

Abstract
Most bacterial toxins derived from chromosomally encoded toxin–antitoxin (TA) systems that have been studied to date appear to protect cells from relatively short pulses of stress by triggering a reversible state of growth arrest. In contrast to many bacterial toxins that are produced as defense mechanisms and secreted from their hosts, TA toxins exert their protective effect from within the cell that produces them. TA toxin-mediated growth arrest is most frequently achieved through their ability to selectively cleave RNA species that participate in protein synthesis. Until very recently, it was thought that the primary conduit for toxin-mediated translation inhibition was cleavage of a single class of RNA, mRNA, thus depleting transcripts and precluding production of essential proteins. This minireview focuses on how the development and implementation of a specialized RNA-seq method to study Mycobacterium tuberculosis TA systems enabled the identification of unexpected RNA targets for toxins, i.e. a handful of tRNAs that are cleaved into tRNA halves. Our result brings to light a new perspective on how these toxins may act in this pathogen and uncovers a striking parallel to signature features of the eukaryotic stress response.
Keywords: toxin, antitoxin, VapC, translation inhibition, angiogenin, Rny1
Recent studies on the emergence of tRNAs as targets for stress inducible bacterial toxin–antitoxin systems are reviewed; these findings are compared to tRNA cleavage in human stress responses and disease.
INTRODUCTION
Bacterial toxin–antitoxin (TA) systems comprise adjacent genes encoding a cognate antitoxin and toxin protein. When harbored in the chromosomes of free-living bacteria, the majority of TA systems appear to coordinately enhance stress survival in their host cell. This is achieved through transcriptional regulation of TA operon expression coupled with posttranslational regulation of antitoxin stability. Stress conditions favor degradation of the antitoxin, resulting in free, active toxin (summarized in Fig. 1). Stress survival is most often imparted through growth arrest after inhibition of translation by an RNA-cleaving toxin. Because most of these endoribonuclease toxins had been shown to cleave mRNAs at specific three-, five- or seven-base recognition sequences using conventional methods (Zhang et al.2003, 2004, 2005; Zhu et al.2008, 2009; Rothenbacher et al.2012; Yamaguchi et al.2012), it was widely believed that mRNAs were the sole target of these TA toxins (Fig. 1). However, our group and others have identified additional RNA targets of TA toxins (Fig. 1), namely rRNA (Vesper et al.2011; Winther and Gerdes 2011; Moll and Engelberg-Kulka 2012; Schifano et al.2013, 2016; Winther et al.2013; Schifano and Woychik 2014) and tRNA (Winther and Gerdes 2011; Cruz et al.2015). These findings suggest that, in general, the spectrum of RNAs that collaborate in TA system-mediated stress survival is more varied than originally proposed.
Figure 1.

Summary of RNA-cleaving TA systems. TA systems are composed of two adjacent genes that are cotranscribed and then translated into a stable toxin protein and a labile antitoxin. Under normal conditions, the toxin and antitoxin proteins can form a stable complex in which the toxin is inactive. This complex, or free antitoxin, can bind to an operator upstream of the toxin and antitoxin genes in order to repress their transcription. Under stress conditions, free active toxin accumulates, cleaves RNA, resulting in bacterial growth arrest. Sustained toxin activity results in cell death. Early studies identified several toxins with activity against mRNA. Thus, mRNA has most often been implicated as a toxin target; however, rRNA and tRNA are recently characterized targets of bacterial TA toxins.
TA systems appear to also facilitate stress survival in the pathogen Mycobacterium tuberculosis. Stress survival is instrumental for the life cycle of this pathogen, and confounds efforts to treat and eradicate tuberculosis. While tuberculosis still causes an estimated 1 million deaths/year worldwide, approximately 90% of those infected with M. tuberculosis—comprising approximately one-third of the world's population—have latent tuberculosis (Dye et al.1999). Individuals with latent tuberculosis harbor M. tuberculosis within granulomas in the lung, created by the immune response to limit growth to a non-replicating persistent state and sequester the infection. However, granulomas serve as a survival niche for the pathogen, because latent infection can revert to active tuberculosis, especially among the immune compromised.
A better understanding of the players in M. tuberculosis growth control is critical to develop antimicrobial therapies that are effective against latent tuberculosis. Since the general function of TA systems is to impart reversible growth inhibition, they are strongly implicated as the molecular switches that enable M. tuberculosis to slow or stop replication and become dormant. Consistent with a role in stress survival, several of the 65 annotated TA systems (Table 1) in the M. tuberculosis genome are induced during stresses relevant to the pathobiology of this organism: heat shock (Stewart et al.2002), hypoxia (Rustad et al.2008; Ramage, Connolly and Cox 2009), DNA damage (Rand et al.2003), nutrient starvation (Betts et al.2002), macrophage infection (Cappelli et al.2006; Korch, Contreras and Clark-Curtiss 2009; Ramage, Connolly and Cox 2009) and antibiotic treatment (Provvedi et al. 2009; Singh, Barry and Boshoff 2010). Also, RNA-seq of nutrient-starved M. tuberculosis revealed that 75% of its TA systems were upregulated to some degree with 25% upregulated 2-fold or higher (Cortes et al.2013).
Table 1.
Toxin-antitoxin systems in M. tuberculosis.
| No. | TA family | Toxin | Mechanism of toxicity | References |
|---|---|---|---|---|
| 50 | VapBC | VapC | Translation; tRNA and rRNA cleavage | Ramage, Connolly and Cox (2009); Ahidjo et al. (2011); Winther et al. (2013); Sala, Bordes and Genevaux. (2014); Cruz et al. (2015) |
| 9 | MazEF | MazF | Translation; mRNA and rRNA cleavage | Pandey and Gerdes (2005); Schifano et al. (2013); Schifano et al. (2014); Tiwari et al. (2015) |
| 3 | RelBE | RelE | Translation; mRNA cleavage at ribosomal A-site | Korch, Contreras and Clark-Curtiss (2009); Singh, Barry and Boshoff (2010); Yang et al. (2010); Miallau et al. (2013); Korch et al. (2015) |
| 2 | ParDE | ParE | Not determined; inhibition of DNA gyrase for ParE from plasmid RK2 and Vibrio cholerae chromosome | Gupta (2009); Ramage, Connolly and Cox (2009); Yuan et al. (2010) |
| 1 | HigBA | HigB | Translation; mRNA and tmRNA cleavage by associating with ribosome | Gupta (2009); Fivian-Hughes and Davis (2010); Schuessler et al. (2013) |
This brief minireview highlights our recent finding that a toxin derived from a VapBC TA system harbored by M. tuberculosis exhibits an activity unique among characterized M. tuberculosis TA toxins. We demonstrated that this toxin, VapC-mt4 (Rv0595c, also known as VapC4), is an isoacceptor-specific tRNase (Cruz et al.2015). VapC-mt4 specifically recognizes and cleaves only 3 of the 45 total tRNAs in M. tuberculosis at a functionally essential site—the anticodon stem-loop—generating stable tRNA halves (Cruz et al.2015). We also integrate our findings into the field overall, discuss the ramifications of this activity in M. tuberculosis cells and examine the striking parallels to the basic features of the eukaryotic stress response.
GENERAL FEATURES OF VapC TOXINS
VapBC TA systems are abundant in bacterial pathogens. In fact, >40% of the approximately 700 TA modules identified in 126 complete genomes of free-living bacteria were VapBC TA family members (Pandey and Gerdes 2005). The multiple copies of VapBC TA systems (though none are identical) are associated with increased virulence, thus they were designated Vap, for virulence associated protein (Katz, Strugnell and Rood 1992). Mycobacterium tuberculosis harbors the highest number of VapBC TA systems among free-living bacteria, with an estimated 50 members in its genome (Ramage, Connolly and Cox 2009; Ahidjo et al.2011). All 50 M. tuberculosis VapC toxins exhibit structure and sequence similarity and possess a PIN (PilT N-terminal) domain. PIN domains are approximately 130 amino acids in length and contain a quartet of acidic residues and a fifth invariant serine or threonine residue that coordinate divalent cation(s) and possess nuclease activity (Arcus, Rainey and Turner 2005; Arcus et al.2011).
Although the presence of the PIN domain provided some clues to VapC toxin function—each of the characterized VapC toxins are endoribonucleases—their primary RNA targets vary. The Shigella/Salmonella (Winther and Gerdes 2011) and Leptospira interrogans (Lopes et al.2014) VapC toxins attack an extremely pivotal player in protein synthesis, tRNAfMet, and inactivate it through cleavage at a single site in the anticodon stem-loop (Fig. 2). Mycobacterium smegmatis VapC cleaves synthetic ssRNAs at a short consensus sequence with some dependence on secondary structure, as well as some mRNAs involved in glycerol uptake and metabolism (McKenzie et al.2012). Among the 50 M. tuberculosis VapC toxins, only two have been characterized in detail. Our laboratory first reported that M. tuberculosis VapC-mt4 cleaves at ACGC or AC(A/U)GC sequences in RNA (Sharp et al.2012), before we were able to pinpoint selected tRNA isoacceptors as targets (Cruz et al.2015). The Gerdes laboratory reported that VapC20 (VapC-mt20 using our convention) cleaves at the highly conserved sarcin–ricin loop of 23S rRNA (Winther et al.2013). Thus, although each VapC targets RNA for cleavage, knowing the function of one VapC toxin does not enable prediction of the targets for the family as a whole. Given that there are 50 of these toxins, the ramifications of differential activity among them are provocative and may shed light on the molecular underpinnings of latent tuberculosis.
Figure 2.

Cleavage of tRNAfMet by VapC inhibits protein synthesis. (A) The VapC proteins from Salmonella enterica serovar LT2 and Shigella flexneri 2a virulence plasmid pMYSH6000 were found to cleave the initiator tRNAfMet within its anticodon stem-loop (yellow arrow). (B) Cleavage of tRNAfMet by VapC results in the generation of tRNA halves and inhibition of protein synthesis.
5′ RNA-Seq TO IDENTIFY RNA TARGETS FOR VapC AND OTHER TA TOXINS
The three VapC-mt4 tRNA targets—tRNAAla2, tRNASer26 and tRNASer24—were identified using a genome-scale approach, a specialized RNA-seq method, 5′ RNA-seq, that we designed specifically for endoribonuclease toxins (Schifano et al.2014; Cruz et al.2015). Conventional bacterial RNA-seq uses massively parallel sequencing technology to sequence the cDNA derived from cellular RNA and is usually intended to survey the entire transcriptome. In contrast, 5′ RNA-seq is designed for differential detection of one or more subpopulations of RNA depending on the modification present at the 5′ end of the transcript. RNAs in bacterial cells have one of three 5′ end modifications: 5′ triphosphate (mRNAs), 5′ monophosphate (rRNAs and tRNAs) or 5′-hydroxyl (non-coding RNA intermediates and products cleaved by certain endoribonucleolytic toxins such as VapC). 5′ RNA-seq enables global analysis of specific populations of RNA transcripts based on their 5′ end; i.e. only those transcripts carrying a 5′-PPP, only those transcripts carrying a 5′-P or only those transcripts carrying a 5′-OH (Goldman et al.2011; Vvedenskaya et al.2012). Therefore, this method enabled us to specifically detect 5′ ends of RNA transcripts that were produced after VapC toxin cleavage (carrying a 5′-OH).
VapC-mt4 REQUIRES A PRECISE SEQUENCE AND STRUCTURE FOR CLEAVAGE
In contrast to the only other characterized M. tuberculosis VapC toxin, VapC-mt20 (which cleaves 23S rRNA at the sarcin–ricin loop; Winther et al.2013), VapC-mt4 specifically targeted just 3 of the 45 total M. tuberculosis tRNAs (tRNAAla2, tRNASer26 and tRNASer24). Each of these tRNAs contains an ACGC or ACUGC cleavage consensus sequence (Sharp et al.2012), indicating that sequence recognition is required for VapC-mt4 cleavage. However, sequence alone cannot account for the selectivity since 13 of the 45 total tRNAs in M. tuberculosis also contain ACGC or ACUGC sequences. VapC-mt4 must also require a specific tertiary fold or recognition of determinants in the sugar-phosphate backbone. Indeed, cleavage efficiency was dependent on an intact anticodon stem-loop; cleavage was precluded upon removal of tRNA secondary structure even though the ACGC sequence was intact (Cruz et al.2015) (Fig. 3A and B).
Figure 3.
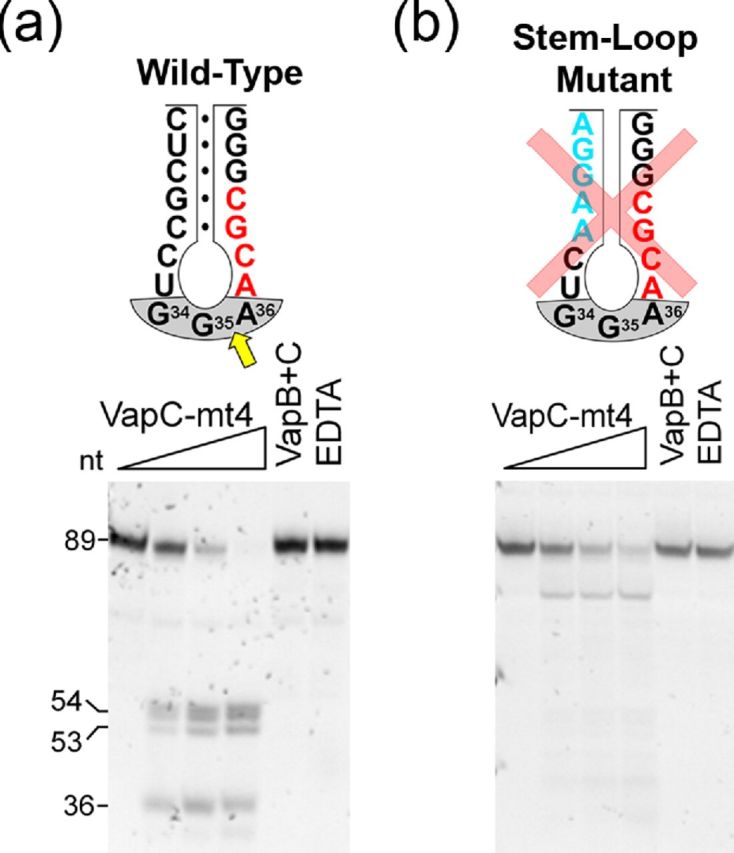
VapC-mt4 requires a consensus sequence within a proper structural context. Cleavage assays comparing wild-type VapC-mt4 cleavage of M. tuberculosis tRNASer24 (A) to a mutant that retains the consensus sequence but removes all base pairing in the stem (B). Mutated bases (blue), cleavage site (yellow arrow), consensus sequence (red), anticodon (gray shaded), base-pairing represented as black dots (•). In vitro synthesized mutant and wild-type tRNASer24 were incubated with increasing amounts of VapC-mt4 (ratios of toxin to RNA were 0:1, 1.25:1, 2.5:1 and 5:1). Control reactions on the right contained the highest concentration of VapC-mt4 preincubated with VapB antitoxin or EDTA before addition of the respective tRNAs. Reactions were incubated at 37°C for 3 h. Sizes of full length and cleaved tRNA products on the left. Convention when numbering tRNA bases dictates that the anticodon is numbered bases 34–36. However, for the actual base numbers for the anticodon are 35–37.
STEM LOOPS AS A COMMON DENOMINATOR FOR ALL VapC TARGETS
Even though VapC toxins do not target the same RNAs, each appears to exhibit a preference for stem-loop structures. 23S rRNA cleavage at the sarcin–ricin loop by VapC-mt20 appears to be more dependent on recognition of the stem-loop structure of the sarcin–ricin loop (Fig. 4A) over sequence alone; although the sequences of each bases 5’ and 3’ of the cleavage site do also matter (Winther et al.2013). Cleavage of tRNAfMet (Fig. 2) occurs in the anticodon stem-loop (Winther and Gerdes 2011; Lopes et al.2014). Although VapC-mt4 does require the presence of the ACGC or ACUGC consensus sequence, it cuts all three M. tuberculosis tRNA targets precisely within the anticodon loop of tRNA (Fig. 4B–D). These studies, in combination with those from the Arcus group (McKenzie et al.2012), revealed that the VapC toxin family is distinct from the majority of TA system toxins that possess endoribonuclease activity (e.g. RelE, MazF, HigB, YafQ) because its members are all predicted to require both sequence and structure elements for target recognition.
Figure 4.

VapC proteins target stem-loop structures in rRNA or tRNA. (A) VapC-mt20 (VapC20) targets the conserved sarcin–ricin loop (SRL) of 23 rRNA. VapC-mt4 cleaves three tRNAs from M. tuberculosis: tRNAAla2 (B), tRNASer26 and (C) tRNASer24. Yellow arrows indicate the position of cleavage by VapC-mt4. tRNA anticodons are shaded in gray and the VapC-mt4 consensus is indicated in red. Note that in tRNASer26 (C) and tRNASer24 (D) the actual base numbers for the anticodon are 35–37.
VapC TOXINS, COLICINS AND OTHER BACTERIAL tRNASES: A COMMON MEANS TO DIFFERENT ENDS
Colicins are bacteriocins produced by strains of Escherichia coli to kill nearby cells and reduce competition for resources (Cascales et al.2007). Colicins are divided into two categories based on their mode of cytotoxicity, pore forming or enzymatic. Enzymatic colicins share mechanistic similarity to VapC toxins because they possess nuclease activity. Two members of this group, colicin D and E5, are tRNases. Death from colicins D and E5 is a consequence of their cleavage of tRNA at a single site within the anticodon stem-loop. Colicin D cleaves all four tRNAArg isoacceptors (Tomita et al.2000); colicin E5 cleaves tRNAHis tRNATyr tRNAAsn tRNAAsp (Ogawa et al.1999). In contrast to colicins, which are defensively secreted from the cell, the E. coli PrrC endoribonuclease initiates cell suicide of its host cell by cleaving tRNALys in response to bacteriophage T4 infection (Amitsur, Levitz and Kaufmann 1987; Kaufmann 2000; Meineke and Shuman 2012).
Therefore, in contrast to the growth regulating (and ostensibly reversible) properties of VapC toxins due to carefully orchestrated association with its cognate antitoxin, colicins D, E5 and PrrC are engineered to kill. Death occurs in neighboring cells that take up colicins D and E5 because these cells do not carry the cognate immunity proteins produced by the colicin-secreting attacking cells (Cascales et al.2007). PrrC-mediated cell death is more subversive. The Stp T4 phage protein alters the association of the ‘antitoxin’ EcoprrI with PrrC, enabling release of free PrrC and unchecked cleavage of lysine tRNA (Amitsur, Levitz and Kaufmann 1987; Kaufmann 2000; Meineke and Shuman 2012).
MIRROR, MIRROR: tRNA CLEAVAGE IN BOTH EUKARYOTIC AND BACTERIAL STRESS RESPONSES
The generation of tRNA halves upon cleavage at the tRNA anticodon stem-loop and smaller fragments derived from these halves are hallmarks of the eukaryotic stress response (reviewed in Thompson and Parker 2009a; Phizicky and Hopper 2010; Gebetsberger and Polacek 2013). tRNA cleavage in response to a diverse array of stresses has been observed in a wide range of organisms. In the pathogenic protozoa Trypanasoma cruzi, tRNA halves are more prevalent upon nutritional stress and associated with specific cytoplasmic granules (Garcia-Silva et al.2010). Other stresses can also induce tRNA cleavage in eukaryotes. Oxidative stress results in the generation of tRNA fragments in Saccharomyces cerevisae, Arabidopsis and human cell lines (Thompson et al.2008). Nitrogen starvation, methionine starvation, heat shock and nearing lag phase growth also result in increased production of tRNA halves in S. cerevisae (Thompson et al.2008). In mammalian cells it is known that heat shock, hypothermia, hypotonic stress and hypoxia also led to the accumulation of tRNA halves (Fu et al.2009; Saikia et al.2012). Note that cleavage of tRNA is not a general response to all stresses. For example, tRNA cleavage is not detected in γ-irradiated, caffeine-treated or etoposide-treated human cells (Thompson et al.2008; Fu et al.2009; Yamasaki et al.2009) or in response to UV irradiation, amino acid or glucose starvation in yeast cells (Thompson et al.2008).
There are two eukaryotic RNases associated with stress-linked generation of tRNA halves, S. cerevisae RNase Rny1p and mammalian angiogenin. Rny1p, a member of the RNase T2 family, cleaves tRNAs within their anticodon stem-loop (Thompson and Parker 2009b). Angiogenin, so named because it was originally found to stimulate angiogenesis, is a member of the RNase A family. Both angiogenin and Rny1p have been proposed to act as sensors of cell damage (Thompson and Parker 2009b). In the absence of stress, Rny1p and angiogenin are sequestered in their respective organelles (secreted or in the vacuole for Rny1p; nucleus/nucleolus for angiogenin) and only released to the cytoplasm upon stress. Thus, angiogenin has been proposed to have opposing roles—either promotion of cell growth or promotion of cell survival—depending on its location in the cell (reviewed in Li and Hu 2012; Saikia and Hatzoglou 2015). To promote cell growth and proliferation, angiogenin translocates to the nucleus/nucleolus where it stimulates rRNA transcription. To promote survival when subjected to certain types of stress, angiogenin accumulates in the cytoplasm and modulates cleavage of tRNA to generate tRNA halves. Although these tRNA halves inhibit most translation, an alternate stress survival mode of translation involving cap-independent internal ribosome entry site initiation is unaffected, resulting in reprogramming of translation (Yamasaki et al.2009; Ivanov et al.2011). Transfection of 5′ tRNA halves alone into cultured cells can also inhibit translation and promote stress granule assembly (Emara et al.2010).
Stress generated tRNA halves or other tRNA-derived fragments studied in a variety of eukaryotes, including humans, have been implicated in a range of important cellular processes, including translational control, apoptosis and RNAi regulation (reviewed in Gebetsberger and Polacek 2013; Raina and Ibba 2014). Therefore, although the signaling pathways and cellular endpoints are clearly distinct, the general features of the stress response are reflected in both the activity of a toxin from the bacterial pathogen M. tuberculosis and eukaryotic tRNases responsible for production of stable tRNA halves and other fragments (Fig. 5).
Figure 5.

tRNA halves are generated in bacteria and eukaryotes. Proteins with known tRNase activity and their general cellular functions are illustrated.
tRNA HALVES, tRNA-DERIVED FRAGMENTS AND DISEASE
There are many intriguing associations between a variety of human diseases and the stable tRNA halves and/or smaller fragments generated by angiogenin (reviewed in Anderson and Ivanov 2014; Kirchner and Ignatova 2015; Saikia and Hatzoglou 2015) (Fig. 6). Angiogenin is important for neuron survival. To date, downregulation of angiogenin is associated with three neurodegenerative diseases—amyotrophic lateral sclerosis (ALS, Lou Gehrig's disease; Komar and Hatzoglou 2011), Parkinson's disease (PD; Komar and Hatzoglou 2011) and Alzheimer's disease (Kim and Kim do 2012). Mutant forms of angiogenin with reduced RNase activity are implicated in the pathogenesis of ALS. Interestingly, some of these angiogenin mutants associated with ALS were also found in patients with PD (van Es et al.2011). Consistent with the connection between angiogenin mutants to these two neurodegenerative diseases, addition of recombinant angiogenin to cultured neurons is protective against hypoxia (Sebastia et al.2009). Angiogenin also increases lifespan and motoneuron survival in an ALS mouse model (Kieran et al.2008).
Figure 6.

Angiogenin can influence mammalian cell growth and survival. Angiogenin (ANG) acts as a stress/damage sensor that can influence cell growth (as demonstrated with certain cancers) or cell survival (as demonstrated with neurons). Angiogenin is upregulated in various cancers and downregulated in the neurodegenerative diseases shown (ALS, amyotrophic lateral sclerosis; PD, Parkinson's disease). The illustration only reflects two of the major processes influenced by angiogenin in this emerging field, not all of its known effects.
Angiogenin is overexpressed in nearly all types of cancer (Li and Hu 2012). However, the significance of this observation is unclear. Genome-scale methods to survey small RNAs are just beginning to track tRNA halves and fragments in certain cancers. For example, deep sequencing of the transcriptomes of prostate cancer patients revealed an increase in tRNA fragments in prostate cancer cells (Martens-Uzunova et al.2012); yet tRNA-derived fragments are downregulated in B-cell lymphoma (Maute et al.2013).
OUTLOOK
Since only two of the estimated 50 VapC toxins from M. tuberculosis have been studied in detail, we have only scratched the surface. VapC-mt4 and VapC-mt20 (VapC20) target different RNAs, tRNA and 23S rRNA sarcin–ricin loop (Winther et al.2013), respectively. Therefore, it is unlikely that the remaining members will be functionally redundant. Instead, each VapC may cleave unique substrates or sites within a given target, thus allowing a defined molecular response to one or more stresses. Given that M. tuberculosis cells can remain dormant yet viable for many years in latent tuberculosis in a granuloma, the resulting activity of each toxin may impart some degree of growth downregulation that is then released before cells reach the point of no return. Thus, through activation of a shifting array of VapC toxins in a calibrated, asynchronous manner, M. tuberculosis may be able to limit growth while never reaching the point of cell death.
M. tuberculosis cells enlist VapC-mt4 toxin for cleavage of a subset of tRNAs to tRNA halves as vehicles for growth regulation. This finding reveals a glimpse into the intricacies of TA toxin selectivity and the power of this toxin activity in determining the fate of a cell. It also brings to light common themes between the stress response in eukaryotes and bacterial pathogens, adding another layer to the daunting diversity of growth modulators that target a finite set of switches within essential molecular machines. Since VapC toxins rely on both sequence and structure determinants for recognition of their target RNAs, they are endowed with exquisite selectivity. Our findings with tRNA cleaving VapC-mt4 portend the same recognition properties for tRNases in humans and other eukaryotes with pivotal roles in a spectrum of essential processes influencing health and disease.
FUNDING
This work was supported by the National Institutes of Health [RO1 GM095693 and R21 AI072399 to N. A. W] and the National Institutes of Health [T32 AI007403 Virus-Host Interactions in Eukaryotic Cells (to J. W. C., awarded to G. Brewer)].
Conflict of interest. None declared.
REFERENCES
- Ahidjo BA, Kuhnert D, McKenzie JL, et al. VapC toxins from Mycobacterium tuberculosis are ribonucleases that differentially inhibit growth and are neutralized by cognate VapB antitoxins. PLoS One. 2011;6:e21738. doi: 10.1371/journal.pone.0021738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amitsur M, Levitz R, Kaufmann G. Bacteriophage T4 anticodon nuclease, polynucleotide kinase and RNA ligase reprocess the host lysine tRNA. EMBO J. 1987;6:2499–503. doi: 10.1002/j.1460-2075.1987.tb02532.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson P, Ivanov P. tRNA fragments in human health and disease. FEBS Lett. 2014;588:4297–4304. doi: 10.1016/j.febslet.2014.09.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arcus VL, McKenzie JL, Robson J, et al. The PIN-domain ribonucleases and the prokaryotic VapBC toxin-antitoxin array. Protein Eng Des Sel. 2011;24:33–40. doi: 10.1093/protein/gzq081. [DOI] [PubMed] [Google Scholar]
- Arcus VL, Rainey PB, Turner SJ. The PIN-domain toxin-antitoxin array in mycobacteria. Trends Microbiol. 2005;13:360–5. doi: 10.1016/j.tim.2005.06.008. [DOI] [PubMed] [Google Scholar]
- Betts JC, Lukey PT, Robb LC, et al. Evaluation of a nutrient starvation model of Mycobacterium tuberculosis persistence by gene and protein expression profiling. Mol Microbiol. 2002;43:717–31. doi: 10.1046/j.1365-2958.2002.02779.x. [DOI] [PubMed] [Google Scholar]
- Cappelli G, Volpe E, Grassi M, et al. Profiling of Mycobacterium tuberculosis gene expression during human macrophage infection: upregulation of the alternative sigma factor G, a group of transcriptional regulators, and proteins with unknown function. Res Microbiol. 2006;157:445–55. doi: 10.1016/j.resmic.2005.10.007. [DOI] [PubMed] [Google Scholar]
- Cascales E, Buchanan SK, Duche D, et al. Colicin biology. Microbiol Mol Biol R. 2007;71:158–229. doi: 10.1128/MMBR.00036-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cortes T, Schubert OT, Rose G, et al. Genome-wide mapping of transcriptional start sites defines an extensive leaderless transcriptome in Mycobacterium tuberculosis. Cell Rep. 2013;5:1121–31. doi: 10.1016/j.celrep.2013.10.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cruz JW, Sharp JD, Hoffer ED, et al. Growth-regulating Mycobacterium tuberculosis VapC-mt4 toxin is an isoacceptor-specific tRNase. Nat Commun. 2015;6:7480. doi: 10.1038/ncomms8480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dye C, Scheele S, Dolin P, et al. Consensus statement. Global burden of tuberculosis: estimated incidence, prevalence, and mortality by country. WHO Global Surveillance and Monitoring Project. J Am Med Assoc. 1999;282:677–86. doi: 10.1001/jama.282.7.677. [DOI] [PubMed] [Google Scholar]
- Emara MM, Ivanov P, Hickman T, et al. Angiogenin-induced tRNA-derived stress-induced RNAs promote stress-induced stress granule assembly. J Biol Chem. 2010;285:10959–68. doi: 10.1074/jbc.M109.077560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fivian-Hughes AS, Davis EO. Analyzing the regulatory role of the HigA antitoxin within Mycobacterium tuberculosis. J Bacteriol. 2010;192:4348–56. doi: 10.1128/JB.00454-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fu Z, Tamber S, Memmi G, et al. Overexpression of MazFsa in Staphylococcusaureus induces bacteriostasis by selectively targeting mRNAs for cleavage. J Bacteriol. 2009;191:2051–9. doi: 10.1128/JB.00907-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garcia-Silva MR, Frugier M, Tosar JP, et al. A population of tRNA-derived small RNAs is actively produced in Trypanosomacruzi and recruited to specific cytoplasmic granules. Mol Biochem Parasitol. 2010;171:64–73. doi: 10.1016/j.molbiopara.2010.02.003. [DOI] [PubMed] [Google Scholar]
- Gebetsberger J, Polacek N. Slicing tRNAs to boost functional ncRNA diversity. RNA Biol. 2013;10:1798–806. doi: 10.4161/rna.27177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldman SR, Sharp JS, Vvedenskaya IO, et al. NanoRNAs prime transcription initiation in vivo. Mol Cell. 2011;42:817–25. doi: 10.1016/j.molcel.2011.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gupta A. Killing activity and rescue function of genome-wide toxin-antitoxin loci of Mycobacterium tuberculosis. FEMS Microbiol Lett. 2009;290:45–53. doi: 10.1111/j.1574-6968.2008.01400.x. [DOI] [PubMed] [Google Scholar]
- Ivanov P, Emara MM, Villen J, et al. Angiogenin-induced tRNA fragments inhibit translation initiation. Mol Cell. 2011;43:613–23. doi: 10.1016/j.molcel.2011.06.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katz ME, Strugnell RA, Rood JI. Molecular characterization of a genomic region associated with virulence in Dichelobacternodosus. Infect Immun. 1992;60:4586–92. doi: 10.1128/iai.60.11.4586-4592.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaufmann G. Anticodon nucleases. Trends Biochem Sci. 2000;25:70–4. doi: 10.1016/s0968-0004(99)01525-x. [DOI] [PubMed] [Google Scholar]
- Kieran D, Sebastia J, Greenway MJ, et al. Control of motoneuron survival by angiogenin. J Neurosci. 2008;28:14056–61. doi: 10.1523/JNEUROSCI.3399-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim YN, Kim do H. Decreased serum angiogenin level in Alzheimer's disease. Prog Neuro-Psychoph. 2012;38:116–20. doi: 10.1016/j.pnpbp.2012.02.010. [DOI] [PubMed] [Google Scholar]
- Kirchner S, Ignatova Z. Emerging roles of tRNA in adaptive translation, signalling dynamics and disease. Nat Rev Genet. 2015;16:98–112. doi: 10.1038/nrg3861. [DOI] [PubMed] [Google Scholar]
- Komar AA, Hatzoglou M. Cellular IRES-mediated translation: the war of ITAFs in pathophysiological states. Cell Cycle. 2011;10:229–40. doi: 10.4161/cc.10.2.14472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Korch SB, Contreras H, Clark-Curtiss JE. Three Mycobacterium tuberculosis Rel toxin-antitoxin modules inhibit mycobacterial growth and are expressed in infected human macrophages. J Bacteriol. 2009;191:1618–30. doi: 10.1128/JB.01318-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Korch SB, Malhotra V, Contreras H, et al. The Mycobacterium tuberculosis relBE toxin:antitoxin genes are stress-responsive modules that regulate growth through translation inhibition. J Microbiol. 2015;53:783–95. doi: 10.1007/s12275-015-5333-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li S, Hu GF. Emerging role of angiogenin in stress response and cell survival under adverse conditions. J Cell Physiol. 2012;227:2822–6. doi: 10.1002/jcp.23051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopes AP, Lopes LM, Fraga TR, et al. VapC from the leptospiral VapBC toxin-antitoxin module displays ribonuclease activity on the initiator tRNA. PLoS One. 2014;9:e101678. doi: 10.1371/journal.pone.0101678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKenzie JL, Robson J, Berney M, et al. A VapBC toxin-antitoxin module is a posttranscriptional regulator of metabolic flux in mycobacteria. J Bacteriol. 2012;194:2189–204. doi: 10.1128/JB.06790-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martens-Uzunova ES, Jalava SE, Dits NF, et al. Diagnostic and prognostic signatures from the small non-coding RNA transcriptome in prostate cancer. Oncogene. 2012;31:978–91. doi: 10.1038/onc.2011.304. [DOI] [PubMed] [Google Scholar]
- Maute RL, Schneider C, Sumazin P, et al. tRNA-derived microRNA modulates proliferation and the DNA damage response and is down-regulated in B cell lymphoma. P Natl Acad Sci USA. 2013;110:1404–9. doi: 10.1073/pnas.1206761110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meineke B, Shuman S. Structure-function relations in the NTPase domain of the antiviral tRNA ribotoxin Escherichia coli PrrC. Virology. 2012;427:144–50. doi: 10.1016/j.virol.2012.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miallau L, Jain P, Arbing MA, et al. Comparative proteomics identifies the cell-associated lethality of M. tuberculosis RelBE-like toxin-antitoxin complexes. Structure. 2013;21:627–37. doi: 10.1016/j.str.2013.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moll I, Engelberg-Kulka H. Selective translation during stress in Escherichia coli. Trends Biochem Sci. 2012;37:493–8. doi: 10.1016/j.tibs.2012.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ogawa T, Tomita K, Ueda T, et al. A cytotoxic ribonuclease targeting specific transfer RNA anticodons. Science. 1999;283:2097–100. doi: 10.1126/science.283.5410.2097. [DOI] [PubMed] [Google Scholar]
- Pandey DP, Gerdes K. Toxin-antitoxin loci are highly abundant in free-living but lost from host-associated prokaryotes. Nucleic Acids Res. 2005;33:966–76. doi: 10.1093/nar/gki201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phizicky EM, Hopper AK. tRNA biology charges to the front. Gene Dev. 2010;24:1832–60. doi: 10.1101/gad.1956510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Provvedi R, Boldrin F, Falciani F, et al. Global transcriptional response to vancomycin in Mycobacterium tuberculosis. Microbiology. 2009;155:1093–102. doi: 10.1099/mic.0.024802-0. [DOI] [PubMed] [Google Scholar]
- Raina M, Ibba M. tRNAs as regulators of biological processes. Front Genet. 2014;5:171. doi: 10.3389/fgene.2014.00171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramage HR, Connolly LE, Cox JS. Comprehensive functional analysis of Mycobacterium tuberculosis toxin-antitoxin systems: implications for pathogenesis, stress responses, and evolution. PLoS Genet. 2009;5:e1000767. doi: 10.1371/journal.pgen.1000767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rand L, Hinds J, Springer B, et al. The majority of inducible DNA repair genes in Mycobacterium tuberculosis are induced independently of RecA. Mol Microbiol. 2003;50:1031–42. doi: 10.1046/j.1365-2958.2003.03765.x. [DOI] [PubMed] [Google Scholar]
- Rothenbacher FP, Suzuki M, Hurley JM, et al. Clostridium difficile MazF toxin exhibits selective, not global, mRNA cleavage. J Bacteriol. 2012;194:3464–74. doi: 10.1128/JB.00217-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rustad TR, Harrell MI, Liao R, et al. The enduring hypoxic response of Mycobacterium tuberculosis. PLoS One. 2008;3:e1502. doi: 10.1371/journal.pone.0001502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saikia M, Hatzoglou M. The many virtues of tRNA derived stress-induced RNAs (tiRNAs): discovering novel mechanisms of stress response and effect on human health. J Biol Chem. 2015;290:29761–8. doi: 10.1074/jbc.R115.694661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saikia M, Krokowski D, Guan BJ, et al. Genome-wide identification and quantitative analysis of cleaved tRNA fragments induced by cellular stress. J Biol Chem. 2012;287:42708–25. doi: 10.1074/jbc.M112.371799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sala A, Bordes P, Genevaux P. Multiple toxin-antitoxin systems in Mycobacterium tuberculosis. Toxins. 2014;6:1002–20. doi: 10.3390/toxins6031002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schifano JM, Woychik NA. 23S rRNA as an a-Maz-ing new bacterial toxin target. RNA Biol. 2014;11:101–5. doi: 10.4161/rna.27949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schifano JM, Cruz JW, Vvedenskaya IO, et al. tRNA is a new target for cleavage by a MazF toxin. Nucleic Acids Res. 2016 doi: 10.1093/nar/gkv1370. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schifano JM, Edifor R, Sharp JD, et al. Mycobacterial toxin MazF-mt6 inhibits translation through cleavage of 23S rRNA at the ribosomal A site. P Natl Acad Sci USA. 2013;110:8501–6. doi: 10.1073/pnas.1222031110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schifano JM, Vvedenskaya IO, Knoblauch JG, et al. An RNA-seq method for defining endoribonuclease cleavage specificity identifies dual rRNA substrates for toxin MazF-mt3. Nat Commun. 2014;5:3538. doi: 10.1038/ncomms4538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schuessler DL, Cortes T, Fivian-Hughes AS, et al. Induced ectopic expression of HigB toxin in Mycobacterium tuberculosis results in growth inhibition, reduced abundance of a subset of mRNAs and cleavage of tmRNA. Mol Microbiol. 2013;90:195–207. doi: 10.1111/mmi.12358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sebastia J, Kieran D, Breen B, et al. Angiogenin protects motoneurons against hypoxic injury. Cell Death Differ. 2009;16:1238–47. doi: 10.1038/cdd.2009.52. [DOI] [PubMed] [Google Scholar]
- Sharp JD, Cruz JW, Raman S, et al. Growth and translation inhibition through sequence-specific RNA binding by Mycobacterium tuberculosis VapC toxin. J Biol Chem. 2012;287:12835–47. doi: 10.1074/jbc.M112.340109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh R, Barry CE, 3rd, Boshoff HI. The three RelE homologs of Mycobacterium tuberculosis have individual, drug-specific effects on bacterial antibiotic tolerance. J Bacteriol. 2010;192:1279–91. doi: 10.1128/JB.01285-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stewart GR, Wernisch L, Stabler R, et al. Dissection of the heat-shock response in Mycobacterium tuberculosis using mutants and microarrays. Microbiology. 2002;148:3129–38. doi: 10.1099/00221287-148-10-3129. [DOI] [PubMed] [Google Scholar]
- Thompson DM, Lu C, Green PJ, et al. tRNA cleavage is a conserved response to oxidative stress in eukaryotes. RNA. 2008;14:2095–103. doi: 10.1261/rna.1232808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson DM, Parker R. Stressing out over tRNA cleavage. Cell. 2009a;138:215–9. doi: 10.1016/j.cell.2009.07.001. [DOI] [PubMed] [Google Scholar]
- Thompson DM, Parker R. The RNase Rny1p cleaves tRNAs and promotes cell death during oxidative stress in Saccharomycescerevisiae. J Cell Biol. 2009b;185:43–50. doi: 10.1083/jcb.200811119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tiwari P, Arora G, Singh M, et al. MazF ribonucleases promote Mycobacterium tuberculosis drug tolerance and virulence in guinea pigs. Nat Commun. 2015;6:6059. doi: 10.1038/ncomms7059. [DOI] [PubMed] [Google Scholar]
- Tomita K, Ogawa T, Uozumi T, et al. A cytotoxic ribonuclease which specifically cleaves four isoaccepting arginine tRNAs at their anticodon loops. P Natl Acad Sci USA. 2000;97:8278–83. doi: 10.1073/pnas.140213797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Es MA, Schelhaas HJ, van Vught PW, et al. Angiogenin variants in Parkinson disease and amyotrophic lateral sclerosis. Ann Neurol. 2011;70:964–73. doi: 10.1002/ana.22611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vesper O, Amitai S, Belitsky M, et al. Selective translation of leaderless mRNAs by specialized ribosomes generated by MazF in Escherichia coli. Cell. 2011;147:147–57. doi: 10.1016/j.cell.2011.07.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vvedenskaya IO, Sharp JS, Goldman SR, et al. Growth phase-dependent control of transcription start site selection and gene expression by nanoRNAs. Gene Dev. 2012;26:1498–507. doi: 10.1101/gad.192732.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winther KS, Brodersen DE, Brown AK, et al. VapC20 of Mycobacterium tuberculosis cleaves the Sarcin-Ricin loop of 23S rRNA. Nat Commun. 2013;4:2796. doi: 10.1038/ncomms3796. [DOI] [PubMed] [Google Scholar]
- Winther KS, Gerdes K. Enteric virulence associated protein VapC inhibits translation by cleavage of initiator tRNA. P Natl Acad Sci USA. 2011;108:7403–7. doi: 10.1073/pnas.1019587108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamaguchi Y, Nariya H, Park JH, et al. Inhibition of specific gene expressions by protein-mediated mRNA interference. Nat Commun. 2012;3:607. doi: 10.1038/ncomms1621. [DOI] [PubMed] [Google Scholar]
- Yamasaki S, Ivanov P, Hu GF, et al. Angiogenin cleaves tRNA and promotes stress-induced translational repression. J Cell Biol. 2009;185:35–42. doi: 10.1083/jcb.200811106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang M, Gao C, Wang Y, et al. Characterization of the interaction and cross-regulation of three Mycobacterium tuberculosis RelBE modules. PLoS One. 2010;5:e10672. doi: 10.1371/journal.pone.0010672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan J, Sterckx Y, Mitchenall LA, et al. Vibrio cholerae ParE2 poisons DNA gyrase via a mechanism distinct from other gyrase inhibitors. J Biol Chem. 2010;285:40397–408. doi: 10.1074/jbc.M110.138776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang J, Zhang Y, Zhu L, et al. Interference of mRNA function by sequence-specific endoribonuclease PemK. J Biol Chem. 2004;279:20678–84. doi: 10.1074/jbc.M314284200. [DOI] [PubMed] [Google Scholar]
- Zhang Y, Zhang J, Hara H, et al. Insights into the mRNA cleavage mechanism by MazF, an mRNA interferase. J Biol Chem. 2005;280:3143–50. doi: 10.1074/jbc.M411811200. [DOI] [PubMed] [Google Scholar]
- Zhang Y, Zhang J, Hoeflich KP, et al. MazF cleaves cellular mRNAs specifically at ACA to block protein synthesis in Escherichia coli. Mol Cell. 2003;12:913–23. doi: 10.1016/s1097-2765(03)00402-7. [DOI] [PubMed] [Google Scholar]
- Zhu L, Inoue K, Yoshizumi S, et al. Staphylococcus aureus MazF specifically cleaves a pentad sequence, UACAU, which is unusually abundant in the mRNA for pathogenic adhesive factor SraP. J Bacteriol. 2009;191:3248–55. doi: 10.1128/JB.01815-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu L, Phadtare S, Nariya H, et al. The mRNA interferases, MazF-mt3 and MazF-mt7 from Mycobacterium tuberculosis target unique pentad sequences in single-stranded RNA. Mol Microbiol. 2008;69:559–69. doi: 10.1111/j.1365-2958.2008.06284.x. [DOI] [PMC free article] [PubMed] [Google Scholar]