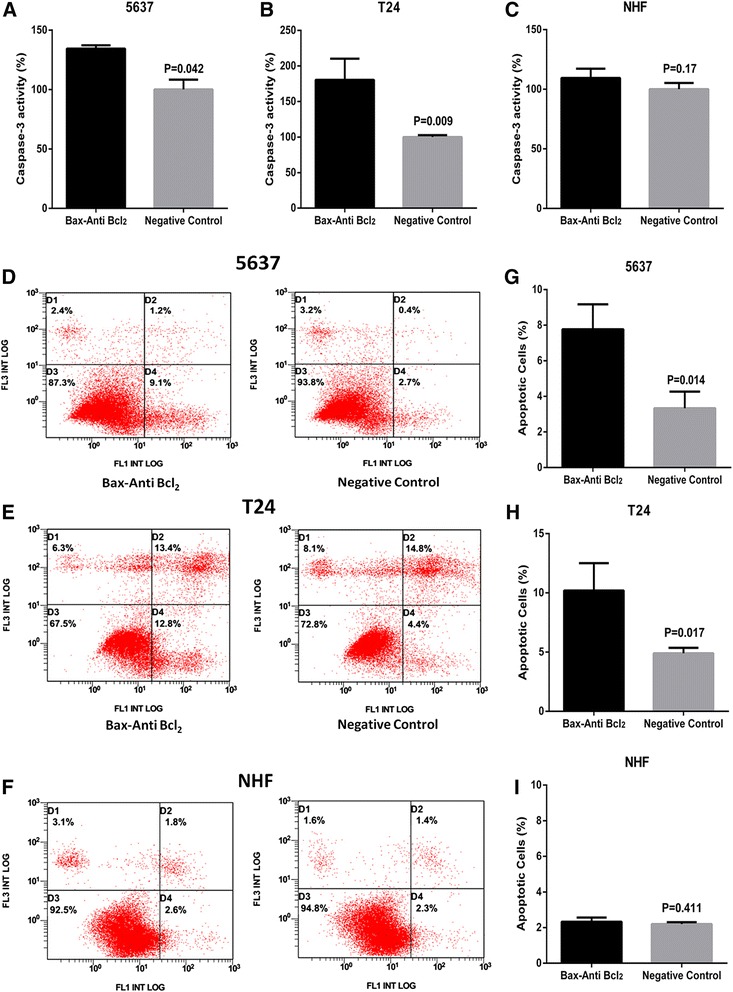
Artificial hTERT promoter-Bax-Anti Bcl2 combination module selectively induced cell apoptosis in bladder cancer cells. The cell apoptosis changes were determined by caspase-3 ELISA assay and flow cytometry analysis. The relative activity of caspase-3 was increased in bladder cancer 5637 (a) cells and T24 (b) cells treated with artificial hTERT promoter-Bax-Anti Bcl2 combination module. Cell apoptosis induction were also observed in bladder cancer 5637 (d) cells and T24 (e) cells treated with artificial hTERT promoter-Bax-Anti Bcl2 combination module using flow cytometry analysis. Representative images of flow cytometry analysis in bladder cancer 5637 cells (g) and T24 cells (h) were also shown. Nevertheless, there was no obvious difference with the relative activity of caspase-3 (c) and cell apoptosis induction (f, i) between artificial hTERT promoter-Bax-Anti Bcl2 combination module and negative control in NHF. D1, cell debris. D2, non-viable apoptotic cell. D3, normal cells. D4, viable apoptotic cell. All data are presented as mean ± SD
Discussion
Rationally designed synthetic systems that are capable of performing complex functions pave the way for translational applications, including disease diagnostics and targeted therapeutics [22]. With millions of new cases for cancer diagnosis, synthetic biology not only can provide new methods for cancer therapy, but also can study pathways, develop novel diagnostic tools, and address drug delivery issues [6]. Engineered elements for the construction of synthetic biology systems could respond to relative metabolites or other relative cues such as DNA, RNA and proteins [6]. In our team, we devote ourselves to translate the basic cancer research with synthetic biology technology. Usually, we would like to use the promoters, terminators and shRNA/miRNA expression scaffolds as the tools to construct complex devices for cancer treatment [23–25].
Our objective for bladder cancer therapy is to kill cancer cells without affecting the normal cells. So it is necessary to use some tumor- specific genetic parts for the construction of treatment devices. For example, above 85 % of malignancies over-express telomerase, whereas the normal somatic cells doesn’t express telomerase [26]. And various studies also reported that telomerase is up-regulated in about 90 % of bladder cancer cells [27]. The hTERT promoter region is the paramount element for the expression of telomerase [28], in which several factors can regulate the expression of telomerase by occupying the binding motifs of hTERT promoter [29–31]. In addition, the mutations of hTERT promoter were found in almost all kinds of cancers but not in most human normal cells [32, 33]. As the two significant genes in the cell apoptotic pathway, the role of Bax and Bcl2 has been well-studied and the ratio of Bcl2/Bax was closely related to the sensitivity of cells apoptosis [13]. Some previous researches also reported that several factors could reduce Bcl2 levels and elevate Bax levels to induce cell apoptosis [14–18]. Reversing the ratio of Bcl2/Bax might affect the bladder cancer cells. All the above information reminds us that we can construct a combination module of Bax over-expression element and Bcl2 interference element driven by an artificial hTERT promoter and this module should have a high transcriptional activity and specificity for the bladder cancer treatment.
In this study, to verify the high transcriptional activity and specificity of the artificial hTERT promoter, we constructed the artificial hTERT promoter-driven reporter module at first. The result indicated that artificial hTERT promoter could efficiently and selectively drive the expression of downstream gene in bladder cancer cell. The mutant hTERT promoter can enhance the expression of downstream gene and maintain its tumor-specific feature. The previous research had reported that the 124G > A and 146G > A mutations strongly enhanced the combination between the hTERT promoter and ETS-1 in the mutant T24 cells [7]. Therefore, to investigate whether the activity of the artificial hTERT promoter was also regulated by ETS-1, we co-transfected ETS-1 shRNA and the artificial hTERT promoter-driven reporter module into bladder cancer cells and tested the expression of the dual luciferase. Silencing ETS-1 observably suppressed the activity of artificial hTERT promoter in bladder cancer cell. From our unpublished data, the expression levels of ETS-1 were significantly up-regulated in bladder cancer tissues compared with matched normal tissues. It demonstrated that ETS-1 was associated with the transcriptional activity of artificial hTERT promoter. ETS-1 might regulate the transcriptional activity of artificial hTERT promoter in bladder cancer cells.
Finally, the artificial hTERT promoter-Bax-Anti Bcl2 combination module was constructed and tested in the bladder cancer cells and normal human fibroblasts. The results showed that this module could inhibit the cell proliferation and induce the cell apoptosis in bladder cancer cells, but not in the normal human fibroblasts. The Bax-Anti Bcl2 combination module driven by artificial hTERT promoter could availably and selectively over-express Bax gene and interference sequence of Bcl2 to reverse the ratio of Bcl2/Bax for intervention of malignant phenotype of bladder cancer cells. In brief, the Bax-Anti Bcl2 combination module driven by artificial hTERT promoter selectively suppresses malignant phenotypes of bladder cancer cells. It would therefore be of great interest to extend this module to clinical research when the technologies for efficient in vivo gene delivery are further developed.
Conclusions
In summary, the artificial hTERT promoter is a cancer-specific promoter that robustly induces the expression of synthetic modules. The oncogenic signal ETS-1 actuates this artificial cancer-specific promoter. The Bax-Anti Bcl2 combination module can effectively inhibits the malignant phenotypes of bladder cancer.
Acknowledgements
The authors are appreciative to the donors, whose names were not included in the author list, but who participated in this program. This work was funded by the National Key Basic Research Program of China (973 Program) (2014CB745201), National Natural Science Foundation of China [81402103,81502684], International S&T Cooperation program of China (ISTCP) (2014DFA31050), the Chinese High-Tech (863) Program (2014AA020607), The National Science Foundation Projects of Guangdong Province (2014A030313717), the Shenzhen Municipal Government of China (ZDSYS201504301722174, JCYJ20150330102720130, GJHZ20150316154912494), and Special Support Funds of Shenzhen for Introduced High-Level Medical Team.
Footnotes
Li Liu, Yuchen Liu, Tianbiao Zhang and Hanwei Wu contributed equally to this work.
Competing interests
There is no conflict of interest between the authors.
Authors’ contributions
LL and YL conceived and designed the experiments. LL, TZ, HW and ML performed the experiments. CW, YZ and QZ coordinated the research and analyzed the data. BQ, XS and GZ supported the experiments and helped to draft the manuscript. QZ and XG performed the statistical analysis. LL, YL and WZ wrote the manuscript. WH provided the financial support and supervised laboratorial processes. All authors read and approved the final manuscript.
Contributor Information
Li Liu, Email: sumc_liuli2013@163.com.
Yuchen Liu, Email: liuyuchenmdcg@163.com.
Tianbiao Zhang, Email: zhangtianbiao215@126.com.
Hanwei Wu, Email: rayman_wu@126.com.
Muqi Lin, Email: muk_lin@163.com.
Chaoliang Wang, Email: wangchaoliang2008@yahoo.com.
Yonghao Zhan, Email: yonghao_zhan@163.com.
Qing Zhou, Email: zqboy008@163.com.
Baoping Qiao, Email: zdyfynksl@126.com.
Xiaojuan Sun, Email: xiaojuan26@gmail.com.
Qiaoxia Zhang, Email: qiaoxiazhang@163.com.
Xiaoqiang Guo, Email: xiaoqiangguo123@163.com.
Weixing Zhang, Email: zhangweixing1957@yahoo.com.
Weiren Huang, Email: pony8980@163.com.
References
- 1.Burger M, Catto JW, Dalbagni G, Grossman HB, Herr H, Karakiewicz P, et al. Epidemiology and risk factors of urothelial bladder cancer. Eur Urol. 2013;63(2):234–41. doi: 10.1016/j.eururo.2012.07.033. [DOI] [PubMed] [Google Scholar]
- 2.Kim WJ, Bae SC. Molecular biomarkers in urothelial bladder cancer. Cancer Sci. 2008;99(4):646–52. doi: 10.1111/j.1349-7006.2008.00735.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Marta GN, Hanna SA, Gadia R, Correa SF, Silva JL, Carvalho HA. The role of radiotherapy in urinary bladder cancer: current status. Int Braz J Urol. 2012;38(2):144–53. doi: 10.1590/S1677-55382012000200002. [DOI] [PubMed] [Google Scholar]
- 4.Racioppi M, D’Agostino D, Totaro A, Pinto F, Sacco E, D’Addessi A, et al. Value of current chemotherapy and surgery in advanced and metastatic bladder cancer. Urol Int. 2012;88(3):249–58. doi: 10.1159/000335556. [DOI] [PubMed] [Google Scholar]
- 5.Weber W, Fussenegger M. Emerging biomedical applications of synthetic biology. Nat Rev Genet. 2012;13(1):21–35. doi: 10.1038/nrg3094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Shankar S, Pillai MR. Translating cancer research by synthetic biology. Mol Bio Syst. 2011;7:1802–10. doi: 10.1039/c1mb05016h. [DOI] [PubMed] [Google Scholar]
- 7.Wu S, Huang P, Li C, Huang Y, Li X, Wang Y, et al. Telomerase reverse transcriptase gene promoter mutations help discern the origin of urogenital tumors: a genomic and molecular study. Eur Urol. 2014;65(2):274–7. doi: 10.1016/j.eururo.2013.10.038. [DOI] [PubMed] [Google Scholar]
- 8.Saeki H, Kuwano H, Kawaguchi H, Ohno S, Suqimachi K. Expression of ets-1 transcription factor is correlated with penetrating tumor progression in patients with squamous cell carcinoma of the esophagus. Cancer. 2000;89(8):1670–6. doi: 10.1002/1097-0142(20001015)89:8<1670::AID-CNCR4>3.0.CO;2-J. [DOI] [PubMed] [Google Scholar]
- 9.Lin Z, Liu Y, Sun Y, He X. Expression of Ets-1, Ang-2 and maspin in ovarian cancer and their role in tumor angiogenesis. J Exp Clin Cancer Res. 2011;30(1):31. doi: 10.1186/1756-9966-30-31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Sari A, Calli A, Gorqel SN, Altinboqa AA, Kara C, Dincel C, et al. Immunohistochemical determination of ETS-1oncoprotein expression in urothelial carcinomas of the urinary bladder. Appl Immunohistochem Mol Morphol. 2012;20(2):153–8. doi: 10.1097/PAI.0b013e31821ba035. [DOI] [PubMed] [Google Scholar]
- 11.Linehan WM, Walther MM, Zbar B. The genetic basis of cancer of the kidney. J Urol. 2003;170(6 Pt 1):2163–72. doi: 10.1097/01.ju.0000096060.92397.ed. [DOI] [PubMed] [Google Scholar]
- 12.Youle RJ, Strasser A. The BCL-2 protein family: opposing activities that mediate cell death. Nat Rev Mol Cell Bio. 2008;9(1):47–59. doi: 10.1038/nrm2308. [DOI] [PubMed] [Google Scholar]
- 13.Oltvai ZN, Milliman CL, Korsmeyer SJ. Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that accelerates programmed cell death. Cell. 1993;74(4):609–19. doi: 10.1016/0092-8674(93)90509-O. [DOI] [PubMed] [Google Scholar]
- 14.Yang G, Zhang S, Zhang Y, Zhou Q, Peng S, Zhang T, et al. The inhibitory effects of extracellular ATP on the growth of nasopharyngeal carcinoma cells via P2Y2 receptor and osteopontin. J Exp Clin Cancer Res. 2014;33(1):53. doi: 10.1186/1756-9966-33-53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Zhao S, Wang Q, Li Z, Ma X, Wu L, Ji H, et al. LDOC1 inhibits proliferation and promotes apoptosis by repressing NF-κB activation in papillary thyroid carcinoma. J Exp Clin Cancer Res. 2015;34:146. doi: 10.1186/s13046-015-0265-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Jaqadish N, Parashar D, Gupta N, Aqarwal S, Purohit S, Kumar V, et al. A-kinase anchor protein 4 (AKAP4) a promising therapeutic target of colorectal cancer. J Exp Clin Cancer Res. 2015;34(1):142. doi: 10.1186/s13046-015-0258-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Wu T, Wang S, Wu J, Lin Z, Sui X, Xu X, et al. Icaritin induces lytic cytotoxicity in extranodal NK/T-cell lymphoma. J Exp Clin Cancer Res. 2015;34(1):17. doi: 10.1186/s13046-015-0133-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Yang T, Shi R, Chang L, Tang K, Chen K, Yu G, et al. Huachansu suppresses human bladder cancer cell growth through the Fas/Fasl and TNF- alpha/TNFR1 pathway in vitro and in vivo. J Exp Clin Cancer Res. 2015;34(1):21. doi: 10.1186/s13046-015-0134-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hollenhorst PC, Chandler KJ, Poulsen RL, Johnson WE, Speck NA, Graves BJ. DNA specificity determinants associate with distinct transcription factor functions. PLoS Genet. 2009;5(12) doi: 10.1371/journal.pgen.1000778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Senft D, Berking C, Graf SA, Kammerbauer C, Ruzicka T, Besch R. Selective induction of cell death in melanoma cell lines through targeting of Mcl-1 and A1. PLoS One. 2012;7(1) doi: 10.1371/journal.pone.0030821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Liu XY, Tang QS, Chen HC, Jiang XL, Fang H. Lentiviral miR30-based RNA interference against heparanase suppresses melanoma metastasis with lower liver and lung toxicity. Int J Biol Sci. 2013;9(6):564–77. doi: 10.7150/ijbs.5425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Chen YY, Smolke CD. From DNA to targeted therapeutics: bringing synthetic biology to the clinic. Sci Transl Med. 2011;3(106):8470. doi: 10.1126/scitranslmed.3002944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Fu X, Liu Y, Zhuang C, Liu L, Cai Z, Huang W. Synthetic artificial microRNAs targeting UCA1-MALAT1 or c-Myc inhibit malignant phenotypes of bladder cancer cells T24 and 5637. Mol Bio Syst. 2015;11(5):1285–9. doi: 10.1039/c5mb00127g. [DOI] [PubMed] [Google Scholar]
- 24.Zhuang C, Fu X, Liu L, Liu YC, Huang WR, Cai ZM. Synthetic miRNA sponges driven by mutant hTERT promoter selectively inhibit the progression of bladder cancer. Tumour Biol. 2015;36(7):5157–63. doi: 10.1007/s13277-015-3169-9. [DOI] [PubMed] [Google Scholar]
- 25.Liu Y, Zeng Y, Liu L, Zhuang C, Fu X, Huang W, et al. Synthesizing AND gate genetic circuits based on CRISPR-Cas9 for identification of bladder cancer cells. Nat Commun. 2014;5:5393. doi: 10.1038/ncomms6393. [DOI] [PubMed] [Google Scholar]
- 26.Kim NW, Piatyszek MA, Prowse KR, Harley CB, West MD, Ho PL, et al. Specific association of human telomerase activity with immortal cells and cancer. Science. 1994;266(5193):2011–5. doi: 10.1126/science.7605428. [DOI] [PubMed] [Google Scholar]
- 27.Harley CB. Telomerase and cancer therapeutics. Nat Rev Cancer. 2008;8(3):167–79. doi: 10.1038/nrc2275. [DOI] [PubMed] [Google Scholar]
- 28.Cong YS, Wen J, Bacchetti S. The human telomerase catalytic subunit hTERT: organization of the gene and characterization of the promoter. Hum Mol Genet. 1999;8(1):137–42. doi: 10.1093/hmg/8.1.137. [DOI] [PubMed] [Google Scholar]
- 29.Gladych M, Wojtyla A, Rubis B. Human telomerase expression regulation. Biochem Cell Biol. 2011;89(4):359–76. doi: 10.1139/o11-037. [DOI] [PubMed] [Google Scholar]
- 30.Wojtyla A, Gladych M, Rubis B. Human telomerase activity regulation. Mol Biol Rep. 2011;38(5):3339–49. doi: 10.1007/s11033-010-0439-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Adamo R, Comandini A, Aquino A, Bonmassar L, Guqlielmi L, Bonmassar E, et al. The antiretroviral agent saquinavir enhances hTERT expression and telomerase activity in human T leukaemia cells in vitro. J Exp Clin Cancer Res. 2013;32(1):38. doi: 10.1186/1756-9966-32-38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Horn S, Figl A, Rachakonda PS, Fischer C, Sucker A, Gast A, et al. TERT promoter mutations in familial and sporadic melanoma. Science. 2013;339(6122):959–61. doi: 10.1126/science.1230062. [DOI] [PubMed] [Google Scholar]
- 33.Huang FW, Hodis E, Xu MJ, Kryukov GV, Chin L, Garraway LA. Highly recurrent TERT promoter mutations in human melanoma. Science. 2013;339(6122):957–9. doi: 10.1126/science.1229259. [DOI] [PMC free article] [PubMed] [Google Scholar]