

Living with ROS through Superoxide Dismutase Enzymes
Since oxygen appeared in the biosphere some 3–5 billion years ago, all organisms have had to deal with the hazards of potentially damaging reactive oxygen species (ROS), such as superoxide, hydrogen peroxide, and hydroxyl radical. Like all organisms, pathogenic microbes produce ROS as byproducts of aerobic metabolism, but the burden of ROS is magnified when these microbes confront the oxidative burst of the host. As part of the innate immune response, macrophages and neutrophils attack invading microbes with toxic superoxide [1]. To counteract this attack, some microbial pathogens express superoxide dismutase enzymes (SOD).
SODs are metalloenzymes that use a redox-active metal to disproportionate two molecules of superoxide to oxygen and hydrogen peroxide, the latter of which is removed by catalase and peroxidase enzymes. SODs have evolved on three separate occasions, yielding a family of Mn and Fe SODs (that use either metal as co-factor), a Cu/Zn SOD family that uses Cu for catalysis, and a rare family of Ni SODs [2]. Why so many SODs? This is best answered in terms of metal bioavailability. In a typical gram-negative bacteria, such as Escherichia coli, the cytosol can have ample Mn and/or Fe, but Cu is extruded into the periplasmic/extracellular space [3]. As a result, Mn and Fe SODs are generally intracellular/cytosolic while Cu/Zn SODs are extracellular/periplasmic (Fig 1). Consistent with the endosymbiosis theory of mitochondrial evolution, this partitioning of SOD enzymes has been retained in eukaryotic mitochondria: The mitochondrial matrix (equivalent to bacterial cytosol) harbors a Mn SOD, while Cu/Zn SOD is in the mitochondrial intermembrane space and cytosol (equivalent to bacterial periplasmic/extracellular) (Fig 1).
Fig 1. The family of SOD enzymes in microbial pathogens.
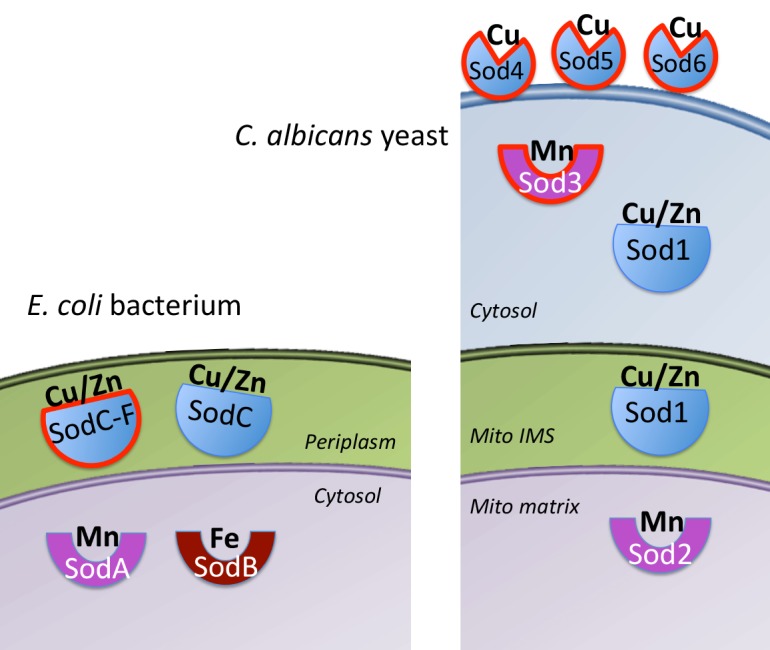
In a gram-negative bacteria such as E. coli (left), the Mn-SodA and Fe-SodA are intracellular/cytosolic where Mn and Fe metals ions are generally bioavailable. Cu is extruded into the periplasmic space, driving the evolution of Cu/Zn-containing SodC in this extra-cytosolic compartment. In eukaryotes (right), the mitochondria thought to evolve from a gram-negative bacteria exhibits the equivalent partitioning of a Mn-Sod2 to the mitochondrial matrix and Cu/Zn Sod1 to the mitochondrial IMS and cytosol. Some pathogenic microbes have acquired additional SODs (lined in red), including the highly stable prophage-derived SodC-F in pathogenic E. coli (left) and in Candida albicans (right), the cytosolic Mn-Sod3 as a backup for Cu/Zn Sod1, and the Cu-only extracellular Sod4, Sod5, and Sod6.
Custom SOD Enzymes for Bacterial Pathogenesis
The accepted nomenclature for bacterial SODs is SodA, SodB, and SodC for the Mn, Fe, and Cu/Zn SODs, respectively. Because superoxide does not generally cross biological membranes, the intracellular SodA and SodB largely remove intracellular or metabolic sources of superoxide while the periplasmic/extracellular SodC directly combats superoxide from the animal host. Not all bacterial pathogens contain extracellular SodC; for example, the Lyme disease pathogen Borrelia burgdorferi contains a single Mn-SodA. This pathogen also does not express Fe-SodB, representing a clever adaptation to limiting Fe supplies in the host [4,5].
The roles of SodA and SodB in bacterial survival and pathogenesis vary greatly depending on the species. In some cases (e.g., Salmonella typhimurium), loss of SodA has no impact on virulence [6], whereas in other instances (e.g., Streptococcus agalactiae or B. burgdorferi), loss of the intracellular SOD attenuates virulence, in spite of there being no growth inhibition in laboratory cultures [7,8]. How could an intracellular SOD promote virulence when host superoxide is extracellular? Although superoxide does not typically cross biological membranes, it can do so when protonated. In low pH environments, such as that of a macrophage phagolysosome, the protonated superoxide might enter the bacteria and serve as substrate for intracellular SodA and SodB [9]. Alternatively, the intracellular SODs may exclusively prevent damage from bacteria-derived superoxide that somehow promotes microbial fitness during infection.
Compared to SodA and SodB, the extracellular Cu/Zn SodC is well known to react with host superoxide and is a documented virulence factor for many bacteria [10]. In certain microbes, nature seems to have improved on the Cu/Zn SOD template to support pathogenesis. SodC was originally identified by Ludmil T. Benov and Irwin Fridovich in non-pathogenic laboratory strains of E. coli [11], but pathogenic E. coli including the highly virulent enteropathogenic O157:H7 serotype has acquired additional SodC genes through horizontal prophage gene transfer (Fig 1). This so-called SodC-F appears superior to chromosomal SodC in terms of protease resistance and enhanced stability for Cu binding [12]. Interestingly, this same pattern is seen with Salmonella, where all strains have a chromosomal SodCII, but highly pathogenic serotypes also carry prophage-derived SodC1 that is resistant to degradation by host proteases and exhibits superior Cu binding stability [13]. The prophage SodCII is more crucial for Salmonella virulence than the chromosomal SodC [14]. The enhanced biochemical stability of these prophage-derived SODs should prove advantageous to the pathogen in the harsh environment of the host.
Up until 2004, all Cu-containing SODs were believed to also contain a Zn co-factor. Zn does not participate directly in enzyme catalysis, but stabilizes the polypeptide and fine-tunes the redox properties of the catalytic Cu. Surprisingly, SodC of Mycobacterium tuberculosis and of the closely related M. leprae and M. avian species is an enzymatically active Cu-SOD that lacks Zn [15]. This Cu-only SOD is well suited to function under Zn-limited conditions. As part of a nutritional immunity response, the host attempts to starve pathogens of nutrient Zn [16], which would have no consequence on a Cu-only SOD.
Fungal Adaptations to Pathogenesis through Specialized SOD Enzymes
As with other eukaryotes, pathogenic fungi express a largely cytosolic Cu/Zn Sod1 and a distinct Mn-containing Sod2 in the mitochondrial matrix (Fig 1). Cu/Zn Sod1 is a documented virulence factor for Cryptococcus neoformans and Candida albicans [17,18]. Activity of fungal Sod1 is limited by the availability of its Cu co-factor [19], and Cu inside the host can vary tremendously. Cu can become very high in activated macrophages [20], and consistent with this, C. albicans mutants defective in Cu detoxification show impairments in macrophage invasion [21]. Cu can also become high in specific host niches, such as in lungs infected with C. neoformans and in the bloodstream during C. albicans and C. neoformans invasion [22,23]. However, in tissues that are targeted by C. neoformans and C. albicans (such as brain and kidney tissues), Cu availability can become very low [23,24]. We have recently shown that C. albicans adapts to such variations in host Cu by adjusting its metal co-factor selection for SODs. When host Cu is high, the yeast expresses Cu/Zn Sod1, but when host Cu is low, C. albicans will switch to a non-Cu alternative, namely a cytosolic Mn-Sod3 (Fig 1) [23]. Cytosolic Mn SODs are extremely rare in biology, and the unusual expression of Mn-Sod3 in the C. albicans cytosol endows this pathogen with uninterrupted SOD activity irrespective of host Cu [23].
In addition to this adaptation with intracellular SODs, certain fungal pathogens possess unusual extracellular Cu/Zn-like SODs that appear tailor-made for host invasion. In C. albicans, three extracellular Cu/Zn-like SODs (Sod4, Sod5, and Sod6) are attached to the cell wall through glycosylphosphatidylinositol (GPI) anchors (Fig 1). Of these, Sod5 has been well characterized and is known to directly remove host cell-derived superoxide and to promote virulence in animal models [25,26]. Upon close inspection, we noted that Sod4, Sod5, and Sod6 can only bind Cu (not Zn), making these SODs akin to Cu-only SodC of M. tuberculosis [15,27]. However, unlike Cu-only SodC, the fungal SODs also lack a region of the SOD protein known as the “electrostatic loop” (ESL), named for its role in electrostatically guiding superoxide to the active site [28]. In spite of having no ESL and no Zn, C. albicans Sod5 is a very active SOD [27]. With no ESL, the Cu site of Sod5 is surface-exposed, as opposed to the buried Cu ion of Cu/Zn SODs and Mycobacterium Cu-only SodC [27]. We propose that this open Cu site allows the fungal SOD to avidly capture Cu from the host (Fig 2). In particular niches, host Cu can become very high, and Sod5-like SODs may use this Cu to mount a powerful defense against the superoxide attack (Fig 2). Sod5-like SODs (Cu-only, no ESL, extracellular GPI-anchored) can be found throughout Basidiomycota and Ascomycota fungi [27], and they are essential for virulence of other fungi, including the pulmonary pathogen Histoplasma capsulatum [29]. Not all fungal pathogens express extracellular Cu-SODs; an example is C. neoformans. Additionally, there are examples of fungi predicted to express Sod5-like SODs that are not established pathogens, such as Podospora anserine [27]. Interestingly, P. anserine produces ROS as part of a differentiation process [30] and may employ its Sod5-like SOD to handle fungal-derived, rather than host-derived, superoxide.
Fig 2. Model for Cu-only fungal SOD5 at the macrophage–pathogen interface.
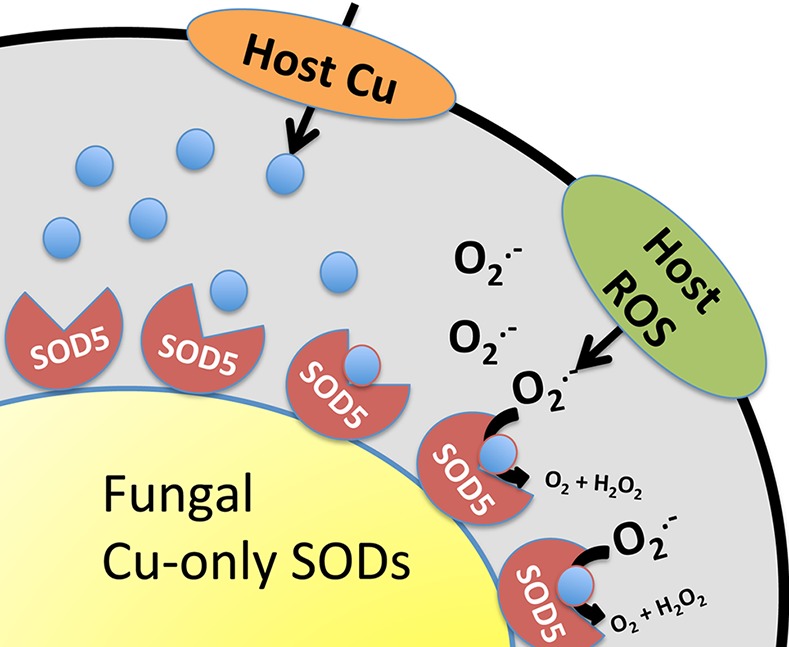
Activated macrophages may attack invading microbes with toxic levels of Cu and superoxide (O2 -). With its open Cu site, Sod5 on the surface of the fungal pathogen may be able to capture this host Cu, fueling the SOD enzyme to remove the superoxide in counterattack.
Concluding Remarks
While all aerobic organisms express SODs for endogenous superoxide, many pathogens have been armed with additional SODs designed to function in the hostile climate of the host–pathogen interface. This is particularly true for extracellular Cu-SODs of bacteria and fungi that lie in the direct line of fire from host superoxide. The phage-acquired SodCs of pathogenic E. coli and Salmonella are resilient towards host proteases and will not readily surrender their Cu co-factor to the host. Additionally, the Cu-only SODs of Mycobacterium and of fungal pathogens appear optimally designed to function in host environments of low Zn and high Cu. As a final thought, it is worth mentioning that the host cell itself must endure the oxidative insult of its own doing. Like the invading microbe, host cells secrete Cu/Zn SODs to manage extracellular superoxide, but how well this host SOD has evolved to endure the infection battleground remains to be determined.
Funding Statement
The preparation of this review was supported by NIH funding from R37 GM50016, RO1 AI119949, T32 CA009110, and F31 GM113637. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Fenlon LA, Slauch JM. Phagocyte roulette in Salmonella killing. Cell Host Microbe. 2014;15(1):7–8. Epub 2014/01/21. doi: 10.1016/j.chom.2014.01.001 S1931-3128(14)00002-X [pii]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Sheng Y, Abreu IA, Cabelli DE, Maroney MJ, Miller AF, Teixeira M, et al. Superoxide dismutases and superoxide reductases. Chem Rev. 2014;114(7):3854–918. Epub 2014/04/02. 10.1021/cr4005296 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Fu Y, Chang FM, Giedroc DP. Copper Transport and Trafficking at the Host-Bacterial Pathogen Interface. Acc Chem Res. 2014;47(12):3605–13. Epub 2014/10/14. 10.1021/ar500300n . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Troxell B, Xu H, Yang XF. Borrelia burgdorferi, a pathogen that lacks iron, encodes manganese-dependent superoxide dismutase essential for resistance to streptonigrin. J Biol Chem. 2012;287(23):19284–93. Epub 2012/04/14. M112.344903 [pii] 10.1074/jbc.M112.344903 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Aguirre JD, Clark HM, McIlvin M, Vazquez C, Palmere SL, Grab D, et al. A Manganese-Rich Environment Supports Superoxide Dismutase Activity in a Lyme Disease Pathogen, Borrelia burgdorferi . J Biol Chem. 2013;288:8468–78. Epub 2013/02/05. M112.433540 [pii] 10.1074/jbc.M112.433540 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Tsolis RM, Baumler AJ, Heffron F. Role of Salmonella typhimurium Mn-superoxide dismutase (SodA) in protection against early killing by J774 macrophages. Infect Immun. 1995;63(5):1739–44. Epub 1995/05/01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Poyart C, Pellegrini E, Gaillot O, Boumaila C, Baptista M, Trieu-Cuot P. Contribution of Mn-cofactored superoxide dismutase (SodA) to the virulence of Streptococcus agalactiae . Infect Immun. 2001;69(8):5098–106. Epub 2001/07/12. 10.1128/IAI.69.8.5098-5106.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Esteve-Gassent MD, Elliott NL, Seshu J. sodA is essential for virulence of Borrelia burgdorferi in the murine model of Lyme disease. Mol Microbiol. 2009;71(3):594–612. Epub 2008/12/02. MMI6549 [pii] 10.1111/j.1365-2958.2008.06549.x . [DOI] [PubMed] [Google Scholar]
- 9. Korshunov SS, Imlay JA. A potential role for periplasmic superoxide dismutase in blocking the penetration of external superoxide into the cytosol of Gram-negative bacteria. Mol Microbiol. 2002;43(1):95–106. Epub 2002/02/19. 2719 [pii]. . [DOI] [PubMed] [Google Scholar]
- 10. Battistoni A. Role of prokaryotic Cu,Zn superoxide dismutase in pathogenesis. Biochem Soc Trans. 2003;31(Pt 6):1326–9. . [DOI] [PubMed] [Google Scholar]
- 11. Benov LT, Fridovich I. Escherichia coli expresses a copper- and zinc-containing superoxide dismutase. J Biol Chem. 1994;269(41):25310–4. Epub 1994/10/14. . [PubMed] [Google Scholar]
- 12. D'Orazio M, Scotti R, Nicolini L, Cervoni L, Rotilio G, Battistoni A, et al. Regulatory and structural properties differentiating the chromosomal and the bacteriophage-associated Escherichia coli O157:H7 Cu, Zn superoxide dismutases. BMC Microbiol. 2008;8:166 Epub 2008/10/03. doi: 10.1186/1471-2180-8-166 1471-2180-8-166 [pii]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Fang FC, DeGroote MA, Foster JW, Baumler AJ, Ochsner U, Testerman T, et al. Virulent Salmonella typhimurium has two periplasmic Cu, Zn-superoxide dismutases. Proc Natl Acad Sci U S A. 1999;96(13):7502–7. Epub 1999/06/23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Ammendola S, Pasquali P, Pacello F, Rotilio G, Castor M, Libby SJ, et al. Regulatory and structural differences in the Cu,Zn-superoxide dismutases of Salmonella enterica and their significance for virulence. J Biol Chem. 2008;283(20):13688–99. Epub 2008/03/26. 10.1074/jbc.M710499200 [pii]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Spagnolo L, Toro I, D'Orazio M, O'Neill P, Pedersen JZ, Carugo O, et al. Unique features of the sodC-encoded superoxide dismutase from Mycobacterium tuberculosis, a fully functional copper-containing enzyme lacking zinc in the active site. J Biol Chem. 2004;279(32):33447–55. Epub 2004/05/25. 10.1074/jbc.M404699200 [pii]. . [DOI] [PubMed] [Google Scholar]
- 16. Kehl-Fie TE, Skaar EP. Nutritional immunity beyond iron: a role for manganese and zinc. Curr Opin Chem Biol. 2010;14(2):218–24. 10.1016/j.cbpa.2009.11.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Cox GM, Harrison TS, McDade HC, Taborda CP, Heinrich G, Casadevall A, et al. Superoxide dismutase influences the virulence of Cryptococcus neoformans by affecting growth within macrophages. Infect Immun. 2003;71(1):173–80. Epub 2002/12/24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Hwang CS, Rhie GE, Oh JH, Huh WK, Yim HS, Kang SO. Copper- and zinc-containing superoxide dismutase (Cu/ZnSOD) is required for the protection of Candida albicans against oxidative stresses and the expression of its full virulence. Microbiology. 2002;148(Pt 11):3705–13. . [DOI] [PubMed] [Google Scholar]
- 19. Gleason JE, Li CX, Odeh HM, Culotta VC. Species-specific activation of Cu/Zn SOD by its CCS copper chaperone in the pathogenic yeast Candida albicans . J Biol Inorg Chem. 2014;19(4–5):595–603. Epub 2013/09/18. 10.1007/s00775-013-1045-x . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. White C, Lee J, Kambe T, Fritsche K, Petris MJ. A role for the ATP7A copper-transporting ATPase in macrophage bactericidal activity. J Biol Chem. 2009;284(49):33949–56. 10.1074/jbc.M109.070201 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Douglas LM, Wang HX, Keppler-Ross S, Dean N, Konopka JB. Sur7 promotes plasma membrane organization and is needed for resistance to stressful conditions and to the invasive growth and virulence of Candida albicans . MBio. 2012;3(1). Epub 2011/12/29. 10.1128/mBio.00254-11 e00254-11 [pii] mBio.00254-11 [pii]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Ding C, Festa RA, Chen YL, Espart A, Palacios O, Espin J, et al. Cryptococcus neoformans copper detoxification machinery is critical for fungal virulence. Cell Host Microbe. 2013;13(3):265–76. Epub 2013/03/19. doi: 10.1016/j.chom.2013.02.002 S1931-3128(13)00068-1 [pii]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Li CX, Gleason JE, Zhang SX, Bruno VM, Cormack BP, Culotta VC. Candida albicans adapts to host copper during infection by swapping metal cofactors for superoxide dismutase. Proc Natl Acad Sci U S A. 2015;112(38):E5336–42. Epub 2015/09/10. 10.1073/pnas.1513447112 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Sun TS, Ju X, Gao HL, Wang T, Thiele DJ, Li JY, et al. Reciprocal functions of Cryptococcus neoformans copper homeostasis machinery during pulmonary infection and meningoencephalitis. Nat Commun. 2014;5:5550 Epub 2014/11/25. 10.1038/ncomms6550 [pii]. . [DOI] [PubMed] [Google Scholar]
- 25. Frohner IE, Bourgeois C, Yatsyk K, Majer O, Kuchler K. Candida albicans cell surface superoxide dismutases degrade host-derived reactive oxygen species to escape innate immune surveillance. Mol Microbiol. 2009;71(1):240–52. 10.1111/j.1365-2958.2008.06528.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Fradin C, De Groot P, MacCallum D, Schaller M, Klis F, Odds FC, et al. Granulocytes govern the transcriptional response, morphology and proliferation of Candida albicans in human blood. Mol Microbiol. 2005;56(2):397–415. . [DOI] [PubMed] [Google Scholar]
- 27. Gleason JE, Galaleldeen A, Peterson RL, Taylor AB, Holloway SP, Waninger-Saroni J, et al. Candida albicans SOD5 represents the prototype of an unprecedented class of Cu-only superoxide dismutases required for pathogen defense. Proc Natl Acad Sci U S A. 2014;111(16):5866–71. Epub 2014/04/09. 10.1073/pnas.1400137111 [pii]. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Getzoff ED, Cabelli DE, Fisher CL, Parge HE, Viezzoli MS, Banci L, et al. Faster superoxide dismutase mutants designed by enhancing electrostatic guidance. Nature. 1992;358(6384):347–51. . [DOI] [PubMed] [Google Scholar]
- 29. Youseff BH, Holbrook ED, Smolnycki KA, Rappleye CA. Extracellular superoxide dismutase protects histoplasma yeast cells from host-derived oxidative stress. PLoS Pathog. 2012;8(5):e1002713 Epub 2012/05/23. 10.1371/journal.ppat.1002713PPATHOGENS-D-11-02651 [pii]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Aguirre J, Rios-Momberg M, Hewitt D, Hansberg W. Reactive oxygen species and development in microbial eukaryotes. Trends Microbiol. 2005;13(3):111–8. Epub 2005/03/02. S0966-842X(05)00030-2 [pii] 10.1016/j.tim.2005.01.007 . [DOI] [PubMed] [Google Scholar]