

Abstract
Objective: To construct a recombinant adenovirus vector-carrying human growth and differentiation factor-5 (GDF-5) gene, investigate the biological effects of adenovirus-mediated GDF-5 (Ad-GDF-5) on extracellular matrix (ECM) expression in human degenerative disc nucleus pulposus (NP) cells, and explore a candidate gene therapy method for intervertebral disc degeneration (IDD). Methods: Human NP cells of a degenerative disc were isolated, cultured, and infected with Ad-GDF-5 using the AdEasy-1 adenovirus vector system. On Days 3, 7, 14, and 21, the contents of the sulfated glycosaminoglycan (sGAG), deoxyribonucleic acid (DNA) and hydroxyproline (Hyp), synthesis of proteoglycan and collagen II, gene expression of collagen II and aggrecan, and NP cell proliferation were assessed. Results: The adenovirus was an effective vehicle for gene delivery with prolonged expression of GDF-5. Biochemical analysis revealed increased sGAG and Hyp contents in human NP cells infected by Ad-GDF-5 whereas there was no conspicuous change in basal medium (BM) or Ad-green fluorescent protein (GFP) groups. Only cells in the Ad-GDF-5 group promoted the production of ECM, as demonstrated by the secretion of proteoglycan and up-regulation of collagen II and aggrecan at both protein and mRNA levels. The NP cell proliferation was significantly promoted. Conclusions: The data suggest that Ad-GDF-5 gene therapy is a potential treatment for IDD, which restores the functions of degenerative intervertebral disc through enhancing the ECM production of human NP cells.
Keywords: Intervertebral disc, Degeneration, Growth and differentiation factor-5 (GDF-5), Adenovirus, Gene therapy, Nucleus pulposus
1. Introduction
The intervertebral disc (IVD) functions as a shock absorber for the body by providing a weight-bearing structure. The IVD maintains the mechanical stability and multidirectional flexibility of the spine through its unique composition of a collagen-rich annulus fibrosus (AF) and a proteoglycan-rich nucleus pulposus (NP) (Melrose et al. 2012). Degeneration of discs induced by biochemical and biomechanical changes traditionally attributed to age trauma >heredity morbid obesity mechanical loading and other factors impairing disc nutrition leads to low back pain (LBP). This disease is a common public socio-economic and health-care problem in western societies (Eskola et al. 2012; Phillips et al. 2013).
The cellular changes associated with disc degeneration consist of alterations in the equilibria of the extracellular matrix (ECM) increased synthesis of collagen I decreased collagen II and proteoglycan (Freemont 2009).The NP tissue as the central component of IVD includes abundant collagen II and proteoglycan. NP loss through cell aging and apoptosis leads to lower moisture and higher swelling pressure to force apart the vertebral bodies and finally leads to LBP (Zhang et al. 2009). In this disease process zinc-based matrix-degrading enzymes such as matrix metalloproteases (MMPs) and the protease family with a disintegrin and metalloproteinase domain with thrombospondin motifs (ADAMTS) play a pivotal role in the up-regulation catabolism of ECM which causes the decrease of collagen II and aggrecan via dependent complex molecular signaling pathways (Kim et al. 2012; Hua et al. 2013).
Traditional treatment methods for degenerated IVD vary but include the conservative involving medications physical therapy and surgery including discectomy and interbody fusion (Smith et al. 2011; Eskola et al. 2012). However conservative treatment and invasive interventions are aimed at alleviating painful symptoms without preserving disc mechanics structure and involving restoration of cellular biological function. Patients undergoing discectomy or interbody fusion lose the natural multi-directionality of the spine which will lead to recurrent pain and degeneration of adjacent levels of the spine potentially requiring additional surgery (Hanley et al. 2010; Smith et al. 2011).
Growth factor gene therapy cell implantation and tissue engineering strategy all have been investigated as promising treatment strategies. In particular growth factor or gene therapy is considered to be a potential method for inhibiting or reversing early intervertebral disc degeneration (IDD) (Stoyanov et al. 2011). Anabolic effects of growth factors on the ECM metabolism of IVD cells have been shown (Eskola et al. 2012). Transforming growth factor-β (TGF-β) bone morphogenetic proteins (BMPs) insulin-like growth factor-1 (IGF-1) basic fibroblast growth factor (bFGF) platelet-derived growth factor (PDGF) growth and differentiation factor-5 (GDF-5) and others have attracted the most attention as candidates for regenerating the matrix of the IVD restoring disc height and reversing IDD disease (Ellman et al. 2008; Masuda 2008). However several studies in vitro and animal studies in vivo have suggested that the biological half-life of cytokine is very short with a transitory biological therapeutic effect (Chubinskaya et al. 2007; Elkasrawy and Hamrick 2010). Gene therapy is used as an effective treatment method due to its ability to offer long-lasting production of the target protein to stimulate the regeneration of matrix components in disc degeneration.
GDF-5 a member of the BMP family is also called cartilage-derived morphogenetic protein-1 (CDMP-1) and BMP-14. GDF-5 could promote proteoglycan synthesis in chondrocyte-like cells. GDF-5 not only plays a crucial role in musculoskeletal processes (Daans et al. 2011) but also could work on other diseases such as tendon and ligament injury (Saiga et al. 2010; Hogan et al. 2011) Parkinson’s disease (PD) (Costello et al. 2012) myocardial infarction (Zaidi et al. 2010) and periodontal disease (Moore et al. 2010). Previous studies suggest that a single-nucleotide polymorphism (SNP) variant rs143383 in GDF-5 is associated with IDD (Williams et al. 2011; Eskola et al. 2012). In addition the study of the levels of collagen II and aggrecan mRNA in GDF-5-deficient mice and molecular changes when treating disc cells with recombinant GDF-5 has identified that GDF-5 deficiency leads to tissue-level abnormalities of the IVD (Li et al. 2004). Recombinant GDF-5 could result in a dose-dependent up-regulation of the collagen II and aggrecan genes which are the important markers of disc health (Li et al. 2004). In our previous work adenovirus-mediated GDF-5 (Ad-GDF-5) was injected into the mice disc degeneration model induced by annulus needle puncture. The results not only confirmed the long-term expression of the target protein in the disc in vivo but also highlighted the physiological improvements that occurred to the disc. Ad-GDF-5 gene therapy could restore the functions of injured discs and has the potential to be an effective treatment (Liang et al. 2010). In other research GDF-5 was transfected with mesenchymal stem cells (MSCs) by electroporation to prepare the seed cells for IVD cell therapy (Bucher et al. 2013). The results showed that the MSCs expressed GDF-5 efficiently for up to 21 d and GDF-5 gene transfer was accompanied by an up-regulation of aggrecan and SOX9 two markers for chondrogenesis.
In this study the Ad-GDF-5 was used to transfect human NP cells to determine whether Ad-GDF-5 could promote ECM metabolism in human NP cells derived from degenerate IVD and try to explore a candidate method of gene therapy for IDD.
2. Materials and methods
2.1. Preparation of recombinant adenovirus
Ad-GDF-5 was generated using the AdEasy-1 adenoviral vector system (Stratagene La Jolla CA USA) according to the manufacturer’s instructions. Briefly human GDF-5 gene obtained by polymerase chain reaction (PCR) was inserted into plasmid pMD19-T and the 1.7 kb GDF-5 cDNA sequence was cloned into the adenoviral shuttle plasmid pShuttle-cytomegalovirus (CMV) of the AdEasy-1 system. The adenoviral GDF-5 vector was produced by using the resultant pShuttle-CMV-GDF-5 to homologous recombination with the adenoviral backbone plasmid pAdEasy-1 in BJ5183 bacterial cells. It was selected by kanamycin and identified by a digestion with HindIII restriction enzyme and amplified in XL10-Gold competent bacteria. Then the DNA of the recombinant adenovirus vector was linearized with PacI and using the adenoviral recombinants generated adenoviruses in HEK293 packaging cells to obtain an Ad-GDF-5. In addition virus stocks were propagated in HEK293 cells purified by the method of cesium chloride ultracentrifugation at 32 000 r/min for 20 h at 4 °C and further titered using agarose-overlayed plaque assays.
2.2. Weatern blotting of GDF-5
Lysates from cells infected or not with Ad-GDF-5 or Ad-green fluorescent protein (GFP) were split by a sample loading buffer (β-mercaptoethanol, Bio-Rad; D L-dithiothreitol, Hercules, USA), and denatured at 100 °C for 5 min. Protein samples were then concentrated by electrophoresis on 5% (50 mg/ml) criterion stacking sodium dodecyl sulphate (SDS)-polyacrylamide gel at 40 V for 20 min and separated on 10% (100 mg/ml) separation gel at 120 V for 70 min under reducing conditions on a mini-protean 3 system apparatus (Bio-Rad, USA). Protein was transferred to pre-wetted polyvinylidene fluoride (PVDF) membrane with semi-dry (Bio-Rad) conditions (15 V for 15 min). The immunoblotting membrane was blocked by incubation with blocking buffer-TBST (Tris-buffered saline (TBS), pH 7.5 with 0.05% Tween-20) containing 5% (50 mg/ml) nonfat dry milk for 2 h at room temperature, then incubated overnight at 4 °C with a primary goat anti-GDF-5 polyclonal antibody diluted 1:200 (v/v) in blocking buffer (5% (50 mg/ml) nonfat dry milk, 3% (30 mg/ml) bovine serum albumin (BSA) in TBST solution). After successive washing with TBS and TBST solution, the membranes were blocked in 1% (10 mg/ml) secondary mouse anti-goat IgG (Sigma) for 2 h at 37 °C, then washed with TBST, and detected using the enhanced chemiluminescence (ECL; Bio-Rad).
2.3. Isolation, culture and identification of NP cells
Human degenerative IVD tissues were obtained from patients (50–65 years old) undergoing discectomy (n=10) after they had provided informed consent (all the IDD patients are in Stage 3 according to Pfirrmann classification theory (Pfirrmann et al., 2001)).
NP tissues were then separated and collected. After sequential enzymatic digestion of collagen II and trypsin, cells from the NP tissues were isolated and washed twice with complete medium (Dulbecco’s modified Eagle’s medium, and Ham’s F-12 medium without NaHCO3 (DMEM/F-12); Gibco, New York, USA) containing 100 U/ml penicillin and streptomycin (Gibco, New York, USA). The NP cells were placed into the sterile culture flask and incubated at 37 °C in 5% CO2 and 5% O2 (Eppendorf, 48R, Germany). The medium contained high glucose DMEM/F-12 supplemented with 20% fetal bovine serum (FBS; Gibco, New York, USA) and 50 U/ml penicillin and streptomycin, and was changed every 2 d.
When the NP cells confused about 80%–90%, the cells were digested by 0.25% (2.5 mg/ml) trypsin, and then the suspension was adjusted about 1×105 ml−1. The four kinds of special NP cells antibody were detected by flow cytometry (FC 500, Beckman, USA) respectively. The antibodies included KRT18, FoxF1, Tie2, and CD24. Each antibody was tested three times. The cell suspension (1 ml) was used for identification each time. The fluorescent monoclonal antibodies were mixed to the cell suspension. The four kinds of antibody were mouse monoclonal cytokeratin 18 (Fitzgerald, USA), monoclonal anti-FoxF1 antibody produced in the mice (Sigma, USA), recombinant Fab monoclonal Tie2 antibody (Fitzgerald, USA), and monoclonal anti-CD24 antibody produced in the mouse (Sigma, USA). After the mixing solution had been incubated in the dark at room temperature for 30 min, the mixing solution was washed by 2 ml phosphate-buffered saline (PBS), and the supernatant was removed. Then 500 μl PBS (including 10 g/L paraformaldehyde) was added, and the cell suspension was used for flow cytometry detection. According to the fluorescence intensity of the same type of antibody, the negative cells group was set. The positive cells’ expression rate was observed.
2.4. Infection of NP cells with Ad-GDF-5 and determination of the optimal multiplicity of infection (MOI)
Passage 3 human NP cells were cultured in 24-well plates at a density of 5×104 per well. Different MOI Ad-GDF-5 virus particle solutions were premixed with 1 ml DMEM/F-12 containing 2% FBS and 1% penicillin/streptomycin and added to the cell culture wells. The medium was then changed every 3 d. The MOIs were 25, 50, 100, 150, and 200, respectively. For determination of the optimal MOI, the cells were digested in 200 μl of sterile papain digestion buffer (125 g/ml in 1×phosphate buffered ethylenediaminetetraacetic acid (EDTA) solution (PBE), pH 6.5) at 60 °C for 24 h. The sulfated glycosaminoglycan (sGAG) concentration was determined by the modified dimethylmethylene blue (DMMB; Sigma, Saint Louis, MO, USA) colorimetric assay (16 mg of DMMB, 3.04 g glycine, NaCl 2.37 g, 0.1 mol/L HCl 95 ml; volume adjusted to 1 L with distilled water and the pH adjusted to 3.0 with formic acid) with chondroitin sulfate as the standard (Farndale et al., 1986). Total DNA content of NP cells was determined using the bisbenzimide Hoechst 33258 DNA quantitation kit (Sigma, Saint Louis, MO, USA) fluorometric dye assay with calf thymus DNA as a standard (Kim et al., 1988).
2.5. Biochemical analyses
Human NP cells were treated in Passage 3 cell cultures and evaluated at four time points. In summary, 5×104 cells were cultured overnight in the medium containing the optimal virus titer dissolution in DMEM/F-12 plus 2% FBS in 24-well plates. The medium was then replaced with basal medium (DMEM/F-12 containing 2% FBS). Cells were treated with Ad-GDF-5 or Ad-GFP (as Ad-GDF-5/GFP group) and cultured in basal medium, which served as a control group (BM group). The contents of sGAG and DNA of NP cells were measured after 3, 7, 14, and 21 d in culture as described previously. The hydroxyproline (Hyp) content was determined after hydrolysis of the cells in 6 mol/L HCl for 24 h at 110 °C using the classical acid hydrolyzation (Hydroxyproline Assay Kit, Sigma, USA). The total DNA content of NP cells was also determined by Hoechst 33258 as described above. For each assay, all cell groups and standards were processed in duplicate as part of a single experiment and repeated a minimum of three times to confirm the results. The following ratios were calculated for each group: sGAG/DNA and Hyp/DNA ratios.
2.6. Histology analysis
For histological analysis, the cell cultures were harvested, washed with PBS to remove the medium, and fixed in 4% paraformaldehyde for 10 min. The cells were stained with hematoxylin and eosin staining. Sequential staining with fast green and Safranin-O binds with sulfated proteoglycans and produces a deep red stain to areas rich in proteoglycan.
2.7. Immunohistochemical staining
Immunochemical staining for collagen II was performed using a collagen staining kit (Chondrex, Redmond, WA, USA) according to the manufacturer’s instructions. For immunohistochemical staining of intracellular proteins, the NP cells cultured on sterile glass cover slips were fixed with glutaraldehyde in PBS for 10 min at 4 °C. Cells were then treated with 20 mg/ml bovine testicular hyaluronidase dissolved in PBS, pH 7.4 for 30 min at 25 °C, then incubated with blocking buffer for 30 min followed by arthrogen-CIA collagen II monoclonal antibodies at a dilution of 1:250 (v/v) overnight at 4 °C. Streptavidin peroxidase (50 ml in 10 ml streptavidin peroxidase-dilution buffer) was applied for 1 h at room temperature. The cells were then developed with a brown specific stain by 3,3'-diaminobenzidine (DAB) substrate solution for 30 min. For visualization and aggrecan assay, the cells were treated as above, blocked with goat IgG (1:250, v/v) for 2 h at room temperature and then incubated with rabbit anti-mouse aggrecan polyclonal antibody (5 mg/ml; Santa Cruz Biotechnology, Santa Cruz, CA, USA) for 12–18 h at 4 °C, followed by incubation with fluorescein isothiocyanate (FITC; for aggrecan) conjugated secondary antibody for 1 h at room temperature in a moist dark box.
2.8. Enzyme-linked immunosorbent assay (ELISA)
Cells were digested by 5 mmol/L cyclopamine at 21th day after infection and then diluted to 106 ml−1 with PBS (pH 7.2–7.4). The cell suspension was lysed by tissue protein extraction reagent to obtain the protein. The supernate was collected by centrifuging at 2500 r/min for 20 min. The ELISA test for collagen II was performed using a human collagen type II ELISA kit (Cusabio, USA) according to the manufacturer’s instructions and the ELISA test for aggrecan was performed by human aggrecan core protein (AGC1) ELISA kit (Cusabio, USA). The standard curve was drawn to calculate the corresponding protein concentration in different groups. Every group was transferred to three wells and each experiment was repeated three times.
2.9. Analysis of mRNA expression levels
Total RNA from the cells was extracted by the RNeasy Mini kit (Qiagen, Valencia, CA, USA). The RNA was reverse-transcribed to cDNA using the first strand cDNA kit (Promega, Madison, WI, USA) according to the manufacturer’s protocol. Real-time PCR analysis was conducted using the QuantiTect SYBR Green PCR master mix (Qiagen, Valencia, CA, USA). Standard curves were plotted and each transcript was quantified by normalizing to human β-actin as an internal control. Table 1 shows the amplification primers.
Table 1.
Primer sequences used for RT-PCR
| Molecule | Primer sequence | Product size (bp) |
| Collagen II | 5'-TCCCAGAACATCACCTACC-3' (sense) | 131 |
| 5'-AACCTGCTATTGCCCTCT-3' (antisense) | ||
| Aggrecan | 5'-CGCTACTCGCTGACCTTT-3' (sense) | 106 |
| 5'-GCTCATAGCCTGCTTCGT-3' (antisense) | ||
| β-Actin | 5'-GAGCTACGAGCTGCCTGACG-3' (sense) | 128 |
| 5'-GTAGTTTCGTGGATGCCACAG-3' (antisense) |
2.10. NP cell proliferation after transfection
For NP cell proliferation test, the cells were cultured as described above in method Section 2.5. The cell proliferation was measured after 3, 7, 14, and 21 d by CCK-8 kit (Cell Counting Kit-8, Dojindo, Japan). The test solution was prepared by diluting the stock solution at 1:10 (volume ratio) with the cell culture medium. The cell medium was then discarded and the cells were washed with PBS (pH 7.2–7.4) three times. The test solution (250 μl) was added to every well and incubated for 2 h, and then 100 μl test solution was transferred to 96-well plates for measuring at 450 nm (Cytation 3, Biotek, USA).
2.11. Statistical analysis
All experiments in vitro were performed in triplicate and statistical evaluation was performed using analysis of variance followed by a post hoc Student’s t-test. A P-value of less than 0.05 was considered significant. All data are expressed as the mean±standard deviation (SD).
3. Results
3.1. Infection with Ad-GDF-5 induces expression of GDF-5
To verify the ability of Ad-GDF-5 to induce expression of GDF-5, HEK293 cells were infected with Ad-GDF-5 adenovirus. Cell culture supernatant was collected 2 d post-transduction, and Western blotting was performed with goat anti-GDF-5 polyclonal antibody. Two bands migrating at approximately 15 and 55 kDa were clearly observed in PVDF membrane blotting by mouse anti-goat IgG (Fig. 1a). The GDF-5 gene codes for 501-amino acid (aa) precursor protein that is cleaved to a 382-aa mature GDF-5 monomeric protein (long: 120 aa (382–501); molecular mass: 13.6 kDa) by subtilisin-like proteases. GDF-5 as a homodimer has two monomers linked by a disulfide bridge under physiological conditions, while mature GDF-5 exhibited a molecular mass of about 15 kDa under semidry conditions. Moreover, the band at 55 kDa probably denotes the full-length precursor protein. As shown in Fig. 1a, the Western blotting results of the BM group and Ad-GFP group were negative. There were no obvious protein bands at 55 and 15 kDa. These data demonstrate that infection with Ad-GDF-5 provides expression of mature GDF-5 protein, which is consistent with previous reports (Feng et al., 2008).
Fig. 1.

Western blotting results
Proteins were electrophoretically separated on 10% criterion separation SDS-polyacrylamide gel on a mini-protean 3 system apparatus. Western blotting was performed with goat anti-GDF-5 polyclonal antibody. (a) Western blotting of GDF-5 recovered from the supernatant of cultured HEK293 cells at the second day post-infection; (b) Western blotting of GDF-5 recovered from the supernatant of NP cells at 21 d post-infection. Lane 1: control group; Lane 2: Ad-GFP group; Lane 3: Ad-GDF-5 group
In order to verify the GDF-5 expression in NP cells, the protein was lysated from NP cells at 21th day after transfected for Western blotting. The results show that there were obvious bands at 15 kDa in the Ad-GDF-5 group. It is notable that there were still protein bands at 55 kDa in the BM group and Ad-GFP group because of GDF-5 self-secretion in NP cells. However, the GDF-5 protein expression amount was significantly higher in the Ad-GDF-5 group than in the other two groups from the bands at 15 kDa (Fig. 1b). These data demonstrate that GDF-5 expression in NP cells has been promoted by Ad-GDF-5.
3.2. Optimal MOI of Ad-GDF-5 for transduction of NP cells
All the NP cells were identified through flow cytometry. Because the NP cells were harvested from the IDD patients, the positive expression rate was less than that for normal NP cells, but the positive rates were all more than 50% (Fig. 2). Since the transduction efficiency depended on the virus dose, NP cells were infected with increasing amounts of the Ad-GDF-5. The NP cells were then harvested at 3, 7, and 14 d after transduction by Ad-GDF-5 (MOIs of 25, 50, 100, 150, and 200).
Fig. 2.

Flow cytometry identification result of NP cells
As shown in Fig. 3, the contents of sGAG generated by all the groups increased with time, and the production of sGAG peaked 14 d after the initial infection. It should be noted that although the sGAG contents increased with time at 3 and 7 d, there was no significant difference among all groups at 3 and 7 d (P>0.05). During the 14 d post-transduction, the concentration of sGAG increased with time and the viral MOI. Cells infected with Ad-GDF-5 at MOIs 150 and 200 gave sGAG per cell (DNA) level at 196.759 and 193.675 after 14 d, respectively. The difference was not significant at an MOI of 200 compared to 150 on Day 14. However, there was a significant difference between MOIs 150, 200, and other groups (P<0.05). Accordingly, the transduction efficiency was optimal at an MOI of 150 at 14 d and the results were used for all the subsequent experiments.
Fig. 3.
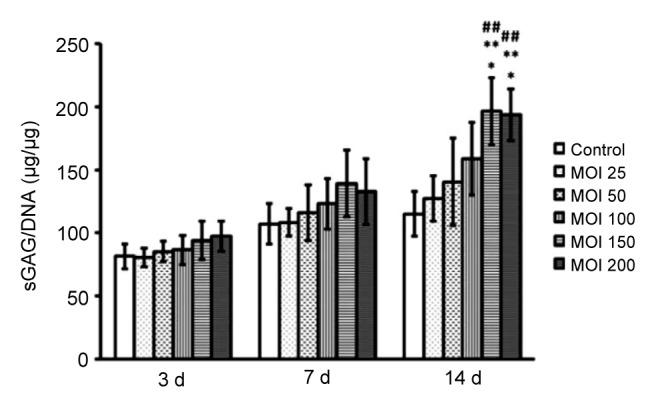
sGAG/DNA ratio for human NP cells treated with Ad-GDF-5
The sGAG and DNA expression levels were measured in NP cells that were infected with increasing multiplicity of infection (MOI) ratios of Ad-GDF-5. Human NP cells were collected at 3, 7, and 14 d after the primal transduction, and the sGAG and DNA levels were detected using the DMMB colorimetric assay and the Hoechst 33258 DNA quantitation kit, respectively. Data points are presented as mean±SD (n=3). * P<0.05 vs. MOI 50; ** P<0.01 vs. control; ## P<0.01 vs. MOI 25
3.3. Ad-GDF-5 promotes the synthesis of sGAG and Hyp
Human NP cells treated with Ad-GDF-5 were cultured in 24-well plates to determine whether gene transfer could be used as an effective means for increasing the synthesis of ECM; infection with Ad-GFP and non-infection were used as control groups.
To verify the promotive effect of Ad-GDF-5 for NP cells, the sGAG was measured to estimate the total proteoglycan level within the cells, and Hyp was used to estimate the total collagen content. Both sGAG and Hyp were normalized to DNA content as described previously. The sGAG secreted by cells cultured in the basal medium was the highest after Day 3, and did not demonstrate a statistically significant increase with time. The level of sGAG in the Ad-GDF-5 group was more than that in the BM or Ad-GFP controls after 7 d, and the level of sGAG gradually increased during each sampling period. As shown in Fig. 4a, the Ad-GDF-5 group exhibited a significantly higher sGAG content than the BM and Ad-GFP groups 14 d post-infection (P<0.05) and particularly at 21 d (P<0.01), whereas the Ad-GFP group did not show a greater effect than BM group (P>0.05), as did total intracellular collagen (Fig. 4b).
Fig. 4.
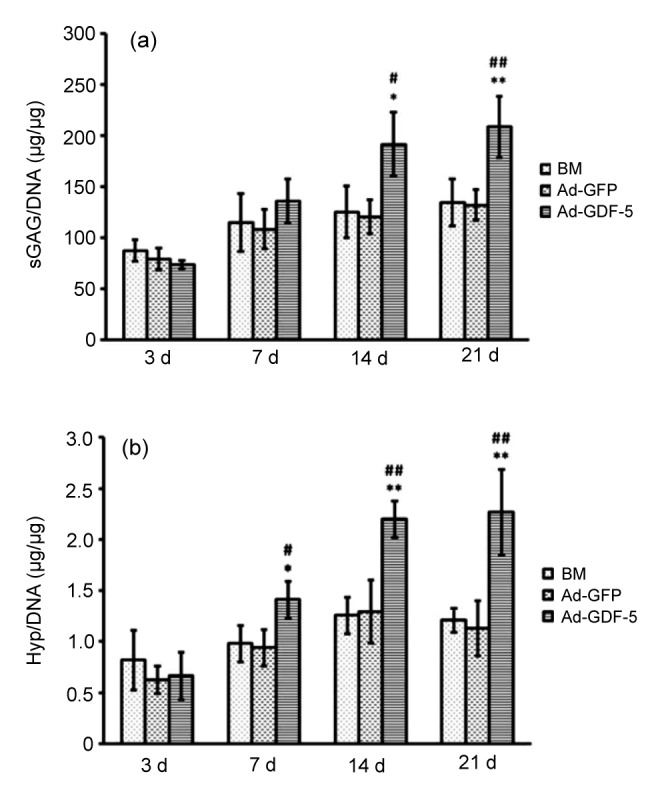
Levels of sulfated glycosaminoglycan (sGAG) and hydroxyproline (Hyp) normalized to DNA content in NP cells
(a) The sGAG/DNA ratio in NP cells infected by Ad-GDF-5, Ad-GFP, or no adenovirus (BM group); (b) The Hyp/DNA ratio in NP cells infected with Ad-GDF-5, Ad-GFP, or no adenovirus (BM group). Data points are presented as mean±SD (n=3). * P<0.05, ** P<0.01, vs. BM group; # P<0.05, ## P<0.01, vs. Ad-GFP group
3.4. Ad-GDF-5 augments NP cells ECM production
Safranin-O allowed us to visually estimate the proteoglycan-rich ECM deposition. Safranin-O staining creates a red/orange appearance in all cell culture lines, allowing histological examination for the presence of proteoglycans in NP cells. Only cells in the Ad-GDF-5 group showed an increase in the deposition of proteoglycans compared with the other two groups of BM or Ad-GFP (Fig. 5).
Fig. 5.
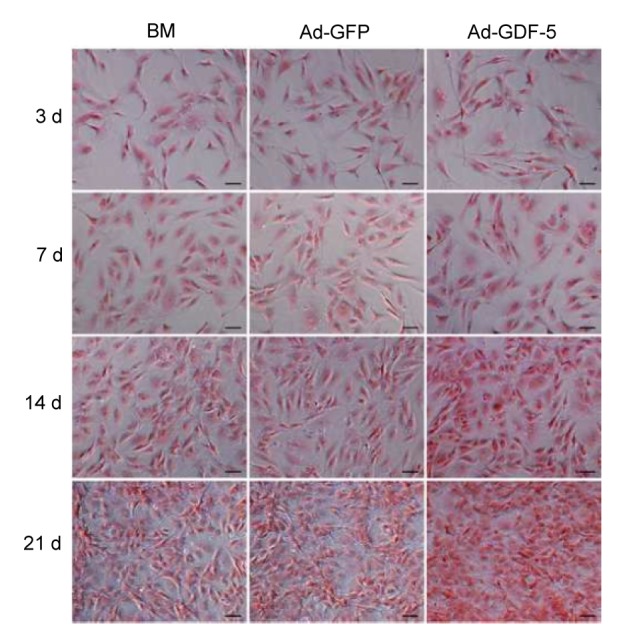
Safranin-O staining (red) of the proteoglycan-rich ECM of human
NP cells NP cells were stained at 3, 7, 14, and 21 d after being infected by Ad-GDF-5, Ad-GFP, or no adenovirus (BM group). Scale bar=50 μm (Note: for interpretation of the references to color in this figure legend, the reader is referred to the web version of this article)
NP cells were harvested and immunostained for collagen II (immunochemical staining) and aggrecan (immunofluorescent staining). NP cells treated with Ad-GDF-5 demonstrated robust staining for collagen II, whereas the cells infected by Ad-GFP or non-infected showed weaker staining (Fig. 6a). Typically, the immunofluorescent staining intensity of aggrecan was higher toward the cytoplasm and lower in the nucleus regions of all cells. Ad-GDF-5 cells exhibited a deeper staining than BM or Ad-GFP cells as shown in Fig. 6b. This, along with the biochemical data and Safranin-O staining, clearly demonstrates that NP cells enhanced the ECM production in the presence of Ad-GDF-5.
Fig. 6.
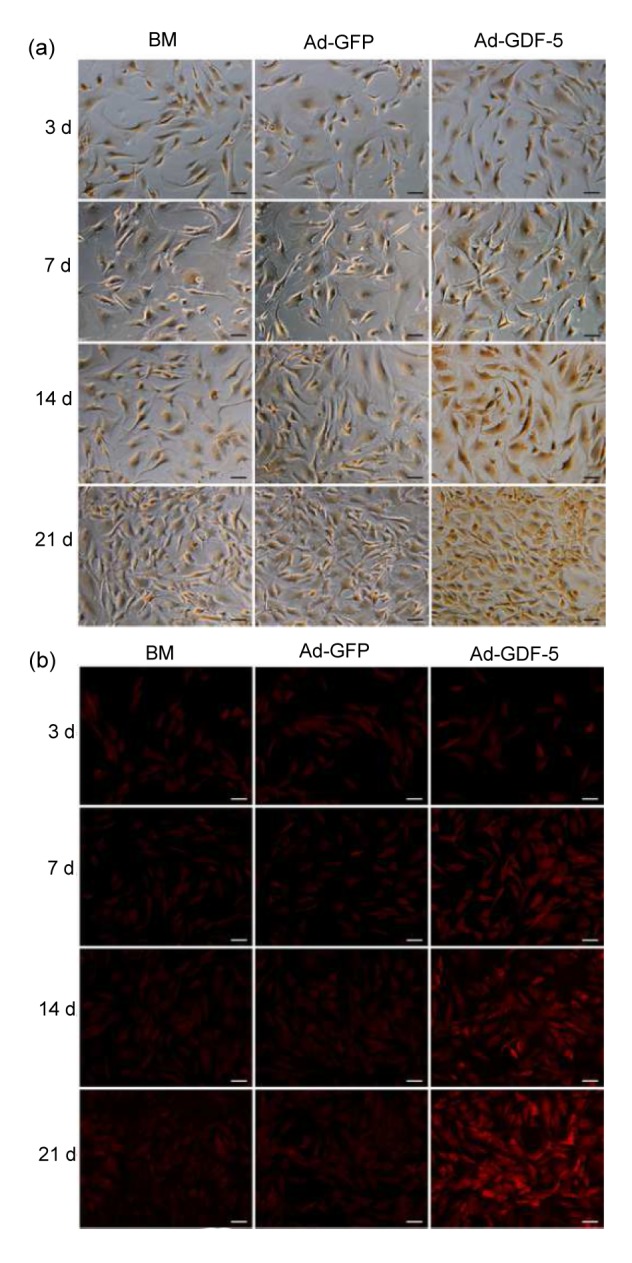
Immunostaining of human NP cells for type-II collagen and aggrecan after they were infected by Ad-GDF-5, Ad-GFP, or no adenovirus (BM group)
Type-II collagen is stained dark brown (a), and aggrecan is shown by red fluorescent staining (b). Scale bar=50 μm (Note: for interpretation of the references to color in this figure legend, the reader is referred to the web version of this article)
To further prove the ECM production in the three group, the collagen II and aggrecan expressions in NP cells at 21st day were measured by ELISA kit. The results showed that the two specific ECM expressions in the Ad-GDF-5 group were significantly higher than those in the other two groups (P<0.05; Fig. 7). This result indicated that Ad-GDF-5 is effective in promoting NP cell ECM secretion.
Fig. 7.
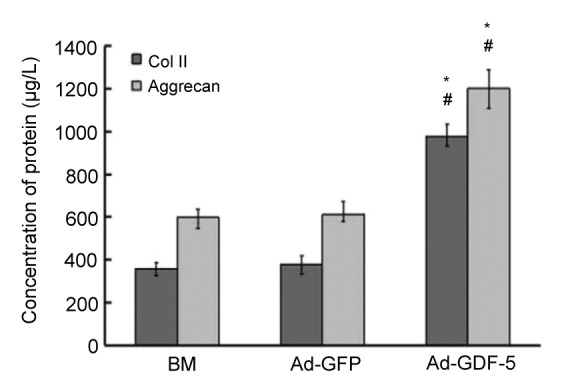
Concentrations of collagen II (Col II) and aggrecan in NP cells at 21th days post-infection by ELISA
Data points are presented as mean±SD (n=3). * P<0.05 vs. BM group; # P<0.05 vs. Ad-GFP group
3.5. Ad-GDF-5 increases expression of collagen II and aggrecan genes
Levels of RNA transcripts for collagen II and aggrecan expression patterns were determined in human NP cells infected by Ad-GDF-5 or Ad-GFP using real-time PCR performed with the SYBR Green PCR kit. In the three groups, mRNA levels of collagen II and aggrecan gradually increased with time (Fig. 8). Compared with BM or Ad-GFP groups, Ad-GDF-5 treatments strongly induced the expression of collagen II and aggrecan. On Days 3 and 7, no significant effect on gene expression was observed among the three cell groups. At the 14th day, the Ad-GDF-5-treated group had significantly up-regulated collagen II and aggrecan expression levels compared with the other groups (P<0.05). By Day 21, all Ad-GDF-5-infected groups demonstrated a significantly greater elevation in collagen II and aggrecan expression levels compared with the other groups (P<0.01). These data indicate that GDF-5 gene transfer mediated adenovirus potently enhances the gene expression levels of collagen II and aggrecan.
Fig. 8.
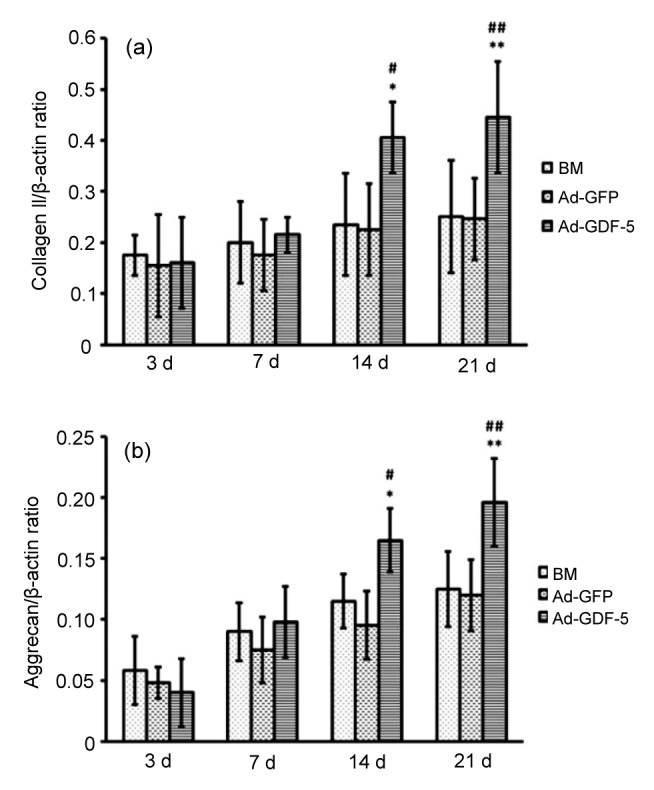
Gene expression patterns in human NP cells following infection by Ad-GDF-5, Ad-GFP, or no adenovirus (BM group) over 3, 7, 14, and 21 d
(a) Collagen II mRNA levels; (b) Aggrecan mRNA levels. Data points are presented as mean±SD (n=6). * P<0.05, ** P<0.01, vs. BM group; # P<0.05, ## P<0.01, vs. Ad-GFP group
3.6. Ad-GDF-5 promotes NP cell proliferation
To verify the Ad-GDF-5 effect on NP cell proliferation, the cells’ survival rate was measured by the CCK-8 method. The higher optical density (OD) value at 450 nm indicated a higher cell survival rate. As shown in Fig. 9, the OD value in the GDF-5 group was the lowest among all groups at 3 d. However, the OD value increased with time and the value was significantly higher than those in the other groups at 14 and 21 d (P<0.01), which suggested that the NP cell proliferation was notably better than that in the other two groups. Compared with the GDF-5 group, although the cell proliferation increased in the GFP group with time, the OD value was still lower than that in the BM group at all time points (P>0.05). Based on these data, it is clear that the NP cell proliferation had been significantly promoted by Ad-GDF-5.
Fig. 9.
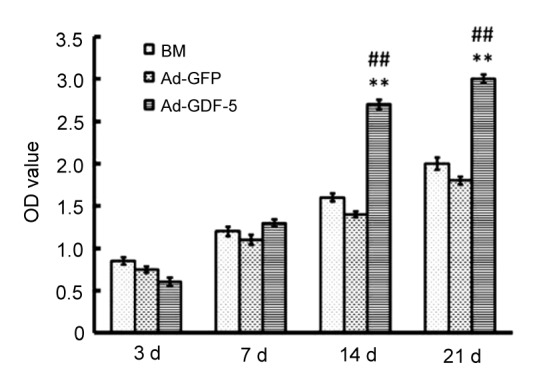
NP cell proliferation after transfection
Data points are presented as mean±SD (n=3). ** P<0.01 vs. BM group; ## P<0.01 vs. Ad-GFP group
4. Discussion
In the present study, the efficacy of Ad-GDF-5 gene therapy on the phenotype of human NP cells derived from degenerate discs in vitro has been investigated. Our results support the following conclusions: (1) Human GDF-5 gene was successfully cloned into an expression vector, and nucleofection of the clone into human NP cells increased gene expression for long-lasting production of the target protein; (2) Infection with Ad-GDF-5 could promote the NP cell ECM accumulation, which was demonstrated by an augmentation of sGAG/DNA and Hyp/DNA ratios, increased synthesis of proteoglycan, and up-regulation of the expression of collagen II and the aggrecan genes.
Sustained expression of exogenous proteins in chronic diseases such as IDD is essential to the success of any treatment approach (Larson III et al., 2006). The previous studies have demonstrated extended expression time of marker transgenes mediated by an adenoviral vector in a rabbit lumbar disc model (Nishida et al., 1998; 1999; Shimer et al., 2004; Sobajima et al., 2004; Larson III et al., 2006; Kaneyama et al., 2008). Therefore, IVD is an appropriate site for adenovirus-mediated transfer of exogenous genes thereby resulting in production of therapeutic growth factors. The IVD is encapsulated, avascular, and “immune privileged”. Owing to this property, no histological changes or cellular immune response occurred when the gene transfect is mediated by adenovinus (Nishida et al., 1998; Shimer et al., 2004). This feature permits a sustained endogenous production of desired gene products in the IVD without immune system interruption.
The GDF-5 gene manipulation for the treatment of IDD is attractive for the following reasons. GDF-5 is important for the development of cartilage and plays an integral role in maintaining the healthy skeletal system. Like cartilage tissue, the NP tissue has similar histological and biological characteristics. Moreover, a known gene mutation in GDF-5 is associated with IDD in Northern European women, as previously delineated (Williams et al., 2011; Eskola et al., 2012). Additionally, previous experiments suggest that GDF-5 can successfully induce collagen II and aggrecan gene expression in the IVD cells of GDF-5-deficient mice cultured in alginate beads (Li et al., 2004).
GDF-5 protein or gene mediated by adenovirus can increase the main ECM, such as collagen and proteoglycan, to enhance the gene expression of collagen II and aggrecan (Wang et al., 2004; Cui et al., 2008). A single injection of Ad-GDF-5 into a degenerated mice disc has shown that the results not only confirmed the long-term expression of the target protein in the disc in vivo but also highlighted the physiological improvements that occurred to the disc (Williams et al., 2011). These observations have been confirmed by other research groups (Walsh et al., 2004; Feng et al., 2009).
In the present study, the recombinant adenovirus vector carrying human GDF-5 gene was successfully constructed using the AdEasy-1 adenovirus vector system. We observed that human NP cells can be engineered by the adenovirus vector to express this growth factor for a prolonged period in vitro. After infecting NP cells with Ad-GDF-5, we measured the synthesis of ECM by analyzing the production of sGAG and Hyp. These measurements were then used to estimate the total proteoglycan level and collagen content respectively. After biochemical analyses, we found no statistical differences at any time in the sGAG/DNA ratio or in the Hyp/DNA ratio between the BM group and AD-GFP group. Between the 14th and 21st days, the NP cells infected with Ad-GDF-5 show a significantly greater sGAG/DNA ratio or Hyp/DNA ratio than those in the BM or Ad-GFP groups. Our data suggest that Ad-GDF-5 promotes ECM accumulation in NP cells via producing the target protein encoded by the GDF-5 gene. Therefore, ex vivo gene transfer of growth factors, particularly GDF-5, may be an attractive approach for achieving sustained growth factor delivery in people with IDD disease.
It has previously been reported that incubated chondrocytes or disc cells with GDF-5 can augment the production of proteoglycans and collagen (Li et al., 2004; Eskola et al., 2012). Consistent with this finding, results from our Safranin-O staining and immunostaining experiments showed deep and robust staining of NP cells infected with Ad-GDF-5. The Western blotting and ELISA results also proved this view at the protein level. Our results indicated that NP cells infected with Ad-GDF-5 increased the syntheses of proteoglycan and collagen II at the protein level. These results prove that Ad-GDF-5 can promote the regeneration of the disc cells matrix. In addition, our gene expression profile confirmed a significant increase in the mRNA levels of collagen II and aggrecan in the cells treated with Ad-GDF-5. We have shown that NP cells infected with Ad-GDF-5 have significantly up-regulated collagen II and aggrecan gene expression at the 21st day. These findings indicate that NP cells can express exogenous GDF-5 efficiently and continuously for a long time. And the increased ECM expression has also promoted the cell proliferation to some extent through the CCK-8 result. Results suggest that gene transfer of GDF-5 mediated by an adenovirus can potently enhance the synthesis of proteoglycan and up-regulation of collagen II and aggrecan at both protein and mRNA levels. Consistent with previous reports, Ad-GDF-5 treatment not only promotes the expression of ECM proteins but also could improve the construction and function of NP cells in vivo via the sustained transcription and translation of ECM proteins (Wang et al., 2004; Feng et al., 2008; Liang et al., 2010).
5. Conclusions
The results of our studies in vitro indicate that NP cells within degenerated discs are the ideal sites for adenovirus-mediated transfer of exogenous genes and long-term expression of transgenic proteins. Results also confirm that gene therapy may be an effective approach for restoring disc functions after they have degenerated. Insertion of the GDF-5 gene shows significant promise for clinical applications in repairing the matrix of degenerated IVD cells. Therefore, this therapeutic intervention may be a promising candidate for halting or reversing the progression of disc degeneration.
Footnotes
Project supported by the National Natural Science Foundation of China (Nos. 81171472 and 81201407), the Innovation Team Project of Sichuan Provincial Education Department (No. 13TD0030), the Major Transformation Cultivation Project of Sichuan Provincial Education Department (No. 15CZ0021), and the Science and Technology Project of Nanchong City (No. 14A0021), China
Compliance with ethics guidelines: Xu-wei LUO, Kang LIU, Zhu CHEN, Ming ZHAO, Xiao-wei HAN, Yi-guang BAI, and Gang FENG declare that they have no conflict of interest.
All procedures followed were in accordance with the ethical standards of the responsible committee on human experimentation (institutional and national) and with the Helsinki Declaration of 1975, as revised in 2008 (5). Informed consent was obtained from all patients for being included in the study. Additional informed consent was obtained from all patients for whom identifying information is included in this article.
References
- 1.Bucher C, Gazdhar A, Benneker LM, et al. Nonviral gene delivery of growth and differentiation factor 5 to human mesenchymal stem cells injected into a 3D bovine intervertebral disc organ culture system. Stem Cells Int. 2013;2013:326828. doi: 10.1155/2013/326828. (Available from: http://dx.doi.org/10.1155/2013/326828) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Chubinskaya S, Hurtig M, Rueger DC. OP-1/BMP-7 in cartilage repair. Int Orthop. 2007;31(6):773–781. doi: 10.1007/s00264-007-0423-9. (Available from: http://dx.doi.org/10.1007/s00264-007-0423-9) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Costello DJ, O'Keeffe GW, Hurley FM, et al. Transplantation of novel human GDF5-expressing CHO cells is neuroprotective in models of Parkinson’s disease. J Cell Mol Med. 2012;16(10):2451–2460. doi: 10.1111/j.1582-4934.2012.01562.x. (Available from: http://dx.doi.org/10.1111/j.1582-4934.2012.01562.x) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Cui M, Wan Y, Anderson DG, et al. Mouse growth and differentiation factor-5 protein and DNA therapy potentiates intervertebral disc cell aggregation and chondrogenic gene expression. Spine J. 2008;8(2):287–295. doi: 10.1016/j.spinee.2007.05.012. (Available from: http://dx.doi.org/10.1016/j.spinee.2007.05.012) [DOI] [PubMed] [Google Scholar]
- 5.Daans M, Luyten FP, Lories RJ. GDF5 deficiency in mice is associated with instability-driven joint damage, gait and subchondral bone changes. Ann Rheum Dis. 2011;70(1):208–213. doi: 10.1136/ard.2010.134619. (Available from: http://dx.doi.org/10.1136/ard.2010.134619) [DOI] [PubMed] [Google Scholar]
- 6.Elkasrawy MN, Hamrick MW. Myostatin (GDF-8) as a key factor linking muscle mass and bone structure. J Musculoskelet Neuronal Interact. 2010;10(1):56–63. [PMC free article] [PubMed] [Google Scholar]
- 7.Ellman MB, An HS, Muddasani P, et al. Biological impact of the fibroblast growth factor family on articular cartilage and intervertebral disc homeostasis. Gene. 2008;420(1):82–89. doi: 10.1016/j.gene.2008.04.019. (Available from: http://dx.doi.org/10.1016/j.gene.2008.04.019) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Eskola PJ, Lemmelä S, Kjaer P, et al. Genetic association studies in lumbar disc degeneration: a systematic review. PLoS ONE. 2012;7(11):e49995. doi: 10.1371/journal.pone.0049995. (Available from: http://dx.doi.org/10.1371/journal.pone.0049995) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Farndale RW, Buttle DJ, Barrett AJ. Improved quantitation and discrimination of sulphated glycosaminoglycans by use of dimethylmethylene blue. Biochim Biophys Acta. 1986;883(2):173–177. doi: 10.1016/0304-4165(86)90306-5. (Available from: http://dx.doi.org/10.1016/0304-4165(86)90306-5) [DOI] [PubMed] [Google Scholar]
- 10.Feng G, Wan Y, Balian G, et al. Adenovirus-mediated expression of growth and differentiation factor-5 promotes chondrogenesis of adipose stem cells. Growth Factors. 2008;26(3):132–142. doi: 10.1080/08977190802105917. (Available from: http://dx.doi.org/10.1080/08977190802105917) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Feng G, Wan Y, Shen FH, et al. Nucleus pulposus explant culture model. J Orthop Res. 2009;27(6):814–819. doi: 10.1002/jor.20803. (Available from: http://dx.doi.org/10.1002/jor.20803) [DOI] [PubMed] [Google Scholar]
- 12.Freemont AJ. The cellular pathobiology of the degenerate intervertebral disc and discogenic back pain. Rheumatology. 2009;48(1):5–10. doi: 10.1093/rheumatology/ken396. (Available from: http://dx.doi.org/10.1093/rheumatology/ken396) [DOI] [PubMed] [Google Scholar]
- 13.Hanley ENJr, Herkowitz HN, Kirkpatrick JS, et al. Debating the value of spine surgery. J Bone Joint Surg Am. 2010;92(5):1293–1304. doi: 10.2106/JBJS.I.01439. (Available from: http://dx.doi.org/10.2106/JBJS.I.01439) [DOI] [PubMed] [Google Scholar]
- 14.Hogan M, Girish K, James R, et al. Growth differentiation factor-5 regulation of extracellular matrix gene expression in murine tendon fibroblasts. J Tissue Eng Regen Med. 2011;5(3):191–200. doi: 10.1002/term.304. (Available from: http://dx.doi.org/10.1002/term.304) [DOI] [PubMed] [Google Scholar]
- 15.Hua G, Haiping Z, Baorong H, et al. Effect of ulinastatin on the expression of iNOS, MMP-2, and MMP-3 in degenerated nucleus pulposus cells of rabbits. Connect Tissue Res. 2013;54(1):29–33. doi: 10.3109/03008207.2012.716882. (Available from: http://dx.doi.org/10.3109/03008207.2012.716882) [DOI] [PubMed] [Google Scholar]
- 16.Kaneyama S, Nishida K, Takada T, et al. Fas ligand expression on human nucleus pulposus cells decreases with disc degeneration processes. J Orthop Sci. 2008;13(2):130–135. doi: 10.1007/s00776-007-1204-4. (Available from: http://dx.doi.org/10.1007/s00776-007-1204-4) [DOI] [PubMed] [Google Scholar]
- 17.Kim JS, Ellman MB, An HS, et al. Lactoferricin mediates anabolic and anti-catabolic effects in the intervertebral disc. J Cell Physiol. 2012;227(4):1512–1520. doi: 10.1002/jcp.22867. (Available from: http://dx.doi.org/10.1002/jcp.22867) [DOI] [PubMed] [Google Scholar]
- 18.Kim YJ, Sah RL, Doong JY, et al. Fluorometric assay of DNA in cartilage explants using Hoechst 33258. Anal Biochem. 1988;174(1):168–176. doi: 10.1016/0003-2697(88)90532-5. (Available from: http://dx.doi.org/10.1016/0003-2697(88)90532-5) [DOI] [PubMed] [Google Scholar]
- 19.Larson III JW, Levicoff EA, Gilbertson LG, et al. Biologic modification of animal models of intervertebral disc degeneration. J Bone Joint Surg Am. 2006;88(Suppl. 2):83–88. doi: 10.2106/JBJS.F.00043. (Available from: http://dx.doi.org/10.2106/JBJS.F.00043) [DOI] [PubMed] [Google Scholar]
- 20.Li X, Leo BM, Beck G, et al. Collagen and proteoglycan abnormalities in the GDF-5-deficient mice and molecular changes when treating disk cells with recombinant growth factor. Spine. 2004;29(20):2229–2234. doi: 10.1097/01.brs.0000142427.82605.fb. (Available from: http://dx.doi.org/10.1097/01.brs.0000142427.82605.fb) [DOI] [PubMed] [Google Scholar]
- 21.Liang H, Ma SY, Feng G, et al. Therapeutic effects of adenovirus-mediated growth and differentiation factor-5 in a mice disc degeneration model induced by annulus needle puncture. Spine J. 2010;10(1):32–41. doi: 10.1016/j.spinee.2009.10.006. (Available from: http://dx.doi.org/10.1016/j.spinee.2009.10.006) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Masuda K. Biological repair of the degenerated intervertebral disc by the injection of growth factors. Eur Spine J. 2008;17(Suppl. 4):441–451. doi: 10.1007/s00586-008-0749-z. (Available from: http://dx.doi.org/10.1007/s00586-008-0749-z) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Melrose J, Shu C, Young C, et al. Mechanical destabilization induced by controlled annular incision of the intervertebral disc dysregulates metalloproteinase expression and induces disc degeneration. Spine. 2012;37(1):18–25. doi: 10.1097/BRS.0b013e31820cd8d5. (Available from: http://dx.doi.org/10.1097/BRS.0b013e31820cd8d5) [DOI] [PubMed] [Google Scholar]
- 24.Moore YR, Dickinson DP, Wikesjö UM. Growth/differentiation factor-5: a candidate therapeutic agent for periodontal regeneration A review of pre-clinical data. J Clin Periodontol. 2010;37(3):288–298. doi: 10.1111/j.1600-051X.2009.01527.x. (Available from: http://dx.doi.org/10.1111/j.1600-051X.2009.01527.x) [DOI] [PubMed] [Google Scholar]
- 25.Nishida K, Kang JD, Suh JK, et al. Adenovirus-mediated gene transfer to nucleus pulposus cells. Implications for the treatment of intervertebral disc degeneration. Spine. 1998;23(22):2437–2442. doi: 10.1097/00007632-199811150-00016. (Available from: http://dx.doi.org/10.1097/00007632-199811150-00016) [DOI] [PubMed] [Google Scholar]
- 26.Nishida K, Kang JD, Gilbertson LG, et al. Modulation of the biologic activity of the rabbit intervertebral disc by gene therapy: an in vivo study of adenovirus-mediated transfer of the human transforming growth factor β1 encoding gene. Spine. 1999;24(23):2419–2425. doi: 10.1097/00007632-199912010-00002. (Available from: http://dx.doi.org/10.1097/00007632-199912010-00002) [DOI] [PubMed] [Google Scholar]
- 27.Pfirrmann CW, Metzdorf A, Zanetti M, et al. Magnetic resonance classification of lumbar intervertebral disc degeneration. Spine. 2001;26(17):1873–1878. doi: 10.1097/00007632-200109010-00011. (Available from: http://dx.doi.org/10.1097/00007632-200109010-00011) [DOI] [PubMed] [Google Scholar]
- 28.Phillips FM, Slosar PJ, Youssef JA, et al. Lumbar spine fusion for chronic low back pain due to degenerative disc disease: a systematic review. Spine. 2013;38(7):E409–E422. doi: 10.1097/BRS.0b013e3182877f11. (Available from: http://dx.doi.org/10.1097/BRS.0b013e3182877f11) [DOI] [PubMed] [Google Scholar]
- 29.Saiga K, Furumatsu T, Yoshida A, et al. Combined use of bFGF and GDF-5 enhances the healing of medial collateral ligament injury. Biochem Biophys Res Commun. 2010;402(2):329–334. doi: 10.1016/j.bbrc.2010.10.026. (Available from: http://dx.doi.org/10.1016/j.bbrc.2010.10.026) [DOI] [PubMed] [Google Scholar]
- 30.Shimer AL, Chadderdon RC, Gilbertson LG, et al. Gene therapy approaches for intervertebral disc degeneration. Spine. 2004;29(23):2770–2778. doi: 10.1097/01.brs.0000146455.11253.08. (Available from: http://dx.doi.org/10.1097/01.brs.0000146455.11253.08) [DOI] [PubMed] [Google Scholar]
- 31.Smith LJ, Nerurkar NL, Choi KS, et al. Degeneration and regeneration of the intervertebral disc: lessons from development. Dis Model Mech. 2011;4(1):31–41. doi: 10.1242/dmm.006403. (Available from: http://dx.doi.org/10.1242/dmm.006403) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Sobajima S, Kim JS, Gilbertson LG, et al. Gene therapy for degenerative disc disease. Gene Ther. 2004;11(4):390–401. doi: 10.1038/sj.gt.3302200. (Available from: http://dx.doi.org/10.1038/sj.gt.3302200) [DOI] [PubMed] [Google Scholar]
- 33.Stoyanov JV, Gantenbein-Ritter B, Bertolo A, et al. Role of hypoxia and growth and differentiation factor-5 on differentiation of human mesenchymal stem cells towards intervertebral nucleus pulposus-like cells. Eur Cell Mater. 2011;21:533–547. doi: 10.22203/ecm.v021a40. [DOI] [PubMed] [Google Scholar]
- 34.Walsh AJ, Bradford DS, Lotz JC. In vivo growth factor treatment of degenerated intervertebral discs. Spine. 2004;29(2):156–163. doi: 10.1097/01.BRS.0000107231.67854.9F. (Available from: http://dx.doi.org/10.1097/01.BRS.0000107231.67854.9F) [DOI] [PubMed] [Google Scholar]
- 35.Wang H, Kroeber M, Hanke M, et al. Release of active and depot GDF-5 after adenovirus-mediated overexpression stimulates rabbit and human intervertebral disc cells. J Mol Med. 2004;82(2):126–134. doi: 10.1007/s00109-003-0507-y. (Available from: http://dx.doi.org/10.1007/s00109-003-0507-y) [DOI] [PubMed] [Google Scholar]
- 36.Williams FM, Popham M, Hart DJ, et al. GDF5 single-nucleotide polymorphism rs143383 is associated with lumbar disc degeneration in Northern European women. Arthritis Rheum. 2011;63(3):708–712. doi: 10.1002/art.30169. (Available from: http://dx.doi.org/10.1002/art.30169) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Zaidi SH, Huang Q, Momen A, et al. Growth differentiation factor 5 regulates cardiac repair after myocardial infarction. J Am Coll Cardiol. 2010;55(2):135–143. doi: 10.1016/j.jacc.2009.08.041. (Available from: http://dx.doi.org/10.1016/j.jacc.2009.08.041) [DOI] [PubMed] [Google Scholar]
- 38.Zhang YG, Sun ZM, Liu JT, et al. Features of intervertebral disc degeneration in rat’s aging process. J Zhejiang Univ-Sci B (Biomed & Biotechnol) 2009;10(7):522–527. doi: 10.1631/jzus.B0820295. (Available from: http://dx.doi.org/10.1631/jzus.B0820295) [DOI] [PMC free article] [PubMed] [Google Scholar]