

Abstract
The trace mineral boron is a micronutrient with diverse and vitally important roles in metabolism that render it necessary for plant, animal, and human health, and as recent research suggests, possibly for the evolution of life on Earth. As the current article shows, boron has been proven to be an important trace mineral because it (1) is essential for the growth and maintenance of bone; (2) greatly improves wound healing; (3) beneficially impacts the body’s use of estrogen, testosterone, and vitamin D; (4) boosts magnesium absorption; (5) reduces levels of inflammatory biomarkers, such as high-sensitivity C-reactive protein (hs-CRP) and tumor necrosis factor μ (TNF-μ); (6) raises levels of antioxidant enzymes, such as superoxide dismutase (SOD), catalase, and glutathione peroxidase; (7) protects against pesticide-induced oxidative stress and heavy-metal toxicity; (8) improves the brains electrical activity, cognitive performance, and short-term memory for elders; (9) influences the formation and activity of key biomolecules, such as S-adenosyl methionine (SAM-e) and nicotinamide adenine dinucleotide (NAD+); (10) has demonstrated preventive and therapeutic effects in a number of cancers, such as prostate, cervical, and lung cancers, and multiple and non-Hodgkin’s lymphoma; and (11) may help ameliorate the adverse effects of traditional chemotherapeutic agents. In none of the numerous studies conducted to date, however, do boron’s beneficial effects appear at intakes < 3 mg/d. No estimated average requirements (EARs) or dietary reference intakes (DRIs) have been set for boron—only an upper intake level (UL) of 20 mg/d for individuals aged ≥ 18 y. The absence of studies showing harm in conjunction with the substantial number of articles showing benefits support the consideration of boron supplementation of 3 mg/d for any individual who is consuming a diet lacking in fruits and vegetables or who is at risk for or has osteopenia; osteoporosis; osteoarthritis (OA); or breast, prostate, or lung cancer.
The trace mineral boron is a micronutrient with diverse and vitally important roles in metabolism that render it necessary for plant, animal, and human health, and, possibly, as recent research suggests, for the evolution of life on Earth. The current review focuses primarily on boron’s most salient effects on human health, including its impact on bone development and regeneration, wound healing, the production and metabolism of sex steroids and vitamin D, and the absorption and use of calcium and magnesium. In addition, boron has anti-inflammatory effects that can help alleviate arthritis and improve brain function and has demonstrated such significant anticancer effects that boronated compounds are now being used in the treatment of several types of cancer. A summary of the evidence suggesting that boron should be given consideration as an essential micronutrient is provided, together with leading dietary sources and intake recommendations.
Salient Effects
Bone Growth and Maintenance
Scientists have known for many years that boron is essential for healthy bones. In 1985, the US Department of Agriculture (USDA) conducted an experiment in which postmenopausal women (n = 12) who had been put on a low-boron diet (0.25 mg/d for 119 d) were supplemented with 3 mg/d of boron during two 28-day trials. In one trial, magnesium intake was low; in the other, magnesium intake was adequate. With boron supplementation, the women’s daily urinary excretion of calcium was reduced by 44%. The reduction in calcium loss resulting from boron supplementation was 52 mg/d when the women were low in magnesium and 22 mg/d when magnesium levels were adequate.1
Boron plays an important role in osteogenesis, and its deficiency has been shown to adversely impact bone development and regeneration.2 Boron influences the production and activity of steroid hormones, actions via which this trace mineral is involved in the prevention of calcium loss and bone demineralization. Boron supplementation has repeatedly been shown to markedly reduce urinary excretion of both calcium and magnesium and to increase serum levels of estradiol and calcium absorption in peri- and postmenopausal women.1,3 Boron also beneficially impacts vitamin-D utilization. Supplementation with boron stimulates bone growth in vitamin-D deficient animals and alleviates dysfunctions in mineral metabolism characteristic of vitamin-D deficiency.4
Animal studies published in 20085 and 20096 found that healing of the alveolar bone—a ridge of compact bone that contains the tooth sockets on the maxillae and mandible (ie, the bones that hold teeth)—was inhibited in boron-deficient rats. Compared with boron-adequate rats (3 mg/kg/d of boron in the diet) when evaluated at 7 and 14 days, boron-deficient animals (0.07 mg/kg/d boron in the diet) had significant reductions in osteoblast surface (57% and 87% at 7 d and 14 d, respectively) concomitant with increases in quiescent surface (120% and 126% at 7 d and 14 d, respectively), indicating that boron deficiency can result in impaired bone healing due to a marked reduction in osteogenesis.
In 2010, Hakki et al7 published research investigating the mechanisms underlying boron’s effects on osteogenesis. Boron was determined to induce mineralization of osteoblasts by regulating the expression of genes related to tissue mineralization and the actions of key hormones (17β-estradiol [E2], testosterone, and vitamin D) involved in bone growth and turnover. Boron’s induction of tissue mineralization also underlies boron’s beneficial effects on wound healing.
Wound Healing
Since 1990, boron has been shown to significantly improve wound healing. Application of a 3% boric acid solution to deep wounds reduced the time required in intensive care by two-thirds.8 In 2000, in vitro research using human fibroblasts showed that a boric-acid solution improved wound healing through action on the extracellular matrix.9 Further in vitro studies published in 2002 revealed that these beneficial effects of boron were due to direct actions on specific enzymes found in fibroblasts: elastase, trypsin-like enzymes, collagenase, and alkaline phosphatase.10 The most common cells in animal connective tissue, fibroblasts synthesize the extracellular matrix and collagen and play a critical role in wound healing. Boron facilitates the activity of these key enzymes in fibroblasts, thus improving extracellular-matrix turnover.
Expression of Extracellular-matrix Proteins
Hakki et al7 extended understanding of the mechanisms behind boron’s effects a step further, showing that boron regulates the messenger RNA (mRNA) expression of a wide range of extracellular-matrix proteins, not only those involved in wound repair but also the mineralized tissue-associated proteins, collagen type 1 (COL1), osteopontin (OPN), bone sialoprotein (BSP), and osteocalcin (OCN). The combined effects of these actions boost osteoblast-cell viability, proliferation, and morphology as well as the mineralization of bone cells.
Further research conducted by other investigators on human bone marrow stromal cells confirmed earlier discoveries of the mechanisms through which boron boosts osteogenic differentiation and continued to add to the list. Boron was also found to increase mRNA expression of alkaline phosphatase and bone morphogenetic proteins (BMPs).11 Several other researchers have now confirmed that boron regulates protein levels of key BMPs—BMP-4, BMP-6, and BMP-7—plus the mRNA expression of runt-related transcription factor 2 (RUNX2).
The BMPs are multifunctional growth factors that belong to the transforming growth factor β (TGF-β) superfamily, and several induce formation of new cartilage and bone. Of the 20 BMP family members that have been identified and characterized to date,12,13 BMP-2, BMP-4, BMP-6, and BMP-7, all of which show transcription and activity regulated by boron, have been found necessary to induce osteoblastic differentiation in primary human, mesenchymal stem cells.14,15
Boron was also determined to regulate production of RUNX2, also known as core-binding factor subunit α-1 (CBF-α1). RUNX2 is essential to osteoblastic differentiation and bone formation and to bone maintenance. It is a transcription factor that works cooperatively with BMPs to stimulate osteoblast gene expression and the differentiation of mesenchymal stem cells into osteoblasts, and it remains active in mature osteoblasts. When active RUNX2 levels are reduced, expression of the genes encoding the main bone-matrix proteins drops, including BSP, OCN, OPN, and COL1.16-19
Regulation of Sex Hormones
Increased levels of sex steroids have been demonstrated in both men and women after boron supplementation.1,20 In 1987, Nielsen et al1 reported that dietary boron repletion in postmenopausal women (n = 13), who were previously on a low-boron diet, significantly increased their serum estradiol (E2) and testosterone levels, particularly for those women whose dietary intake of magnesium was low. In women on a low-magnesium diet, E2 almost doubled, increasing from an average of 21.1 pg/mL to 41.4 pg/mL. Testosterone more than doubled, rising from an average of 0.31 ng/mL to 0.83 ng/mL. Similar increases were seen in the women on an adequate-magnesium diet: E2 rose from an average of 15.5 pg/mL to 38.0 pg/mL, and testosterone increased from 0.38 ng/mL to 0.65 ng/mL. In 1997, Naghii et al21 published findings of a similar increase in serum levels of E2 in healthy males (n = 18) after 4 weeks of dietary supplementation with boron.
After only 1 week of boron supplementation of 6 mg/d, a further study by Naghii et al20 of healthy males (n = 8) found (1) a significant increase in free testosterone, which rose from an average of 11.83 pg/mL to 15.18 pg/mL; and (2) significant decreases in E2, which dropped from 42.33 pg/mL to 25.81 pg/mL. All of the inflammatory biomarkers that were measured also decreased: (1) interleukin (IL) 6, from 1.55 pg/mL to 0.87 pg/mL; (2) high-sensitivity C-reactive protein (hs-CRP) by approximately 50%, a remarkable decrease, from 1460 ng/mL to 795 ng/mL; and (3) tumor necrosis factor α (TNF-α) by approximately 30%, from 12.32 to 9.97 pg/mL. Levels of dihydrotestosterone, cortisol, and vitamin D increased slightly.
The significant decrease in the men’s plasma E2 after 1 week of boron supplementation suggests a higher rate of conversion of total testosterone (T) to free testosterone (FT) in the testosterone metabolic pathway. In support, the ratios of FT/T, T/E2, and FT/E2 were all significantly increased, indicating boron had androgen amplifier effects: (1) FT/T (pg/mL/ng/mL) increased from 3.62 to 4.66; (2) T/E2 (ng/mL) rose from 91.68 to 148; and (3) FT/E2 (ng/mL) from 0.31 to 0.67.
It is well known that approximately 98% of testosterone molecules are bound to proteins in the blood, principally to sex hormone–binding globulin (SHBG), and are not bioavailable because bound hormones cannot exit capillaries.22 Thus, the elevation of unbound free testosterone seen with boron supplementation may have significant beneficial ramifications, particularly in aging men in whom, typically, levels of SHBG increase and levels of FT decrease.23
Prevention of Vitamin-D Deficiency
Boron has been shown to increase serum levels of 25-hydroxyvitamin D3 (25[OH]D3) in animal studies4,24 and of vitamin D–deficient individuals in human studies.25,26 In a clinical trial25 in which middle-aged men and women (n = 15) were placed on a low-boron diet, which was also marginal in magnesium and copper status, for 63 days (0.23 mg B/2000 kcal), 25(OH)D3 rose significantly after boron supplementation (3 mg/d as sodium borate) for an additional 49 days. Levels of 25(OH)D3 rose from an average of 44.9 nM after the 63 days of boron deprivation to 62.4 nM after the 49 days of boron repletion, a 39% increase.
Similar results were seen in an open pilot study of middle-aged individuals (n = 13) predetermined to be vitamin D deficient (serum 25[OH]D3 < 12 ng/mL). Levels of 25(OH)D3 were studied during boron supplementation of 6 mg/d for 60 days using calcium fructoborate, Ca([C6H10O6]2B)2·4H2O, a boron-containing complex that occurs naturally in fruit.26 The study took place in Serbia with supplementation beginning in October and concluding by January; in other words, the study occurred during the fall transition to winter, a time when vitamin-D status would be expected to worsen. Yet, with boron supplementation, 25(OH)D3 levels rose significantly, with an average rise of 20%.27
How does boron exert its hormonal effects? In sum, boron increases the biological half-life and bioavailability of E2 and vitamin D.
Boron Increases Half-life and Bioavailability of Sex Hormones and Vitamin D.
Boron’s beneficial effects on bone metabolism are due in part to the roles it plays in both producing E2 and in increasing its biological half-life and that of vitamin D. Regarding 17β-estradiol, the simplest and preferred pathway for its production is reduction of the keto group of estrone by a tetrahydroborate salt, potassium borohydride.28
Regarding vitamin D, Miljkovic et al27 proposed in an excellent paper in Medical Hypotheses that boron suppresses the activity of 24-hydroxylase, the microsomal enzyme primarily responsible for catabolism of 25(OH)D3. A number of recent papers, which are discussed in the following text, have provided evidence to support this hypothesis.
The hypothesis by Milijkovic et al27 also accounts for boron’s well-recognized upregulation of 17β-estradiol levels in women, including postmenopausal women receiving hormone replacement therapy, because catabolism of 17β-estradiol is also achieved by microsomal enzymes catalyzing vicinal hydroxylations (eg, 24-hydroxylase). This suggests a more general hypothesis: Nutritional boron can inhibit a range of microsomal enzymes that insert hydroxyl groups vicinal to existing hydroxyls in steroids, which include enzymes that catabolize 17β-estradiol, 25(OH)D3, and 1α,25-dihydroxyvitamin D3 (1α,25[OH]2D3).29 As noted by Miljkovic et al27:
Boron readily forms covalent complexes with cis-vicinal dihydroxy compounds. Thus, it is conceivable that it can form such a complex with 24,25-dihydroxyvitamin D, the end product of the reaction of 25(OH)D3 with 24-hydroxylase. This postulated complex might either act as a competitive inhibitor of the 24-hydroxylase reaction, or alternatively, perhaps could act to down-regulate expression of the enzyme. Another possibility is that boron is a direct inhibitor of the enzyme at very modest concentrations; indeed, boron can inhibit numerous enzymes …
Therefore, boron’s beneficial hormonal effects are likely to be a result of its general impact on vicinal hydroxylations of steroids.
In chemistry, vicinal stands for any 2 functional groups that are bonded to 2 adjacent carbon atoms. Hydroxylation is the addition of a hydroxyl group (-OH). Boronic acids’ unique feature is their ability to form reversible covalent complexes with molecules containing vicinal hydroxyl groups. Boronic acids act as Lewis acids, substances that can accept a lone pair of electrons from another molecule in completing the stable group of one of their own atoms. For example, H+ is a Lewis acid because it can accept a lone pair, completing its stable form, which requires 2 electrons. The current definition of a Lewis acid is:
Deactivation of Vitamin D.
Cholecalciferol (vitamin D3) is both activated and degraded via hydroxylation. Synthesis of vitamin D begins in the skin as a nonenzymatic process when the absorption of ultraviolet B (UVB) radiation in sunlight results in conversion of 7-dehydrocholesterol, a metabolite of cholesterol stored in the skin, to precholecalciferol. Precholecalciferol is immediately converted into cholecalciferol (vitamin D3) and transported to the liver where it undergoes hydroxylation by 25-hydroxylase. Actually, 75% of serum 25(OH)D3 arises from the action of 1,25-dihydroxyvitamin D3 24-hydroxylase (CYP2R1), and an, as yet, unidentified enzyme is responsible for the remaining 25% of 25-hydroxylation of cholecalciferol.30
The result of this 25-hydroxylation is the formation of calcidiol (25[OH]D3). A second hydroxylation takes place in the kidney, where calcidiol undergoes hydroxylation by 1α-hydroxylase (CYP27B1) at the C-1 position to form the hormonal, most-active metabolite, calcitriol (1,25[OH]2D3). The 24-hydroxylation of 25(OH)D3 and calcitriol, which is accomplished by 1,25-dihydroxyvitamin D3 24-hydroxylase (CYP24A1), leads to the degradation of these vitamin-D metabolites.31
Boron May Improve the Effects of Vitamin D Supplementation in “Nonresponders”.
Support for boron’s potential for beneficial effects on vitamin D levels is suggested by recent research investigating the relationship between levels of DNA methylation of CYP24A1, a 24-hydroxylase enzyme, and CYP2R1, a 25-hydroxylase enzyme, and also by examining postmenopausal women’s ability to increase serum 25(OH)D3 in response to vitamin-D supplementation.32
Researchers randomly assigned 446 white, postmenopausal women to a calcium and vitamin D intervention, 1100 IU/day, for at least 12 months. From these subjects, 18 with the highest 12-month increase in serum 25(OH)D3 were selected as responders, and 18 with the lowest 12-month increase in serum 25(OH)D3 were selected as nonresponders. Levels of deoxyribonucleic acid (DNA) methylation for the 2 groups were compared.
Methylation silences or shuts down activity in these CpG sites; thus, greater methylation leads to lessened production of the hydroxylase. Responders had significantly lower baseline levels of DNA methylation in the promoter region of CYP2R1—a 25-hydroxylase that activates vitamin D—than nonresponders had, 8% versus 30%, respectively. That finding indicated a greater production of 25-hydroxylase, and, thus, greater potential for vitamin-D activation.
Responders also had lower baseline levels of DNA methylation in the promoter region of CYP24A1, a 24-hydroxylase that degrades vitamin D, than the nonresponders had, 13% versus 32%, respectively. This finding possibly indicates a balance in responders’ ability to both activate and then degrade vitamin D. Although nonresponders’ levels of DNA methylation of CYP24A1 were greater, it is unknown whether the 24-hydroxylase enzymes that they produced had a higher-than-average rate of activity. Boron was not included in this investigation, but it would have been germane to have assayed subjects’ boron intake and serum levels because boron inhibits the activity of 24-hydroxylase.
… a molecular entity (and the corresponding chemical species) that is an electron-pair acceptor, and therefore, able to react with a Lewis base to form a Lewis adduct by sharing the electron pair furnished by the Lewis base. A Lewis base is any species that donates a pair of electrons to a Lewis acid to form a Lewis adduct.33
As already mentioned, serum concentrations of both 25(OH)D3 and 17β-estradiol increase when boron is supplemented. Because this effect is seen in postmenopausal women receiving hormone replacement therapy, a reduction in E2 catabolism, rather than increased E2 synthesis, is responsible. Each of the major routes of E2 catabolism involve hydroxylations (ie, the introduction of a vicinal hydroxyl group at the 2, 4, or 16 position of 17β-estradiol), which is already hydroxylated at the 3 and 17 positions. This addition indicates that boron is a potent inhibitor for microsomal enzymes that catalyze the insertion of hydroxyl groups vicinal to existing hydroxyl groups in steroids.
Two specific examples of that process are the E2 hydroxylases and 24-hydroxylase, the enzymes that catalyze the conversion of 25(OH)D3 and 1,25-dihydroxyvitamin D3 (1,25[OH]2D3) into 24-hydroxylated products, which constitute the degradation products of the vitamin-D molecule. A summary of the activation and deactivation of vitamin D.34
Magnesium Absorption
Boron significantly improves magnesium absorption and deposition in bone, yet another beneficial effect of boron’s inhibition of 17β-estradiol degradation. Thus, boron is a factor in magnesium’s myriad beneficial effects. Magnesium’s importance, in bone alone, is illustrative of the widespread ramifications of boron insufficiency.
Approximately 60% of the magnesium in the human body is found in bone, where it is a cofactor for key enzymes that regulate calcium metabolism. The majority of magnesium in bone resides on cortical bone, an integral part of the structure of apatite crystal.35 Apart from its structural role in apatite crystals, magnesium is required in osteoblasts and osteoclasts and in all living cells, within which magnesium is fundamental for adenosine triphosphate (ATP) production and serves as the cofactor of more than 300 enzymes involved in lipid, protein, and nucleic acid synthesis. Because of its positive charge, magnesium stabilizes cell membranes, balances the actions of calcium, and functions as a signal transducer.31
Anti-inflammatory Effects
A number of papers have indicated that boron reduces levels of inflammatory biomarkers.20,36,37 In a recent human trial involving healthy male volunteers (n = 8), a significant increase in concentrations of plasma boron occurred 6 hours after supplementation with 11.6 mg of boron, coupled with significant decreases in levels of hs-CRP and TNF-α. One week of boron supplementation 10 mg/d resulted in a 20% decrease in the plasma concentration of TNF-α, from 12.32 to 9.97 pg/mL, and in remarkable decreases (approximately 50%) in plasma concentration of hs-CRP, from 1460 to 795 ng/mL, and of IL-6, from 1.55 to 0.87 pg/mL.
Is boron adequacy important? Consider that elevated hs-CRP is associated with an increased risk for breast cancer,38 obesity and metabolic syndrome (MetS) in children,39 atherosclerosis, unstable angina, insulin resistance, type 2 diabetes,40,41 nonalcoholic fatty liver disease (NAFLD),42,43 metastatic prostate cancer,44 lung cancer,45 adult depression, depression in childhood and psychosis in young adult life,46,47 coronary heart disease, and stroke.48-51
Anti-inflammatory Effects in Osteoarthritis
Epidemiological evidence, case reports, and controlled animal and human studies have provided evidence for the use of boron as a safe and effective treatment for osteoarthritis (OA).52-54 Examining the relationship between boron administration and OA prevalence around the world, researchers discovered that in areas where boron intake is greater than or equal to 1 mg/d, the estimated incidence of arthritis ranges from 20% to 70%. In contrast, in areas where boron intake is usually 3 to 10 mg/d, estimated incidence of arthritis ranges from 0% to 10%.55 The boron concentration has been found to be lower in the femur heads, bones, and synovial fluid of OA patients compared with individuals without OA.56
Analysis of animal studies, in which rats with induced arthritis benefited from orally or intraperitoneally administered boron, revealed that boron downregulates production and activity of serine protease enzymes involved in the inflammatory response.57 Human studies of boron deprivation and repletion have shown that boron significantly increases erythrocyte superoxide dismutase (SOD) activity. In a study in which 63 days of boron depletion were followed by 49 days of boron supplementation 3 mg/d, SOD rose from 3091 U/g Hb to 3231 U/g Hb in men older than 45 years, from 2666 U/g Hb to 3169 U/g Hb in postmenopausal women, and from 2520 U/g Hb to 3327 U/g Hb in postmenopausal women on estrogen therapy.58
Human clinical evidence for boron’s use in the treatment of OA patients was first provided by a double-blind, placebo-controlled supplementation trial conducted in Australia, in which a significantly favorable response to a supplement of 6 mg of boron per day (sodium tetraborate decahydrate) was seen in 20 subjects with OA; 50% of subjects receiving supplemental boron improved compared with only 10% of those receiving the placebo.61
Clinical Effects of Hypomagnesemia.
In addition to its direct effects on the structure and cells of the skeleton, magnesium impacts bone indirectly by affecting the homeostasis of the 2 master regulators of calcium homeostasis (ie, parathyroid hormone [PTH] and 1,25[OH]2D3).59 PTH signaling involves the increase of cyclic adenosine monophosphate (cAMP) through the activation of adenylate cyclase, which requires magnesium.
Hypomagnesemia impairs secretion of PTH and renders target organs refractory to PTH. Reduced secretion of PTH or impaired peripheral response to the hormone leads to low serum concentrations of 25(OH)D3. In addition, the hydroxylase enzyme 25-hydroxycholecalciferol-1-hydroxylase, which is responsible for production of the most active, hormonal form of vitamin D, calcitriol, requires magnesium as its cofactor. Consequently magnesium deficiency impairs calcitriol production.
Hypomagnesemia promotes inflammation.60 Substance P is released at high levels in magnesium deficiency, and the levels of proinflammatory cytokines—TNF-α, IL-1, and IL-6—increase both in the serum and the bone marrow. Magnesium also plays an important role in carbohydrate metabolism; its deficiency provokes and worsens insulin resistance.
Insulin resistance and resulting high glucose concentrations (>12 mM) alter the biomineralization process in osteoblastic cells so that although a number of factors promoting osteoblast activity are triggered, less calcium is deposited. High glucose increases mRNA expression of the receptor activator of nuclear factor kappa B ligand (RANKL), OCN, BSP, and RUNX2, but it decreases expression of the decoy for RANKL (ie, osteoprotegerin) and increases mRNA expression of proinflammatory cytokines (eg, IL-6, IL-10, and TNF-α). Osteoblastic cells cultured in normal glucose conditions had a Ca/P ratio of 1.48 and 1.60 at 7 days and 14 days, respectively. In contrast, mineral deposits formed by osteoblastic cells cultured with high d-(+)-glucose concentrations had Ca/P ratios of 0.78 and 1.29 at 7 days and 14 days, respectively. These findings represent just a few of the ways in which hypomagnesemia causes accelerated bone loss. Adequate boron intake of 3 mg/d can help prevent hypomagnesemia.
In vitro studies have shown anti-inflammatory effects of calcium fructoborate—a naturally occurring, plant-based, boron-carbohydrate complex—on cellular cultures,62 and an open-label pilot study (n = 20) found that calcium fructoborate had highly positive effects on OA symptoms.26 In the 8-week pilot study, OA patients were divided into 2 groups, those with mild and medium forms of OA and those with severe cases. Two assessment measures were used, the Western Ontario and McMaster Universities Arthritis Index (WOMAC) and the Newnham criteria. Mild-to-medium cases received 6 mg/d of supplemental boron; severe cases received 12 mg/d. In subjects with mild and medium OA, average pain reduction was 62.5% in 4 weeks and 70.8% in 8 weeks. In the first 4 weeks, 80% of mild-to-medium OA subjects reduced or eliminated their analgesic (ibuprofen) use. By 8 weeks, 67% had stopped using the nonsteroidal anti-inflammatory drug (NSAID) medication. Joint rigidity disappeared in one-half of the mild and medium OA patients in the first 4 weeks and decreased in the remaining one-half by 87.5% on average. By week 8, all subjects with mild- and medium-level OA were rigidity free. Mobility and flexibility were significantly improved in 71.4% of subjects at 4 weeks and in 77.8% at 8 weeks.
High-sensitivity C-reactive Protein.
C-reactive protein (CRP) is an acute-phase protein produced in the liver in response to increased immune-cell production of proinflammatory cytokines (eg, IL-1, IL-6, IL-8, and TNF-α). CRP is a known predictor of risk for cardiovascular disease (CVD), heart attack, and stroke. Studies have also shown a clear link between above-normal levels of CRP and OA, periodontal disease, and risk for fracture. In Japan, where women typically have substantially lower levels of CRP than their Caucasian counterparts, women whose CRP levels were above what is normal for the Japanese had more than double the risk for fracture—2.22 if in the medium quartile and 2.40 if in the quartile with the highest CRP levels.63
Two tests can be used to evaluate CRP: CRP and hs-CRP. Both tests measure the same molecule in the blood. CRP is the standard test and measures a much wider, higher range of CRP levels than hs-CRP, but CRP does not do well in capturing the lower ranges, which are important in determining whether low-level, chronic inflammation is present and which may be a contributing factor to an ongoing disease process. The test for hs-CRP accurately detects the lower concentrations of CRP. People with chronically elevated inflammation (eg, individuals with conditions such as arthritis or periodontal disease) should have the CRP, not the hs-CRP test run. Their CRP levels will be very high, often too high to be meaningful or even measured using the hs-CRP test. The hs-CRP test is the better choice for apparently healthy people to determine whether they have low-grade chronic inflammation. The hs-CRP test measures CRP in the range from 0.5 to 10 mg/L. The standard CRP test measures CRP in the range from 10 to 1000 mg/L.
Hs-CRP usually is ordered as one of several tests in a cardiovascular risk profile, often together with tests for cholesterol and triglycerides, because the best way to predict cardiovascular risk is to combine a good marker for inflammation, such as hs-CRP, with the lipid profile. Research has now shown that healthy people whose hs-CRP results are in the high end of the normal range, above 3.0 mg/L, have 1.5 to 4 times the risk of having a heart attack compared with those whose hs-CRP values are at the low end of the normal range, 1.0 mg/L or less.
Because the hs-CRP test serves as a marker for inflammation, it is important that the patient is not recently recovering from a cold, flu, other infection, or injury when the test is run. As noted earlier, CRP is an acute-phase protein, so levels dramatically rise in response to any recent illness, infection, or trauma, including dental work. For that reason, any acute inflammation will raise the amount of CRP and give a falsely elevated estimate of risk. Also, patients should be instructed not to take an NSAID, as those medications will temporarily depress CRP levels, as will statins, giving a falsely lowered estimate of risk.
Women on conventional hormone replacement therapy (HRT) (eg, Premarin or Provera), but not on bioidentical hormone replacement therapy (BHRT), have been shown to have elevated hs-CRP levels, yet another reason to consider use of BHRT rather than HRT.
In the severe OA group, average pain reduction was 47.9% at 4 weeks and 64.5% at 8 weeks. In the first 4 weeks, 40% of subjects with severe OA reduced or eliminated their analgesic (ibuprofen) use. By week 8, 75% had quit using their NSAID medication (ibuprofen). Joint rigidity disappeared in one-half of the severe OA patients in the first 4 weeks. In the remaining one-half, joint rigidity decreased significantly, an average rigidity reduction of 50%. Mobility and flexibility were significantly improved in 50% of severe OA subjects at 4 weeks and in 62.5% at 8 weeks.
Based on the results of animal studies,62 the researchers hypothesized that boron’s anti-inflammatory effects result from inhibition of the oxidative burst by scavenging cells (leukocytes) and excessive activity by neutrophils (ie, white cells that scavenge debris) and invaders outside of the circulatory system. Boron also boosts free-radical scavenging by raising levels of a triumvirate of antioxidant enzymes in blood and cells: SOD, catalase, and glutathione peroxidase.
Scorei et al54 then conducted a double-blind, placebo-controlled pilot study to evaluate the effects of different dosages of calcium fructoborate on systemic inflammation and dyslipidemia markers in middle-aged people with primary OA. Study subjects (n = 72) were randomly assigned to 4 groups: (1) group 1 received 1.5 mg of boron, 2 ×/d; (2) group 2 was given 3 mg, 2 ×/d; (3) group 3 received 6 mg, 2 ×/d; and (4) group 4 received placebo, 2 ×/d. Fifteen days of dietary supplementation with calcium fructoborate lowered inflammatory biomarkers—CRP, fibrinogen (FBR), and erythrocyte sedimentation rate (ESR)—in all groups except the placebo group; no effect was seen on lipids. In groups 1, 2, and 3 (ie, those supplemented with boron in the form of calcium fructoborate), significant decreases occurred in ESR levels: a drop of -10.25%, -11.9%, and -8.5% in groups 1, 2, and 3, respectively, compared with baseline. In the placebo group, the ESR level rose 36.36%. FBR also dropped in those supplemented: -13.73 in group 1, -2.05 in group 2, and -4.18 in group 3. In the placebo group, FBR rose 4.10. CRP also dropped in all supplemented groups: -60.25 in group 1, -26.66 in group 2, and -17.54 in group 3. In the placebo group, CRP increased 5.47.
Most recently, calcium fructoborate 110 mg 2 ×/d, which provides approximately 3 mg of boron 2 ×/d or 6 mg/d, was shown to improve knee discomfort within the first 14 days of treatment. In this study, subjects with self-reported knee discomfort (n = 60) were randomized into 2 groups given either calcium fructoborate or placebo.64 At both 7 days and 14 days, significant reductions in the mean, within-subject changes in scores, as measured using the WOMAC index and the McGill Pain Questionnaire (MPQ), were seen in the group supplemented with calcium fructoborate when compared with the placebo group. Estimated treatment differences for the MPQ score were -5.8 and -8.9 at days 7 and 14, respectively. Estimated differences for the WOMAC score were -5.3 and -13.73 at days 7 and 14, respectively. Negative values indicate greater reductions in reported discomfort. No changes in the WOMAC or MPQ scores were seen in the placebo group.65
CRP levels are typically higher in patients with OA compared with normal controls, and CRP levels with reference values above 0.5 mg/dL in OA patients are associated with disease progression.66,67 As noted earlier, in addition to OA and CVD, higher levels of CRP are also associated with NAFLD, MetS, type 2 diabetes, obesity, depression, kidney disease, and osteoporosis. Boron, in combination with plant-sourced calcium, as calcium fructoborate, has been shown to significantly reduce blood levels of CRP in humans.68
Protection Against Malathion-induced Oxidative Stress
In a recent study, boron protected animals chronically exposed to low levels of malathion, a widely used pesticide that causes oxidative stress even at the low levels at which humans are exposed to it in the food supply.69 Malathion administration 100 mg/kg/d by gastric gavage increased malondialdehyde, nitric oxide and 8-hydroxy-2’-deoxyguanosine (8-OHdG) levels, and markers of liver damage. It decreased acetylcholinesterase and reduced glutathione; superoxide dismutase; and catalase activities in blood, liver, kidney, and brain tissues. Administration of boron 5, 10, and 20 mg/kg/d reversed malathion-induced oxidative stress, lipid peroxidation, and suppression of antioxidant enzyme activity. Boron decreased malathion-induced oxidative stress, enhanced antioxidant defense mechanisms, and regenerated damaged liver, kidney, and brain tissues in rats.
Brain Activation and Psychological Function
Assessments of brain electrical activity in both animals and humans have shown that boron deprivation—0.12 μg/g in the diet as boric acid—results in decreased brain electrical activity.70 In mature rats, boron deprivation was associated with decreased high-frequency and increased low-frequency brain electrical activity, consistent with decreased arousal, suggesting that boron may play an important role in the maintenance of brain activation in animals. In humans, boron deprivation (<0.3 mg/d) resulted in poorer performance on tasks of motor speed and dexterity, attention, and short-term memory. A series of experiments conducted in otherwise healthy older men and women found that relatively short periods, 42 to 73 days, of restricted boron intake adversely affected brain function and cognitive performance. The most consistent EEG finding was that low boron intake resulted in a shift toward more activity in the low frequencies and less activity in the high, dominant frequencies of the EEG spectrum, the same effect typically observed in response to nonspecific malnutrition and heavy-metal toxicity. Increased low frequency activity is characteristic of states of reduced mental alertness, is associated with lowered ability to perform vigilance and psychomotor tasks, and has been related to impaired memory performance.
Heavy-metal Toxicity
The effectiveness of some boron compounds—boric acid, borax, colemanite, and ulexite—on the genotoxicity induced by heavy metals—arsenic trioxide, colloidal bismuth subcitrate, cadmium chloride, mercury chloride, and lead chloride—was assessed in human blood cultures.71 Sister chromatid exchange (SCE) and micronuclei (MN) assays were performed to establish DNA damage in lymphocytes, and oxidative stress was evaluated by estimating the changes in the main, antioxidant, enzyme activities and in the levels of total glutathione in erythrocytes. Heavy-metal treatments increased the frequency of both SCE and MN and the plasma levels of malondialdehyde, a marker of oxidative stress, and decreased the antioxidant enzyme activities and the level of total glutathione compared to controls. All boron-tested compounds (5-20 ppm) significantly reduced all genotoxic effects that were induced by low doses of heavy metals.
Production of Key Biomolecules
Not only is boron an inhibitor of 24-hydroxylase, but it also influences the formation and activity of boroesters in biomolecules containing cis-hydroxyl groups. Such boron-containing biomolecules include those that contain ribose (eg, S-adenosyl methionine [SAM-e], diadenosine phosphates, and nicotinamide adenine dinucleotide [NAD+]). These biomolecules are key players required in a multitude of fundamental biochemical processes.72,73
First, let us examine SAM-e. A key methyl donor, SAM-e is one of the most frequently used enzyme substrates in human metabolism.74 Approximately 95% of SAM-e is used in methylation reactions that influence the activity of DNA, RNA, proteins, phospholipids, hormones, and transmitters. SAM-e methylation reactions result in the formation of S-adenosyl homocysteine, which can be hydrolyzed into homocysteine. The hypothesis that boron bioactivity occurs in part through an effect on SAM-e formation and/or use is supported by findings that plasma homocysteine increased and liver SAM-e decreased in rats fed 0.05 to 0.15 mg/kg of boron compared with rats supplemented with 3 mg/kg in their diets.75 High-circulating homocysteine and depleted SAM-e have been implicated in many of the disorders that can be beneficially affected by intakes of boron of greater than or equal to 3 mg/d, including arthritis, osteoporosis, cancer, diabetes, and impaired brain function.
Boron strongly binds oxidized NAD+,76 and, thus, might influence reactions in which NAD+ is involved, which include ATP production, calcium signaling, and the actions of the sirtuins, which are NAD-dependent deacetylases. Blocking NAD+ utilization, yet another beneficial action of boron, boosts NAD+ levels, activates sirtuins, and promotes healthy aging.77
Boron also forms boroester complexes with phosphoinositides, glycoproteins, and glycolipids. Glycolipids are biomolecules that act as calcium chelators and redox modifiers and affect cell-membrane integrity and function.78 Phosphoinositides are involved in the regulation of lipid kinases.79 Glycoproteins are important, integral membrane proteins, where they play a role in cell-to-cell interactions. Glycolipids, which are found on the outer surface of all eukaryotic cell membranes, extend from the phospholipid bilayer into the aqueous environment outside the cell and act as a recognition site for specific chemicals, help maintain the stability of the membrane, and attach cells to one another to form tissues. Through the boroester complexes formed with these biomolecules, boron impacts all the above actions.
A Possible Histone Deacetylase Inhibitor?
As research into the chemistry of boron-containing compounds has increased, they have been shown to be potent antiosteoporotic, anti-inflammatory, and antineoplastic agents both in vitro and in vivo. One potential explanation for the diversity in beneficial effects is that some boronated compounds are histone deacetylase inhibitors (HDIs).
HDIs are potential therapeutic agents for cancer and neurological diseases because of their abilities to alter gene expression, induce growth arrest or apoptosis of tumor cells, and stimulate normal cell differentiation. Several HDIs promote osteoblast maturation by enhancing RUNX2-dependent transcriptional activation, causing an increase in the expression levels of osteoblast-maturation genes, type 1 collagen, OPN, BSP, and OCN.80 What do all of these increases have in common? All have been shown, as noted earlier, to be activities of boron. HDIs are being considered as a new class of bone anabolic agents for the treatment of diseases associated with bone loss, such as osteoporosis and cancer.
Anticancer Effects
An increasing number of papers have indicated that boron possesses anticarcinogenic properties. Boron-rich diets and regions where the soil and water are rich in boron correlate with lower risks of several types of cancer, including prostate, breast, cervical, and lung cancers. Boron-enriched diets have been found to result in significant decreases in risk for prostate and cervical cancer and to decrease risk of lung cancer in smoking women. In the last few years, the use of natural and synthetic boron-containing compounds as anticancer agents has increased, particularly in inoperable cancers and those with high malignancy. Boron-containing compounds interfere with the physiology and reproduction of cancer cells through diverse mechanisms, including inhibition of serine proteases, NAD-dehydrogenases, mRNA splicing and cell division, receptor binding mimicry, and induction of apoptosis.81
Prostate Cancer.
Dietary boron is inversely correlated with prostate cancer incidence.82 According to National Health and Nutrition Examination Survey (NHANES) III data, risk of prostate cancer was 52% lower in men whose diets supplied more than 1.8 mg/d of boron compared with those whose dietary boron intake was less than or equal to 0.9 mg/d. High correlation (r = 0.63) was found between boron concentration in subsurface water and the distribution of prostate cancer in Texas.83 Analysis of NHANES III data also found that increased dietary intake of boron was associated with a decreased risk of prostate cancer, with a dose-response pattern. The adjusted odds ratio was 0.46 for the highest quartile of boron intake compared with the lowest quartile.82 No correlation with prostate cancer frequency has been observed when boron consumption was less than 1.17 mg/d.84
Boric acid inhibits human prostate cancer cell proliferation in vitro.85,86 In one study, boric acid decreased the size of prostate tumors in mice and markedly reduced levels of insulin-like growth factor 1 (IGF-1) in tumor tissue and of serum prostate specific antigen (PSA).87 In this study, 2 groups with 10 animals per group were dosed with boric-acid solutions (1.7, 9.0 mg boron/kg/d) by gavage. The control group received only water. Tumor sizes were measured weekly for 8 weeks. The size of tumors decreased in mice exposed to the low and high dose of boric acid by 38% and 25%, respectively. Serum PSA levels for the two dosages decreased by 88.6% and 86.4%, respectively, compared with the control group. In boron-dosed animals, a significantly lower incidence of mitotic figures, the term used in cellular pathology to describe the microscopic appearance of a cell undergoing mitosis, was seen. Circulating IGF-1 levels did not differ among groups, but expression of IGF-1 in the tumors was significantly reduced by boron treatment.
PSA is an androgen-regulated serine protease (enzyme) produced by both normal and cancerous prostate epithelial cells88 and is still the most commonly used serum marker for prostate cancer. Boronic acid has been shown to inhibit PSA activity.89,90
CLINICAL PEARL: Several modifications to PSA biomarker detection have been suggested to improve its sensitivity and specificity, including PSA density, free:total PSA, PSA velocity/doubling time, and different PSA isoforms. In addition, new biomarkers, including genetic and blood or urine based biomarkers, are now available. The most advanced is prostate cancer gene 3, which is found in urine and was developed into a commercial test in 2006.91
The IGF-1 signaling pathway promotes cancer progression; its downregulation is associated with lowered risk.92
Additional insight into the cellular mechanisms underlying boron’s antiprostate-cancer effects is provided by Barranco et al,93,94 whose work revealed that boric acid inhibits the growth of prostate-cancer cells both by decreasing expression of A-E cyclin and through boron’s influence on estrogen and testosterone. Cells treated with boric acid also showed decreased adhesion and migration, indicating lowered metastatic potential.21,95-97
It has been thought that high estradiol levels correlate with low prostate-cancer risks,97 a supposition that forms the basis of aromatase-inhibition therapy. Recently, however, papers have begun to appear that suggest estrogen’s role in the prostate is complicated by the differential actions of the estrogen receptors (ERs), ER-α and ER-β. Stimulation of ER-α promotes aberrant proliferation, inflammation, and premalignant pathology, whereas activation of ER-β has beneficial effects regarding cellular proliferation and plays a protective role against carcinogenesis.98 Research in this area is just beginning, but it appears that boron-containing compounds may beneficially modulate estrogen receptors, selectively binding to ER-β.99,100
Cervical Cancer.
Cervical cancer is the second most frequent cancer in women worldwide, yet in Turkey it ranks only ninth, which is 2 to 5 times lower than in Europe and North America.101 Although the reasons for this difference surely involve a combination of factors, including sociocultural differences, lack of population-based screening programs, or a lower human papillomavirus (HPV) prevalence rate in Turkey, it has been suggested that the low incidence of cervical cancer in Turkey correlates with its boron-rich soil.102 HPVs are the main cause of cervical cancer. HPV-16 and HPV-18 cause approximately 95% of all cervical cancers, and boron interferes in the life cycle of HPV.
Serine protease inhibitors reduce the immortalizing and transforming capacity of the HPV E7 oncogene.103-106 Boron exists in the human body mostly in the form of boric acid, a serine protease inhibitor. After conducting research that revealed incidence of cervical cancer-related histopathological findings correlated with boron-rich and boron-poor areas, Korkmaz et al107 suggested that higher amounts of boron in drinking water may help inhibit HPV transformation, reducing incidence of cervical cancer.
In that study, 107 incidence of adverse cytological findings in cervical smears was evaluated for 1059 women of low socioeconomic status who were living in boron-rich (472 women) and boron-poor (587 women) regions in Turkey. Mean dietary intake of boron was 8.41 mg/d for women from the boron-rich regions and 1.26 mg/d for women living in the boron-poor regions. None of the women from the boron-rich regions had any cytopathological indications of cervical cancer; cytopathological findings were present in 15 women from the boron-poor areas.
Lung Cancer.
Boron may have cancer-preventive actions similar to those of HRT, which is known to reduce lung cancer.108,109 A 10-year study (1995-2005), conducted at the University of Texas’s MD Anderson Cancer Center, in Houston, on the joint effects of boron intake and HRT use on lung-cancer risk, found that boron intake was inversely associated with lung cancer in women. Women whose intake of boron was low and who did not use HRT were at substantially increased risk. For all women, decreased boron intake was associated with increasing odds of lung cancer corresponding to a 39%, 64%, and 95% increase by decreasing quartile of intake. Compared with women with high dietary boron intake who used HRT, the odds ratio for lung cancer for low dietary boron intake and no HRT use was 2.07.110 One explanation proposed by the researchers for boron’s reduction in lung cancer risk was that postmenopausal women with high dietary boron intakes, as well as HRT users, have higher levels of estradiol competing for estrogen receptors with polycyclic aromatic hydrocarbons (PAHs) from cigarette smoke carcinogens. If correct, then increasing boron intake during HRT will also reduce the carcinogenic potential of PAHs from cigarette smoke.
Inhibition of Tumor-induced Angiogenesis.
Boronic acid, and a series of boron-containing phenoxyacetanilide derivatives, have also been shown to greatly inhibit hypoxia-inducible factor (HIF) 1.111 HIFs are heterodimeric (α/β) transcriptional factors and are major physiological stimuli for expression of angiogenesis factors. Angiogenesis, the formation of new blood vessels sprouting from existing host capillaries, must occur for tumors to grow beyond a certain critical size. Inhibition of tumor-induced angiogenesis prevents growth of many types of solid tumors and provides a novel approach for cancer treatment; thus, HIF-1 is a target of antineoplastic therapy.
Induction of Cancer-cell Apoptosis.
Sugar-borate esters act as boron vehicles, increasing the concentration of borate inside cancer cells relative to normal cells. Increased intracellular concentration of borate not only activates borate transporters but also leads to growth inhibition and apoptosis. In normal cells, the 2 latter, cell-destructive effects do not occur because the amount of borate present in a healthy diet, 1 to 10 mg/d, is easily exported from normal cells. Cancer cells, however, commonly overexpress sugar transporters and/or underexpress borate export, rendering sugar-borate esters as promising chemopreventive agents.112
Treatment of Multiple Myeloma and Non-Hodgkin’s Lymphoma.
Boron-based drugs are now being developed for use as therapeutic agents with anticancer, antiviral, antibacterial, antifungal, and other disease-specific activities. Bortezomib (marketed as Velcade by Millennium Pharmaceuticals and Cytomib by Venus Remedies), which contains boron as an active element, has been approved as a proteasome inhibitor for the treatment of multiple myeloma and non-Hodgkin’s lymphoma,113 and several other boron-based compounds are in various phases of clinical trials, with the expectation of better efficacy and potency than existing drugs.114
Lessening Side Effects of Traditional Chemotherapeutic Agents.
Traditional chemotherapeutic agents are cytotoxic (ie, they act by killing cells that divide rapidly), a key feature of most cancer cells. Unfortunately, other rapidly dividing cells, including cells produced in the bone marrow, are also destroyed, resulting in myelosuppression (ie, decreased production of blood cells, and, hence, a number of adverse effects, including immunosuppression).115 An in vitro study found that boric acid can help protect against genotoxicity and cytotoxicity that are induced in lymphocytes by paclitaxel, an anticancer drug commonly used to treat breast, ovarian, and lung cancers.116
Paclitaxel (10 or 20 μg/L) was added to human peripheral lymphocytes in blood cultures, and its genotoxic and cytotoxic effects were assessed by SCE and MN tests. Compared with controls, the frequencies of SCEs and the formation of MNs (ie, the genotoxic effects) were greatly increased, and the nuclear-division index (ie, the cytotoxic effect) was decreased in lymphocyte cultures exposed to paclitaxel. The addition of boric acid (2.5 or 5 mg/L) significantly diminished the paclitaxel-induced increases in genotoxicity, returning levels to close to control values and fully restoring cytotoxicity indices to control values.81
Life on Earth
Due to its importance in the so-called sugars world, boron may have played an essential role in the prebiotic origins of genetic material. Borates (ie, boron compounds such as boric acid) influence the formation of ribofuranose from formaldehyde, which feeds the prebiotic metabolic cycle. Borates, by forming complexes with organic cis-diols, are thought to have provided the thermal and chemical stability necessary for the development of life on Earth.117
The development of the RNA world, in which RNA acted as a catalyst as well as an informational macromolecule, assumes a large prebiotic (pre-RNA) source of ribose. However, the generally accepted prebiotic synthesis of ribose, the formose reaction, yields numerous sugars without any selectivity, and even if ribose were selectively synthesized, ribose and other sugars are unstable. Thus, it has been thought that the first genetic material could not have contained ribose or other sugars because of their instability.118
Nonetheless, during evolution, ribose was selected as the exclusive sugar component of nucleic acids. Ribose’s choice is explained by using molecular models to look at the chemical structure of aldopentoses (common sugars) and envisioning how they would fit in a nucleic-acid model. Comparisons indicate that ribose was not randomly selected but was the only choice because β-D-ribose best fits into the structure of the physiological forms of nucleic acids.119 Ribose is the only sugar present in both early RNA-based biochemistry and contemporary DNA-based life. Therefore, the stability of ribose is of fundamental concern for determining the origin of early RNA-based biochemistry. Borates stabilize ribose and can form borate-ester nucleotides.
Borate selectively increases the stability of ribose over all other aldopentoses. Without boron’s stabilizing influence, ribose is the least stable of all the aldopentoses produced by the formose reaction. A current theory is that ribose accumulation occurred in borate-rich environments in the pre-RNA world, setting the stage for the development of ribose-based nucleotides (RNA).120 This borate-complex formation might have sequestered ribose from the isomerization and decomposition reactions, resulting in its selective stabilization. These findings indicate that ribose could have accumulated in borate-rich environments on the early Earth and suggest that ribose-based nucleotides, combined with phosphate and nucleobases, formed abiotically.
Typical Intake of Boron/Supplemental Recommendations
Table 1 lists the boron content of its richest food sources. Unfortunately, the numbers are thought to be inaccurate, with an estimation of their being up to 3 to 4 times higher than the values found using chemical analysis.121
Table 1.
(Food Processor/Overestimated) Boron Content of Richest Food Sources
| Food | mg/100 g | mg in a Typical Serving |
|---|---|---|
| Avocado | 2.06 | 2.06 |
| Apricots (dried) | 2.11 | 0.53 |
| Currants | 1.74 | 0.26 |
| Grapes (red) | 0.50 | 0.50 |
| Peach | 0.52 | 0.57 |
| Prunes | 1.88 | 0.94 |
| Raisins | 4.51 | 0.67 |
| Red kidney beans | 1.4 | 1.82 |
| Lentils | 0.74 | 0.96 |
| Almonds | 2.82 | 0.42 |
| Brazil nutsa | 1.72 | 0.34 |
| Cashew nuts | 1.15 | 0.17 |
| Hazelnuts | 2.77 | 0.68 |
| Peanut butter | 1.92 | 0.38 |
| Pistachio nuts | 1.20 | 0.18 |
| Walnuts (California) | 1.63 | 0.24 |
| Wine (Shiraz Cabernet) | 0.86 | 0.86 |
Note: Adapted from Naghii et al.121
aBrazil nuts are the richest food source of selenium. Consumption of more than 2 Brazil nuts/d may result in selenium toxicity.
Meacham et al122,123 have shown that currently available computer software databases such as Food Processor (ESHA, Salem, OR, USA) greatly overestimate the boron content of foods. When a chemical analysis of foods was compared with analysis of dietary records in Food Processor, chemical analysis of the foodstuffs noted in the dietary records found 1.2 mg/d of boron, whereas Food Processor reported boron content as 4.5 mg/d (version 7.32), 5.0 mg/d (version 8.1), and 5.3 mg/d (version 9.9). Using chemical analysis, the researchers reported the boron content of the 10 foods richest in boron (Table 2).
Table 2.
Chemical Analysis of Boron Content (mg/100 g) in the Top 10 Foods
| Food | mg/100 g |
|---|---|
| Avocado | 1.43 |
| Peanut butter | 0.59 |
| Peanuts, dry | 0.58 |
| Prune juice | 0.56 |
| Chocolate powder | 0.43 |
| Red wine | 0.36 |
| Granola-raisin cereal | 0.36 |
| Grape juice | 0.34 |
| Pecans | 0.26 |
| Raisin bran | 0.26 |
Note: Adapted Meacham et al.122
Boron is part of the normal human diet, but daily intake varies widely depending on the proportions of various food groups in the diet and on the boron concentrations in the soil.124 Reported values for mean overall boron intake vary: 1.7 to 7.0 mg/d in the United States; 1.75 to 2.12 mg/d in Mexico; 0.8 to 1.9 mg/d in the European Union; 2.16 to 2.28 mg/d in Australia; and approximately 0.93 mg/d in Korea.125,126 The dissimilarities correlate with regional differences in the soil and consumption of fiber and protein-rich plant foods.
A diverse, plant-food-rich diet is estimated to provide approximately 1.5 to 3 mg/d of boron.125,127 Foods of plant origin, especially fruits, leafy vegetables, nuts, and legumes, are rich in boron as are plant-derived fermented beverages (ie, wine, cider, and beer). Meat, fish, and dairy products, however, are poor sources. Peanuts and peanut butter, other nuts, raisins, wine, and avocado are also top contributors to boron intake. Although coffee and milk are low in boron, they provide 12% of the total boron intake in the United States due to the volume consumed and to the fact that the standard American diet contains so few servings of fruits, vegetables, and legumes.125,128 No recommended levels have been set for boron, only an upper intake level (UL) of 20 mg/d.129
Although Rainey et al125 stated that “very large variations in boron intakes occur across the US adult population,” ranging as high as 9 mg/d, the researchers’ estimates of mean and 95th-percentile intakes of boron in the United States were 1.17 to 2.42 mg/d for men and 0.96 to 1.94 mg/d for women. It is important to note that the US population is consuming fewer boron-rich plant foods today than in 1998, when the research conducted by Rainey et al125 was done. It seems likely that average boron intake has dropped.
In 1988 to 1994, only 27% of adults met the USDA guidelines for fruit—2 or more servings, and only 35% met the guidelines for vegetables—3 or more servings; in 1999 to 2002, 28% and 32% of adults met fruit and vegetable guidelines, respectively. Only 11% met USDA guidelines for both fruits and vegetables in 1988 to 1994 and 1999 to 2002, indicating no change in consumption.130 By 1999 to 2002, only 40% ± 2% of the US population met the then-current recommendation to eat an average of 5 or more servings of fruits and vegetables per day. Among subpopulations, the percentages consuming at least that much ranged from a low of 10% ± 3% of girls aged 4 to 8 years to a high of 60% ± 4% of men aged 51 to 70 years.
As of 2006, the new combined recommended amounts of fruits and vegetables ranged from 2 to 6.5 cups. Assuming 2 servings per cup, the proportions of the subpopulations meeting the combined new recommendations for fruits and vegetables are estimated to range from a low of 0.7% ± 0.4% of boys aged 14 to 18 years, whose combined recommendation is 5 cups, to a high of 48% ± 4% of children aged 2 to 3 years, whose combined recommendation is 1 cup. Only 17% ± 3% of women aged 51 to 70 years meet their combined recommendation, and all other sex–age groups have fewer than 11% meeting them.131
Human requirements for boron remain undefined. The only guideline is a tolerable UL, which for adults aged 18 years or older is approximately 20 mg/d.132 The current author hopes that increasing knowledge about boron’s diverse biological functions will promote recognition of the importance of specifying what constitutes adequate, and, even possibly, a consideration of what might be optimal boron intake. Although it has been suggested that humans need at least 0.2 mg/d of boron and that the diet should contain approximately 1 to 2 mg/d of boron, numerous papers indicate that the standard American diet does not do so, and boron’s beneficial effects on bone,133 sex steroids,20 and vitamin D26 do not appear at boron intakes of less than 3 mg/d. All the human studies showing beneficial effects for boron supplementation discussed in this review used supplemental boron at a dose of greater than or equal to 3 mg/d. Studies of residents in boron-rich areas of the world indicated that 3 mg/d is a conservative amount that would produce benefits with a very remote risk of adverse effects. For example, in Turkey, daily boron intake for workers in a boric acid production plant averaged 12.6 mg/d, with no adverse effects.134,135
Conclusions
Boron has been proven to be an important trace mineral because it (1) is essential for the growth and maintenance of bone; (2) greatly improves wound healing; (3) beneficially impacts the body’s use of estrogen, testosterone, and vitamin D; (4) boosts magnesium absorption; (5) reduces levels of inflammatory biomarkers, such as hs-CRP and TNF-α; (6) raises levels of antioxidant enzymes, such as SOD, catalase, and glutathione peroxidase; (7) protects against pesticide-induced oxidative stress and heavy-metal toxicity; (8) improves brain electrical activity, cognitive performance, and short-term memory in elders; (9) influences the formation and activity of key biomolecules, such as SAM-e and NAD+; (10) has demonstrated preventive and therapeutic effects in a number of cancers, such as prostate, cervical, and lung cancers and multiple and non-Hodgkin’s lymphoma; and (11) may help ameliorate the adverse effects of traditional chemotherapeutic agents. Americans’ daily dietary intake of boron was estimated to be approximately 1 mg/d in 1999.
In none of the numerous studies conducted to date, however, do boron’s beneficial effects appear at intakes of less than 3 mg/d. No EARs or DRIs have been set for boron; only a UL of 20 mg/d for individuals aged 18 years or older. The absence of studies showing harm in conjunction with the substantial number of articles showing benefits support the consideration of boron supplementation of 3 mg/d for any individual who is consuming a diet low in fruits and vegetables or who is at risk for or has osteopenia; osteoporosis; OA; or breast, prostate, or lung cancer.
Figure 1.
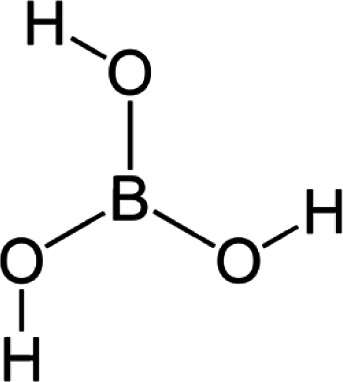
Structure of boric acid.
References
- 1.Nielsen FH, Hunt CD, Mullen LM, Hunt JR. Effect of dietary boron on mineral, estrogen, and testosterone metabolism in postmenopausal women. FASEB J. 1987;1(5):394-397. [PubMed] [Google Scholar]
- 2.Nielsen FH. Is boron nutritionally relevant? Nutr Rev. 2008;66(4):183-191. [DOI] [PubMed] [Google Scholar]
- 3.Beattie JH, Peace HS. The influence of a low-boron diet and boron supplementation on bone, major mineral and sex steroid metabolism in postmenopausal women. Br J Nutr. 1993;69(3):871-884. [DOI] [PubMed] [Google Scholar]
- 4.Hunt CD. The biochemical effects of physiologic amounts of dietary boron in animal nutrition models. Environ Health Perspect. 1994;102(suppl 7):35-43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Gorustovich AA, Steimetz T, Nielsen FH, Guglielmotti MB. A histomorphometric study of alveolar bone modelling and remodeling in mice fed a boron-deficient diet. Arch Oral Biol. 2008;53(7):677-682. [DOI] [PubMed] [Google Scholar]
- 6.Nielsen FH, Stoecker BJ. Boron and fish oil have different beneficial effects on strength and trabecular microarchitecture of bone. J Trace Elem Med Biol. 2009;23(3):195-203. [DOI] [PubMed] [Google Scholar]
- 7.Hakki SS, Bozkurt BS, Hakki EE. Boron regulates mineralized tissue-associated proteins in osteoblasts (MC3T3-E1). J Trace Elem Med Biol. 2010;24(4):243-250. [DOI] [PubMed] [Google Scholar]
- 8.Blech MF, Martin C, Borrelly J, Hartemann P. Treatment of deep wounds with loss of tissue: value of a 3 percent boric acid solution [in French]. Presse Med. 1990;19(22):1050-1052. [PubMed] [Google Scholar]
- 9.Benderdour M, Van Bui T, Hess K, Dicko A, Belleville F, Dousset B. Effects of boron derivatives on extracellular matrix formation. J Trace Elem Med Biol. 2000;14(3):168-173. [DOI] [PubMed] [Google Scholar]
- 10.Nzietchueng RM, Dousset B, Franck P, Benderdour M, Nabet P, Hess K. Mechanisms implicated in the effects of boron on wound healing. J Trace Elem Med Biol. 2002;16(4):239-244. [DOI] [PubMed] [Google Scholar]
- 11.Ying X, Cheng S, Wang W, et al. Effect of boron on osteogenic differentiation of human bone marrow stromal cells. Biol Trace Elem Res. 2011;144(1-3):306-315. [DOI] [PubMed] [Google Scholar]
- 12.Rosen V, Wozney JM. Bone morphogenetic proteins. In: Bilezikian JP, Raisz LG, Rodan GA, eds. Principles of Bone Biology. Vol 2. 2nd ed. San Diego, CA: Academic Press; 2002:919-928. [Google Scholar]
- 13.Chen D, Zhao M, Mundy GR. Bone morphogenetic proteins. Growth Factors. 2004;22(4):233-241. [DOI] [PubMed] [Google Scholar]
- 14.Lavery K, Swain P, Falb D, Alaoui-Ismaili MH. BMP-2/4 and BMP-6/7 differentially utilize cell surface receptors to induce osteoblastic differentiation of human bone marrow-derived mesenchymal stem cells. J Biol Chem. 2008;283(30):20948-20958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Martinovic S, Borovecki F, Miljavac V, et al. Requirement of a bone morphogenetic protein for the maintenance and stimulation of osteoblast differentiation. Arch Histol Cytol. 2006;69(1):23-36. [DOI] [PubMed] [Google Scholar]
- 16.Phimphilai M, Zhao Z, Boules H, Roca H, Franceschi RT. BMP signaling is required for RUNX2-dependent induction of the osteoblast phenotype. J Bone Miner Res. 2006;21(4):637-646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Franceschi RT, Ge C, Xiao G, Roca H, Jiang D. Transcriptional regulation of osteoblasts. Cells Tissues Organs. 2009;189(1-4):144-152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Ducy P, Starbuck M, Priemel M, et al. A Cbfa1-dependent genetic pathway controls bone formation beyond embryonic development. Genes Dev. 1999;13(8):1025-1036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Gersbach CA, Byers BA, Pavlath GK, García AJ. Runx2/Cbfa1 stimulates transdifferentiation of primary skeletal myoblasts into a mineralizing osteoblastic phenotype. Exp Cell Res. 2004;300(2):406-417. [DOI] [PubMed] [Google Scholar]
- 20.Naghii MR, Mofid M, Asgari AR, Hedayati M, Daneshpour MS. Comparative effects of daily and weekly boron supplementation on plasma steroid hormones and proinflammatory cytokines. J Trace Elem Med Biol. 2011;25(1):54-58. [DOI] [PubMed] [Google Scholar]
- 21.Naghii MR, Samman S. The effect of boron supplementation on its urinary excretion and selected cardiovascular risk factors in healthy male subjects. Biol Trace Elem Res. 1997;56(3):273-286. [DOI] [PubMed] [Google Scholar]
- 22.Morgentaler A. Testosterone for Life: Recharge Your Vitality, Sex Drive, Muscle Mass, and Overall Health. New York, NY: McGraw-Hill; 2009:65. [Google Scholar]
- 23.Chueh KS, Huang SP, Lee YC, et al. The comparison of the aging male symptoms (AMS) scale and androgen deficiency in the aging male (ADAM) questionnaire to detect androgen deficiency in middle-aged men. J Androl. 2012;33(5):817-823. [DOI] [PubMed] [Google Scholar]
- 24.Dupre JN, Keenan MJ, Hegsted M, Brudevold AM. Effects of dietary boron in rats fed a vitamin D-deficient diet. Environ Health Perspect. 1994;102(suppl 7):55-58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Nielsen FH, Mullen LM, Gallegher SK. Effect of boron depletion and repletion on blood indicators of calcium status in humans fed a magnesium-low diet. J Trace Elem Exp Med. 1990;3:45-54. [Google Scholar]
- 26.Miljkovic D, Scorei RI, Cimpoiaşu VM, Scorei ID. Calcium fructoborate: plant-based dietary boron for human nutrition. J Diet Suppl. 2009;6(3):211-226. [DOI] [PubMed] [Google Scholar]
- 27.Miljkovic D, Miljkovic N, McCarty MF. Up-regulatory impact of boron on vitamin D function—does it reflect inhibition of 24-hydroxylase? Med Hypotheses. 2004;63(6):1054-1056. [DOI] [PubMed] [Google Scholar]
- 28.Nuklear. Estradiol Synthesis. Licensed under CC BY-SA 4.0 via Wikimedia Commons, work by Lara Pizzorno, http://commons.wikimedia.org/wiki/File:Estradiol_synthesis.svg#mediaviewer/File:Estradiol_synthesis.svg. Accessed July 8, 2015.
- 29.Henry HL. The 25(OH)D(3)/1alpha,25(OH)(2)D(3)-24R-hydroxylase: a catabolic or biosynthetic enzyme? Steroids. 2001;66(3-5):391-398. [DOI] [PubMed] [Google Scholar]
- 30.DeLuca HF. Is there more to learn about functional vitamin D metabolism [published online ahead of print September 4, 2014]? J Steroid Biochem Mol Biol. April 2015;148:3-6. [DOI] [PubMed] [Google Scholar]
- 31.Stubbs JR, Zhang S, Friedman PA, Nolin TD. Decreased conversion of 25-hydroxyvitamin D3 to 24,25-dihydroxyvitamin D3 following cholecalciferol therapy in patients with CKD. Clin J Am Soc Nephrol. 2014;9(11):1965-1973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Zhou Y, Zhao LJ, Xu X, et al. DNA methylation levels of CYP2R1 and CYP24A1 predict vitamin D response variation. J Steroid Biochem Mol Biol. 2014;144 Pt A:207-214. doi: 10.1016/j.jsbmb.2013.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Silberberg MS. e-Study Guide for Chemistry: The Molecular Nature of Matter and Change. ISBN:9780077216504 (Google eBook). [Google Scholar]
- 34.Jones G, Prosser DE, Kaufmann M. 25-Hydroxyvitamin D-24-hydroxylase (CYP24A1): its important role in the degradation of vitamin D. Arch Biochem Biophys. 2012;523(1):9-18. [DOI] [PubMed] [Google Scholar]
- 35.Zofková I, Nemcikova P, Matucha P. Trace elements and bone health. Clin Chem Lab Med. 2013;51(8):1555-1561. [DOI] [PubMed] [Google Scholar]
- 36.Nielsen FH. The emergence of boron as nutritionally important throughout the life cycle. Nutrition. 2000;16(7-8):512-514. [DOI] [PubMed] [Google Scholar]
- 37.Armstrong TA, Spears JW. Effect of boron supplementation of pig diets on the production of tumor necrosis factor-alpha and interferon-gamma. J Anim Sci. 2003;81(10):2552-2561. [DOI] [PubMed] [Google Scholar]
- 38.Wang G, Li N, Chang S, et al. A prospective follow-up study of the relationship between C-reactive protein and human cancer risk in the Chinese Kailuan female cohort [published online ahead of print December 9, 2014]. Cancer Epidemiol Biomarkers Prev. 2015;24(2):459-465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Chen F, Wang W, Teng Y, et al. Relationship between high-sensitivity C-reactive protein and obesity/metabolic syndrome in children [in Chinese]. Zhonghua Liu Xing Bing Xue Za Zhi. 2014;35(6):621-625. [PubMed] [Google Scholar]
- 40.Kaur R, Matharoo K, Sharma R, Bhanwer AJ. C-reactive protein + 1059 G>C polymorphism in type 2 diabetes and coronary artery disease patients. Meta Gene. November 2013;1:82-92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Svensson E, Mor A, Rungby J, et al. Lifestyle and clinical factors associated with elevated C-reactive protein among newly diagnosed type 2 diabetes mellitus patients: a cross-sectional study from the nationwide DD2 cohort. BMC Endocr Disord. August 2014;14:74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Hamirani YS, Katz R, Nasir K, et al. Association between inflammatory markers and liver fat: the multi-ethnic study of atherosclerosis. J Clin Exp Cardiolog. 2014;5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Zhao X, Luo J, Li B, Liu S, Li D. The association between preoperative serum C-reactive protein and hepatocellular carcinoma recurrence in patients with chronic hepatitis B virus (HBV) infection—a retrospective study. PLoS One. 2015;10(1):e0116909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Rocha P, Morgan CJ, Templeton AJ, Pond GR, Naik G, Sonpavde G. Prognostic impact of C-reactive protein in metastatic prostate cancer: a systematic review and meta-analysis. Oncol Res Treat. 2014;37(12):772-776. [DOI] [PubMed] [Google Scholar]
- 45.Jin Y, Sun Y, Shi X, Zhao J, Shi L, Yu X. Prognostic value of circulating C-reactive protein levels in patients with non-small cell lung cancer: a systematic review with meta-analysis. J Cancer Res Ther. 2014;10(suppl):C160-C166. [DOI] [PubMed] [Google Scholar]
- 46.Au B, Smith KJ, Gariépy G, Schmitz N. C-reactive protein, depressive symptoms, and risk of diabetes: results from the English Longitudinal Study of Ageing (ELSA). J Psychosom Res. 2014;77(3):180-186. [DOI] [PubMed] [Google Scholar]
- 47.Khandaker GM, Pearson RM, Zammit S, Lewis G, Jones PB. Association of serum interleukin 6 and C-reactive protein in childhood with depression and psychosis in young adult life: a population-based longitudinal study. JAMA Psychiatry. 2014;71(10):1121-1128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kaptoge S, Di Angelantonio E, Lowe G, et al. Emerging Risk Factors Collaboration . C-reactive protein concentration and risk of coronary heart disease, stroke, and mortality: an individual participant meta-analysis. Lancet. 2010;375(9709):132-140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Danesh J, Kaptoge S, Mann AG, et al. Long-term interleukin-6 levels and subsequent risk of coronary heart disease: two new prospective studies and a systematic review. PLoS Med. 2008;5(4):e78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Arkhipova SV, Zorin NA, Iankin Mlu, et al. Cytokine and acute phase inflammation reactant levels in men with myocardial infarction [in Russian]. Klin Med (Mosk). 2009;87(12):20-23. [PubMed] [Google Scholar]
- 51.Gustafson B. Adipose tissue, inflammation and atherosclerosis. J Atheroscler Thromb. 2010;17(4):332-341. [DOI] [PubMed] [Google Scholar]
- 52.Newnham RE. Essentiality of boron for healthy bones and joints. Environ Health Perspect. 1994;102(suppl 7):83-85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Newnham R. Discovering the cure for arthritis. Nutr Health. 2004;17(4):281-284. [DOI] [PubMed] [Google Scholar]
- 54.Scorei R, Mitrut P, Petrisor I, Scorei I. A double-blind, placebo-controlled pilot study to evaluate the effect of calcium fructoborate on systemic inflammation and dyslipidemia markers for middle-aged people with primary osteoarthritis. Biol Trace Elem Res. 2011;144(1-3):253-263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Korkmaz M, Sayli U, Sayli BS, et al. Estimation of human daily boron exposure in a boron-rich area. Br J Nutr. 2007;98(3):571-575. [DOI] [PubMed] [Google Scholar]
- 56.Helliwell TR, Kelly SA, Walsh HP, et al. Elemental analysis of femoral bone from patients with fractured neck of femur or osteoarthrosis. Bone. 1996;18(2):151-157. [DOI] [PubMed] [Google Scholar]
- 57.Hunt CD, Idso JP. Dietary boron as a physiological regulator of the normal inflammatory response: a review and current research progress. J Trace Elem Exp Med. 1999;12(3):221-233. [Google Scholar]
- 58.Nielsen FH. Biochemical and physiologic consequences of boron deprivation in humans. Environ Health Perspect. 1994;102(suppl 7):59-63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Castiglioni S, Cazzaniga A, Albisetti W, Maier JA. Magnesium and osteoporosis: current state of knowledge and future research directions. Nutrients. 2013;5(8):3022-3033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.García-Hernández A, Arzate H, Gil-Chavarría I, Rojo R, Moreno-Fierros L. High glucose concentrations alter the biomineralization process in human osteoblastic cells. Bone. 2012;50(1):276-288. [DOI] [PubMed] [Google Scholar]
- 61.Travers RL, Rennie GC, Newnham RE. Boron and arthritis: the results of a double-blind pilot study. J Nutr Environ Med. 1990;1(2):127-132. [Google Scholar]
- 62.Scorei RI, Ciofrangeanu C, Ion R, et al. In vitro effects of calcium fructoborate upon production of inflammatory mediators by LPS-stimulated RAW 264.7 macrophages. Biol Trace Elem Res. 2010;135(1-3):334-344. [DOI] [PubMed] [Google Scholar]
- 63.Nakamura K, Saito T, Kobayashi R, et al. C-reactive protein predicts incident fracture in community-dwelling elderly Japanese women: the Muramatsu study. Osteoporos Int. 2011;22(7):2145-2150. [DOI] [PubMed] [Google Scholar]
- 64.Scorei RI, Rotaru P. Calcium fructoborate—potential anti-inflammatory agent. Biol Trace Elem Res. 2011;143(3):1223-1238. [DOI] [PubMed] [Google Scholar]
- 65.Pietrzkowski Z, Phelan MJ, Keller R, Shu C, Argumedo R, Reyes-Izquierdo T. Short-term efficacy of calcium fructoborate on subjects with knee discomfort: a comparative, double-blind, placebo-controlled clinical study. Clin Interv Aging. June 2014;9:895-899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Wolfe F. The C-reactive protein but not erythrocyte sedimentation rate is associated with clinical severity in patients with osteoarthritis of the knee or hip. J Rheumatol. 1997;24(8):1486-1488. [PubMed] [Google Scholar]
- 67.Pearle AD, Scanzello CR, George S, et al. Elevated high-sensitivity C-reactive protein levels are associated with local inflammatory findings in patients with osteoarthritis. Osteoarthritis Cartilage. 2007;15(5):516-523. [DOI] [PubMed] [Google Scholar]
- 68.Scorei ID, Scorei RI. Calcium fructoborate helps control inflammation associated with diminished bone health. Biol Trace Elem Res. 2013;155(3):315-321. [DOI] [PubMed] [Google Scholar]
- 69.Coban FK, Ince S, Kucukkurt I, Demirel HH, Hazman O. Boron attenuates malathion-induced oxidative stress and acetylcholinesterase inhibition in rats [published online ahead of print October 24, 2014]. Drug Chem Toxicol. [DOI] [PubMed] [Google Scholar]
- 70.Penland JG. The importance of boron nutrition for brain and psychological function. Biol Trace Elem Res. 1998;66(1-3):299-317. [DOI] [PubMed] [Google Scholar]
- 71.Turkez H, Geyikoglu F, Tatar A, Keles MS, Kaplan I. The effects of some boron compounds against heavy metal toxicity in human blood. Exp Toxicol Pathol. 2012;64(1-2):93-101. [DOI] [PubMed] [Google Scholar]
- 72.Nielsen FH. Update on human health effects of boron. J Trace Elem Med Biol. 2014;28(4):383-387. [DOI] [PubMed] [Google Scholar]
- 73.Bolaños L, Lukaszewski K, Bonilla I, Blevins D. Why boron? Plant Physiol Biochem. 2004;42(11):907-912. [DOI] [PubMed] [Google Scholar]
- 74.Loenen WA. S-adenosylmethionine: jack of all trades and master of everything? Biochem Soc Trans. April 2006;34(pt 2):330-333. [DOI] [PubMed] [Google Scholar]
- 75.Nielsen FH. Boron deprivation decreases liver S-adenosylmethionine and spermidine and increases plasma homocysteine and cysteine in rats. J Trace Elem Med Biol. 2009;23(3):204-213. [DOI] [PubMed] [Google Scholar]
- 76.Ralston NV, Hunt CD. Diadenosine phosphates and S-adenosylmethionine: novel boron binding biomolecules detected by capillary electrophoresis. Biochim Biophys Acta. 2001;1527(1-2):20-30. [DOI] [PubMed] [Google Scholar]
- 77.Mouchiroud L, Houtkooper RH, Auwerx J. NAD+ metabolism: a therapeutic target for age-related metabolic disease. Crit Rev Biochem Mol Biol. 2013;48(4):397-408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Wimmer MA, Lochnit G, Bassil E, Mühling KH, Goldbach HE. Membrane-associated, boron-interacting proteins isolated by boronate affinity chromatography. Plant Cell Physiol. 2009;50(7):1292-1304. [DOI] [PubMed] [Google Scholar]
- 79.Grabon A, Khan D, Bankaitis VA. Phosphatidylinositol transfer proteins and instructive regulation of lipid kinase biology [published online ahead of print January 12, 2015]. Biochim Biophys Acta. doi:10.1016/j.bbalip.2014.12.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Schroeder TM, Westendorf JJ. Histone deacetylase inhibitors promote osteoblast maturation. J Bone Miner Res. 2005;20(12):2254-2263. [DOI] [PubMed] [Google Scholar]
- 81.Scorei RI, Popa R, Jr. Boron-containing compounds as preventive and chemotherapeutic agents for cancer. Anticancer Agents Med Chem. 2010;10(4):346-351. [DOI] [PubMed] [Google Scholar]
- 82.Cui Y, Winton MI, Zhang ZF, et al. Dietary boron intake and prostate cancer risk. Oncol Rep. 2004;11(4):887-892. [PubMed] [Google Scholar]
- 83.Barranco WT, Hudak PF, Eckhert CD. Evaluation of ecological and in vitro effects of boron on prostate cancer risk (United States). Cancer Causes Control. 2007;18(1):71-77. [DOI] [PubMed] [Google Scholar]
- 84.Gonzalez A, Peters U, Lampe JW, White E. Boron intake and prostate cancer risk. Cancer Causes Control. 2007;18(10):1131-1140. [DOI] [PubMed] [Google Scholar]
- 85.Gallardo-Williams MT, Maronpot RR, Wine RN, Brunssen SH, Chapin RE. Inhibition of the enzymatic activity of prostate-specific antigen by boric acid and 3-nitrophenyl boronic acid. Prostate. 2003;54(1):44-49. [DOI] [PubMed] [Google Scholar]
- 86.Barranco WT, Eckhert CD. Boric acid inhibits human prostate cancer cell proliferation. Cancer Lett. 2004;216(1):21-29. [DOI] [PubMed] [Google Scholar]
- 87.Gallardo-Williams MT, Chapin RE, King PE, et al. Boron supplementation inhibits the growth and local expression of IGF-1 in human prostate adenocarcinoma (LNCaP) tumors in nude mice. Toxicol Pathol. 2004;32(1):73-78. [DOI] [PubMed] [Google Scholar]
- 88.Balk SP, Ko YJ, Bubley GJ. Biology of prostate-specific antigen. J Clin Oncol. 2003;21(2):383-391. [DOI] [PubMed] [Google Scholar]
- 89.Smoum R, Rubinstein A, Dembitsky VM, Srebnik M. Boron containing compounds as protease inhibitors. Chem Rev. 2012;112(7):4156-4220. [DOI] [PubMed] [Google Scholar]
- 90.Diederich WE, Wegscheid-Gerlach C. The medicinal chemistry of protease inhibitors. Curr Top Med Chem. 2010;10(3):231. [DOI] [PubMed] [Google Scholar]
- 91.Artibani W. Landmarks in prostate cancer diagnosis: the biomarkers. BJU Int. 2012;110(suppl 1):8-13. [DOI] [PubMed] [Google Scholar]
- 92.Arnaldez F, Helman LJ. Targeting the insulin growth factor receptor 1. Hematol Oncol Clin North Am. 2012;26(3):527-542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Barranco WT, Eckhert CD. Cellular changes in boric acid-treated DU-145 prostate cancer cells. Br J Cancer. 2006;94(6):884-890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Barranco WT, Kim DH, Stella SL, Jr, Eckhert CD. Boric acid inhibits stored Ca2+ release in DU-145 prostate cancer cells. Cell Biol Toxicol. 2009;25(4):309-320. [DOI] [PubMed] [Google Scholar]
- 95.Black A, Pinsky PF, Grubb RL, III, et al. Sex steroid hormone metabolism in relation to risk of aggressive prostate cancer. Cancer Epidemiol Biomarkers Prev. 2014;23(11):2374-2382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Carruba G. Estrogen and prostate cancer: an eclipsed truth in an androgendominated scenario. J Cell Biochem. 2007;102(4):899-911. [DOI] [PubMed] [Google Scholar]
- 97.Gann PH, Hennekens CH, Ma J, Longcope C, Stampfer MJ. Prospective study of sex hormone levels and risk of prostate cancer. J Natl Cancer Inst. 1996;88(16):1118-1126. [DOI] [PubMed] [Google Scholar]
- 98.Ellem SJ, Risbridger GP. Aromatase and regulating the estrogen:androgen ratio in the prostate gland. J Steroid Biochem Mol Biol. 2010;118(4-5):246-251. [DOI] [PubMed] [Google Scholar]
- 99.Hirata M, Inada M, Matsumoto C, et al. A novel carborane analog, BE360, with a carbon-containing polyhedral boron-cluster is a new selective estrogen receptor modulator for bone. Biochem Biophys Res Commun. 2009;380(2):218-222. [DOI] [PubMed] [Google Scholar]
- 100.Ohta K, Ogawa T, Kaise A, Endo Y. Enhanced estrogen receptor beta (ERβ) selectivity of fluorinated carborane-containing ER modulators. Bioorg Med Chem Lett. 2013;23(24):6555-6558. [DOI] [PubMed] [Google Scholar]
- 101.Turkish Cervical Cancer and Cervical Cytology Research Group. Prevalence of cervical cytological abnormalities in Turkey. Int J Gynaecol Obstet. 2009;106(3):206-209. [DOI] [PubMed] [Google Scholar]
- 102.Simsek A, Velioglu YS, Coskun AL, Sayli BS. Boron concentrations in selected foods from borate-producing regions in Turkey. J Sci Food Agric. 2003;83(6):586-592. [Google Scholar]
- 103.Drubin DA, McLaughlin-Drubin ME, Clawson GA, Meyers C. A protease inhibitor specifically inhibits growth of HPV-infected keratinocytes. Mol Ther. 2006;13(6):1142-1148. [DOI] [PubMed] [Google Scholar]
- 104.Stöppler H, Stöppler MC, Adduci A, Koval D, Schlegel R. The serine protease inhibitors TLCK and TPCK react with the RB-binding core of HPV-18 E7 protein and abolish its RB-binding capability. Virology. 1996;217(2):542-553. [DOI] [PubMed] [Google Scholar]
- 105.Stöppler H, Koval D, Schlegel R. The serine protease inhibitors TLCK and TPCK inhibit the in vitro immortalization of primary human keratinocytes by HPV-18 DNA. Oncogene. 1996;13(7):1545-1548. [PubMed] [Google Scholar]
- 106.Wise-Draper TM, Wells SI. Papillomavirus E6 and E7 proteins and their cellular targets. Front Biosci. January 2008;13:1003-1017. [DOI] [PubMed] [Google Scholar]
- 107.Korkmaz M, Uzgören E, Bakirdere S, Aydin F, Ataman OY. Effects of dietary boron on cervical cytopathology and on micronucleus frequency in exfoliated buccal cells. Environ Toxicol. 2007;22(1):17-25. [DOI] [PubMed] [Google Scholar]
- 108.Pesatori AC, Carugno M, Consonni D, et al. Hormone use and risk for lung cancer: a pooled analysis from the International Lung Cancer Consortium (ILCCO). Br J Cancer. 2013;109(7):1954-1964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Yao Y, Gu X, Zhu J, Yuan D, Song Y. Hormone replacement therapy in females can decrease the risk of lung cancer: a meta-analysis. PLoS One. 2013;8(8):e71236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Mahabir S, Spitz MR, Barrera SL, Dong YQ, Eastham C, Forman MR. Dietary boron and hormone replacement therapy as risk factors for lung cancer in women. Am J Epidemiol. 2008;167(9):1070-1080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Shimizu K, Maruyama M, Yasui Y, Minegishi H, Ban HS, Nakamura H. Boron-containing phenoxyacetanilide derivatives as hypoxia-inducible factor (HIF)-1alpha inhibitors. Bioorg Med Chem Lett. 2010;20(4):1453-1456. [DOI] [PubMed] [Google Scholar]
- 112.Scorei RI, Popa R. Sugar-borate esters—potential chemical agents in prostate cancer chemoprevention. Anticancer Agents Med Chem. 2013;13(6):901-909. [DOI] [PubMed] [Google Scholar]
- 113.Genadieva-Stavric S, Cavallo F, Palumbo A. New approaches to management of multiple myeloma. Curr Treat Options Oncol. 2014;15(2):157-170. [DOI] [PubMed] [Google Scholar]
- 114.Das BC, Thapa P, Karki R, et al. Boron chemicals in diagnosis and therapeutics. Future Med Chem. 2013;5(6):653-676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Javeed A, Ashraf M, Riaz A, Ghafoor A, Afzal S, Mukhtar MM. Paclitaxel and immune system. Eur J Pharm Sci. 2009;38(4):283-290. [DOI] [PubMed] [Google Scholar]
- 116.Turkez H, Tatar A, Hacimuftuoglu A, Ozdemir E. Boric acid as a protector against paclitaxel genotoxicity. Acta Biochim Pol. 2010;57(1):95-97. [PubMed] [Google Scholar]
- 117.Scorei R. Is boron a prebiotic element? A mini-review of the essentiality of boron for the appearance of life on earth. Orig Life Evol Biosph. 2012;42(1):3-17. [DOI] [PubMed] [Google Scholar]
- 118.Larralde R, Robertson MP, Miller SL. Rates of decomposition of ribose and other sugars: implications for chemical evolution. Proc Natl Acad Sci U S A. 1995;92(18):8158-8160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Banfalvi G. Why ribose was selected as the sugar component of nucleic acids. DNA Cell Biol. 2006;25(3):189-196. [DOI] [PubMed] [Google Scholar]
- 120.Furukawa Y, Horiuchi M, Kakegawa T. Selective stabilization of ribose by borate. Orig Life Evol Biosph. 2013;43(4-5):353-361. [DOI] [PubMed] [Google Scholar]
- 121.Naghii MR, Wall PM, Samman S. The boron content of selected foods and the estimation of its daily intake among free-living subjects. J Am Coll Nutr. 1996;15(6):614-619. [DOI] [PubMed] [Google Scholar]
- 122.Meacham S, Karakas S, Wallace A, Altun F. Boron in human health: evidence for dietary recommendations and public policies. Open Miner Process J. 2010;3(1):36-53. [Google Scholar]
- 123.Naghii MR, Samman S. The effect of boron on plasma testosterone and plasma lipids in rats. Nutr Res. 1997;17(3):523-531. [Google Scholar]
- 124.Nielsen FH. Boron—an overlooked element of potential nutritional importance. Nutr Today. 1988;23(1):4-7. [Google Scholar]
- 125.Rainey C, Nyquist L. Multicountry estimation of dietary boron intake. Biol Trace Elem Res. 1998;66(1-3):79-86. [DOI] [PubMed] [Google Scholar]
- 126.Kim MH, Bae YJ, Lee YS, Choi MK. Estimation of boron intake and its relation with bone mineral density in free-living Korean female subjects. Biol Trace Elem Res. 2008;125(3):213-222. [DOI] [PubMed] [Google Scholar]
- 127.Richold M. Boron exposure from consumer products. Biol Trace Elem Res. 1998;66(1-3):121-129. [DOI] [PubMed] [Google Scholar]
- 128.Rainey CJ, Nyquist LA, Christensen RE, Strong PL, Culver BD, Coughlin JR. Daily intake of boron from the American diet. J Am Diet Assoc. 1999;99(3):335-340. [DOI] [PubMed] [Google Scholar]
- 129.Panel on Micronutrients, Subcommittees on Upper Reference Levels of Nutrients and of Interpretation and Uses of Dietary Reference Intakes; Standing Committee on the Scientific Evaluation of Dietary Reference Intakes; Food and Nutrition Board; Institute of Medicine. Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc. Washington, DC: National Academy Press; 2002. [Google Scholar]
- 130.Casagrande SS, Wang Y, Anderson C, Gary TL. Have Americans increased their fruit and vegetable intake? The trends between 1988 and 2002. Am J Prev Med. 2007;32(4):257-263. [DOI] [PubMed] [Google Scholar]
- 131.Guenther PM, Dodd KW, Reedy J, Krebs-Smith SM. Most Americans eat much less than recommended amounts of fruits and vegetables. J Am Diet Assoc. 2006;106(9):1371-1379. [DOI] [PubMed] [Google Scholar]
- 132.Food and Nutrition Board. Institute of Medicine. National Academie, dietary reference intakes (DRIs): Recommended intakes for individuals, vitamins, 2001. [Google Scholar]
- 133.Meacham SL, Taper LJ, Volpe SL. Effects of boron supplementation on bone mineral density and dietary, blood, and urinary calcium, phosphorus, magnesium, and boron in female athletes. Environ Health Perspect. 1994;102(suppl 7):79-82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Başaran N, Duydu Y, Bolt HM. Reproductive toxicity in boron exposed workers in Bandirma, Turkey. Trace Elem Med Biol. 2012;26(2-3):165-167. [DOI] [PubMed] [Google Scholar]
- 135.Cöl M, Cöl C. Environmental boron contamination in waters of Hisarcik area in the Kutahya province of Turkey. Food Chem Toxicol. 2003;41(10):1417-1420. [DOI] [PubMed] [Google Scholar]